The Plant Sesquiterpene Nootkatone Efficiently Reduces Heterodera schachtii Parasitism by Activating Plant Defense

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
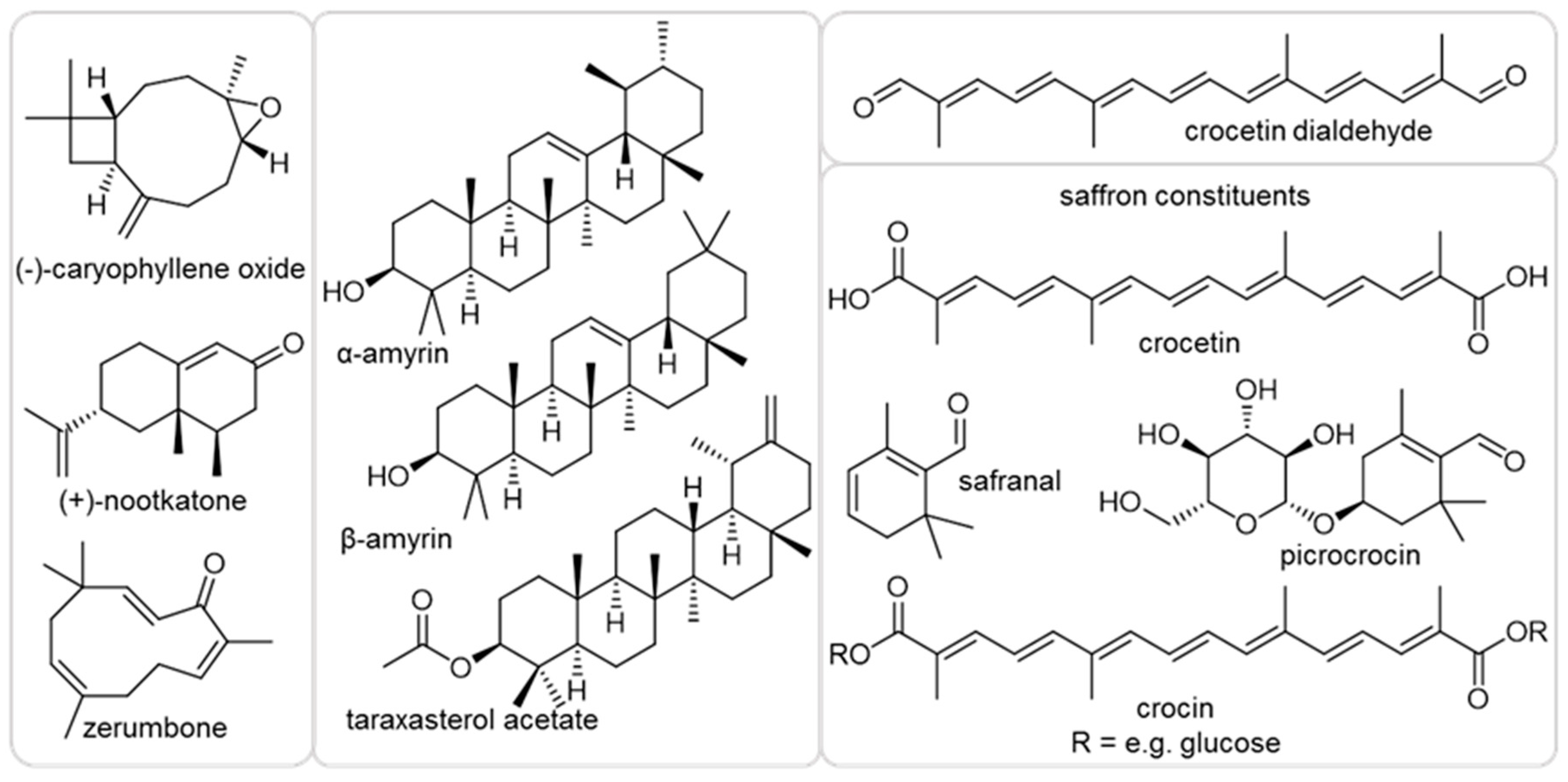
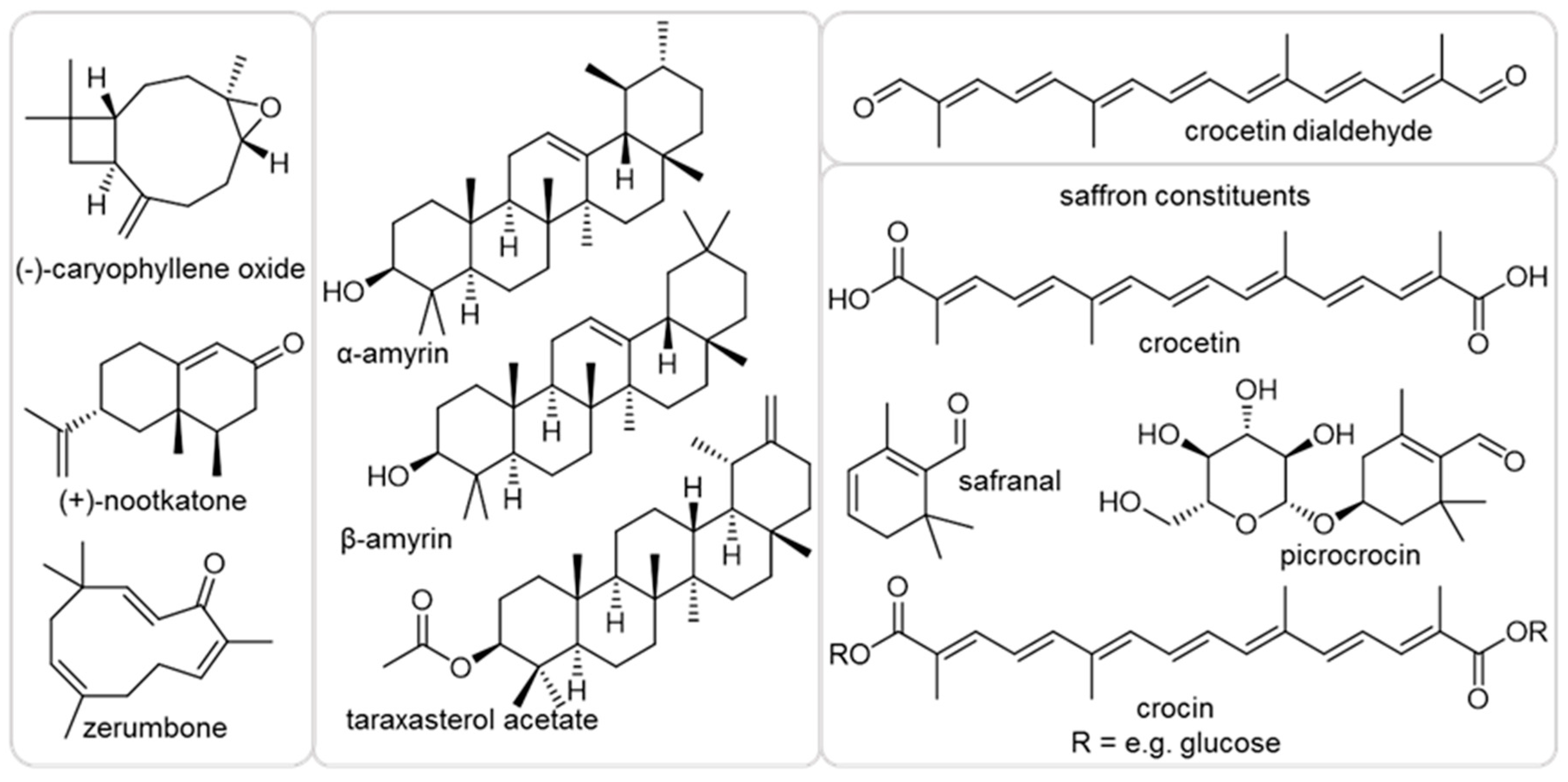
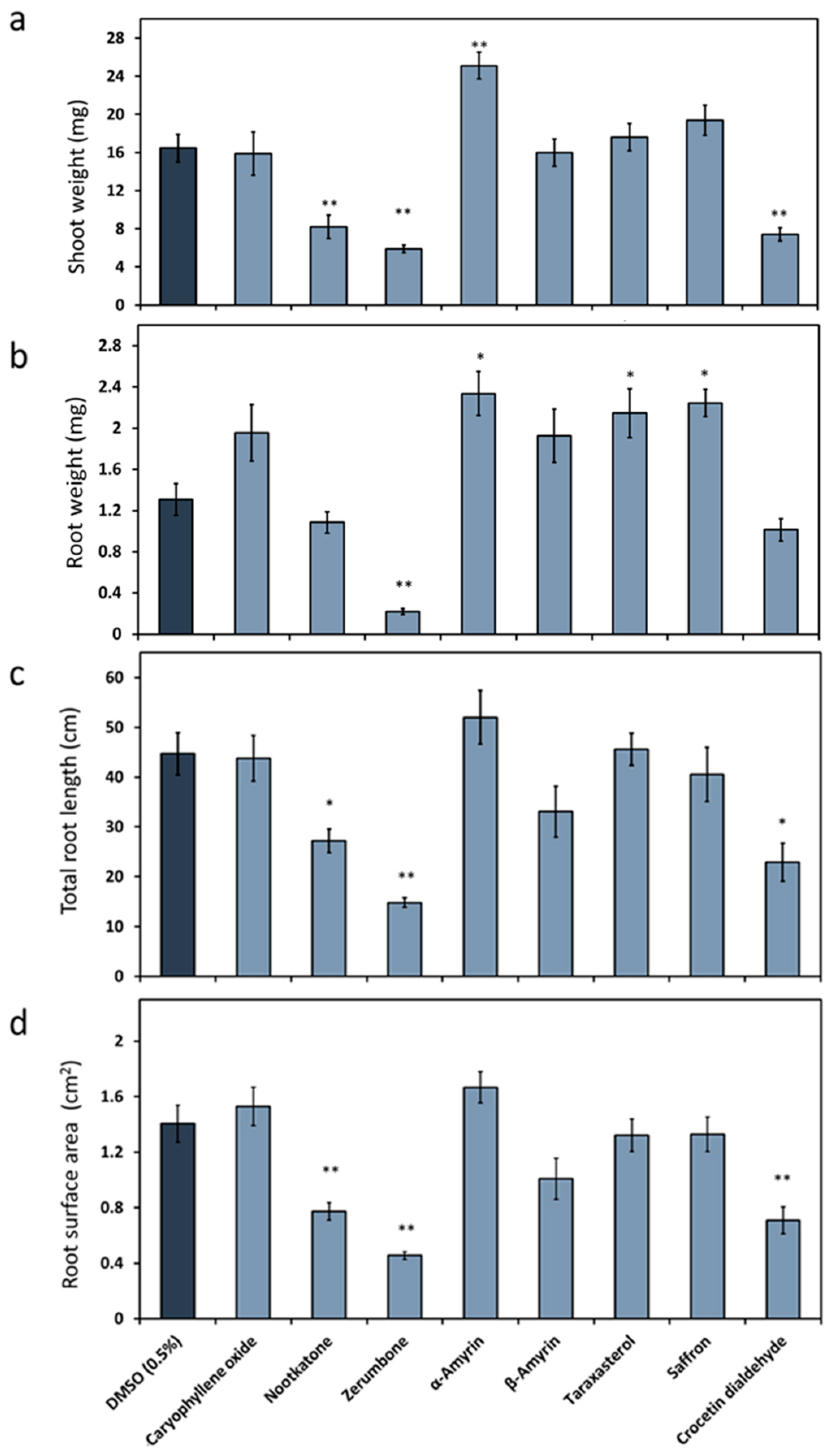
2.1. Effect of Plant Terpenoids on A. thaliana Growth
2.2. Effect of Plant Metabolites on H. schachtii Parasitism at A. thaliana
2.3. Nootkatone Has No Direct Effect on J2s of H. schachtii
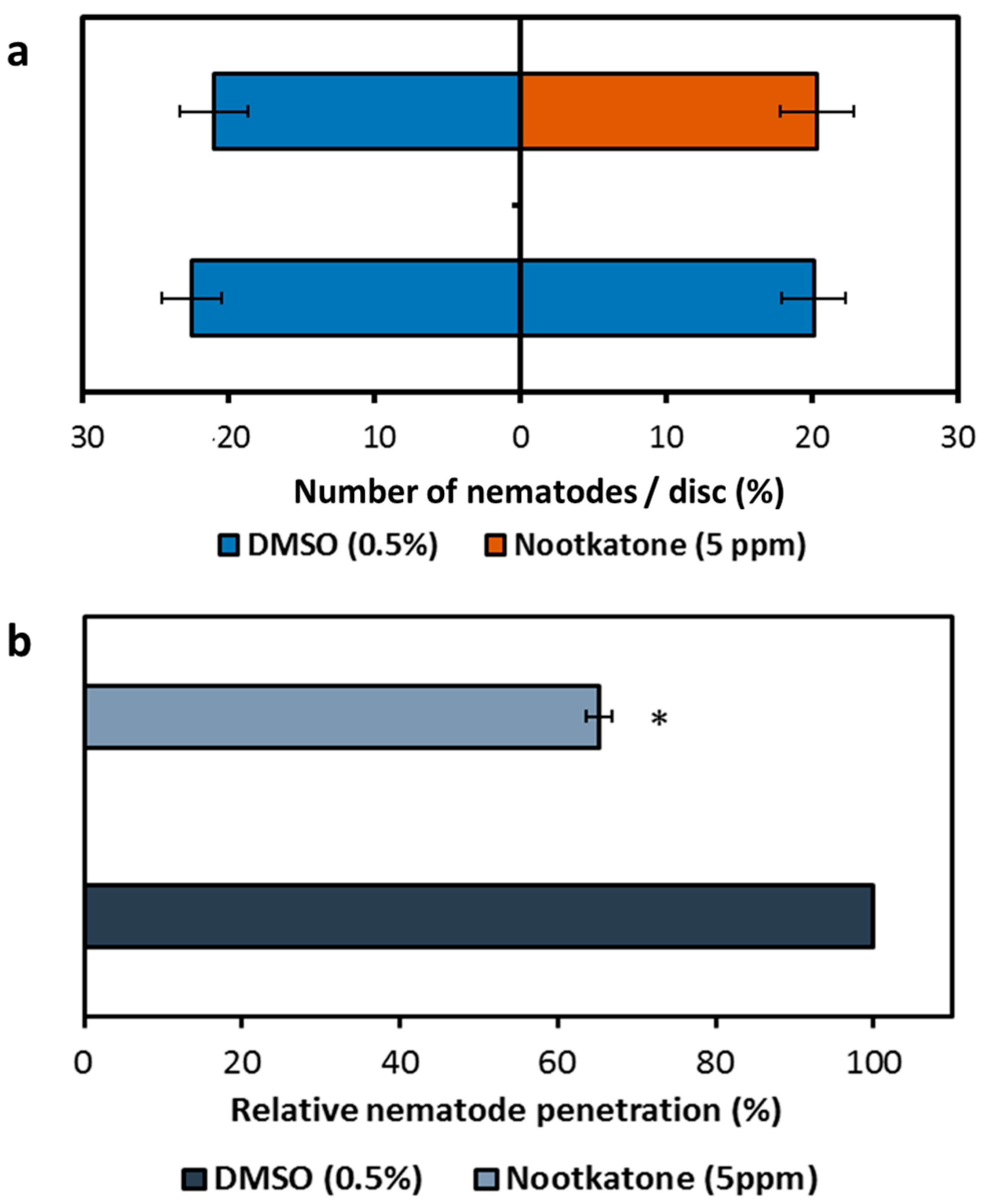
2.4. Nootkatone Does Not Affect Nematode Attraction to the Plant but Inhibits Nematode Root Penetration
2.5. Nootkatone Activates Plant Defense
2.6. Nootkatone Does Not Induce the Biosynthesis of Sesquiterpenes in A. thaliana
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Nematode Culture
4.3. Effect of Terpenoids on A. thaliana Growth
4.4. Infection Assay
4.5. Attraction Assay
4.6. Root Penetration Assay
4.7. Mortality Assay
4.8. Reactive Oxygen Species (ROS) Measurements
4.9. Quantitative Real-Time PCR
4.10. HS–SPME–GC×GC–TOF–MS Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Aury, J.; Castagnone-Sereno, P.; Danchin, E.G.J.; Abad, P.; Deleury, E.; Perfus-Barbeoch, L.; Coutinho, P.M.; Dasilva, C.; De Luca, F.; Blok, V.C.; et al. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, J.E.; Wesemael, W.M.L.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef]
- Steele, A.E. The host range of the sugar beet nematode, Heterodera schachtii Schmidt. JSBR 1965, 13, 574–603. [Google Scholar] [CrossRef] [Green Version]
- Muller, J. The economic importance of Heterodera schachtii in Europe. Helminthologia 1999, 36, 205–213. [Google Scholar]
- Wyss, U.; Grundler, F.M.W. Feeding-behavior of sedentary plant parasitic nematodes. Netherlands J. Plant Pathol. 1992, 98, 165–173. [Google Scholar] [CrossRef]
