The Emerging Role of Mechanosensitive Piezo Channels in Migraine Pain



Abstract
:1. Introduction
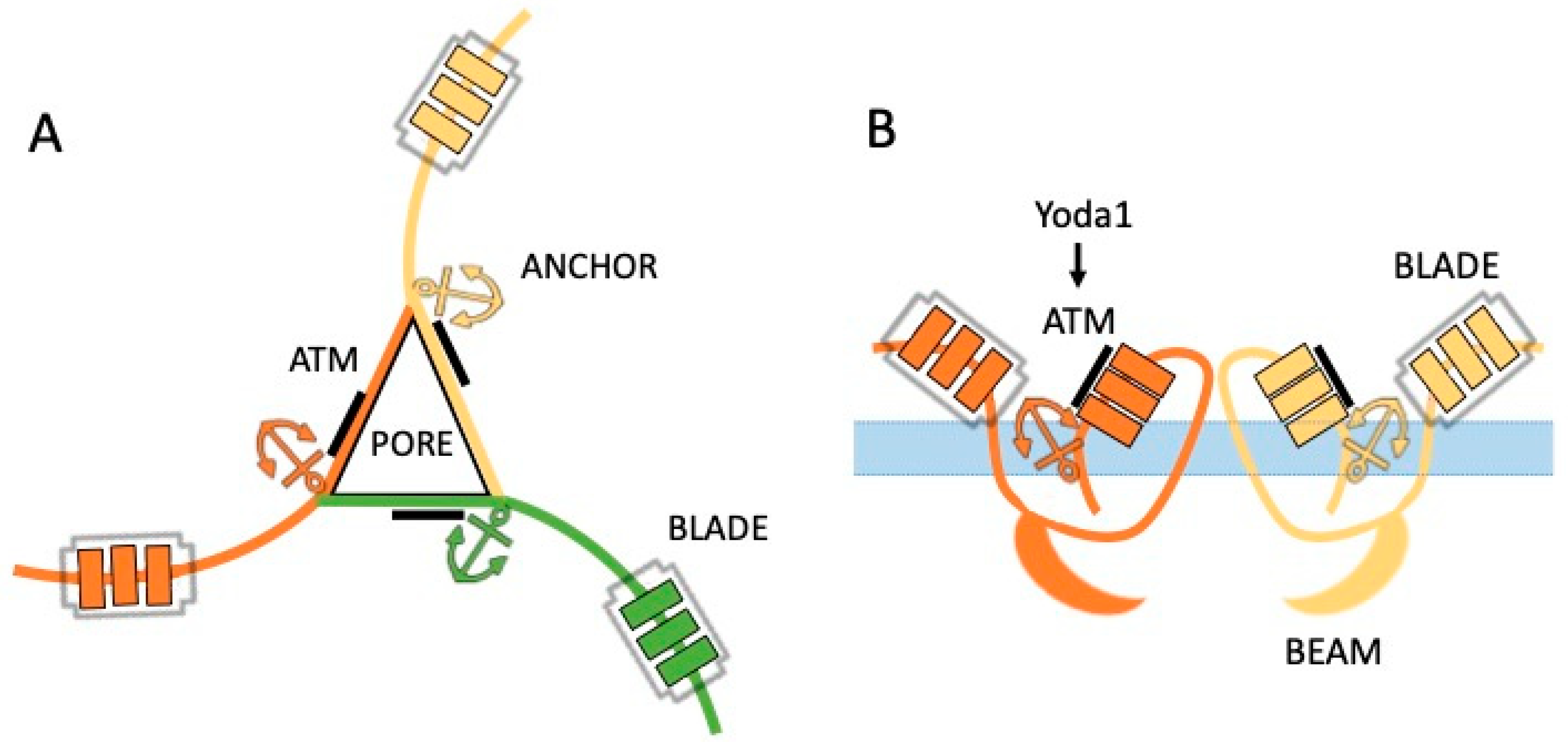
2. Complex Structure of Gigantic Piezo Channels
3. Functional Properties and the Role of Piezo1 vs. Piezo2 in Nociception
4. Unusual Chemical Activation of Piezo Channels
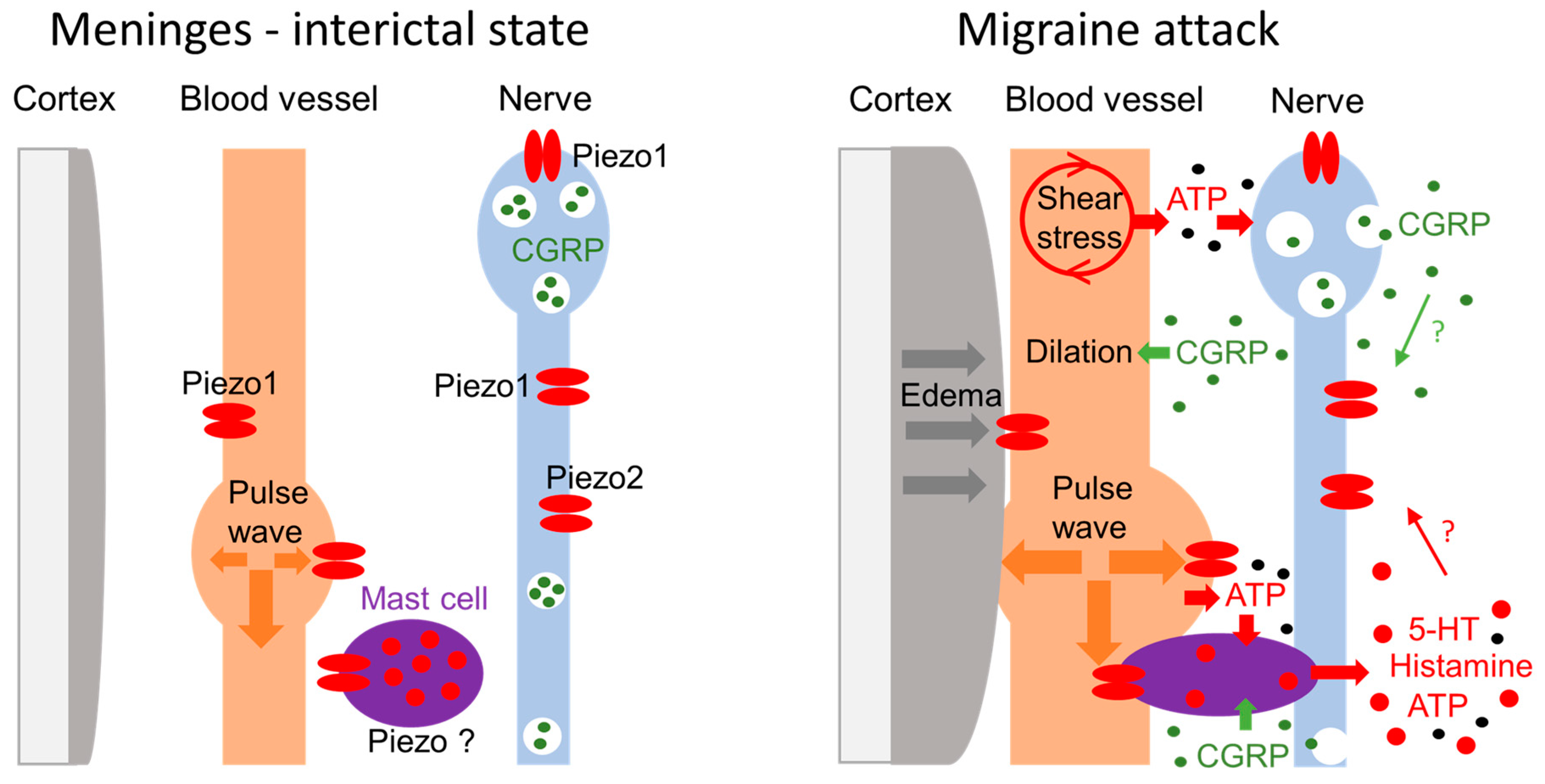
5. Puzzling Phenomenon of Pulsatile Pain: Role of Piezo?
6. New Model of Mechanosensation in Meninges during Migraine Attack
7. How to Alleviate Migraine Pain Through Piezo Channels?
8. Summary and Outlook
Funding
Conflicts of Interest
Abbreviations
| MSCs | Mechanosensitive ion channels |
| K2P | Two-pore-domain potassium channels |
| TMEM63 | Transmembrane protein 63 |
| OSCA | Hyperosmolality-gated calcium-permeable channels |
| TMC1/2 | Transmembrane channel-like protein 1/2 |
| DRG | Dorsal Root Ganglion |
| CSD | Cortical spreading depression |
| CGRP | Calcitonin gene-related peptide |
| NO | Nitric Oxides |
| Epac | Exchange protein activated by cAMP |
| cAMP | Cyclic Adenosine Monophosphate |
| TGVS | Trigeminovascular system |
References
- Douguet, D.; Honore, E. Mammalian mechanoelectrical transduction: Structure and function of force-gated ion channels. Cell 2019, 179, 340–354. [Google Scholar] [CrossRef]
- Benatto, M.T.; Florencio, L.L.; Carvalho, G.F.; Dach, F.; Bigal, M.E.; Chaves, T.C.; Bevilaqua-grossi, D. Cutaneous allodynia is more frequent in chronic migraine, and its presence and severity seems to be more associated with the duration of the disease. Arq. Neuropsiquiatr. 2017, 75, 153–159. [Google Scholar] [CrossRef] [Green Version]
- LaPaglia, D.M.; Sapio, M.R.; Burbelo, P.D.; Thierry-Mieg, J.; Thierry-Mieg, D.; Raithel, S.J.; Ramsden, C.E.; Iadarola, M.J.; Mannes, A.J. RNA-Seq investigations of human post-mortem trigeminal ganglia. Cephalalgia 2018, 38, 912–932. [Google Scholar] [CrossRef]
- Nguyen, M.Q.; Wu, Y.; Bonilla, L.S.; Von Buchholtz, L.J.; Nicholas, J.; Ryba, P. Diversity amongst trigeminal neurons revealed by high throughput single cell sequencing. PLoS ONE 2017, 12, e0185543. [Google Scholar] [CrossRef] [Green Version]
- Mikhailov, N.; Leskinen, J.; Fagerlund, I.; Poguzhelskaya, E.; Giniatullina, R.; Gafurov, O.; Malm, T.; Karjalainen, T.; Gröhn, O.; Giniatullin, R. Mechanosensitive meningeal nociception via Piezo channels: Implications for pulsatile pain in migraine? Neuropharmacology 2019, 149, 113–123. [Google Scholar] [CrossRef]
