Genome-Wide Identification of Metal Tolerance Protein Genes in Populus trichocarpa and Their Roles in Response to Various Heavy Metal Stresses

Abstract
:1. Introduction
2. Results
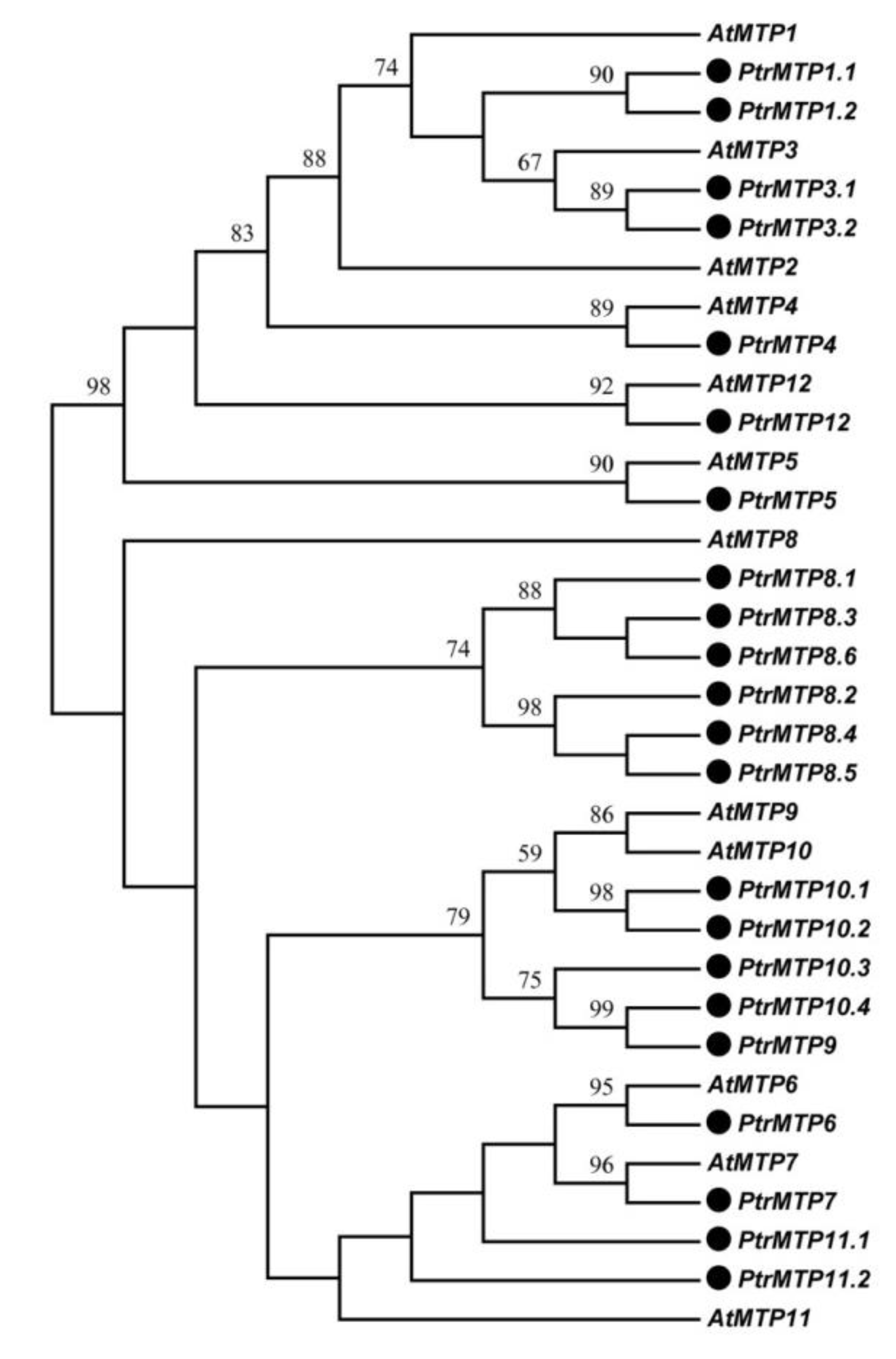
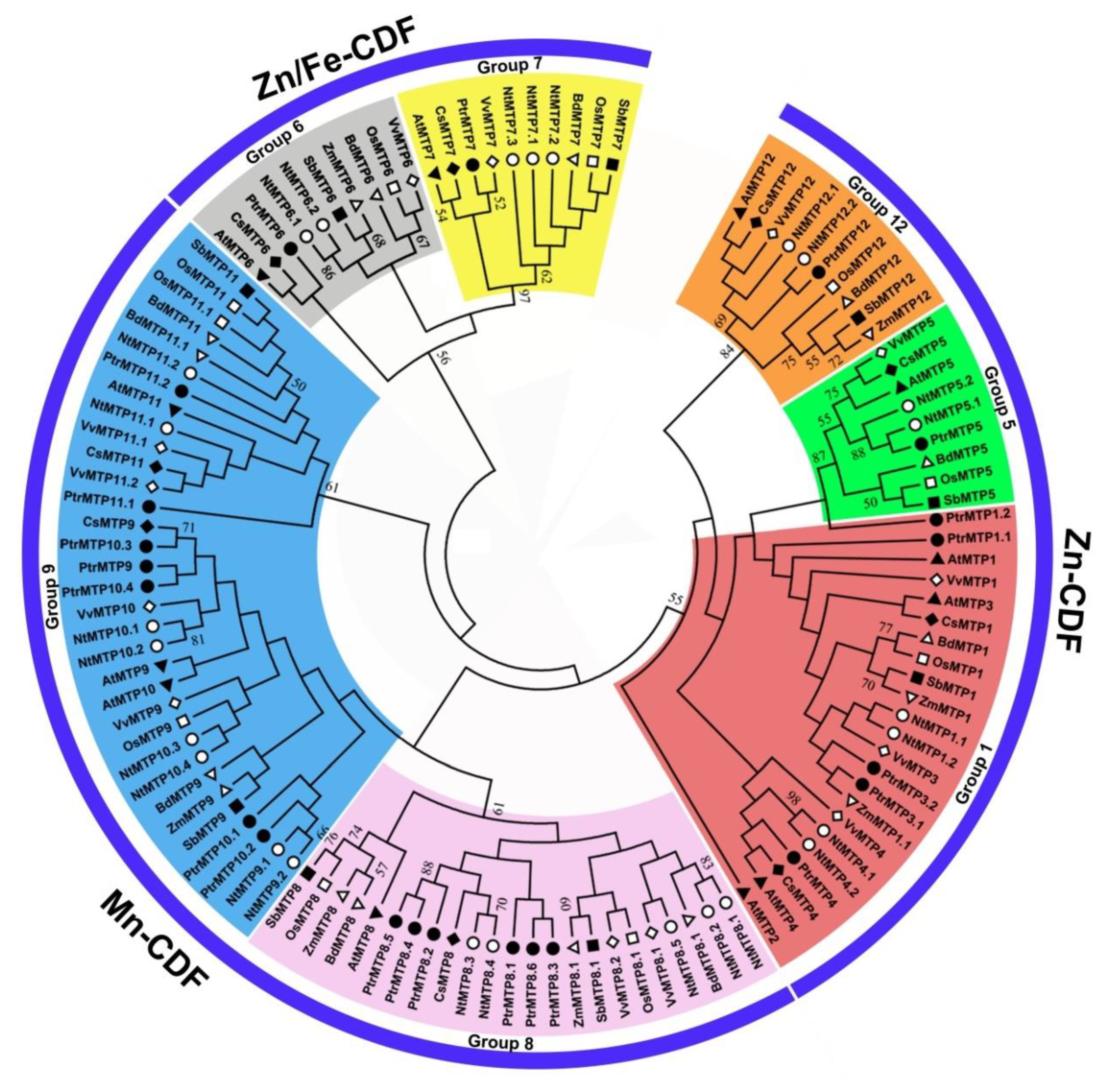
2.1. Identification and Classification of MTP Genes in P. trichocarpa Genome
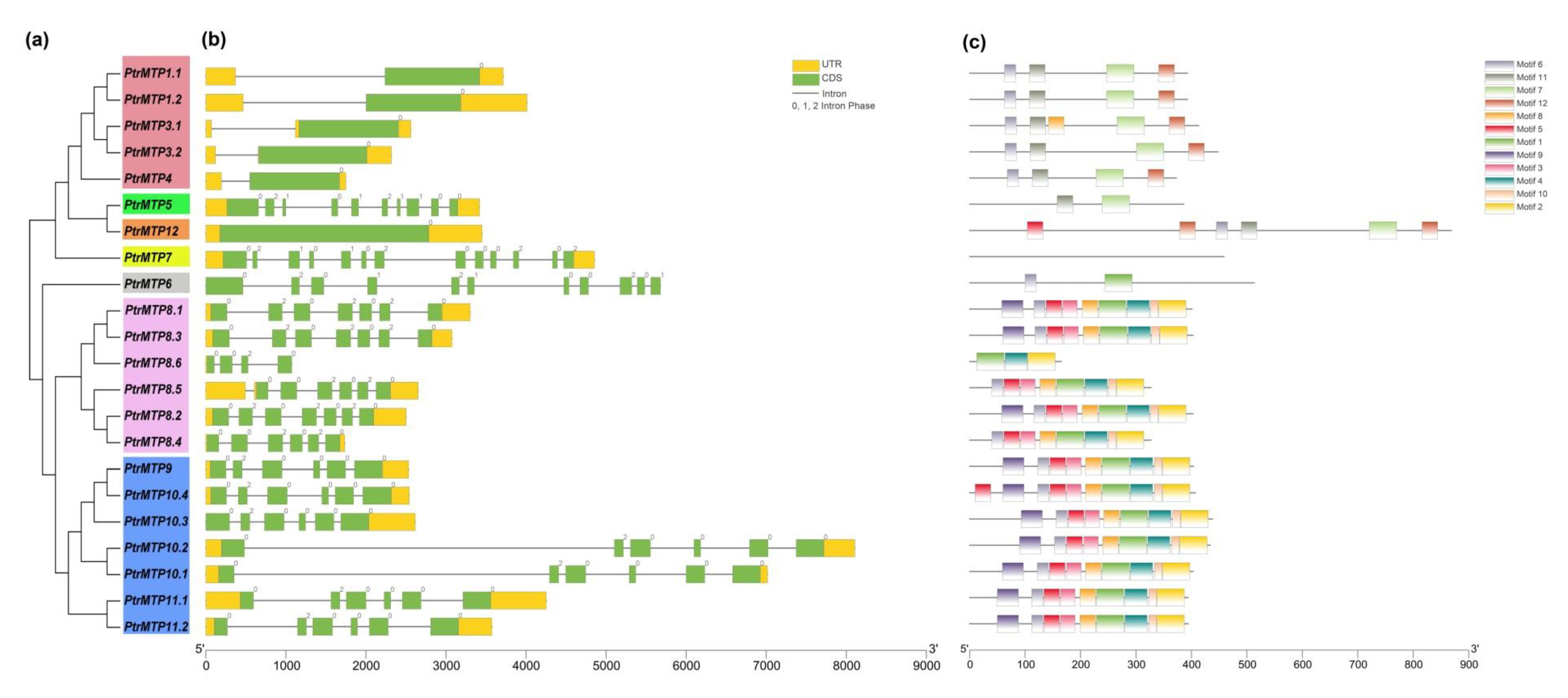
2.2. Structure and Characteristics Analysis of PtrMTP Genes
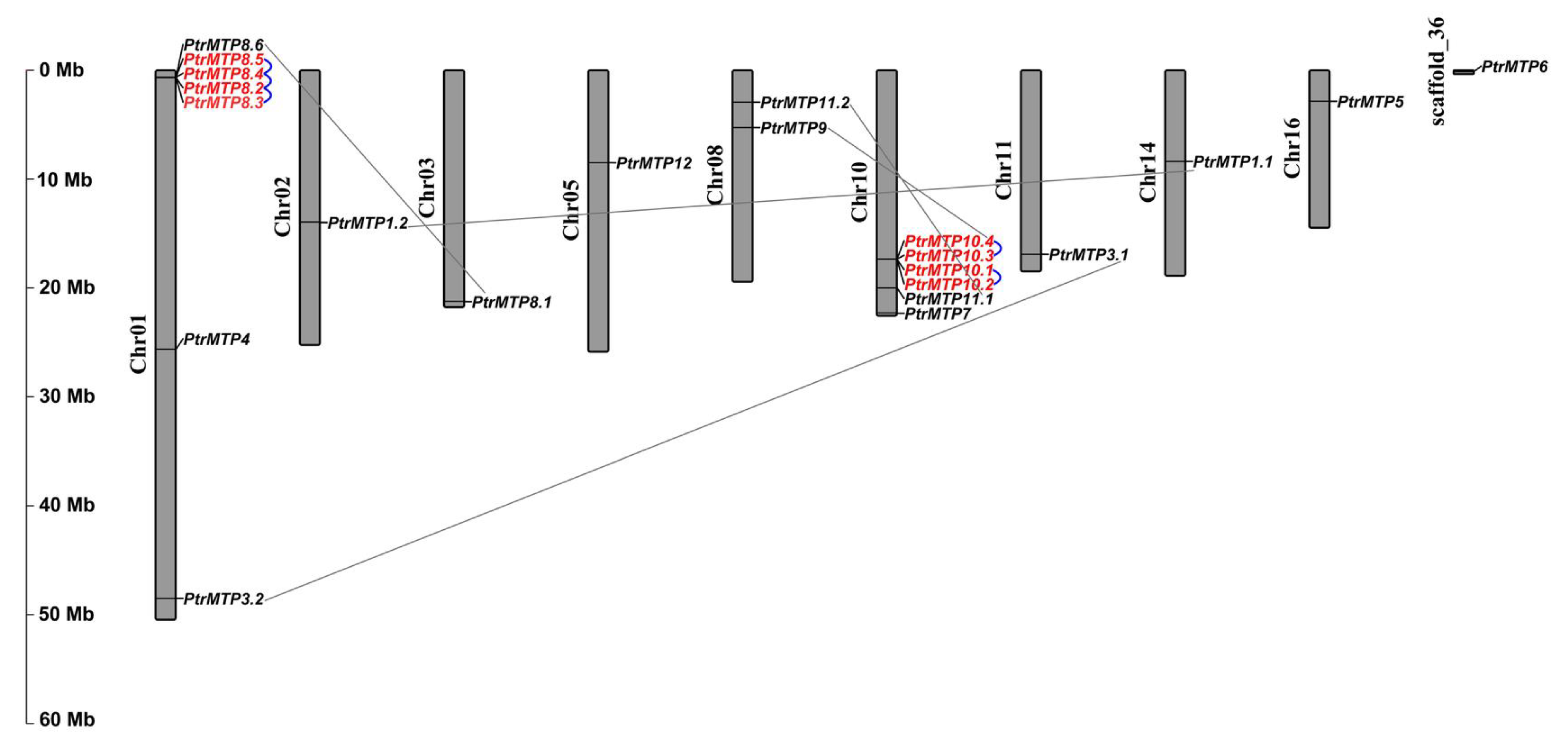
2.3. Chromosomal Localization and Gene Duplication Analysis of PtrMTP Genes
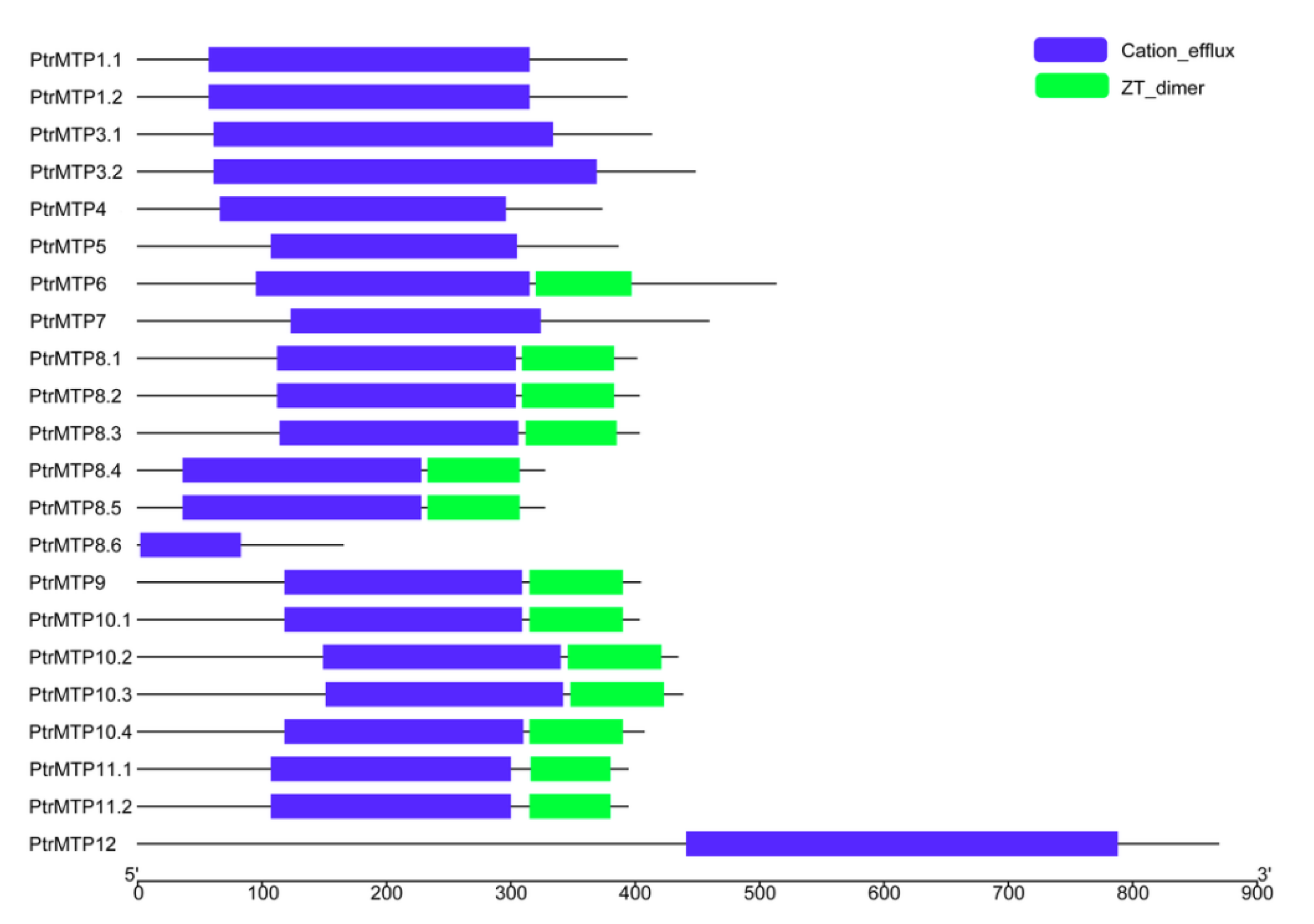
2.4. Conserved Motif and Domain Architectures Analysis of PtrMTP Proteins
2.5. Cis-Acting Elements in the Promoter Regions of PtrMTP Genes
2.6. Potential miRNA Target Sites in PtrMTP Genes
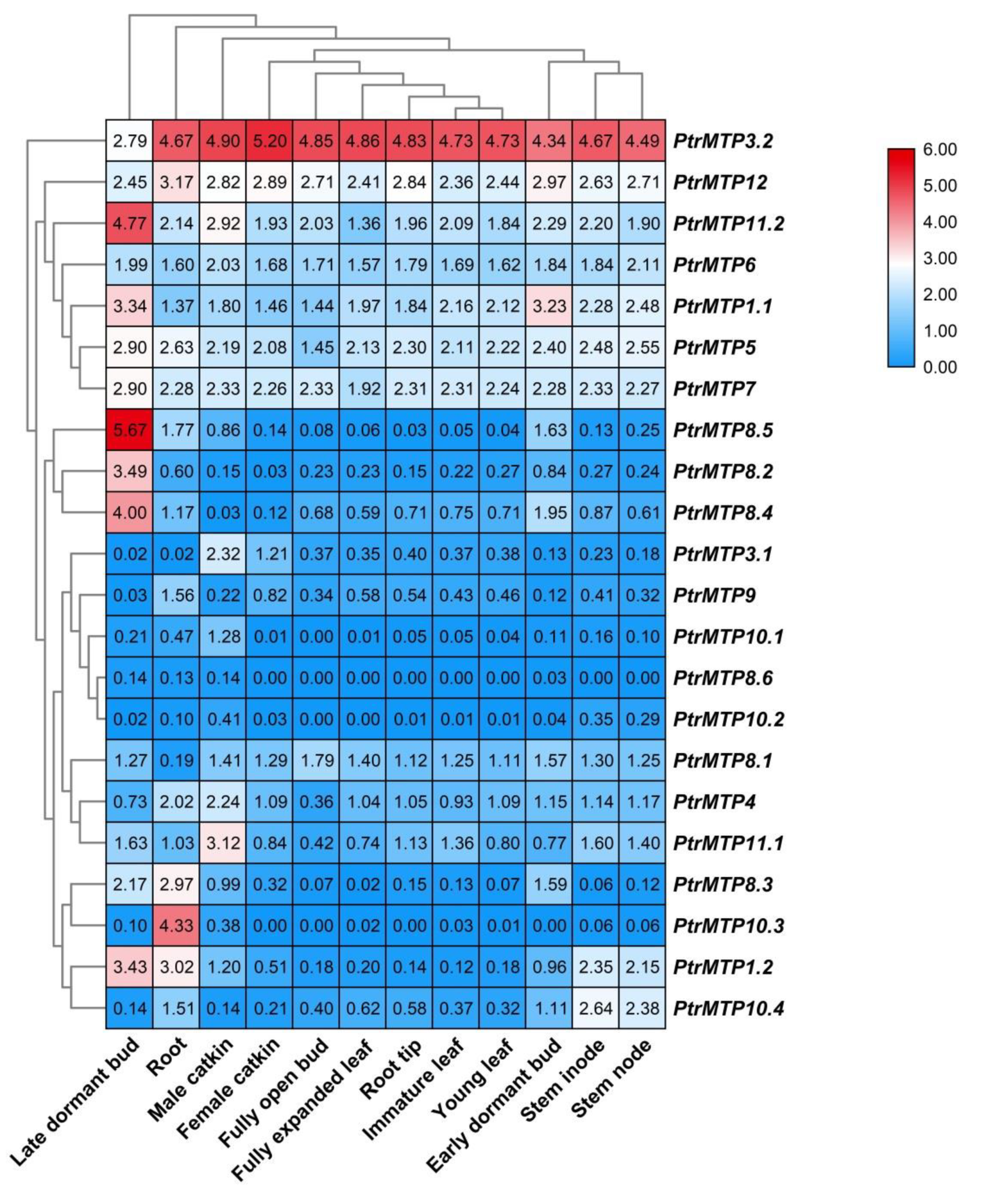
2.7. The Temporal and Spatial Expression Patterns of PtrMTP Genes
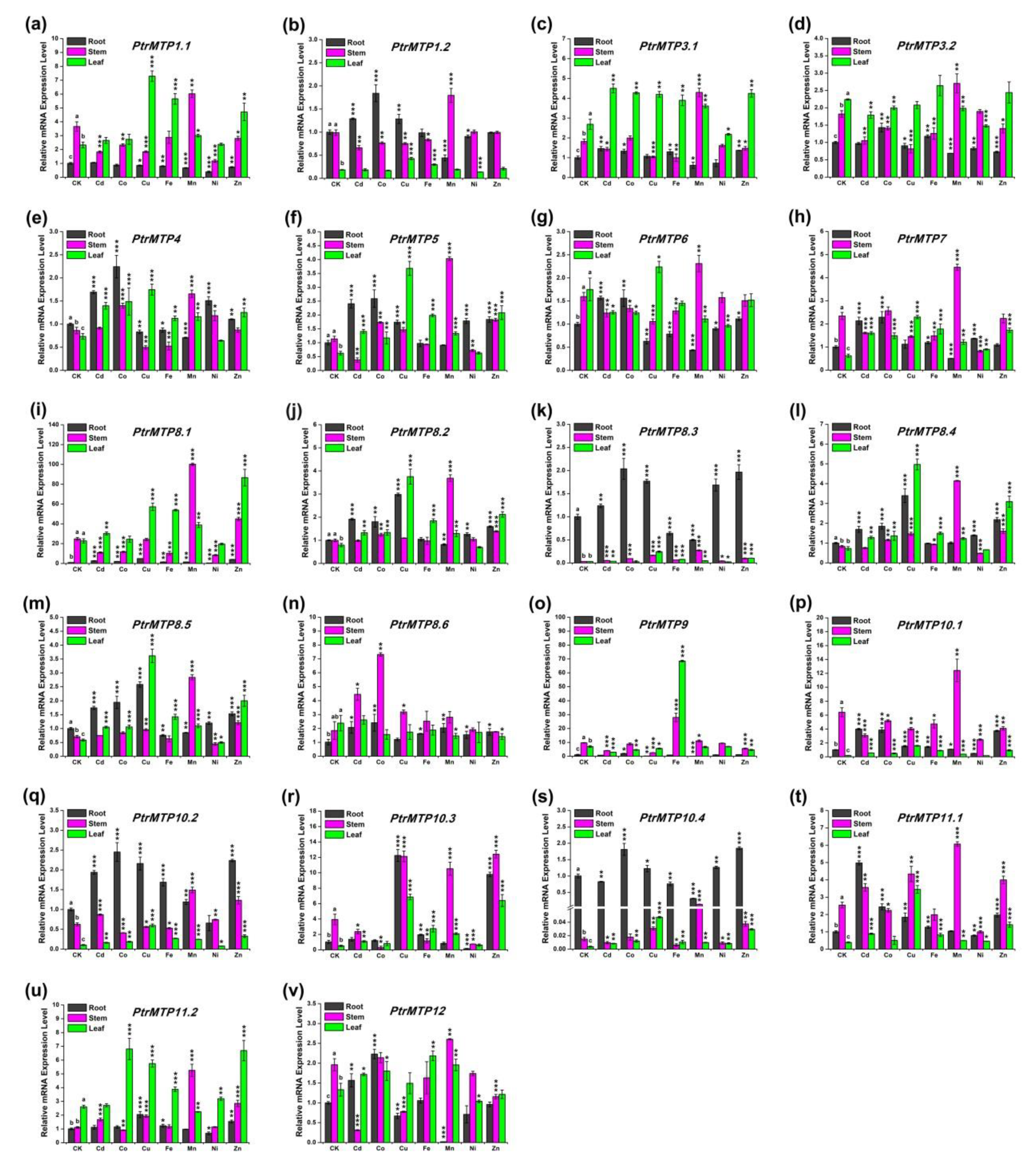
2.8. Expression Profiles of PtrMTPs under Different Heavy Metal Treatments
2.9. Effect of PtrMTP Genes on Yeast Growth
3. Discussion
3.1. Evolution and Differentiation of PtrMTP Genes as well as Their Proteins Architectures
3.2. Regulation of PtrMTP Gene Expression in P. trichocarpae
3.3. The Diverse Expression Patterns of PtrMTP Genes
3.4. Some PtrMTPs were Co, Fe and Mn Transporters in Yeast Cells
4. Materials and Methods
4.1. Identification and Phylogenetic Analysis of the MTPs in P. Trichocarpa
4.2. Analysis of Genomic Structure, Chromosomal Localization, Gene Duplication and Ka/Ks of MTPs in P. trichocarpa
4.3. Amino Acid Properties and Structure Characteristics of PtrMTP Proteins
4.4. Prediction of Cis-Acting Regulatory Elements and MicroRNA (miRNA) Target Sites of PtrMTP Genes
4.5. Tissue Expression Pattern Based on RNA-seq Data
4.6. Growth Condictions and Heavy Metal Treatments
4.7. RNA Extraction and qRT-PCR Analysis
4.8. Yeast Transformation and Growth Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MTP | Metal tolerance protein |
| CDFs | Cation diffusion facilitators |
| TMDs | Transmembrane domains |
| MW | Molecular weight |
| UTR | Untranslated regions |
| CDS | Coding sequence |
| pI | Isoelectric point |
| GRAVY | Grand average of hydropathicity |
| Chr | Chromosomes |
| Ka | The number of nonsynonymous substitutions per nonsynonymous site |
| Ks | The number of synonymous substitutions per synonymous site |
| FPKM | Fragments Per Kilobase of exon model per Million mapped reads |
| qRT-PCR | quantitative RealTime-PCR |
| SD | Synthetic drop-out |
References
- Vareda, J.P.; Valente, A.J.M.; Duraes, L. Assessment of heavy metal pollution from anthropogenic activities and remediation strategies: A review. J. Environ. Manag. 2019, 246, 101–118. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals—Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Clemens, S. Molecular mechanisms of plant metal tolerance and homeostasis. Planta 2000, 212, 475–486. [Google Scholar] [CrossRef]
- Hall, J.L.; Williams, L.E. Transition metal transporters in plants. J. Exp. Bot. 2003, 54, 2601–2613. [Google Scholar] [CrossRef]
