Proanthocyanidins Ameliorated Deficits of Lipid Metabolism in Type 2 Diabetes Mellitus Via Inhibiting Adipogenesis and Improving Mitochondrial Function

 and
and
Abstract
:1. Introduction
2. Results
2.1. Spectrophotometric Analysis of the Proanthocyanidins
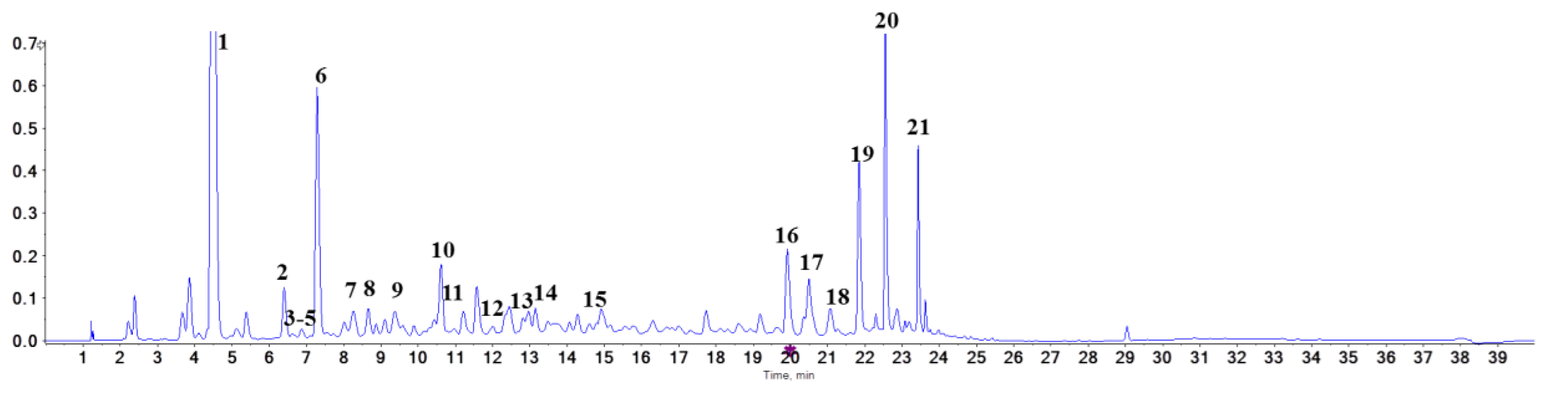
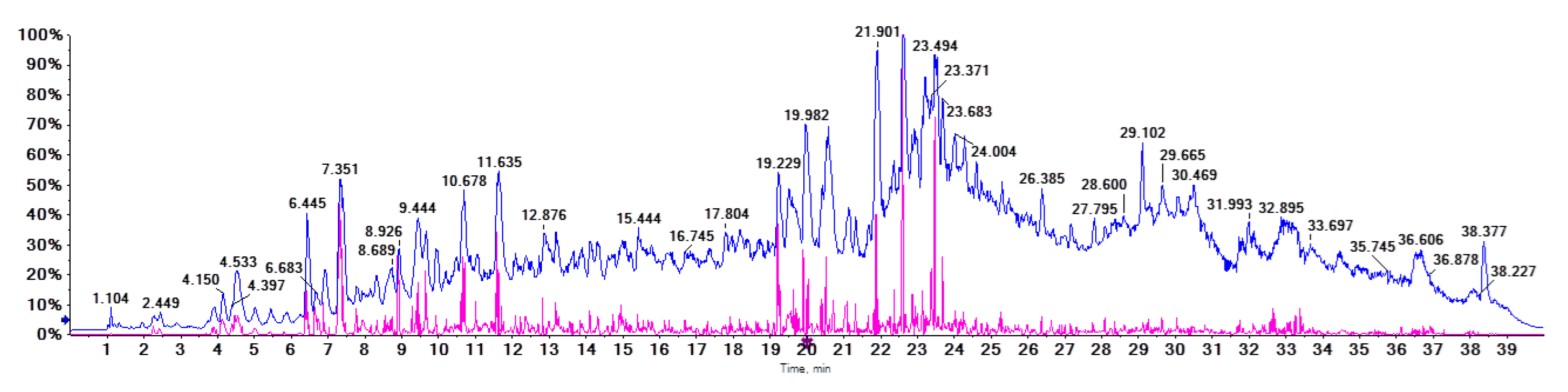