- Da Silva, F.F.A.; Alves, C.C.F.; de Oliveira Filho, J.G.; Vieira, T.M.; Crotti, A.E.M.; Miranda, M.L.D. Chemical constituents of essential oil from Murraya paniculata leaves and its application to in vitro biological control of the fungus Sclerotinia sclerotiorum. Food Sci. Technol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.J.; Gray, F.A.; Koch, D.W. Reproduction of Heterodera schachtii Schmidt on resistant mustard, radish, and sugar beet cultivars. J. Nematol. 2004, 36, 123–130. [Google Scholar]
- Hauer, M.; Koch, H.J.; Krüssel, S.; Mittler, S.; Märländer, B. Integrated control of Heterodera schachtii Schmidt in Central Europe by trap crop cultivation, sugar beet variety choice and nematicide application. Appl. Soil Ecol. 2016, 99, 62–69. [Google Scholar] [CrossRef]
- Müller, J. Detection of pathotypes by assessing the virulence of Heterodera schachtii populations. Nematologica 1992, 38, 50–64. [Google Scholar] [CrossRef]
- Cheng, A.X.; Lou, Y.G.; Mao, Y.B.; Lu, S.; Wang, L.J.; Chen, X.Y. Plant terpenoids: Biosynthesis and ecological functions. J. Integr. Plant Biol. 2007, 49, 179–186. [Google Scholar] [CrossRef]
- Cheng, A.X.; Xiang, C.Y.; Li, J.X.; Yang, C.Q.; Hu, W.L.; Wang, L.J.; Lou, Y.G.; Chen, X.Y. The rice (E)-β-caryophyllene synthase (OsTPS3) accounts for the major inducible volatile sesquiterpenes. Phytochemistry 2007, 68, 1632–1641. [Google Scholar] [CrossRef]
- Echeverrigaray, S.; Zacaria, J.; Beltrão, R. Nematicidal activity of monoterpenoids against the root-knot nematode Meloidogyne incognita. Phytopathology 2010, 100, 199–203. [Google Scholar] [CrossRef] [Green Version]
- Zengin, H.; Baysal, A.H. Antibacterial and antioxidant activity of essential oil terpenes against pathogenic and spoilage-forming bacteria and cell structure-activity relationships evaluated by SEM microscopy. Molecules 2014, 19, 17773–17798. [Google Scholar] [CrossRef] [Green Version]
- Dambolena, J.S.; Zunino, M.P.; Herrera, J.M.; Pizzolitto, R.P.; Areco, V.A.; Zygadlo, J.A. Terpenes: Natural products for controlling insects of importance to human health—A structure-activity relationship study. Psyche J. Entomol. 2016. [Google Scholar] [CrossRef] [Green Version]
