Spreading of Isolated Ptch Mutant Basal Cell Carcinoma Precursors Is Physiologically Suppressed and Counteracts Tumor Formation in Mice

 , , and
, , and
Abstract
1. Introduction
2. Results
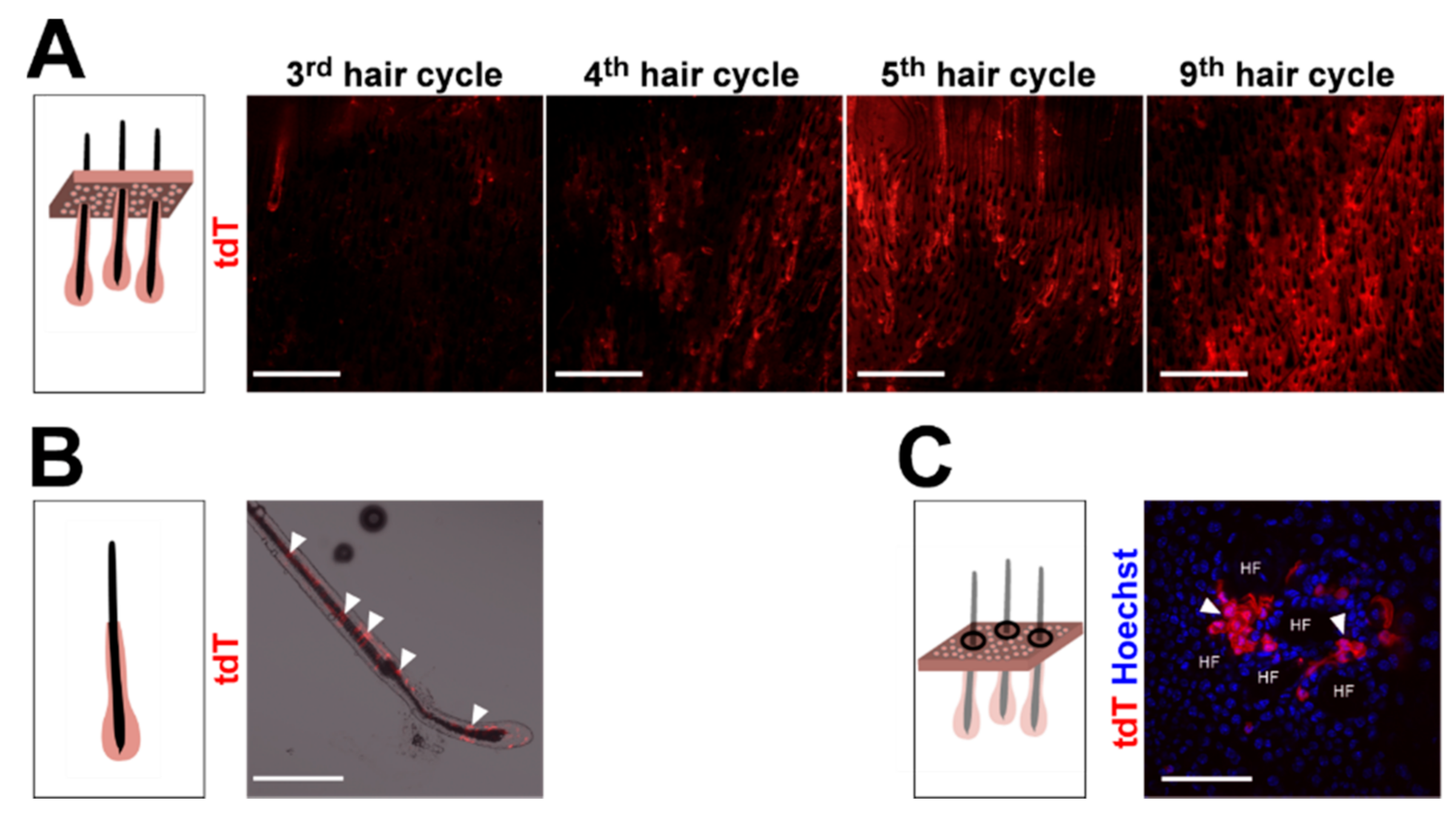
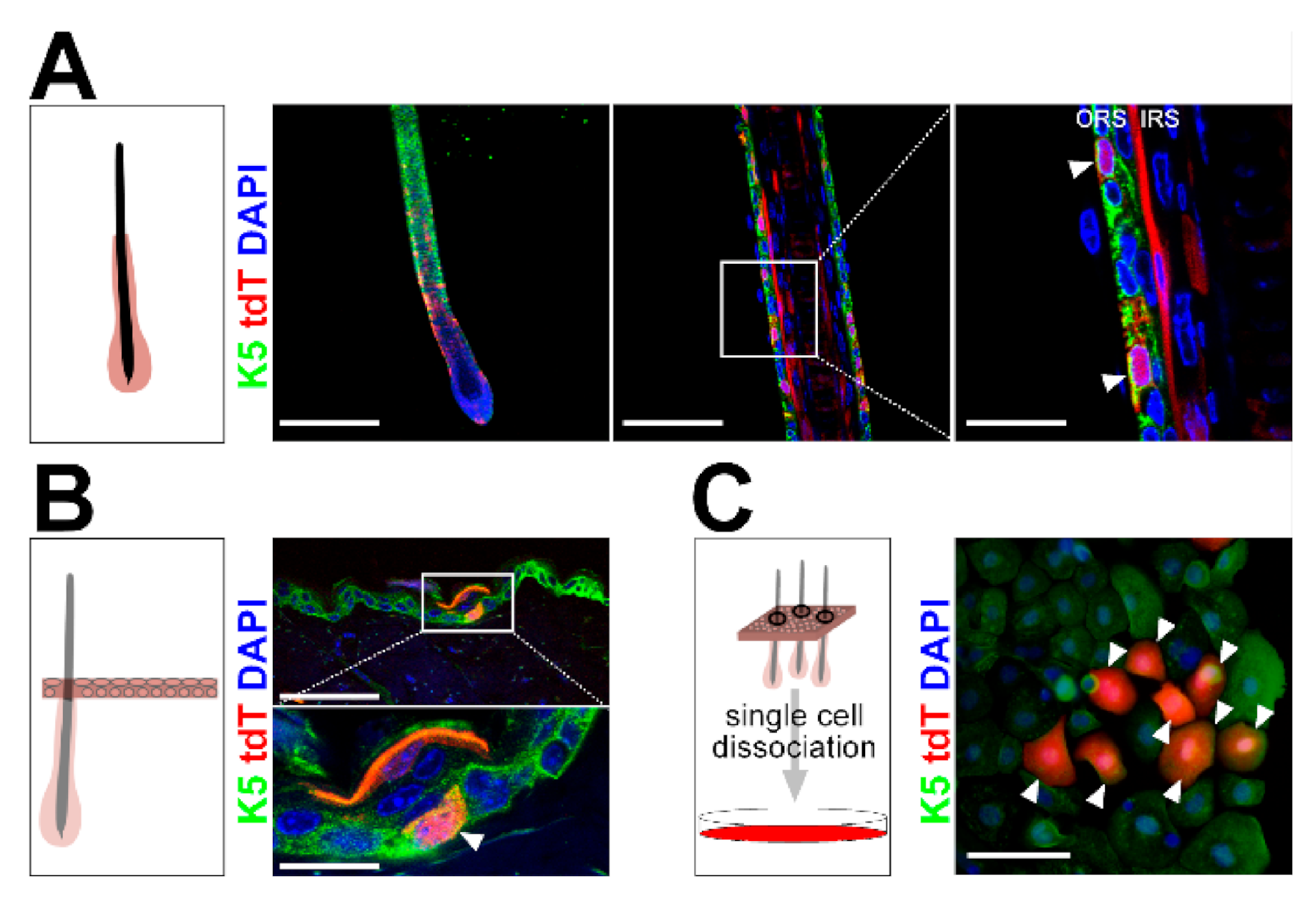
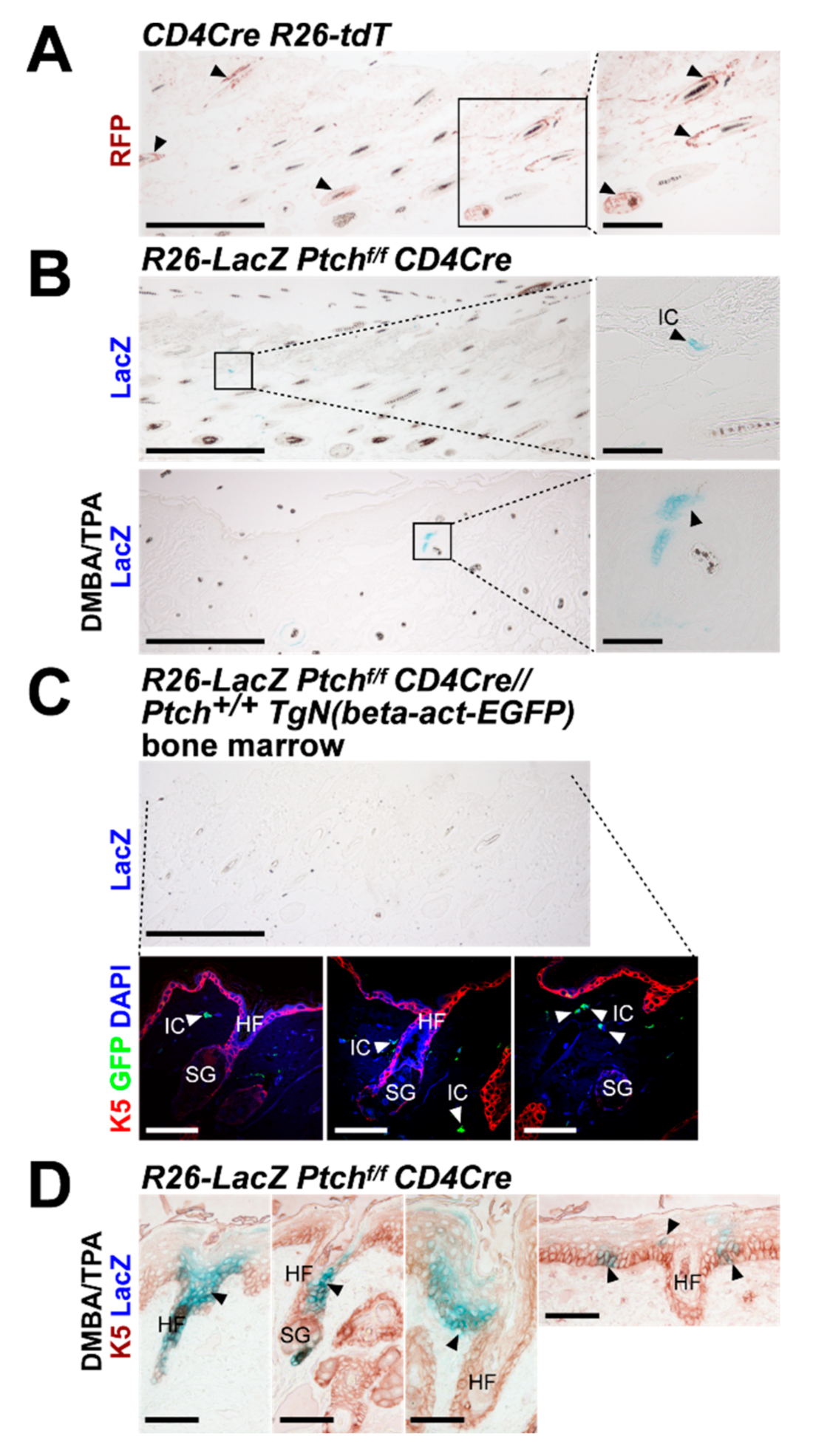
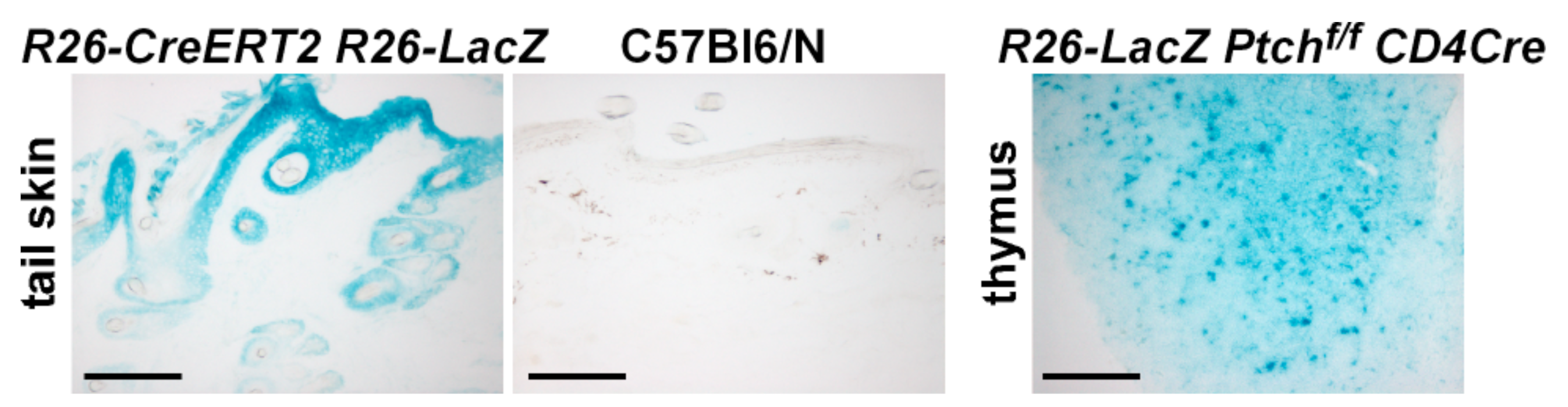
2.1. Wildtype Ptch Progeny of CD4Cre-Targeted Cells Spread over the HF/Skin Complex with Increasing Mouse Age
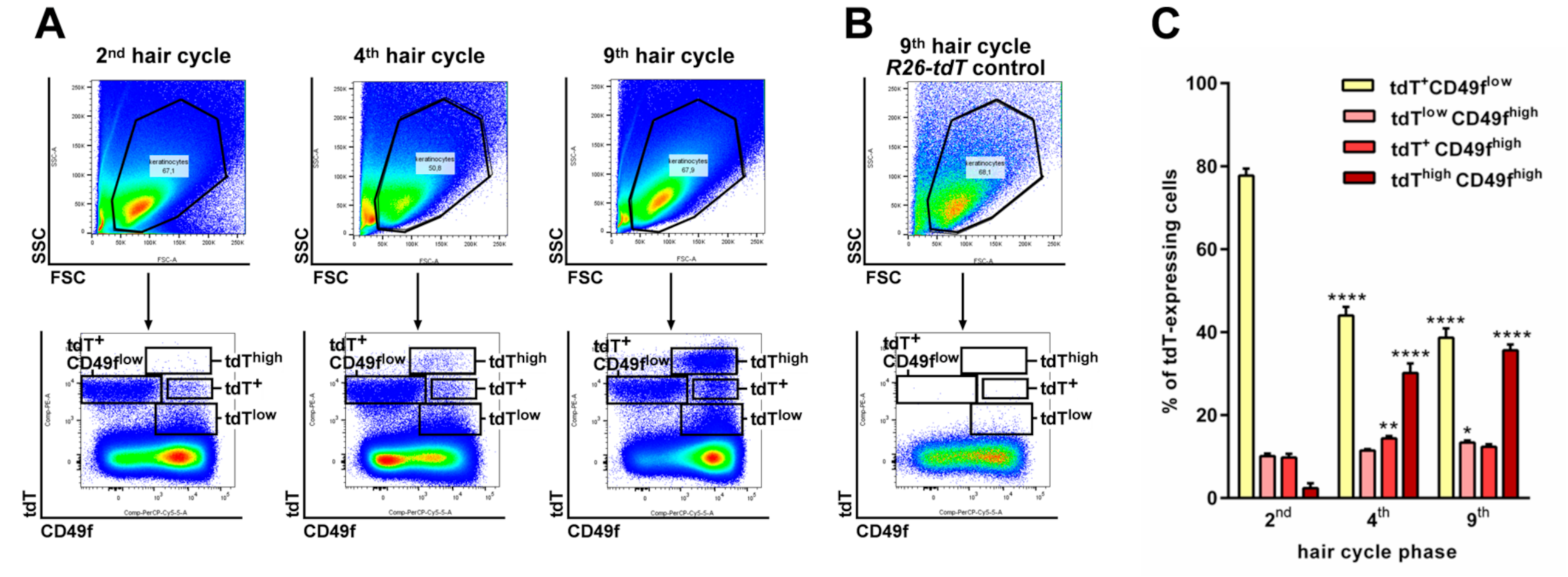
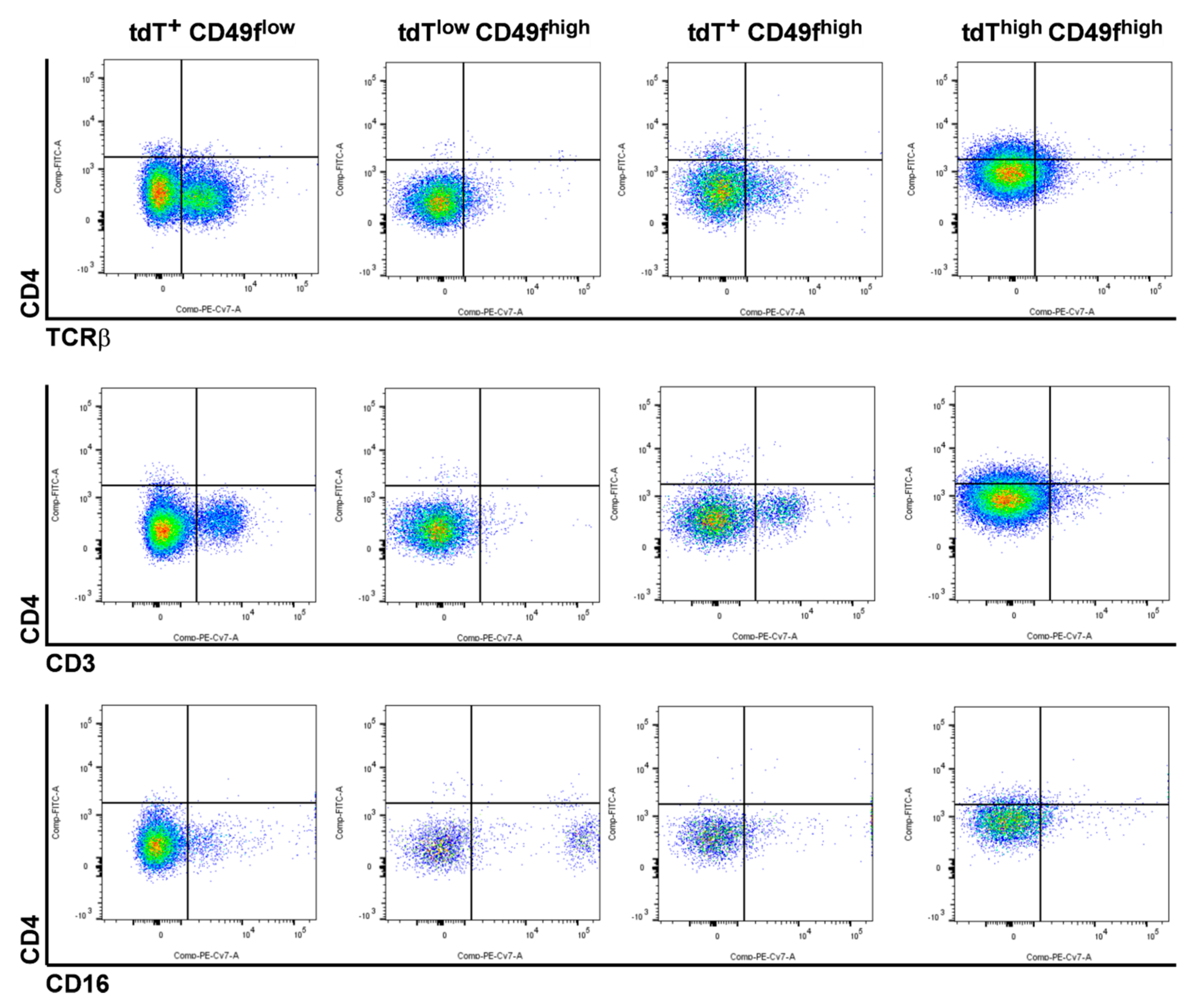
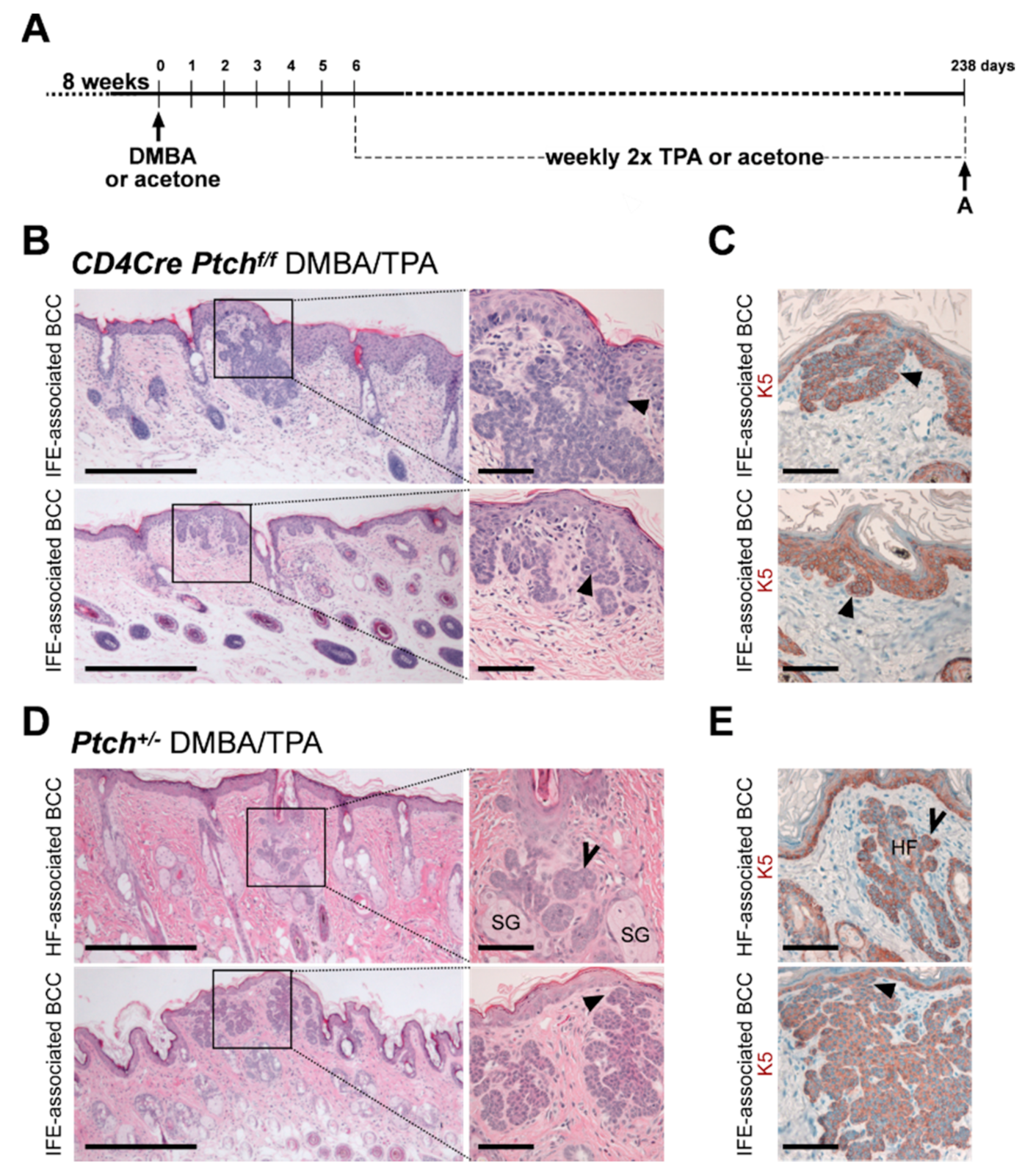
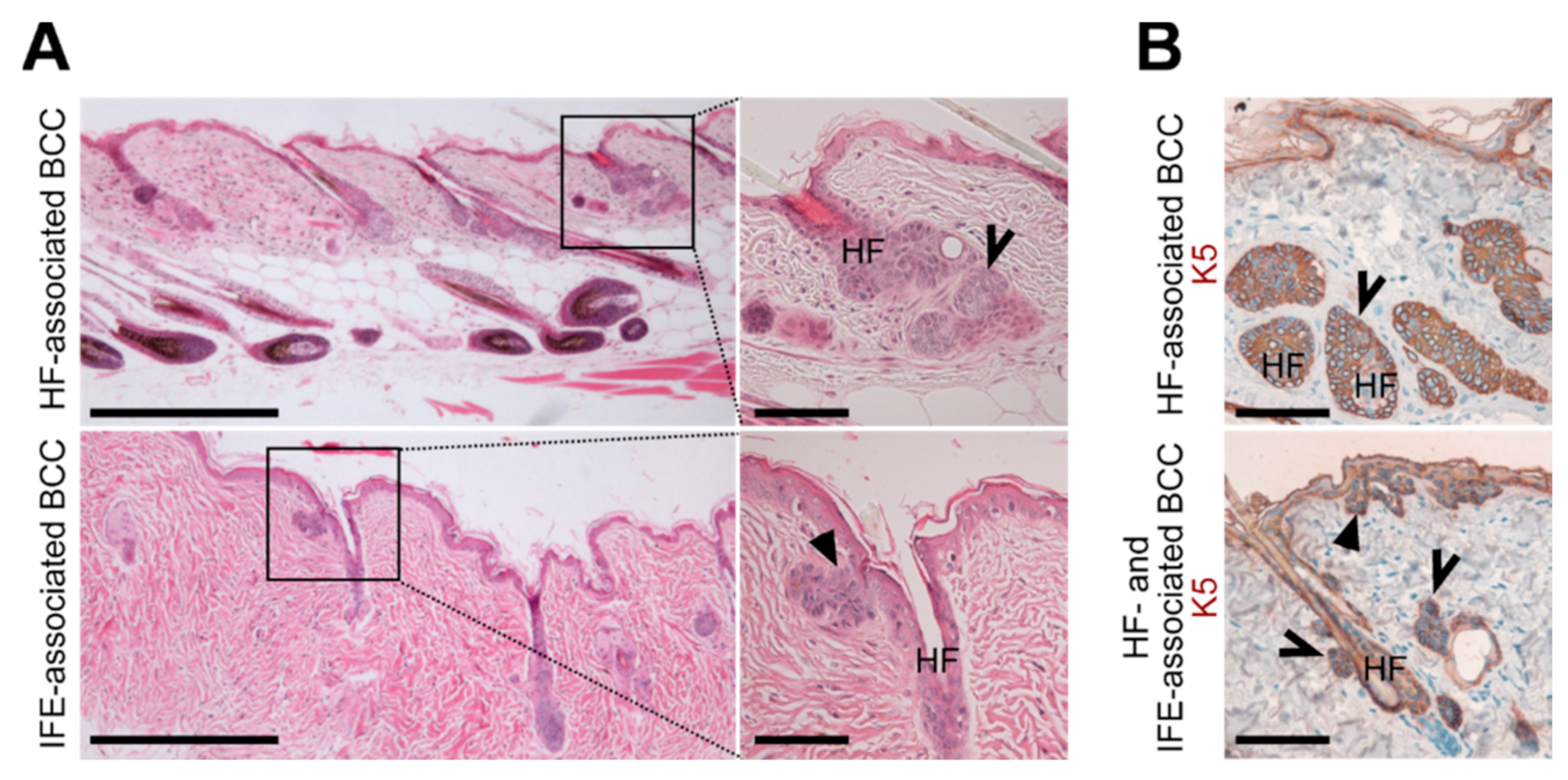
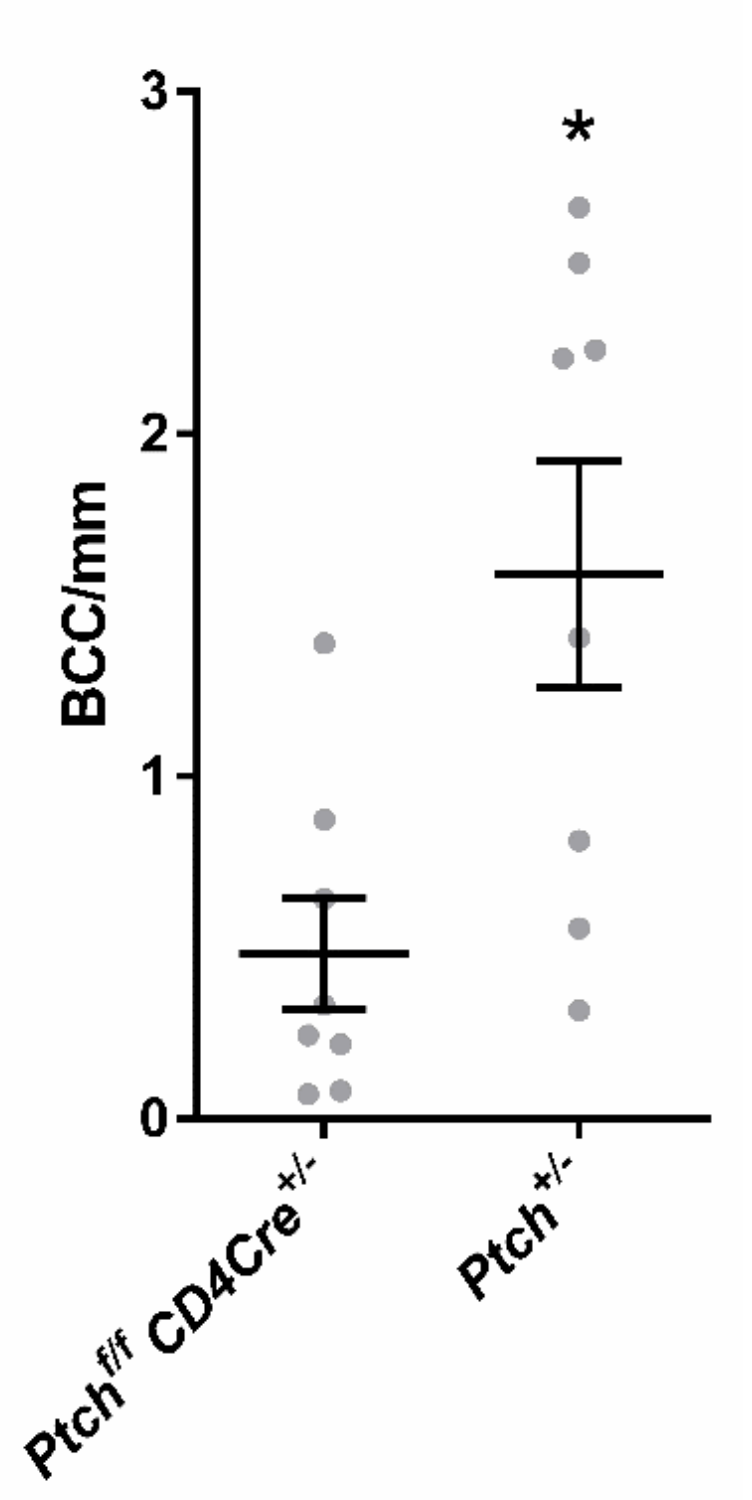
2.2. The CD4Cre-Deleter Targets Keratin 5+ Epidermal Cells That Are the Origin of DMBA/TPA-Induced BCC in Ptchf/f CD4Cre Mice
2.3. Isolated Ptch Mutant Epidermal Cells do Not Spread Like Their Wildtype Ptch Counterparts
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Isolation of Keratinocytes, Epidermal Sheets and Individual Hair Follicles
4.3. Tissue Embedding and Sectioning
4.4. Primary Keratinocyte Culture
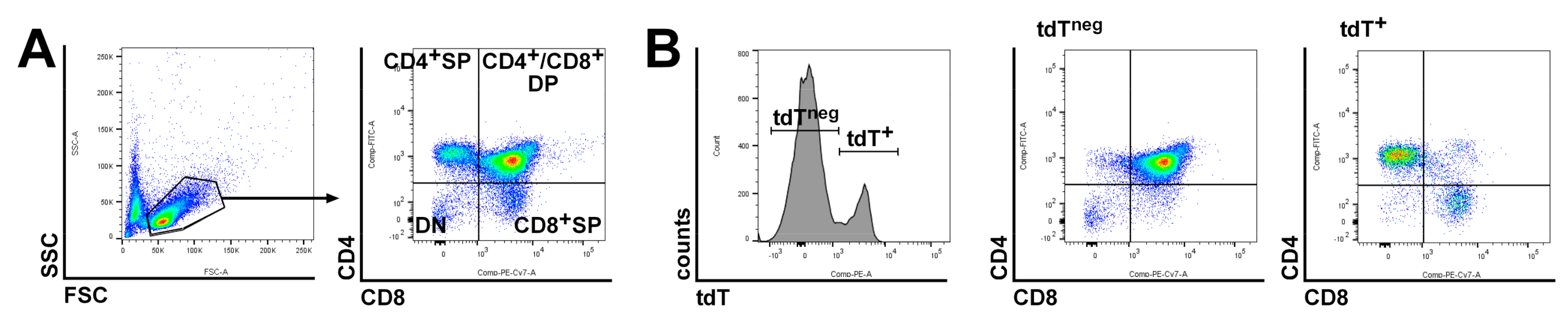
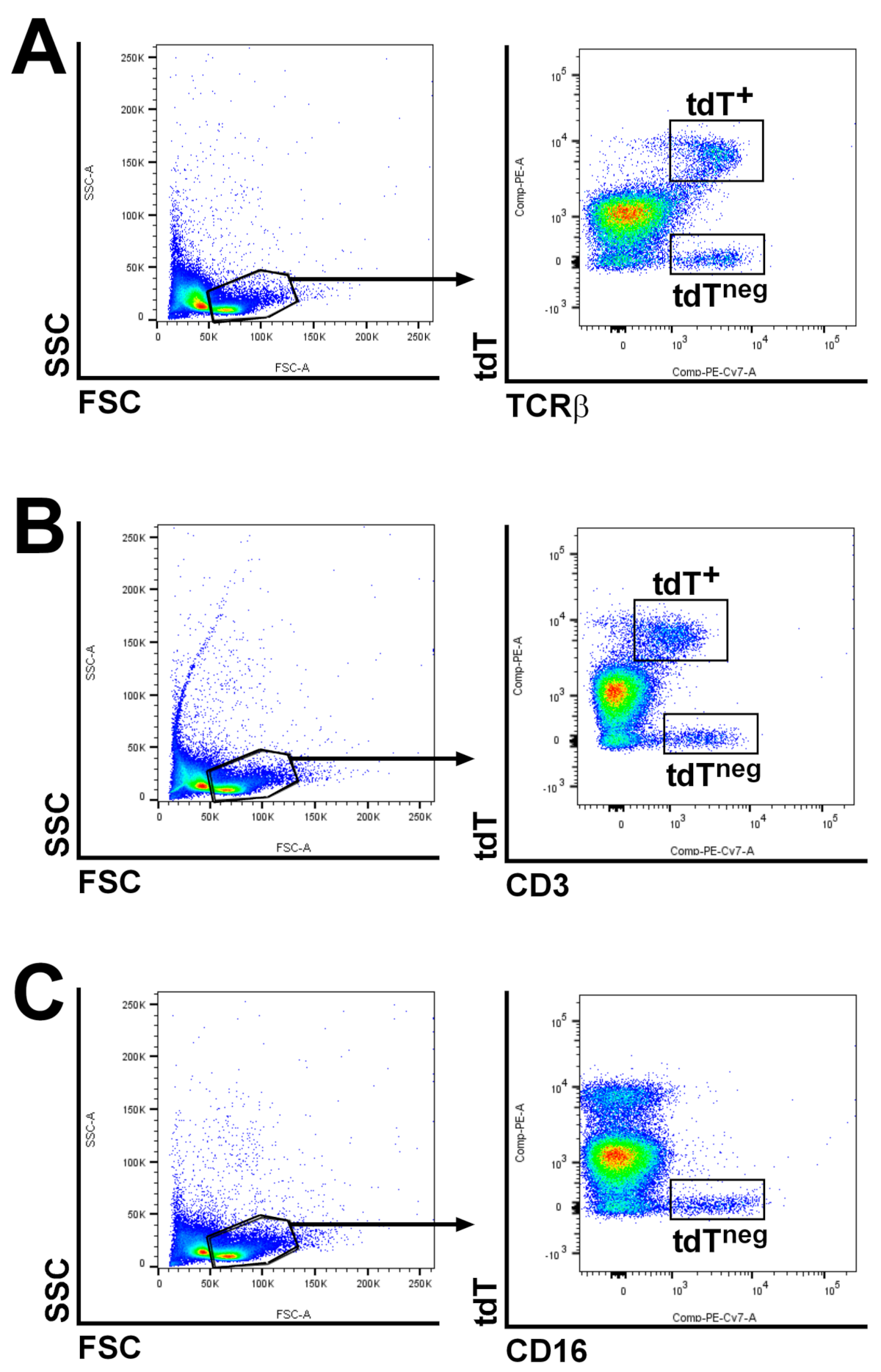
4.5. LacZ Staining, Antibody Staining, Flowcytometric Analyses and Microscopy
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AEC | 3-amino-9-ethylcarbazole |
| BCC | basal cell carcinoma |
| cSCC | cutaneous squamous cell carcinoma |
| CTL | cytotoxic T cells |
| DK-SFM | defined keratinocyte-serum-free medium |
| DMBA | 7,12-Dimethylbenz[a]anthracene |
| DN | double-negative |
| DP | double-positive |
| EDAC | epithelial defense against cancer |
| EGFP | enhanced GFP |
| FITC | fluorescein isothiocyanate |
| FSC | forward scatter |
| GFP | green fluorescent protein |
| H&E | hematoxylin and eosin |
| HF | hair follicle |
| Hh | hedgehog |
| IFE | interfollicular epidermis |
| IRS | inner root sheath |
| K5 | Keratin 5 |
| ORS | outer root sheath |
| PBS | phosphate buffered saline |
| PE | phycoerythrin |
| PerCP | peridinin-chlorophyll-protein |
| Ptch | Patched |
| RFP | red fluorescent protein |
| SG | sebaceous gland |
| SP | single-positive |