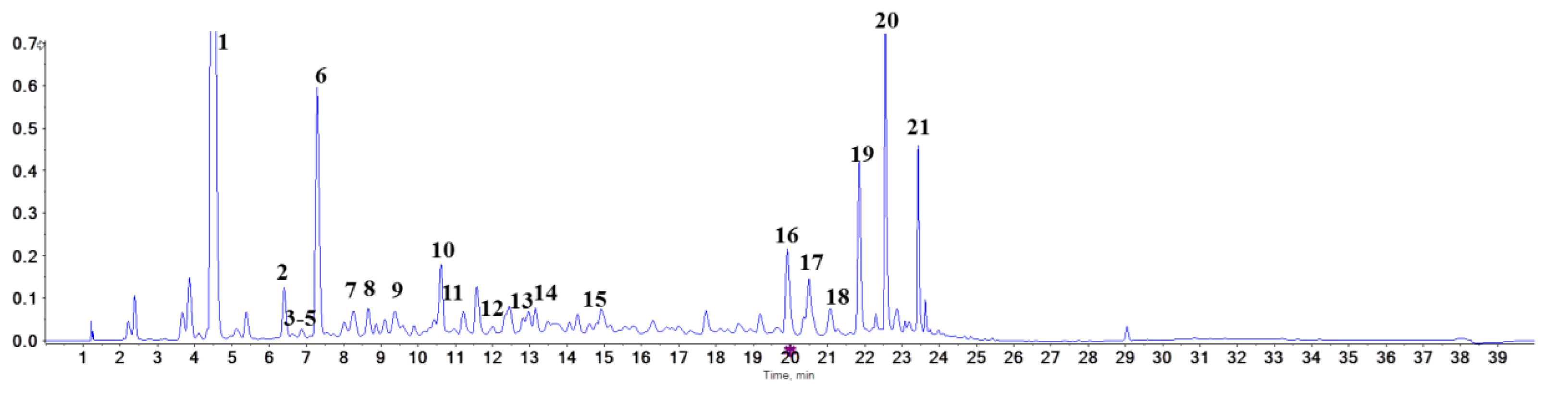
2.2. Identification of Proanthocyanidins with UPLC-Triple-TOF/MS Analysis
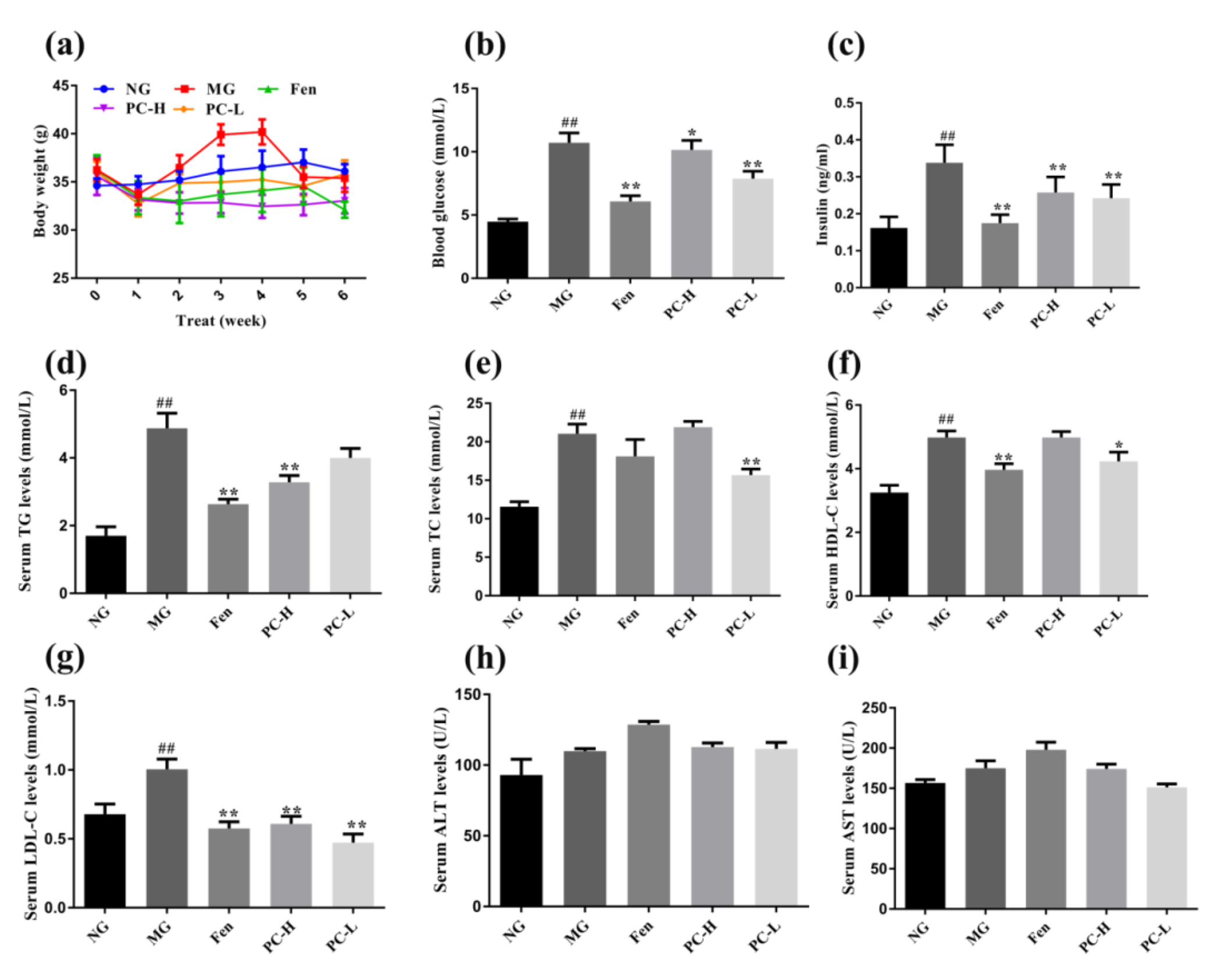