- Bagriantsev, S.N.; Gracheva, E.O.; Gallagher, P.G. Piezo proteins: Regulators of mechanosensation and other cellular processes. J. Biol. Chem. 2014, 289, 31673–31681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Zhou, H.; Chi, S.; Wang, Y.; Wang, J.; Geng, J.; Wu, K.; Liu, W.; Zhang, T.; Dong, M.Q.; et al. Structure and mechanogating mechanism of the Piezo1 channel. Nature 2018, 554, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottlieb, P.A. A tour de force: The discovery, properties, and function of Piezo channels. Curr. Top. Membr. 2017, 79, 1–36. [Google Scholar] [PubMed]
- Wang, Y.; Chi, S.; Guo, H.; Li, G.; Wang, L.; Zhao, Q.; Rao, Y.; Zu, L.; He, W.; Xiao, B. A lever-like transduction pathway for long-distance chemical- and mechano-gating of the mechanosensitive Piezo1 channel. Nat. Commun. 2018, 9, 1300. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, P.A.; Sachs, F. Properties of a cation selective mechanical channel. Landes Bioscence 2012, 6, 1–6. [Google Scholar]
- Anderson, E.O.; Schneider, E.R.; Bagriantsev, S.N. Piezo2 in cutaneous and proprioceptive mechanotransduction in vertebrates. Curr. Top. Membr. 2017, 79, 197–217. [Google Scholar]
- Ge, J.; Li, W.; Zhao, Q.; Li, N.; Chen, M.; Zhi, P.; Li, R.; Gao, N.; Xiao, B.; Yang, M. Architecture of the mammalian mechanosensitive Piezo1 channel. Nature 2015, 527, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hou, B.; Tumova, S.; Muraki, K.; Bruns, A.; Ludlow, M.J.; Sedo, A.; Hyman, A.J.; McKeown, L.; Young, R.S.; et al. Piezo1 integration of vascular architecture with physiological force. Nature 2014, 515, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.H.; Cui, A.F.; McDonald, M.F.; Grandl, J. Transduction of repetitive mechanical stimuli by Piezo1 and Piezo2 ion channels. Cell Rep. 2017, 19, 2572–2585. [Google Scholar] [CrossRef] [Green Version]
- Romero, L.O.; Massey, A.E.; Mata-Daboin, A.D.; Sierra-Valdez, F.J.; Chauhan, S.C.; Cordero-Morales, J.F.; Vásquez, V. Dietary fatty acids fine-tune Piezo1 mechanical response. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Ranade, S.S.; Woo, S.; Dubin, A.E.; Moshourab, R.A.; Wetzel, C.; Petrus, M.; Mathur, J.; Bégay, V.; Coste, B.; Mainquist, J.; et al. Piezo2 is the major transducer of mechanical forces for touch sensation in mice. Nature 2014, 516, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; La, J.; Hamill, O.P. Piezo1 is selectively expressed in small diameter mouse DRG neurons distinct from neurons strongly expressing TRPV1. Front. Mol. Neurosci. 