| SSC | side scatter |
| TBS | tris buffered saline |
| TCR | T cell receptor |
| tdT | tdTomato |
| TPA | 12-O-tetradecanoylphorbol-13-acetate |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Antibody | Reactivity | Manufacturer (Clone) | Antigen Retrieval | Dilution | Fluorochrome-Labeled Secondary Antibody (Dilution, Manufacturer, Catalogue Code) |
|---|---|---|---|---|---|---|
| GFP | g anti-GFP | Novus Biologicals, Littleton, CO, USA (NB100-1770) | citric acid, pH6 | 1:200 | bov anti-gt-Alexa488 (1:200; Jackson ImmunoResearch Laboratories, Inc., West Grove, PA, USA; #805-545-150) | |
| K5 | rb anti-K5 | m/h | Bio Legend, San Diego, CA, USA (Poly19055) | citric acid, pH6 | 1:1000 | d anti-rb-Alexa488 (1:200; Jackson ImmunoResearch; #711-545-152) d anti-rb-Cy3 (1:400; Jackson ImmunoResearch; #711-165-152) |
| RFP * | g anti-RFP | My BioSource, San Diego, CA, USA (MBS448122) | boric acid or citric acid, pH6 | 1:200 | d anti-g-FITC (1:200; Jackson ImmunoResearch; #705-095-147) bovine anti-g-Alexa488 (1:1000, Jackson ImmunoResearch; #805-545-150) | |
| RFP * | rb anti-RFP | Rockland Immunochemicals, Inc., Gilbertsville, PA, USA (600-401-379S) | boric acid or citric acid, pH6 | 1:500 | d anti-rb-Cy3 (1:400; Jackson ImmunoResearch; #711-165-152) g anti-rb-Cy3 (1:400; Jackson ImmunoResearch; #111-165-003) |
| Antigen | Antibody | Reactivity | Clone/Catalogue Code |
|---|---|---|---|
| CD16/CD32 | r anti-CD16/CD32-PE_Cy7 | m | 2.4G2 |
| CD3ε | ha anti-CD3e-PE-Cy7 | m | 145-2C11 |
| CD4 | r anti-CD4-FITC | m | RM4-5 |
| CD49f | r anti-CD49f-PerCP-Cy5.5 | m/h | GoH3 |
| TCRβ | ha anti-TCRβ-PE-Cy7 | m | H57-597 |





References
- Grachtchouk, M.; Pero, J.; Yang, S.H.; Ermilov, A.N.; Michael, L.E.; Wang, A.; Wilbert, D.; Patel, R.M.; Ferris, J.; Diener, J.; et al. Basal Cell Carcinomas in Mice Arise from Hair Follicle Stem Cells and Multiple Epithelial Progenitor Populations. J. Clin. Investig. 2011, 121, 1768–1781. [Google Scholar] [CrossRef]