- Araniti, F.; Sánchez-Moreiras, A.M.; Graña, E.; Reigosa, M.J.; Abenavoli, M.R. Terpenoid trans-caryophyllene inhibits weed germination and induces plant water status alteration and oxidative damage in adult Arabidopsis. Plant Biol. 2017, 19, 79–89. [Google Scholar] [CrossRef]
- Marisco, G.; Santos, R.X.; Aguiar, R.; Brendel, M.; Pungartnik, C. Antifungal potential of terpenes from Spondias purpurea L. leaf extract against Moniliophthora perniciosa that causes witches broom disease of Theobroma cacao. Int. J. Complement. Altern. Med. 2017, 7. [Google Scholar] [CrossRef]
- Macías, F.A.; Lacret, R.; Varela, R.M.; Nogueiras, C.; Molinillo, J.M.G. Isolation and phytotoxicity of terpenes from Tectona grandis. J. Chem. Ecol. 2010, 36, 396–404. [Google Scholar] [CrossRef]
- Ntalli, N.G.; Ferrari, F.; Giannakou, I.; Menkissoglu-Spiroudi, U. Synergistic and antagonistic interactions of terpenes against Meloidogyne incognita and the nematicidal activity of essential oils from seven plants indigenous to Greece. Pest Manag. Sci. 2011, 67, 341–351. [Google Scholar] [CrossRef]
- Abdel-Rahman, F.H.; Alaniz, N.M.; Saleh, M.A. Nematicidal activity of terpenoids. J. Environ. Sci. Heal. Part B Pestic. Food Contam. Agric. Wastes 2013, 48, 16–22. [Google Scholar] [CrossRef]
- Kammerhofer, N.; Radakovic, Z.; Regis, J.M.A.; Dobrev, P.; Vankova, R.; Grundler, F.M.W.; Siddique, S.; Hofmann, J.; Wieczorek, K. Role of stress-related hormones in plant defence during early infection of the cyst nematode Heterodera schachtii in Arabidopsis. New Phytol. 2015, 207, 778–789. [Google Scholar] [CrossRef] [Green Version]
- Rohloff, J.; Bones, A.M. Volatile profiling of Arabidopsis thaliana—Putative olfactory compounds in plant communication. Phytochemistry 2005, 66, 1941–1955. [Google Scholar] [CrossRef] [PubMed]
- Gränicher, F.; Christen, P.; Kapetanidis, I. Essential oils from normal and hairy roots of Valeriana officinalis var. sambucifolia. Phytochemistry 1995, 40, 1421–1424. [Google Scholar]
- Duquesnoy, E.; Castola, V.; Casanova, J. Composition and chemical variability of the twig oil of Abies alba Miller from Corsica. Flavour Fragr. J. 2007, 22, 293–299. [Google Scholar] [CrossRef]