2019, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Wang, Y.; Geng, J.; Zhou, S.; Xiao, B. Mechanically activated Piezo channels mediate touch and suppress acute mechanical pain response in mice. Cell Rep. 2019, 26, 1419–1431. [Google Scholar] [CrossRef] [Green Version]
- Bolay, H.; Reuter, U.; Dunn, A.K.; Huang, Z.; Boas, D.A.; Moskowitz, M.A. Intrinsic brain activity triggers trigeminal meningeal afferents in a migraine model. Nat. Med. 2002, 8, 136–142. [Google Scholar] [CrossRef]
- Messlinger, K. Migraine: Where and how does the pain originate? Exp. Brain Res. 2009, 196, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wei, X.; Bischoff, C.; Edelmayer, R.M.; Dussor, G. Research submission pH-evoked dural afferent signaling is mediated by ASIC3 and is sensitized by mast cell mediators. Headache 2013, 53, 1250–1261. [Google Scholar] [CrossRef] [Green Version]
- Noseda, R.; Burstein, R. Migraine pathophysiology: Anatomy of the trigeminovascular pathway and associated neurological symptoms, CSD, sensitization and modulation of pain. Pain 2013, 154, 44–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syeda, R.; Xu, J.; Dubin, A.E.; Coste, B.; Mathur, J.; Huynh, T.; Matzen, J.; Lao, J.; Tully, D.C.; Engels, I.H.; et al. Chemical activation of the mechanotransduction channel Piezo1. Elife 2015, 4, 1–11. [Google Scholar] [CrossRef]
- Lacroix, J.J.; Botello-Smith, W.M.; Luo, Y. Probing the gating mechanism of the mechanosensitive channel Piezo1 with the small molecule Yoda. Nat. Commun. 2018, 9, 2029. [Google Scholar] [CrossRef] [Green Version]
- Botello-smith, W.M.; Jiang, W.; Lacroix, J.J.; Luo, Y. A mechanism for the activation of the mechanosensitive Piezo1 channel by the small molecule Yoda1. Nat. Commun. 2019, 10, 4503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabbretti, E.; Arco, M.D.; Fabbro, A.; Simonetti, M.; Nistri, A.; Giniatullin, R. Delayed upregulation of ATP P2X 3 receptors of trigeminal sensory neurons by calcitonin gene-related peptide. J. Neurosci. 2006, 26, 6163–6171. [Google Scholar] [CrossRef] [Green Version]
- D’Arco, M.; Giniatullin, R.; Simonetti, M.; Fabbro, A.; Nair, A.; Nistri, A. Neutralization of nerve growth factor induces plasticity of ATP-sensitive P2X 3 receptors of nociceptive trigeminal ganglion neurons. J. Neurosci. 2007, 27, 8190–8201. [Google Scholar] [CrossRef] [PubMed]