- Lapouge, G.; Youssef, K.K.; Vokaer, B.; Achouri, Y.; Michaux, C.; Sotiropoulou, P.A.; Blanpain, C. Identifying the Cellular Origin of Squamous Skin Tumors. Proc. Natl. Acad. Sci. USA 2011, 108, 7431–7436. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.Y.; Wang, J.; Mancianti, M.-L.; Epstein, E. Basal Cell Carcinomas Arise from Hair Follicle Stem Cells in Ptch1+/− Mice. Cancer Cell 2011, 19, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Kasper, M.; Jaks, V.; Are, A.; Bergström, Å.; Schwäger, A.; Svärd, J.; Teglund, S.; Barker, N.; Toftgård, R. Wounding Enhances Epidermal Tumorigenesis by Recruiting Hair Follicle Keratinocytes. Proc. Natl. Acad. Sci. USA 2011, 108, 4099–4104. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.C.; Eberl, M.; Vagnozzi, A.N.; Belkadi, A.; Veniaminova, N.A.; Verhaegen, M.E.; Bichakjian, C.K.; Ward, N.L.; Dlugosz, A.A.; Wong, S.Y. Basal Cell Carcinoma Preferentially Arises from Stem Cells Within Hair Follicle and Mechanosensory Niches. Cell Stem Cell 2015, 16, 400–412. [Google Scholar] [CrossRef]
- Wong, S.Y.; Reiter, J.F. From the Cover: Wounding mobilizes hair follicle stem cells to form tumors. Proc. Natl Acad Sci USA 2011, 108, 4093–4098. [Google Scholar] [CrossRef] [PubMed]
- Puig, S.; Berrocal, A. Management of High-Risk and Advanced Basal Cell Carcinoma. Clin. Transl. Oncol. 2015, 17, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Epstein, E. Basal Cell Carcinomas: Attack of the Hedgehog. Nat. Rev. Cancer 2008, 8, 743–754. [Google Scholar] [CrossRef]
- Michel, K.D.; Uhmann, A.; Dressel, R.; Brandt, J.V.D.; Hahn, H.; Reichardt, H.M. The Hedgehog Receptor Patched1 in T Cells Is Dispensable for Adaptive Immunity in Mice. PLoS ONE 2013, 8, e61034. [Google Scholar] [CrossRef]
- Uhmann, A.; Brandt, J.V.D.; Dittmann, K.; Heß, I.; Dressel, R.; Binder, C.; Lühder, F.; Christiansen, H.; Fassnacht, M.; Bhandoola, A.; et al. T Cell Development Critically Depends on Prethymic Stromal Patched Expression. J. Immunol. 2011, 186, 3383–3391. [Google Scholar] [CrossRef]
- Uhmann, A.; Heß, I.; Frommhold, A.; König, S.; Zabel, S.; Nitzki, F.; Dittmann, K.; Lühder, F.; Christiansen, H.; Reifenberger, J.; et al. DMBA/TPA Treatment Is Necessary for BCC Formation from Patched Deficient Epidermal Cells in Ptch flox/Flox CD4Cre +/− Mice. J. Investig. Dermatol. 2014, 134, 2620–2629. [Google Scholar] [CrossRef] [PubMed]