- Montanini, B.; Blaudez, D.; Jeandroz, S.; Sanders, D.; Chalot, M. Phylogenetic and functional analysis of the Cation Diffusion Facilitator (CDF) family: Improved signature and prediction of substrate specificity. BMC Genom. 2007, 8, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nies, D.H.; Silver, S. Ion efflux system involved in bacterial metal resistances. J. Ind. Microbiol. 1995, 14, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, I.T.; Saier, M.H. A novel family of ubiquitous heavy metal ion transport proteins. J. Membr. Biol. 1997, 156, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Gustin, J.L.; Zanis, M.J.; Salt, D.E. Structure and evolution of the plant cation diffusion facilitator family of ion transporters. BMC Evol. Biol. 2011, 11, 76. [Google Scholar] [CrossRef] [Green Version]
- Kobae, Y.; Uemura, T.; Sato, M.H.; Ohnishi, M.; Mimura, T.; Nakagawa, T.; Maeshima, M. Zinc transporter of Arabidopsis thaliana AtMTP1 is localized to vacuolar membranes and implicated in zinc homeostasis. Plant Cell Physiol. 2004, 45, 1749–1758. [Google Scholar] [CrossRef] [Green Version]
- Desbrosses-Fonrouge, A.G.; Voigt, K.; Schröder, A.; Arrivault, S.; Thomine, S.; Krämer, U. Arabidopsis thaliana MTP1 is a Zn transporter in the vacuolar membrane which mediates Zn detoxification and drives leaf Zn accumulation. FEBS Lett. 2005, 579, 4165–4174. [Google Scholar] [CrossRef]
- Arrivault, S.; Senger, T.; Kramer, U. The Arabidopsis metal tolerance protein AtMTP3 maintains metal homeostasis by mediating Zn exclusion from the shoot under Fe deficiency and Zn oversupply. Plant J. 2006, 46, 861–879. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, T.; Kawachi, M.; Sato, Y.; Mori, H.; Kutsuna, N.; Hasezawa, S.; Maeshima, M. A high molecular mass zinc transporter MTP12 forms a functional heteromeric complex with MTP5 in the Golgi in Arabidopsis thaliana. FEBS J. 2015, 282, 965–1979. [Google Scholar] [CrossRef] [PubMed]
- Eroglu, S.; Meier, B.; vonWirén, N.; Peiter, E. The vacuolar manganese transporter MTP8 determines tolerance to iron deficiency-induced chlorosis in Arabidopsis. Plant Physiol. 2016, 170, 1030–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, H.H.; Car, S.; Socha, A.L.; Hindt, M.N.; Punshon, T.; Guerinot, M.L. The Arabidopsis MTP8 transporter determines the localization of manganese and iron in seeds. Sci. Rep. 2017, 7, 11024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eroglu, S.; Giehl, R.F.H.; Meier, B.; Takahashi, M.; Terada, Y.; Ignatyev, K.; Andresen, E.; Küpper, H.; Peiter, E.; von Wirén, N. Metal tolerance protein 8 mediates manganese homeostasis and iron reallocation during seed development and germination. Plant Physiol. 2017, 174, 1633–1647. [Google Scholar] [CrossRef] [Green Version]
- Delhaize, E.; Gruber, B.D.; Pittman, J.K.; White, R.G.; Leung, H.; Miao, Y.; Jiang, L.; Ryan, P.R.; Richardson, A.E. A role for the AtMTP11 gene of Arabidopsis in manganese transport and tolerance. Plant J. 2007, 51, 198–210. [Google Scholar] [CrossRef]
- Peiter, E.; Montanini, B.; Gobert, A.; Pedas, P.; Husted, S.; Maathuis, F.J.; Blaudez, D.; Chalot, M.; Sanders, D. A secretory pathway-localized cation diffusion facilitator confers plant manganese tolerance. Proc. Natl. Acad. Sci. USA 2007, 104, 8532–8537. [Google Scholar] [CrossRef] [Green Version]
- Yadav, R.; Arora, P.; Kumar, S.; Chaudhury, A. Perspectives for genetic engineering of poplars for enhanced phytoremediation abilities. Ecotoxicology 2010, 19, 1574–1588. [Google Scholar] [CrossRef]