- Gonny, M.; Cavaleiro, C.; Salgueiro, L.; Casanova, J. Analysis of Juniperus communis subsp. alpina needle, berry, wood and root oils by combination of GC, GC/MS and 13C-NMR. Flavour Fragr. J. 2006, 21, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Cavalli, J.F.; Tomi, F.; Bernardini, A.F.; Casanova, J. Chemical variability of the essential oil of Helichrysum faradifani Sc. Ell. from Madagascar. Flavour Fragr. J. 2006, 21, 111–114. [Google Scholar] [CrossRef]
- Bader, A.; Caponi, C.; Cioni, P.L.; Flamini, G.; Morelli, I. Composition of the essential oil of Ballota undulata, B. nigra ssp. foetida and B. saxatilis. Flavour Fragr. J. 2003, 18, 502–504. [Google Scholar] [CrossRef]
- Riu-Aumatell, M.; López-Tamames, E.; Buxaderas, S. Assessment of the volatile composition of juices of apricot, peach, and pear according to two pectolytic treatments. J. Agric. Food Chem. 2005, 53, 7837–7843. [Google Scholar] [CrossRef] [PubMed]
- Sahin, F.; Güllüce, M.; Daferera, D.; Sökmen, A.; Sökmen, M.; Polissiou, M.; Agar, G.; Özer, H. Biological activities of the essential oils and methanol extract of Origanum vulgare ssp. vulgare in the Eastern Anatolia region of Turkey. Food Control 2004, 15, 549–557. [Google Scholar] [CrossRef]
- Sacchetti, G.; Maietti, S.; Muzzoli, M.; Scaglianti, M.; Manfredini, S.; Radice, M.; Bruni, R. Comparative evaluation of 11 essential oils of different origin as functional antioxidants, antiradicals and antimicrobials in foods. Food Chem. 2005, 91, 621–632. [Google Scholar] [CrossRef]
- Malik, S.; de Mesquita, L.; Silva, C.; de Mesquita, J.; de Sá Rocha, E.; Bose, J.; Abiri, R.; de Maria Silva Figueiredo, P.; Costa-Júnior, L. Chemical profile and biological activities of essential oil from Artemisia vulgaris L. cultivated in Brazil. Pharmaceuticals 2019, 12, 49. [Google Scholar] [CrossRef] [Green Version]
- Sijmons, P.C.; Grundler, F.M.W.; von Mende, N.; Burrows, P.R.; Wyss, U. Arabidopsis thaliana as a new model host for plant-parasitic nematodes. Plant J. 1991, 1, 245–254. [Google Scholar] [CrossRef]