- Adolphe, C.; Hetherington, R.; Ellis, T.; Wainwright, B.; Thebault, S.; Flourakis, M.; Vanoverberghe, K.; Vandermoere, F.; Roudbaraki, M.; Lehen’Kyi, V.; et al. Patched1 Functions As a Gatekeeper by Promoting Cell Cycle Progression. Cancer Res. 2006, 66, 2081–2088. [Google Scholar] [CrossRef] [PubMed]
- Adolphe, C.; Nieuwenhuis, E.; Villani, R.; Li, Z.J.; Kaur, P.; Hui, C.-C.; Wainwright, B. Patched 1 and Patched 2 Redundancy Has a Key Role in Regulating Epidermal Differentiation. J. Investig. Dermatol. 2014, 134, 1981–1990. [Google Scholar] [CrossRef] [PubMed]
- Reeves, M.Q.; Kandyba, E.; Harris, S.; Del Rosario, R.; Balmain, A. Multicolour Lineage Tracing Reveals Clonal Dynamics of Squamous Carcinoma Evolution from Initiation to Metastasis. Nat. Cell Biol. 2018, 20, 699–709. [Google Scholar] [CrossRef]
- Nitzki, F.; Becker, M.; Frommhold, A.; Schulz-Schaeffer, W.; Hahn, H. Patched Knockout Mouse Models of Basal Cell Carcinoma. J. Ski. Cancer 2012, 2012, 1–11. [Google Scholar] [CrossRef]
- Pyczek, J.; Khizanishvili, N.; Kuzyakova, M.; Zabel, S.; Bauer, J.; Nitzki, F.; Emmert, S.; Schön, M.P.; Boukamp, P.; Schildhaus, H.-U.; et al. Regulation and Role of GLI1 in Cutaneous Squamous Cell Carcinoma Pathogenesis. Front. Genet. 2019, 10, 1185. [Google Scholar] [CrossRef]
- Pellegrini, C.; Maturo, M.G.; Di Nardo, L.; Ciciarelli, V.; García-Rodrigo, C.G.; Fargnoli, M.C. Understanding the Molecular Genetics of Basal Cell Carcinoma. Int. J. Mol. Sci. 2017, 18, 2485. [Google Scholar] [CrossRef]
- Tanimura, N.; Fujita, Y. Epithelial Defense against Cancer (EDAC). Semin. Cancer Biol. 2020, 63, 44–48. [Google Scholar] [CrossRef]
- Kong, Y.-H.; Xu, S.-P. Salidroside Prevents Skin Carcinogenesis Induced by DMBA/TPA in a Mouse Model through Suppression of Inflammation and Promotion of Apoptosis. Oncol. Rep. 2018, 39, 2513–2526. [Google Scholar] [CrossRef]
- Athar, M. Inhibition of Smoothened Signaling Prevents Ultraviolet B-Induced Basal Cell Carcinomas through Regulation of Fas Expression and Apoptosis. Cancer Res. 2004, 64, 7545–7552. [Google Scholar] [CrossRef]