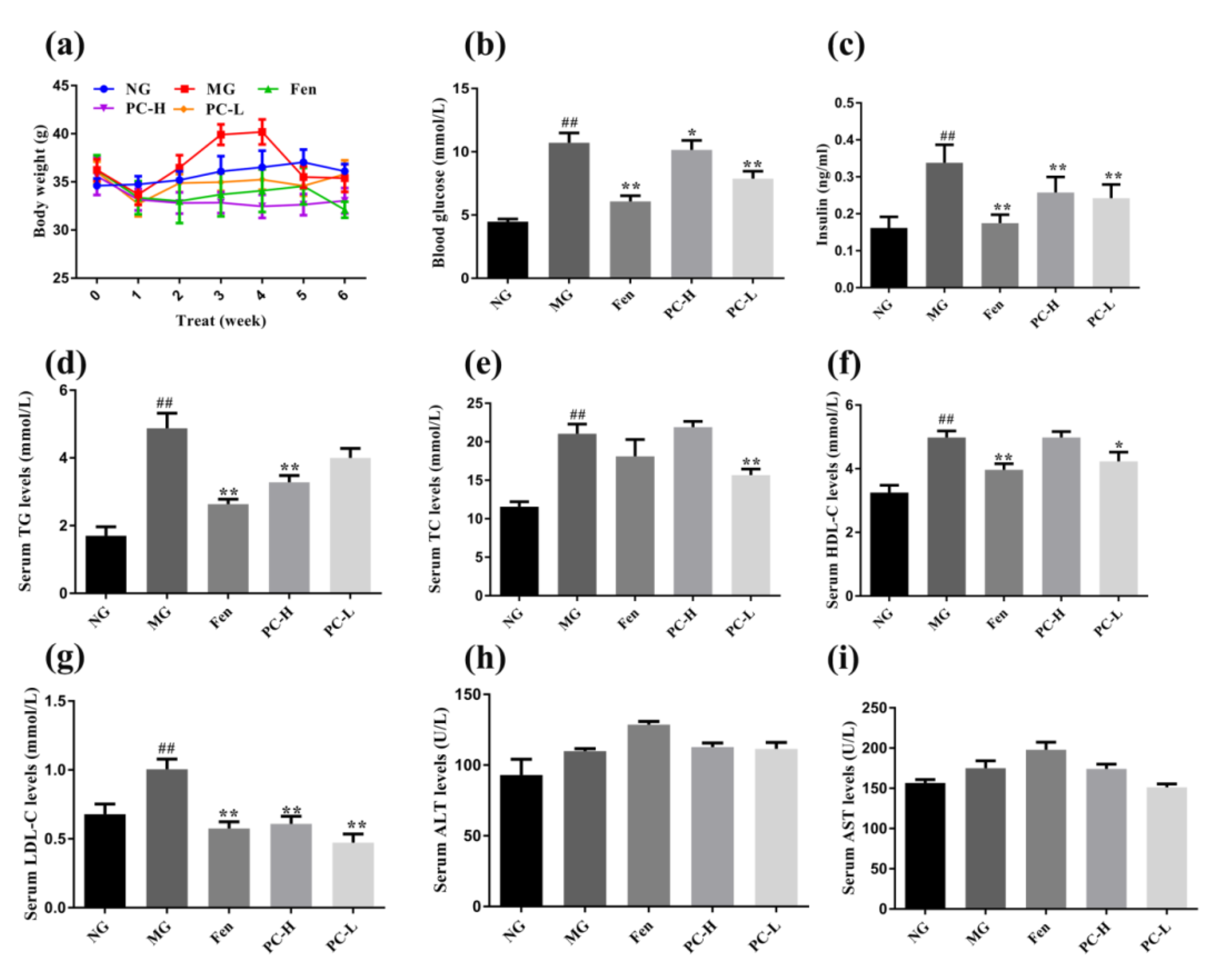
2.3. Effects of Proanthocyanidins on Body Weight and Metabolic Markers in Serum
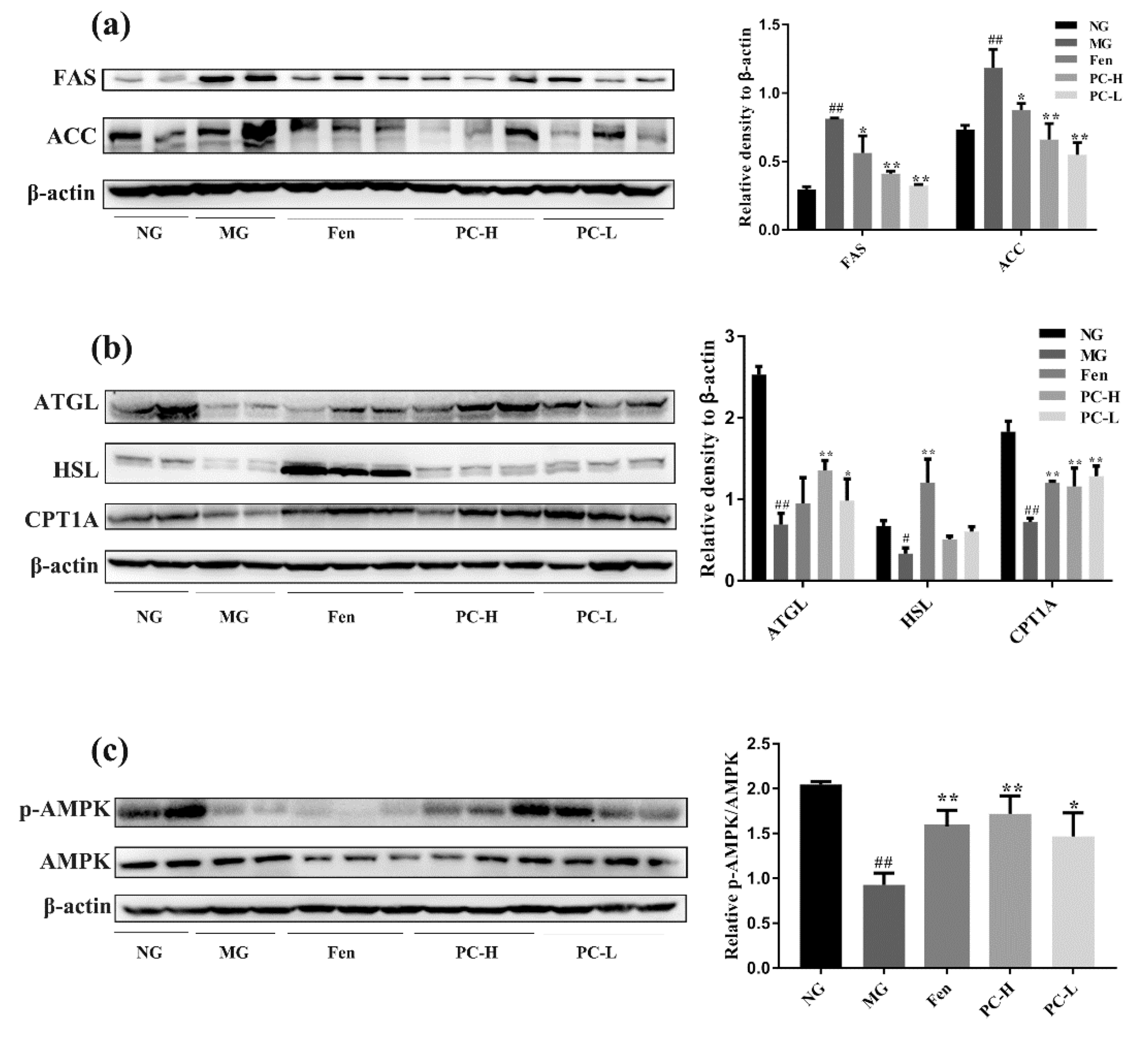