- Blümel, R.C.; Fischer, D.F.; Grundler, F.M.W. Effects of exogenous amino acid applications on the plant-parasitic nematode Heterodera schachtii. Nematology 2018, 20, 713–727. [Google Scholar] [CrossRef]
- Panella, N.A.; Dolan, M.C.; Karchesy, J.J.; Xiong, Y.; Peralta-Cruz, J.; Khasawneh, M.; Montenieri, J.A.; Maupin, G.O. Use of novel compounds for pest control: Insecticidal and acaricidal activity of essential oil components from heartwood of Alaska Yellow Cedar. J. Med. Entomol. 2005, 42, 352–358. [Google Scholar] [CrossRef]
- Dolan, M.C.; Jordan, R.A.; Schulze, T.L.; Schulze, C.J.; Manning, M.C.; Ruffolo, D.; Schmidt, J.P.; Piesman, J.; Karchesy, J.J. Ability of two natural products, nootkatone and carvacrol, to suppress Ixodes scapularis and Amblyomma americanum (Acari: Ixodidae) in a lyme disease endemic area of New Jersey. J. Econ. Entomol. 2009, 102, 2316–2324. [Google Scholar] [CrossRef]
- Jordan, R.A.; Schulze, T.L.; Dolan, M.C. Efficacy of plant-derived and synthetic compounds on clothing as repellents against Ixodes scapularis and Amblyomma americanum (Acari: Ixodidae). J. Med. Entomol. 2012, 49, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Pretel, A.G.; Del Pulgar, H.P.; Olmeda, A.S.; Gonzalez-Coloma, A.; Barrero, A.F.; Del Moral, J.F.Q. Novel insect antifeedant and ixodicidal nootkatone derivatives. Biomolecules 2019, 9, 742. [Google Scholar] [CrossRef] [Green Version]
- Hiltpold, I.; Baroni, M.; Toepfer, S.; Kuhlmann, U.; Turlings, T.C.J. Selective breeding of entomopathogenic nematodes for enhanced attraction to a root signal did not reduce their establishment or persistence after field release. Plant Signal. Behav. 2010, 5, 1450–1452. [Google Scholar] [CrossRef] [Green Version]
- Hiltpold, I.; Baroni, M.; Toepfer, S.; Kuhlmann, U.; Turlings, T.C.J. Selection of entomopathogenic nematodes for enhanced responsiveness to a volatile root signal helps to control a major root pest. J. Exp. Biol. 2010, 213, 2417–2423. [Google Scholar] [CrossRef] [Green Version]