- Biehs, B.; Dijkgraaf, G.J.P.; Piskol, R.; Alicke, B.; Boumahdi, S.; Peale, F.; Gould, S.E.; De Sauvage, F.J. A Cell Identity Switch Allows Residual BCC to Survive Hedgehog Pathway Inhibition. Nat. Cell Biol. 2018, 562, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, S.C.; Tang, X.; Arumugam, A.; Li, C.; Srivastava, R.K.; Weng, Z.; Xu, J.; Zhang, X.; Kim, A.L.; McKay, K.; et al. Shh and p50/Bcl3 Signaling Crosstalk Drives Pathogenesis of BCCs in Gorlin Syndrome. Oncotarget 2015, 6, 36789–36814. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.L.; Back, J.H.; Zhu, Y.; Tang, X.; Yardley, N.P.; Kim, K.J.; Athar, M.; Bickers, D.R. AKT1 Activation Is Obligatory for Spontaneous BCC Tumor Growth in a Murine Model That Mimics Some Features of Basal Cell Nevus Syndrome. Cancer Prev. Res. 2016, 9, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.A.; Sundram, U.; Chang, A.L.S. Two Different Scenarios of Squamous Cell Carcinoma within Advanced Basal Cell Carcinomas. JAMA Dermatol. 2014, 150, 970. [Google Scholar] [CrossRef] [PubMed]
- Orouji, A.; Goerdt, S.; Utikal, J.; Leverkus, M. Multiple Highly and Moderately Differentiated Squamous Cell Carcinomas of the Skin during Vismodegib Treatment of Inoperable Basal Cell Carcinoma. Br. J. Dermatol. 2014, 171, 431–433. [Google Scholar] [CrossRef] [PubMed]
- Aasi, S.; Silkiss, R.; Tang, J.Y.; Wysong, A.; Liu, A.; Epstein, E.; Oro, A.E.; Chang, A.L.S. New Onset of Keratoacanthomas after Vismodegib Treatment for Locally Advanced Basal Cell Carcinomas: A Report of 2 Cases. JAMA Dermatol. 2013, 149, 242–243. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Ponomaryov, T.; Ornell, K.J.; Zhou, P.; Dabral, S.K.; Pak, E.; Li, W.; Atwood, S.X.; Whitson, R.J.; Chang, A.L.S.; et al. RAS/MAPK Activation Drives Resistance to Smo Inhibition, Metastasis, and Tumor Evolution in Shh Pathway-Dependent Tumors. Cancer Res. 2015, 75, 3623–3635. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.J.; Sanborn, Z.; Arnett, K.L.; Bayston, L.J.; Liao, W.; Proby, C.M.; Leigh, I.M.; Collisson, E.A.; Gordon, P.B.; Jakkula, L.; et al. Loss-of-Function Mutations in Notch Receptors in Cutaneous and Lung Squamous Cell Carcinoma. Proc. Natl. Acad. Sci. USA 2011, 108, 17761–17766. [Google Scholar] [CrossRef]