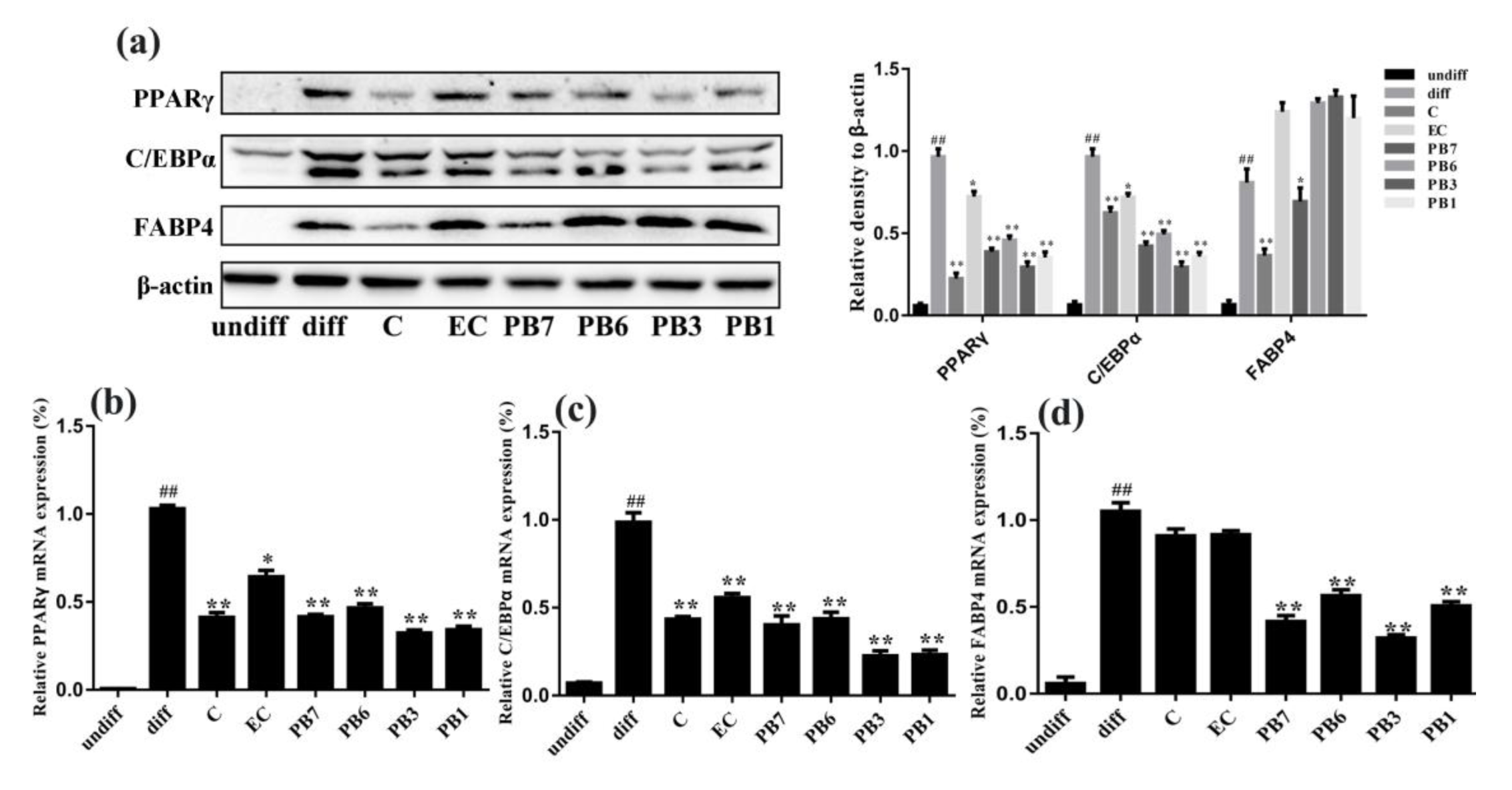
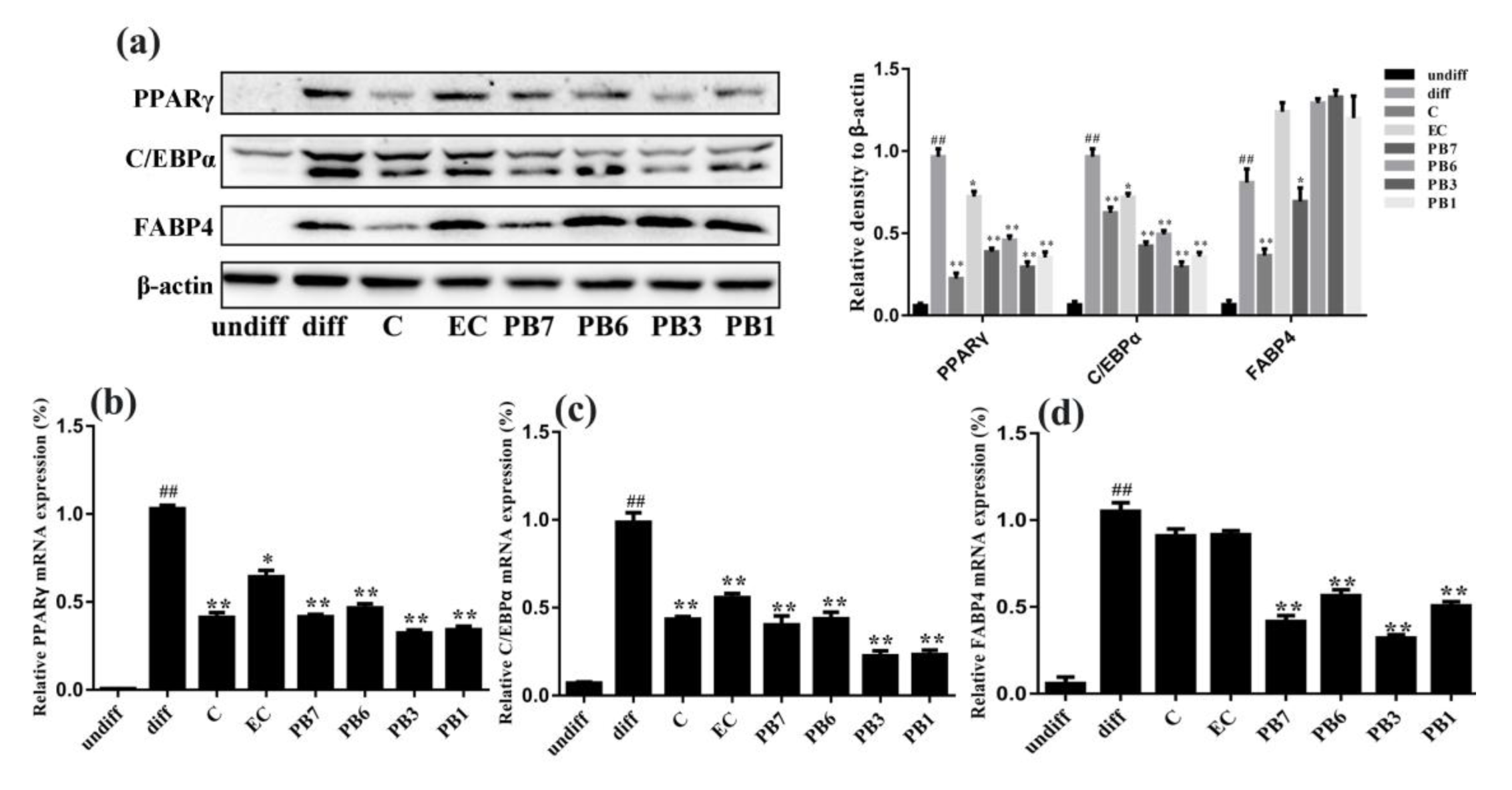
2.4. Effects of Proanthocyanidins on Expression of Lipid Metabolism-Associated Proteins