- Mcallister, J.C.; Adams, M.F. Mode of action for natural products isolated from essential oils of two trees is different from available mosquito adulticides. J. Med. Entomol. 2010, 47, 1123–1126. [Google Scholar] [CrossRef] [Green Version]
- Torres, M.A.; Jones, J.D.G.; Dangl, J.L. Reactive oxygen species signaling in response to pathogens. Plant Physiol. 2006, 141, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Torres, M.A.; Dangl, J.L.; Jones, J.D.G. Arabidopsis gp91phox homologues Atrbohd and Atrbohf are required for accumulation of reactive oxygen intermediates in the plant defense response. Proc. Natl. Acad. Sci. USA 2002, 99, 517–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melillo, M.T.; Leonetti, P.; Bongiovanni, M.; Castagnone-Sereno, P.; Bleve-Zacheo, T. Modulation of reactive oxygen species activities and H2O 2 accumulation during compatible and incompatible tomato-root-knot nematode interactions. New Phytol. 2006, 170, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Sanchez-Moreiras, A.M.; Abel, C.; Sohrabi, R.; Lee, S.; Gershenzon, J.; Tholl, D. The major volatile organic compound emitted from Arabidopsis thaliana flowers, the sesquiterpene (E)-β-caryophyllene, is a defense against a bacterial pathogen. New Phytol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tholl, D.; D´Auria, J.C.; Farooq, A.; Pichersky, E.; Gershenzon, J. Biosynthesis and emission of terpenoid volatiles from Arabidopsis flowers. Plant Cell. 2003, 15, 481–494. [Google Scholar] [CrossRef] [Green Version]
- Wubben, M.J.E.; Jin, J.; Baum, T.J. Cyst nematode parasitism of Arabidopsis thaliana is inhibited by salicylic acid (SA) and elicits uncoupled SA-independent pathogenesis-related gene expression in roots. Mol. Plant Microb. Interact. 2008, 21, 424–432. [Google Scholar] [CrossRef] [Green Version]
- Kfoury, M.; Landy, D.; Ruellan, S.; Auezova, L.; Greige-Gerges, H.; Fourmentin, S. Nootkatone encapsulation by cyclodextrins: Effect on water solubility and photostability. Food Chem. 2017, 236, 41–48. [Google Scholar] [CrossRef]