- Pickering, C.R.; Zhou, J.H.; Lee, J.J.; Drummond, J.A.; Peng, S.A.; Saade, R.E.; Tsai, K.Y.; Curry, J.L.; Tetzlaff, M.T.; Lai, S.Y.; et al. Mutational Landscape of Aggressive Cutaneous Squamous Cell Carcinoma. Clin. Cancer Res. 2014, 20, 6582–6592. [Google Scholar] [CrossRef]
- Dong, X.; Meng, L.; Liu, P.; Ji, R.; Su, X.; Xin, Y.; Jiang, X. YAP/TAZ: A Promising Target for Squamous Cell Carcinoma Treatment. Cancer Manag. Res. 2019, 11, 6245–6252. [Google Scholar] [CrossRef]
- Kato, H.; Kurosawa, K.; Inoue, Y.; Tanuma, N.; Momoi, Y.; Hayashi, K.; Ogoh, H.; Nomura, M.; Sakayori, M.; Kakugawa, Y.; et al. Loss of Protein Phosphatase 6 in Mouse Keratinocytes Increases Susceptibility to Ultraviolet-B-Induced Carcinogenesis. Cancer Lett. 2015, 365, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.D.; Lynch, C.N.S.; Craythorne, E.; Liakath-Ali, K.; Mallipeddi, R.; Barker, J.N.; Watt, F.M. Spatial Constraints Govern Competition of Mutant Clones in Human Epidermis. Nat. Commun. 2017, 8, 1119. [Google Scholar] [CrossRef]
- Wolfer, A.; Bakker, T.; Wilson, A.; Nicolas, M.; Ioannidis, V.; Littman, D.R.; Wilson, C.B.; Held, W.; Macdonald, H.R.; Radtke, F. Inactivation of Notch1 in Immature Thymocytes Does Not Perturb CD4 or CD8 T Cell Development. Nat. Immunol. 2001, 2, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.P.; Fitzpatrick, D.R.; Beard, C.; Jessup, H.K.; Lehar, S.; Makar, K.W.; Pérez-Melgosa, M.; Sweetser, M.T.; Schlissel, M.S.; Nguyen, S.; et al. A Critical Role for Dnmt1 and DNA Methylation in T Cell Development, Function, and Survival. Immun. 2001, 15, 763–774. [Google Scholar] [CrossRef]
- Sawada, S.; Scarborough, J.D.; Killeen, N.; Littman, D.R. A Lineage-Specific Transcriptional Silencer Regulates CD4 Gene Expression During T Lymphocyte Development. Cell 1994, 77, 917–929. [Google Scholar] [CrossRef]