- Giniatullin, R.; Nistri, A.; Fabbretti, E. Molecular mechanisms of sensitization of pain-transducing P2X 3 receptors by the migraine mediators CGRP and NGF. Mol. Neurobiol. 2008, 37, 83–90. [Google Scholar] [CrossRef]
- Vincent, M.; Wang, S. Headache classification committee of the International Headache Society (IHS) The International Classification of Headache Disorders, 3rd edition. Cephalalgia 2018, 38, 1–211. [Google Scholar]
- Wolff, H.G. Wolff’s Headache and Other Head Pain, 1st ed.; Oxford University Press: Oxford, UK, 1948. [Google Scholar]
- Edvinsson, L. CGRP as the target of new migraine therapies—Successful translation from bench to clinic. Nat. Rev. Neurol. 2018, 14, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, R.; Olesen, J. Calcitonin gene-related peptide modulators–The history and renaissance of a new migraine drug class. Headache Curr. 2019, 59, 951–970. [Google Scholar] [CrossRef] [PubMed]
- Waeber, C.; Moskowitz, M.A. Migraine as an inflammatory disorder. Neurology 2005, 64, S9–S15. [Google Scholar] [CrossRef] [PubMed]
- Rahmann, A.; Wienecke, T.; Hansen, J.M.; Fahrenkrug, J.; Olesen, J.; Ashina, M. Vasoactive intestinal peptide causes marked cephalic vasodilation, but does not induce migraine. Cephalalgia 2008, 28, 226–236. [Google Scholar] [CrossRef]
- Zakharov, A.; Vitale, C.; Kilinc, E.; Koroleva, K.; Fayuk, D.; Shelukhina, I. Hunting for origins of migraine pain: Cluster analysis of spontaneous and capsaicin-induced firing in meningeal trigeminal nerve fibers. Front. Cell. Neurosci. 2015, 9, 1–14. [Google Scholar] [CrossRef]
- Yegutkin, G.G.; Guerrero-toro, C.; Kilinc, E.; Koroleva, K. Nucleotide homeostasis and purinergic nociceptive signaling in rat meninges in migraine-like conditions. Purinergic Signal. 2016, 12, 561–574. [Google Scholar] [CrossRef] [Green Version]
- Levy, D. Migraine pain, meningeal inflammation, and mast cells. Curr. Pain Headache Rep. 2009, 13, 237–240. [Google Scholar] [CrossRef]