- Blaudez, D.; Kohler, A.; Martin, F.; Sanders, D.; Chalot, M. Poplar metal tolerance protein 1 confers zinc tolerance and is an oligomeric vacuolar zinc transporter with an essential leucine zipper motif. Plant Cell 2003, 15, 2911–2928. [Google Scholar] [CrossRef] [Green Version]
- Migeon, A.; Blaudez, D.; Wilkins, O.; Montanini, B.; Campbell, M.M.; Richaud, P.; Thomine, S.; Chalot, M. Genome-wide analysis of plant metal transporters, with an emphasis on poplar. Cell. Mol. Life Sci. 2010, 67, 3763–3784. [Google Scholar] [CrossRef]
- Hurst, L.D. The Ka/Ks ratio: Diagnosing the form of sequence evolution. Trends Genet. 2002, 18, 486. [Google Scholar] [CrossRef]
- Migocka, M.; Papierniak, A.; Maciaszczyk-Dziubi’nska, E.; Po’zdzik, P.; Posyniak, E.; Garbiec, A.; Filleur, S. Cucumber metal transport protein MTP8 confers increased tolerance to manganese when expressed in yeast and Arabidopsis thaliana. J. Exp. Bot. 2014, 65, 5367–5384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatansever, R.; Filiz, E.; Froglu, S. Genome-wide exploration of metaltolerance protein (MTP) genes in common wheat (Triticum aestivum): Insightsinto metal homeostasis and biofortification. Biometals 2017, 30, 217–235. [Google Scholar] [CrossRef]
- Fu, X.; Tong, Y.; Zhou, X.; Ling, L.; Chun, C.; Cao, L.; Zeng, M.; Peng, L.Z. Genome-wide identification of sweet orange (Citrus sinensis) metal tolerance proteins and analysis of their expression patterns under zinc, manganese, copper, and cadmium toxicity. Gene 2017, 629, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, Y.; Li, B.; He, W.; Yang, Y.; Yang, Y. Genome-wide identification and expression analysis of the cation diffusion facilitator gene family in Turnip under diverse metal ion stresses. Front. Genet. 2018, 9, 103. [Google Scholar] [CrossRef]
- Liu, J.; Gao, Y.; Tang, Y.; Wang, D.; Chen, X.; Yao, Y.; Guo, Y. Genome-wide identification, comprehensive gene feature, evolution, and expression analysis of plant metal tolerance proteins in tobacco under heavy metal toxicity. Front. Genet. 2019, 24, 345. [Google Scholar] [CrossRef] [PubMed]
- Shirazi, Z.; Abedi, A.; Kordrostami, M.; Burritt, D.J.; Hossain, M.A. Genome-wide identification and characterization of the metal tolerance protein (MTP) family in grape (Vitis vinifera L.). 3 Biotech 2019, 9, 199. [Google Scholar] [CrossRef] [PubMed]
- Ram, H.; Kaur, A.; Gandass, N.; Singh, S.; Deshmukh, R.; Sonah, H.; Sharma, T.R. Molecular characterization and expression dynamics of MTP genes under various spatio-temporal stages and metal stress conditions in rice. PLoS ONE 2019, 14, e0217360. [Google Scholar] [CrossRef] [PubMed]
- Hittinger, C.T.; Carroll, S.B. Gene duplication and the adaptive evolution of a classic genetic switch. Nature 2007, 449, 677–681. [Google Scholar] [CrossRef]
- Hofmann, K.; Stoffel, W. TMbase-A database of membrane spanning proteins segments. Biol. Chem. Hoppe Seyler 1993, 374, 166–170. [Google Scholar]
- Lu, M.; Fu, D. Structure of the zinc transporter YiiP. Science 2007, 317, 1746–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laloum, T.; De Mita, S.; Gamas, P.; Baudin, M.; Niebel, A. CCAAT box binding transcription factors in plants: Y so many? Trends Plant. Sci. 2013, 18, 157–166. [Google Scholar] [CrossRef]