- Indra, A.K.; Warot, X.; Brocard, J.; Bornert, J.-M.; Xiao, J.-H.; Chambon, P.; Metzger, D. Temporally-Controlled Site-Specific Mutagenesis in the Basal Layer of the Epidermis: Comparison of the Recombinase Activity of the Tamoxifen-Inducible Cre-ERT and Cre-ERT2 Recombinases. Nucleic Acids Res. 1999, 27, 4324–4327. [Google Scholar] [CrossRef]
- Madisen, L.; Zwingman, T.A.; Sunkin, S.M.; Oh, S.W.; Zariwala, H.A.; Gu, H.; Ng, L.L.; Palmiter, R.D.; Hawrylycz, M.J.; Jones, A.R.; et al. A Robust and High-Throughput Cre Reporting and Characterization System for the Whole Mouse Brain. Nat. Neurosci. 2010, 13, 133–140. [Google Scholar] [CrossRef]
- Soriano, P. Generalized LacZ Expression With the ROSA26 Cre Reporter Strain. Nat. Genet. 1999, 21, 70–71. [Google Scholar] [CrossRef]
- Ventura, A.; Kirsch, D.G.; McLaughlin, M.E.; Tuveson, D.A.; Grimm, J.; Lintault, L.; Newman, J.C.J.; Reczek, E.E.; Weissleder, R.; Jacks, T. Restoration of p53 Function Leads to Tumour Regression in Vivo. Nat. Cell Biol. 2007, 445, 661–665. [Google Scholar] [CrossRef]
- Okabe, M.; Ikawa, M.; Kominami, K.; Nakanishi, T.; Nishimune, Y. ‘Green mice’ As a Source of Ubiquitous Green Cells. FEBS Lett. 1997, 407, 313–319. [Google Scholar] [CrossRef]
- Hahn, H.; Wojnowski, L.; Zimmer, A.M.; Hall, J.; Miller, G.; Zimmer, A. Rhabdomyosarcomas and Radiation Hypersensitivity in a Mouse Model of Gorlin Syndrome. Nat. Med. 1998, 4, 619–622. [Google Scholar] [CrossRef] [PubMed]
- Uhmann, A.; Dittmann, K.; Nitzki, F.; Dressel, R.; Koleva, M.; Frommhold, A.; Zibat, A.; Binder, C.; Adham, I.; Nitsche, M.; et al. The Hedgehog Receptor Patched Controls Lymphoid Lineage Commitment. Blood 2007, 110, 1814–1823. [Google Scholar] [CrossRef] [PubMed]
- Braun, K.M.; Niemann, C.; Jensen, U.B.; Sundberg, J.P.; Silva-Vargas, V.; Watt, F.M. Manipulation of Stem Cell Proliferation and Lineage Commitment: Visualisation of Label-Retaining Cells in Wholemounts of Mouse Epidermis. Dev. 2003, 130, 5241–5255. [Google Scholar] [CrossRef] [PubMed]
- Sequeira, I.; Legue, E.; Capgras, S.; Nicolas, J.-F. Microdissection and Visualization of Individual Hair Follicles for Lineage Tracing Studies. Advanced Structural Safety Studies 2013, 1195, 247–258. [Google Scholar] [CrossRef]