- Mekori, Y.A.; Metcalfe, D.D. Mast cells in innate immunity. Immunol. Rev. 2000, 173, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Demartini, C.; Greco, R.; Maria, A.; Sances, G.; De Icco, R.; Borsook, D.; Tassorelli, C.; Foundation, I.C.M. Progress in neurobiology nitroglycerin as a comparative experimental model of migraine pain: From animal to human and back. Prog. Neurobiol. 2019, 177, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Strassman, A.M.; Raymond, S.A.; Burstein, R. Sensitization of meningeal sensory neurons and the origin of headaches. Nature 1996, 384, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Kilinc, E.; Guerrero-toro, C.; Zakharov, A.; Vitale, C.; Gubert-olive, M.; Koroleva, K.; Timonina, A.; Luz, L.L.; Shelukhina, I.; Giniatullina, R.; et al. Neuropharmacology serotonergic mechanisms of trigeminal meningeal nociception: Implications for migraine pain. Neuropharmacology 2017, 116, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Koroleva, K.; Gafurov, O.; Guselnikova, V.; Nurkhametova, D. Meningeal mast cells contribute to ATP-induced nociceptive firing in trigeminal nerve terminals: Direct and indirect purinergic mechanisms triggering migraine pain. Front. Cell. Neurosci. 2019, 13, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Burstein, R.; Yamamura, H.; Malick, A.; Strassman, A.M. Chemical stimulation of the intracranial dura induces enhanced responses to facial stimulation in brain stem trigeminal neurons. J. Neurophysiol. 2017, 79, 964–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Chennupati, R.; Kaur, H.; Iring, A.; Wettschureck, N.; Offermanns, S. Endothelial cation channel PIEZO1 controls blood pressure by mediating flow-induced ATP release. J. Clin. Investig. 2016, 126, 4527–4536. [Google Scholar] [CrossRef] [PubMed]
- Somjen, G.G. Mechanisms of spreading depression and hypoxic spreading depression-like depolarization. Physiol. Rev. 2001, 81, 1065–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aristides, A.P.L. Spreading depression of activity in the cerebral cortex. J. Neurophysiol. 1944, 7, 359–390. [Google Scholar]
- Karatas, H.; Erdener, S.E.; Gursoy-Ozdemir, Y.; Lule, S.; Eren-Koçak, E.; Sen, Z.D.; Dalkara, T. Spreading depression triggers headache by activating neuronal Panx1 channels. Science 2013, 350, 1092–1096. [Google Scholar] [CrossRef]
- Anderson, T.R.; Andrew, R.D. Spreading depression: Imaging and blockade in the rat neocortical brain slice. J. Neurophysiol. 2002, 88, 2713–2725. [Google Scholar] [CrossRef] [Green Version]
- Viola, S.; Viola, P.; Buongarzone, M.P.; Fiorelli, L.; Litterio, P. The increased distensibility of the wall of cerebral arterial network may play a role in the pathogenic mechanism of migraine headache. Neurol. Sci. 2014, 35, 163–166. [Google Scholar] [CrossRef]