- Gielen, H.; Remans, T.; Vangronsveld, J.; Cuypers, A. MicroRNAs in metal stress: Specific roles or secondary responses? Int. J. Mol. Sci. 2012, 13, 15826–15847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barakat, A.; Sriram, A.; Park, J.; Zhebentyayeva, T.; Main, D.; Abbott, A. Genome wide identification of chilling responsive microRNAs in Prunus persica. BMC Genom. 2012, 13, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, F.; He, C.; Yan, X.; Bai, F.; Pan, Z.; Deng, X.; Xiao, S. Small RNA profiling reveals involvement of microRNA-mediated gene regulation in response to mycorrhizal symbiosis in Poncirus trifoliata L. Raf. Tree Genet. Genomes 2018, 14, 42. [Google Scholar] [CrossRef]
- Shuai, P.; Liang, D.; Zhang, Z.; Yin, W.; Xia, X. Identification of drought-responsive and novel Populus trichocarpa microRNAs by high-throughput sequencing and their targets using degradome analysis. BMC Genom. 2013, 14, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, L.C.; Lin, S.I.; Shih, A.C.; Chen, J.; Lin, W.; Tseng, C.Y.; Li, W.H.; Chiou, T.J. Uncovering small RNA-mediated responses to phosphate deficiency in Arabidopsis by deep sequencing. Plant Physiol. 2009, 15, 2120–2132. [Google Scholar] [CrossRef] [Green Version]
- Tsikou, D.; Yan, Z.; Holt, D.B.; Abel, N.B.; Reid, D.E.; Madsen, L.H.; Bhasin, H.; Sexauer, M.; Stougaard, J.; Markmann, K. Systemic control of legume susceptibility to rhizobial infection by a mobile microRNA. Science 2018, 362, 233–235. [Google Scholar] [CrossRef]
- Ma, C.; Qi, Y.; Liang, W.; Yang, L.; Lu, Y.; Guo, P.; Ye, X.; Chen, L.S. MicroRNA Regulatory Mechanisms on Citrus sinensis leaves to Magnesium-Deficiency. Front. Plant. Sci. 2016, 7, 207. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Wang, Y.; Wang, L.; Yang, L.; Wang, R.; Li, X. miR172b Controls the Transition to Autotrophic Development Inhibited by ABA in Arabidopsis. PLoS ONE 2013, 8, e64770. [Google Scholar] [CrossRef] [Green Version]
- Migocka, M.; Kosieradzka, A.; Papierniak, A.; Maciaszczyk-Dziubinska, E.; Posyniak, E.; Garbiec, A.; Filleur, S. Two metal-tolerance proteins, MTP1 and MTP4, are involved in Zn homeostasis and Cd sequestration in cucumber cells. J. Exp. Bot. 2015, 66, 1001–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Migocka, M.; Małas, K.; Maciaszczyk-Dziubinska, E.; Posyniak, E.; Migdal, I.; Szczech, P. Cucumber Golgi protein CsMTP5 forms a Zn-transporting heterodimer with high molecular mass protein CsMTP12. Plant. Sci. 2018, 277, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Culotta, V.C. Post-translation control of Nramp metal transport in yeast: Role of metal ions and the bsd2 gene. J. Biol. Chem. 1999, 274, 4863–4868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutherford, J.C.; Bird, A.J. Metal-Responsive Transcription Factors That Regulate Iron, Zinc, and Copper Homeostasis in Eukaryotic Cells. Eukaryot. Cell 2004, 3, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Culotta, V.C.; Yang, M.; Hall, M.D. Manganese transport and trafficking: Lessons learned from Saccharomyces cerevisiae. Eukaryot. Cell 2005, 4, 1159–1165. [Google Scholar] [CrossRef] [Green Version]