- Pyczek, J.; Buslei, R.; Schult, D.; Hölsken, A.; Buchfelder, M.; Heß, I.; Hahn, H.; Uhmann, A. Hedgehog Signaling Activation Induces Stem Cell Proliferation and Hormone Release in the Adult Pituitary Gland. Sci. Rep. 2016, 6, 24928. [Google Scholar] [CrossRef]
- Dräger, J.; Simon-Keller, K.; Pukrop, T.; Klemm, F.; Wilting, J.; Sticht, C.; Dittmann, K.; Schulz, M.; Leuschner, I.; Marx, A.; et al. LEF1 Reduces Tumor Progression and Induces Myodifferentiation in a Subset of Rhabdomyosarcoma. Oncotarget 2017, 8, 3259–3273. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brandes, N.; Mitkovska, S.H.; Botermann, D.S.; Maurer, W.; Müllen, A.; Scheile, H.; Zabel, S.; Frommhold, A.; Heß, I.; Hahn, H.; et al. Spreading of Isolated Ptch Mutant Basal Cell Carcinoma Precursors Is Physiologically Suppressed and Counteracts Tumor Formation in Mice. Int. J. Mol. Sci. 2020, 21, 9295. https://doi.org/10.3390/ijms21239295
Brandes N, Mitkovska SH, Botermann DS, Maurer W, Müllen A, Scheile H, Zabel S, Frommhold A, Heß I, Hahn H, et al. Spreading of Isolated Ptch Mutant Basal Cell Carcinoma Precursors Is Physiologically Suppressed and Counteracts Tumor Formation in Mice. International Journal of Molecular Sciences. 2020; 21(23):9295. https://doi.org/10.3390/ijms21239295
Chicago/Turabian StyleBrandes, Nadine, Slavica Hristomanova Mitkovska, Dominik Simon Botermann, Wiebke Maurer, Anna Müllen, Hanna Scheile, Sebastian Zabel, Anke Frommhold, Ina Heß, Heidi Hahn, and et al. 2020. "Spreading of Isolated Ptch Mutant Basal Cell Carcinoma Precursors Is Physiologically Suppressed and Counteracts Tumor Formation in Mice" International Journal of Molecular Sciences 21, no. 23: 9295. https://doi.org/10.3390/ijms21239295
APA StyleBrandes, N., Mitkovska, S. H., Botermann, D. S., Maurer, W., Müllen, A., Scheile, H., Zabel, S., Frommhold, A., Heß, I., Hahn, H., & Uhmann, A. (2020). Spreading of Isolated Ptch Mutant Basal Cell Carcinoma Precursors Is Physiologically Suppressed and Counteracts Tumor Formation in Mice. International Journal of Molecular Sciences, 21(23), 9295. https://doi.org/10.3390/ijms21239295