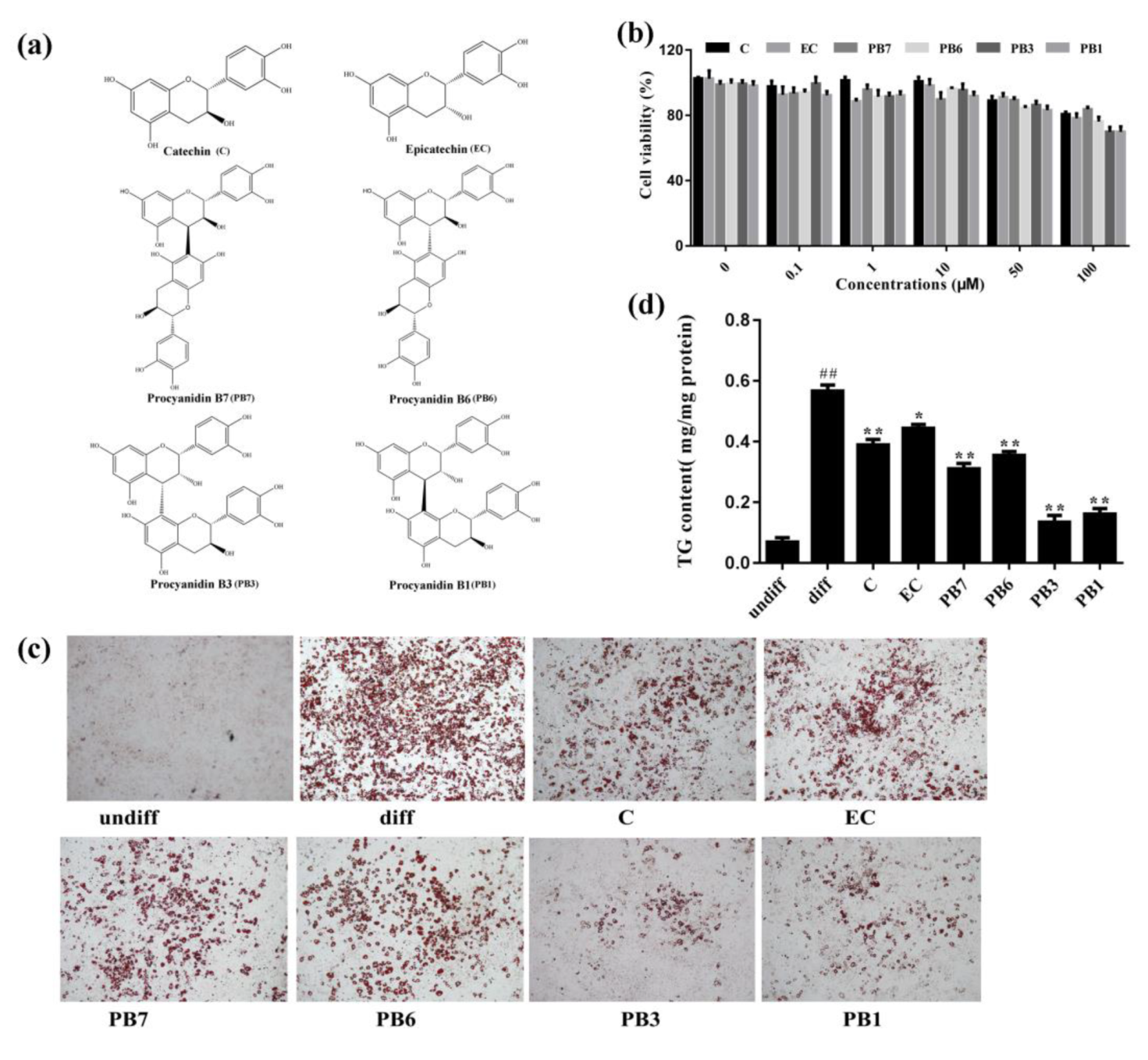
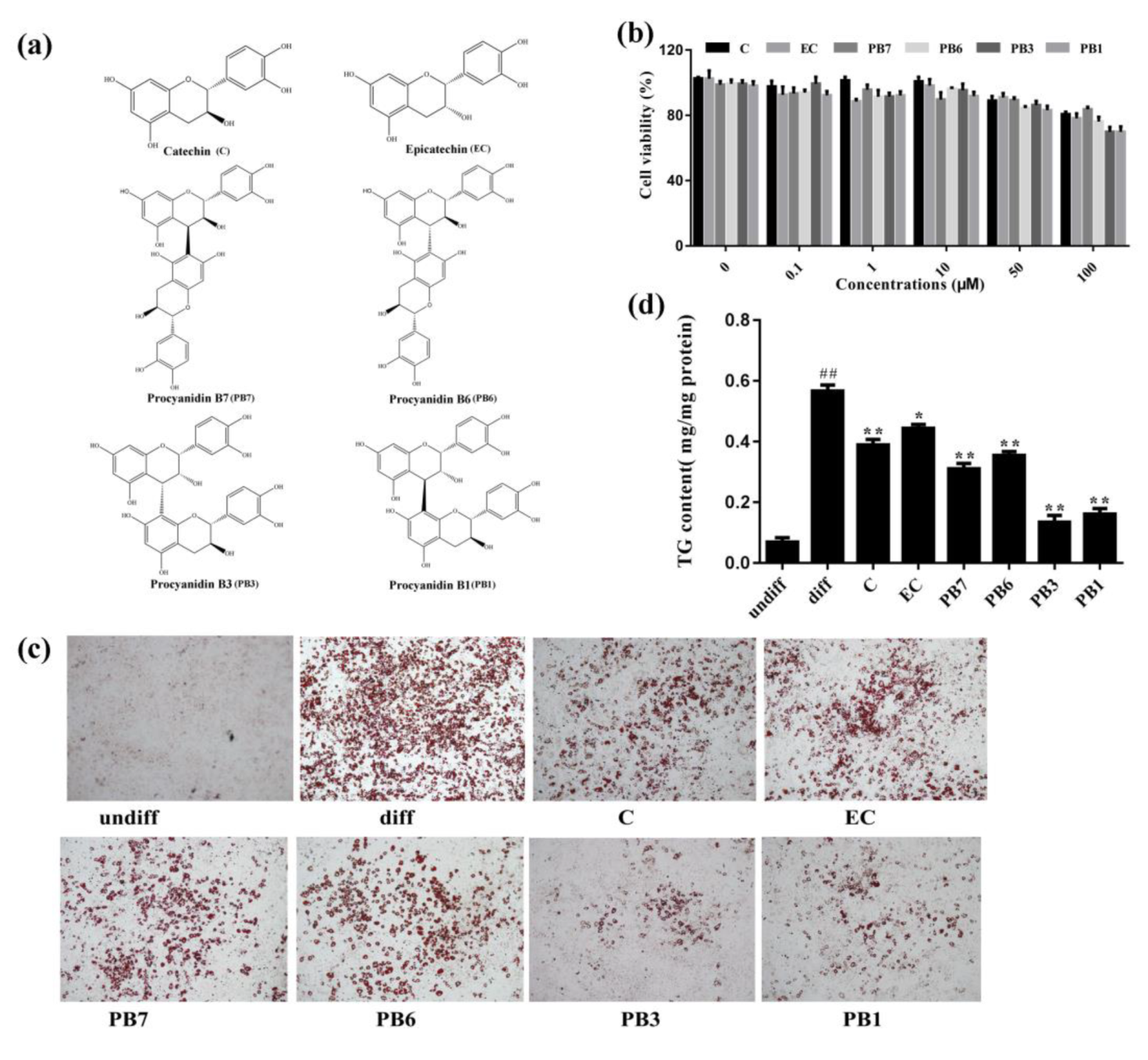
2.5. Effects of Flavan-3-Ols on Intracellular Lipid Accumulation in 3T3-L1 Adipocytes