- Migocka, M.; Maciaszczyk-Dziubinska, E.; Małas, K.; Posyniak, E.; Garbiec, A. Metal tolerance protein MTP6 affects mitochondrial iron and manganese homeostasis in cucumber. J. Exp. Bot. 2019, 70, 285–300. [Google Scholar] [CrossRef]
- Chen, Z.; Fujii, Y.; Yamaji, N.; Masuda, S.; Takemoto, Y.; Kamiya, T.; Yusuyin, Y.; Iwasaki, K.; Kato, S.; Maeshima, M.; et al. Mn tolerance in rice is mediated by MTP8.1, a member of the cation diffusion facilitator family. J. Exp. Bot. 2013, 64, 4375–4387. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Li, J.; Wang, L.; Ma, G.; Zhang, W. A mutagenic study identifying critical residues for the structure and function of rice manganese transporter OsMTP8.1. Sci. Rep. 2016, 6, 32073. [Google Scholar] [CrossRef] [Green Version]
- Migocka, M.; Papierniak, A.; Kosieradzka, A.; Posyniak, E.; Maciaszczyk-Dziubinska, E.; Biskup, R.; Garbiec, A.; Marchewka, T. Cucumber metal tolerance protein CsMTP9 is a plasma membrane H+-coupled antiporter involved in the Mn2+ and Cd2+ efflux from root cells. Plant. J. 2015, 84, 1045–1058. [Google Scholar] [CrossRef] [Green Version]
- Erbasol, I.; Bozdag, G.O.; Koc, A.; Pedas, P.; Karakaya, H. C Characterization of two genes encoding metal tolerance proteins from Beta vulgarissubspeciesmaritimathat confers manganese tolerance in yeast. Biometals 2013, 26, 795–804. [Google Scholar] [CrossRef] [Green Version]
- Tuskan, G.A.; Difazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of Black Cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [PubMed] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal Wand Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a toolkit for biologists integrating various HTS-data handling tools with a user-friendly interface. BioRxiv 2018. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, J.; Yang, F.; Zhang, G.; Wang, D.; Zhang, L.; Ou, Y.B.; Yao, Y.A. The WRKY transcription factor WRKY8 promotes resistance to pathogen infection and mediates drought and salt stress tolerance in Solanum lycopersicum. Physiol. Plant. 2019, 168, 98–117. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name. | Gene ID | Chromosome Location | Strand | CDS (bp) | Protein Size(aa) | MW (KDa) | PI | GRAVY | Sub-Cellular Localization | TMD Number |
|---|---|---|---|---|---|---|---|---|---|---|
| PtrMTP1.1 | Potri.014G106200 | Chr14:8357551..8361095 | + | 1182 | 393 | 43.47 | 5.81 | 0.074 | Vacuole | 6/In-In |
| PtrMTP1.2 | Potri.002G180100 | Chr02:13987567..13990838 | + | 1182 | 393 | 43.55 | 5.9 | 0.07 | Vacuole | 6/In-In |
| PtrMTP3.1 | Potri.011G150600 | Chr11:16906810..16909370 | − | 1242 | 413 | 45.24 | 6.02 | 0.064 | Vacuole | 6/In-In |
| PtrMTP3.2 | Potri.001G450900 | Chr01:48519109..48521426 | + | 1347 | 448 | 48.94 | 5.85 | −0.174 | Vacuole | 6/In-In |
| PtrMTP4 | Potri.001G245800 | Chr01:25633268..25635016 | + | 1122 | 373 | 41.43 | 5.47 | 0.329 | Vacuole | 6/Out-Out |
| PtrMTP5 | Potri.016G045200 | Chr16:2844025..2847434 | − | 1161 | 386 | 43.24 | 6 | 0.173 | Vacuole | 6/In-In |
| PtrMTP6 | Potri.T034500 | scaffold_36:109770..116219 | + | 1542 | 513 | 55.86 | 6.58 | −0.012 | Vacuole | 0 |
| PtrMTP7 | Potri.010G251300 | Chr10:22330375..22335229 | − | 1380 | 459 | 50.51 | 7.24 | −0.017 | Vacuole | 4/In-In |