- Bharadwaj, A.; Stafford, K.C., III; Behle, R.W. Efficacy and Environmental Persistence of Nootkatone for the Control of the Blacklegged Tick (Acari: Ixodidae) in Residential Landscapes. J. Med. Entomol. 2012, 49, 1035–1044. [Google Scholar] [CrossRef]
- Troost, K.; Loeschcke, A.; Hilgers, F.; Özgür, A.Y.; Weber, T.M.; Santiago-Schübel, B.; Svensson, V.; Hage-Hülsmann, J.; Habash, S.S.; Grundler, F.M.W.; et al. Engineered Rhodobacter capsulatus as a Phototrophic Platform Organism for the Synthesis of Plant Sesquiterpenoids. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Habash, S.S.; Sobczak, M.; Siddique, S.; Voigt, B.; Elashry, A.; Grundler, F.M.W. Identification and characterization of a putative protein disulfide isomerase (HsPDI) as an alleged effector of Heterodera schachtii. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Prince, D.C.; Drurey, C.; Zipfel, C.; Hogenhout, S.A. The leucine-rich repeat receptor-like kinase brassinosteroid insensitive 1-associated kinase 1 and the cytochrome P450 phytoalexin deficient 3 contribute to innate immunity to aphids in Arabidopsis. Plant Physiol. 2014, 164, 2207–2219. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucl. Acid. Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Welke, J.E.; Manfroi, V.; Zanus, M.; Lazarotto, M.; Alcaraz Zini, C. Characterization of the volatile profile of Brazilian Merlot wines through comprehensive two dimensional gas chromatography time-of-flight mass spectrometric detection. J. Chromatogr. A 2012, 1226, 124–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound a | Ib | I (lit.) c | Identification d |
|---|---|---|---|---|
| 12 | α-cedrene | 1586 | 1582 [21] | ms (936, 942), ri, std |
| 15 | α-barbatene | 1596 | 1572 [21] | ms (844, 864), ri |
| 5 | β-elemene | 1600 | 1605 [22] | ms (876, 876), ri, std |
| 13 | β-cedrene | 1607 | 1599 [23] | ms (820, 834), ri |
| 6 | (E)-β-caryophyllene | 1613 | 1619 [22] | ms (909, 913), ri, std |