- Ridone, P.; Vassalli, M.; Martinac, B. Piezo1 mechanosensitive channels: What are they and why are they important. Biophys. Rev. 2019, 11, 795–805. [Google Scholar] [CrossRef] [Green Version]
- Ranade, S.S.; Qiu, Z.; Woo, S.-H.; Hur, S.S.; Murthy, S.E.; Cahalan, S.M.; Xu, J.; Mathur, J.; Bandell, M.; Coste, B.; et al. Piezo1, a mechanically activated ion channel, is required for vascular development in mice. Proc. Natl. Acad. Sci. USA 2014, 111, 10347–10352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borbiro, I.; Rohacs, T. Regulation of Piezo channels by cellular signaling pathways. Curr. Top. Membr. 2017, 79, 245–261. [Google Scholar] [PubMed]
- Eijkelkamp, N.; Linley, J.E.; Torres, J.M.; Bee, L.; Dickenson, A.H.; Gringhuis, M.; Minett, M.S. A role for Piezo2 in EPAC1-dependent mechanical allodynia. Nat. Commun. 2013, 4, 1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Agonists | Antagonists | Modulators | ||||
|---|---|---|---|---|---|---|
| Yoda1 | Jedi1/2 | Dooku1 | Gadolinium | GsMTx4 | Margaric acid (saturated) | Docosahexaenoic acid (unsaturated) |
| Selective | Selective | Nonselective | Nonselective | |||
| Acting sites | ||||||
| ATM region in C-terminus [24,25] | L15-16/L19-20 regions [10] | Yoda1 binding site [51] | Ion channel pore [51] | Changes in membrane lipid environment [16] | ||
| accelerated inactivation | reduced inactivation | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Della Pietra, A.; Mikhailov, N.; Giniatullin, R. The Emerging Role of Mechanosensitive Piezo Channels in Migraine Pain. Int. J. Mol. Sci. 2020, 21, 696. https://doi.org/10.3390/ijms21030696
Della Pietra A, Mikhailov N, Giniatullin R. The Emerging Role of Mechanosensitive Piezo Channels in Migraine Pain. International Journal of Molecular Sciences. 2020; 21(3):696. https://doi.org/10.3390/ijms21030696
Chicago/Turabian StyleDella Pietra, Adriana, Nikita Mikhailov, and Rashid Giniatullin. 2020. "The Emerging Role of Mechanosensitive Piezo Channels in Migraine Pain" International Journal of Molecular Sciences 21, no. 3: 696. https://doi.org/10.3390/ijms21030696
APA StyleDella Pietra, A., Mikhailov, N., & Giniatullin, R. (2020). The Emerging Role of Mechanosensitive Piezo Channels in Migraine Pain. International Journal of Molecular Sciences, 21(3), 696. https://doi.org/10.3390/ijms21030696