| PtrMTP8.1 | Potri.003G215600 | Chr03:21264228..21267534 | + | 1206 | 401 | 45.08 | 5.24 | 0.038 | Vacuole | 5/In-Out |
| PtrMTP8.2 | Potri.001G010200 | Chr01:654690..657190 | − | 1212 | 403 | 45.31 | 6.04 | −0.033 | Vacuole | 5/In-Out |
| PtrMTP8.3 | Potri.001G010300 | Chr01:657997..661070 | − | 1212 | 403 | 45.62 | 5.2 | −0.034 | Vacuole | 4/Out-Out |
| PtrMTP8.4 | Potri.001G010100 | Chr01:651043..652778 | − | 984 | 327 | 36.95 | 5.51 | 0.205 | Vacuole | 5/In-Out |
| PtrMTP8.5 | Potri.001G010000 | Chr01:647003..649031 | − | 984 | 327 | 37.11 | 5.78 | 0.149 | Vacuole | 5/In-Out |
| PtrMTP8.6 | Potri.001G009900 | Chr01:643504..644577 | − | 498 | 165 | 18.38 | 5.48 | 0.048 | Vacuole | 0 |
| PtrMTP9 | Potri.008G083600 | Chr08:5257637..5260169 | − | 1215 | 404 | 46.52 | 6.68 | −0.165 | Cell membrane/Vacuole | 5/In-Out |
| PtrMTP10.1 | Potri.010G172800 | Chr10:17367355..17374280 | + | 1212 | 403 | 46.52 | 6.77 | −0.235 | Vacuole | 6/In-In |
| PtrMTP10.2 | Potri.010G172900 | Chr10:17376155..17384476 | + | 1305 | 434 | 50.17 | 6.8 | −0.198 | Cell membrane/Vacuole | 4/Out-Out |
| PtrMTP10.3 | Potri.010G172700 | Chr10:17359356..17361845 | + | 1317 | 438 | 49.89 | 6.24 | −0.064 | Cell membrane/Vacuole | 5/In-Out |
| PtrMTP10.4 | Potri.010G172600 | Chr10:17355982..17358522 | + | 1224 | 407 | 46.73 | 6.88 | −0.076 | Cell membrane/Vacuole | 5/In-Out |
| PtrMTP11.1 | Potri.010G211300 | Chr10:19986602..19990567 | + | 1185 | 394 | 44.87 | 5.05 | −0.053 | Vacuole | 4/Out-Out |
| PtrMTP11.2 | Potri.008G049600 | Chr08:2924867..2928439 | − | 1185 | 394 | 44.74 | 4.88 | −0.055 | Vacuole | 3/Out-In |
| PtrMTP12 | Potri.005G110300 | Chr05:8489679..8492954 | − | 2610 | 869 | 97.5 | 6.95 | −0.026 | Vacuole | 12/In-In |
| Duplicatedpair | Duplicate Type | Ka | Ks | Ka/Ks | Positive Selection |
|---|---|---|---|---|---|
| PtrMTP1.1/PtrMTP1.2 | Segmental | 0.0752 | 0.2006 | 0.374875 | No |
| PtrMTP3.1/PtrMTP3.2 | Segmental | 0.0591 | 0.3208 | 0.184227 | No |
| PtrMTP8.1/PtrMTP8.6 | Segmental | 0.0683 | 0.2624 | 0.26029 | No |
| PtrMTP9/PtrMTP10.4 | Segmental | 0.0627 | 0.2097 | 0.298999 | No |
| PtrMTP11.1/PtrMTP11.2 | Segmental | 0.0388 | 0.2849 | 0.136188 | No |
| PtrMTP8.2/PtrMTP8.3 | Tandem | 0.1652 | 1.1124 | 0.148508 | No |
| PtrMTP8.2/PtrMTP8.4 | Tandem | 0.0027 | 0.0044 | 0.613636 | No |
| PtrMTP8.4/PtrMTP8.5 | Tandem | 0.016 | 0.0313 | 0.511182 | No |
| PtrMTP10.1/PtrMTP10.2 | Tandem | 0.0152 | 0.0257 | 0.59144 | No |
| PtrMTP10.3/PtrMTP10.4 | Tandem | 0.1409 | 0.7069 | 0.199321 | No |
| Gene Name | Gene Transcription | Abiotic Stress | Biotic Stress | Tissue Expression | Secondary Metabolism | Phytohormonal Responsive | Light Response | Circadian Control | Site-Binding |
|---|---|---|---|---|---|---|---|---|---|
| PtrMTP1.1 | 43 | 0 | 0 | 0 | 2 | 0 | 2 | 0 | 0 |
| PtrMTP1.2 | 63 | 4 | 0 | 1 | 0 | 0 | 5 | 0 | 1 |
| PtrMTP3.1 | 33 | 3 | 0 | 0 | 0 | 1 | 3 | 0 | 1 |
| PtrMTP3.2 | 73 | 4 | 1 | 1 | 1 | 1 | 7 | 0 | 3 |
| PtrMTP4 | 41 | 3 | 0 | 1 | 0 | 5 | 12 | 0 | 1 |
| PtrMTP5 | 30 | 1 | 0 | 0 | 0 | 4 | 2 | 0 | 1 |
| PtrMTP6 | 58 | 1 | 0 | 1 | 1 | 3 | 5 | 0 | 1 |
| PtrMTP7 | 34 | 4 | 0 | 0 | 0 | 2 | 4 | 0 | 0 |
| PtrMTP8.1 | 57 | 1 | 0 | 0 | 0 | 6 | 12 | 0 | 4 |
| PtrMTP8.2 | 20 | 3 | 0 | 0 | 0 | 2 | 5 | 0 | 1 |
| PtrMTP8.3 | 32 | 3 | 0 | 0 | 0 | 7 | 13 | 0 | 3 |