| 14 | cis-thujopsene | 1640 | 1620 [24] | ms (924, 927), ri |
| 2 | (E)-β-farnesene | 1666 | 1666 [25] | ms (913, 923), ri |
| 16 | β-barbatene | 1669 | n.a. e | ms (874, 876) |
| 10 | 4-epi-α-acoradiene | 1692 | n.a. e | ms (885, 895) |
| 9 | β-chamigrene | 1722 | 1740 [26] | ms (888, 905), ri |
| 3 | β-bisabolene * | 1734 | 1729 [21] | ms (889, 893), ri |
| 1 | (E,E)-α-farnesene * | 1750 | 1747 [27] | ms (902, 922), ri |
| 8 | α-chamigrene | 1755 | 1753 [21] | ms (845, 894), ri |
| 4 | α-curcumene | 1780 | 1780 [22] | ms (876, 892), ri |
| 7 | Nootkatene | 1829 | 1815 [24] | ms (872, 887), ri |
| 11 | Cuparene | 1838 | 1825 [24] | ms (906, 915), ri |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habash, S.S.; Könen, P.P.; Loeschcke, A.; Wüst, M.; Jaeger, K.-E.; Drepper, T.; Grundler, F.M.W.; Schleker, A.S.S. The Plant Sesquiterpene Nootkatone Efficiently Reduces Heterodera schachtii Parasitism by Activating Plant Defense. Int. J. Mol. Sci. 2020, 21, 9627. https://doi.org/10.3390/ijms21249627
Habash SS, Könen PP, Loeschcke A, Wüst M, Jaeger K-E, Drepper T, Grundler FMW, Schleker ASS. The Plant Sesquiterpene Nootkatone Efficiently Reduces Heterodera schachtii Parasitism by Activating Plant Defense. International Journal of Molecular Sciences. 2020; 21(24):9627. https://doi.org/10.3390/ijms21249627
Chicago/Turabian StyleHabash, Samer S., Philipp P. Könen, Anita Loeschcke, Matthias Wüst, Karl-Erich Jaeger, Thomas Drepper, Florian M. W. Grundler, and A. Sylvia S. Schleker. 2020. "The Plant Sesquiterpene Nootkatone Efficiently Reduces Heterodera schachtii Parasitism by Activating Plant Defense" International Journal of Molecular Sciences 21, no. 24: 9627. https://doi.org/10.3390/ijms21249627
APA StyleHabash, S. S., Könen, P. P., Loeschcke, A., Wüst, M., Jaeger, K.-E., Drepper, T., Grundler, F. M. W., & Schleker, A. S. S. (2020). The Plant Sesquiterpene Nootkatone Efficiently Reduces Heterodera schachtii Parasitism by Activating Plant Defense. International Journal of Molecular Sciences, 21(24), 9627. https://doi.org/10.3390/ijms21249627