| PtrMTP8.4 | 20 | 2 | 0 | 0 | 0 | 3 | 7 | 1 | 2 |
| PtrMTP8.5 | 39 | 2 | 0 | 0 | 0 | 2 | 9 | 2 | 0 |
| PtrMTP8.6 | 20 | 3 | 0 | 0 | 0 | 3 | 6 | 2 | 2 |
| PtrMTP9 | 20 | 1 | 0 | 1 | 0 | 1 | 5 | 0 | 0 |
| PtrMTP10.1 | 33 | 2 | 0 | 1 | 0 | 7 | 15 | 0 | 4 |
| PtrMTP10.2 | 33 | 2 | 0 | 0 | 0 | 7 | 14 | 0 | 2 |
| PtrMTP10.3 | 27 | 1 | 0 | 1 | 0 | 5 | 4 | 0 | 1 |
| PtrMTP10.4 | 45 | 1 | 0 | 1 | 0 | 1 | 6 | 0 | 0 |
| PtrMTP11.1 | 25 | 5 | 0 | 0 | 0 | 11 | 12 | 0 | 5 |
| PtrMTP11.2 | 60 | 1 | 0 | 0 | 0 | 3 | 11 | 0 | 3 |
| PtrMTP12 | 111 | 5 | 0 | 0 | 1 | 6 | 9 | 0 | 0 |
| miRNA Acc. | Target Acc. | Expectation | UPE | miRNA Length | Target Start-End | miRNA Aligned Fragment | Target Aligned Fragment | Inhibition |
|---|---|---|---|---|---|---|---|---|
| ptc-miR473b | PtrMTP1.1 | 2.5 | 15.471 | 20 | 932–951 | GCUCUCCCUCAGGGCUUCCA | UUGAAGUCCUGAUGGAGAGC | Cleavage |
| ptc-miR2111a | PtrMTP11.2 | 3 | 16.046 | 21 | 550–571 | UAAUCUGC-AUCCUGAGGUUUG | GCAACUUUAGGAUUGCAGAUUA | Cleavage |
| ptc-miR2111a | PtrMTP11.1 | 3 | 13.919 | 21 | 550–571 | UAAUCUGC-AUCCUGAGGUUUG | GCAACUUUAGGAUUGCAGAUUA | Cleavage |
| ptc-miR2111b | PtrMTP11.2 | 3 | 16.046 | 21 | 550–571 | UAAUCUGC-AUCCUGAGGUUUG | GCAACUUUAGGAUUGCAGAUUA | Cleavage |
| ptc-miR2111b | PtrMTP11.1 | 3 | 13.919 | 21 | 550–571 | UAAUCUGC-AUCCUGAGGUUUG | GCAACUUUAGGAUUGCAGAUUA | Cleavage |
| ptc-miR6426a | PtrMTP8.1 | 3 | 19.01 | 21 | 162–182 | GUGGAGACAUGGAAGUGAAGA | UUUUCACUUUAAUGUCUCUAA | Translation |
| ptc-miR6426b | PtrMTP8.1 | 3 | 19.01 | 21 | 162–182 | GUGGAGACAUGGAAGUGAAGA | UUUUCACUUUAAUGUCUCUAA | Translation |
| ptc-miR6427-3p | PtrMTP12 | 3 | 15.602 | 21 | 900–920 | GUGGGAAUGAACAUUAUGAGA | AAUUAUACUGUUUAUUCCUGC | Cleavage |
| ptc-miR172b-5p | PtrMTP12 | 3.5 | 16.256 | 21 | 1705–1725 | GGAGCAUCAUCAAGAUUCACA | GGUGGCUCUGGAUCAUGCUCC | Cleavage |
| ptc-miR172g-5p | PtrMTP12 | 3.5 | 16.256 | 21 | 1705–1725 | GGAGCAUCAUCAAGAUUCACA | GGUGGCUCUGGAUCAUGCUCC | Cleavage |
| ptc-miR473b | PtrMTP3.1 | 3.5 | 18.76 | 20 | 989–1008 | GCUCUCCCUCAGGGCUUCCA | UGGAGGUUCUCAUGGAGAGC | Translation |
| ptc-miR480 | PtrMTP7 | 3.5 | 13.362 | 24 | 1122–1145 | ACUACUACAUCAUUGACGUUGAAC | AAUAGAUUUCAAUGGAGUAGUGGU | Translation |
| ptc-miR6464 | PtrMTP7 | 3.5 | 14.684 | 21 | 344–364 | UGAUUGCUUGUUGGAUAUUAU | AACAUAGUCAACGAGCAGUCA | Cleavage |
| ptc-miR6466-3p | PtrMTP10.3 | 3.5 | 19.17 | 21 | 1009–1029 | UAUCAAUCAUCAAAUGUUCGU | GAGAACGUUUGGUCGUUGAUC | Cleavage |
| Gene Name | In Roots | In Stems | In Leaves | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cd | Co | Cu | Fe | Mn | Ni | Zn | Cd | Co | Cu | Fe | Mn | Ni | Zn | Cd | Co | Cu | Fe | Mn | Ni | Zn | |
| PtrMTP1.1 | No | No | No | No | No | - | No | - | No | No | No | No | - | No | No | No | + | + | No | No | + |
| PtrMTP1.2 | No | No | No | No | - | No | No | No | No | No | No | No | No | No | No | No | + | No | No | No | No |
| PtrMTP3.1 | No | No | No | No | No | No | No | No | No | No | No | + | No | No | No | No | No | No | No | No | No |
| PtrMTP3.2 | No | No | No | No | No | No | No | No | No | - | No | No | No | No | No | No | No | No | No | No | No |
| PtrMTP4 | No | + | No | No | No | No | No | No | No | No | No | No | No | No | No | + | + | No | No | No | No |
| PtrMTP5 | + | + | No | No | No | No | No | - | No | No | No | + | No | No | + | No | ++ | + | + | No | + |
| PtrMTP6 | No | No | No | No | - | No | No | No | No | No | No | No | No | No | No | No | No | No | No | No | No |
| PtrMTP7 | + | + | No | No | No | No | No | No | No | No | No | No | - | No | + | + | + | + | No | No | + |
| PtrMTP8.1 | + | No | ++ | No | No | No | + | - | - | No | - | ++ | - | No | No | No | + | + | No | No | + |
| PtrMTP8.2 | No | No | + | No | No | No | No | No | No | No | No | + | No | No | No | No | ++ | + | No | No | + |
| PtrMTP8.3 | No | + | No | No | - | No | No | No | + | ++ | No | +++ | No | + | No | No | ++ | + | No | No | + |
| PtrMTP8.4 | No | No | + | No | No | No | + | No | No | No | No | ++ | No | No | No | No | ++ | + | No | No | ++ |
| PtrMTP8.5 | No | No | + | No | No | No | No | No | No | No | No | ++ | No | No | No | No | ++ | + | No | No | + |
| PtrMTP8.6 | + | + | No | No | + | No | No | + | ++ | No | No | No | No | No | No | No | No | No | No | No | No |
| PtrMTP9 | No | No | -- | No | --- | No | No | - | No | - | + | No | No | No | - | No | No | +++ | No | No | No |
| PtrMTP10.1 | + | + | No | No | No | - | + | - | No | No | No | No | - | No | + | + | +++ | ++ | + | No | ++ |
| PtrMTP10.2 | No | + | + | No | No | No | + | No | No | No | No | + | No | No | No | No | ++ | + | + | No | + |
| PtrMTP10.3 | No | No | +++ | No | No | -- | +++ | No | --- | + | - | + | -- | + | + | No | +++ | ++ | ++ | No | +++ |
| PtrMTP10.4 | No | No | No | No | - | No | No | No | No | + | - | +++ | No | + | + | + | +++ | + | + | + | +++ |
| PtrMTP11.1 | ++ | + | No | No | No | No | No | No | No | No | No | + | - | No | + | No | +++ | + | No | No | + |
| PtrMTP11.2 | No | No | + | No | No | No | No | No | No | No | No | ++ | No | + | No | + | + | No | No | No | + |
| PtrMTP12 | No | + | No | No | ---- | No | No | -- | No | - | No | No | No | No | No | No | No | No | No | No | No |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Yang, F.; Liu, J.; Xie, W.; Zhang, L.; Chen, Z.; Peng, Z.; Ou, Y.; Yao, Y. Genome-Wide Identification of Metal Tolerance Protein Genes in Populus trichocarpa and Their Roles in Response to Various Heavy Metal Stresses. Int. J. Mol. Sci. 2020, 21, 1680. https://doi.org/10.3390/ijms21051680
Gao Y, Yang F, Liu J, Xie W, Zhang L, Chen Z, Peng Z, Ou Y, Yao Y. Genome-Wide Identification of Metal Tolerance Protein Genes in Populus trichocarpa and Their Roles in Response to Various Heavy Metal Stresses. International Journal of Molecular Sciences. 2020; 21(5):1680. https://doi.org/10.3390/ijms21051680
Chicago/Turabian StyleGao, Yongfeng, Fengming Yang, Jikai Liu, Wang Xie, Lin Zhang, Zihao Chen, Zhuoxi Peng, Yongbin Ou, and Yinan Yao. 2020. "Genome-Wide Identification of Metal Tolerance Protein Genes in Populus trichocarpa and Their Roles in Response to Various Heavy Metal Stresses" International Journal of Molecular Sciences 21, no. 5: 1680. https://doi.org/10.3390/ijms21051680
APA StyleGao, Y., Yang, F., Liu, J., Xie, W., Zhang, L., Chen, Z., Peng, Z., Ou, Y., & Yao, Y. (2020). Genome-Wide Identification of Metal Tolerance Protein Genes in Populus trichocarpa and Their Roles in Response to Various Heavy Metal Stresses. International Journal of Molecular Sciences, 21(5), 1680. https://doi.org/10.3390/ijms21051680