2.6. Effects of Flavan-3-Ols on Adipogenesis
2.7. Effects of Flavan-3-Ols on Glucose Uptake and Phosphorylation of AKT in Insulin-Resistant 3T3-L1 Adipocytes
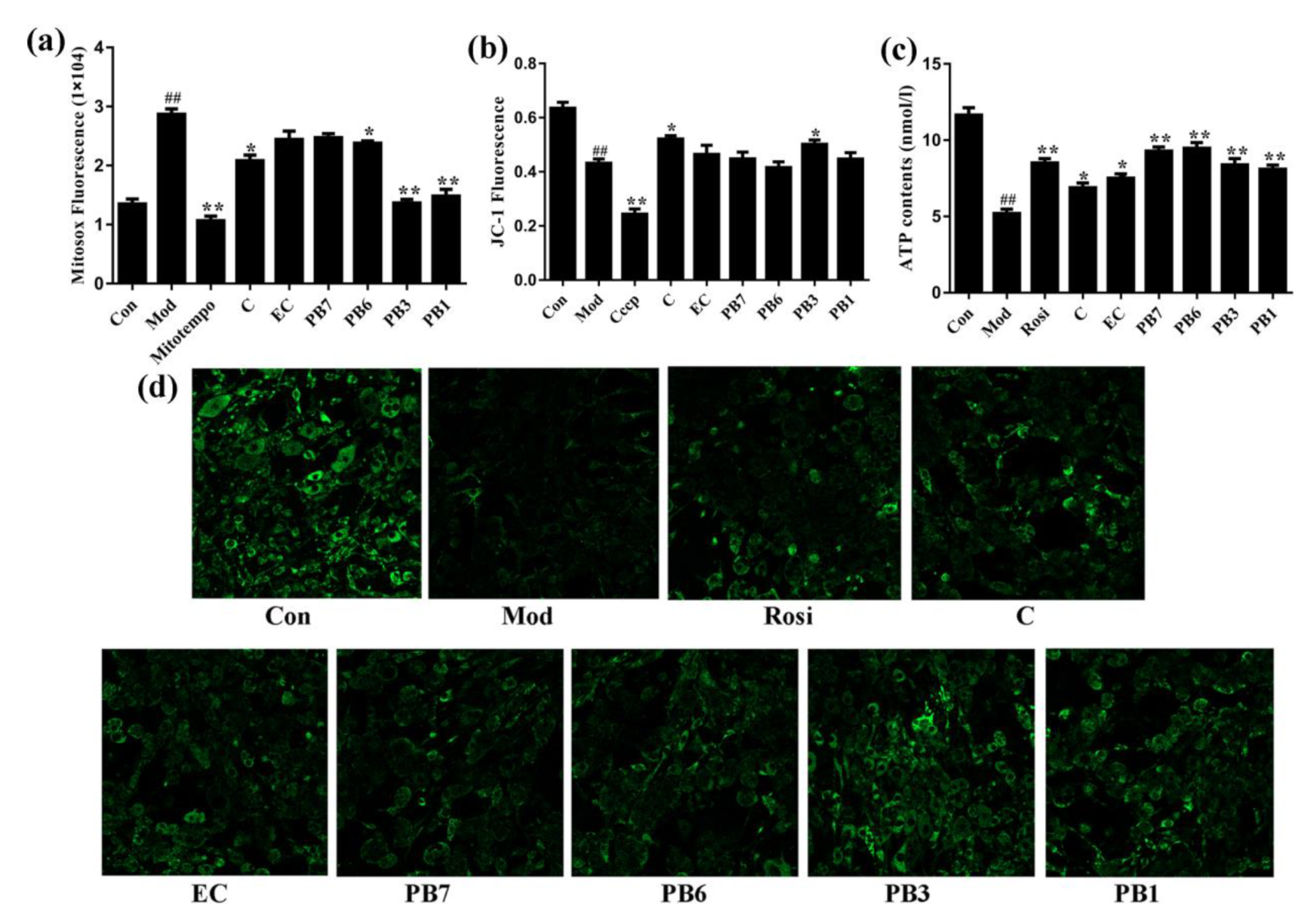
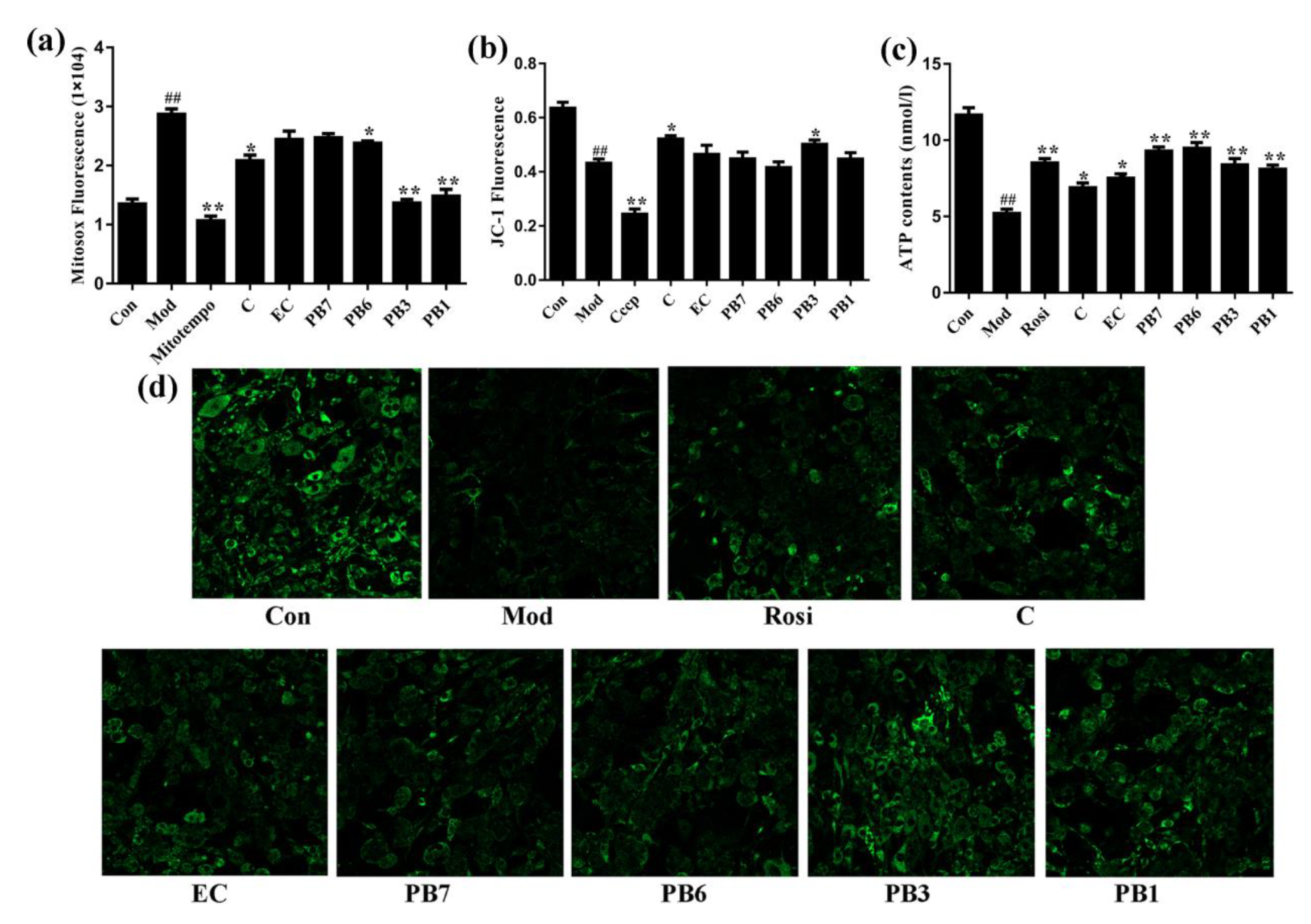
2.8. Effects of Flavan-3-Ols on Mitochondrial Function in Insulin-Resistant 3T3-L1 Adipocytes
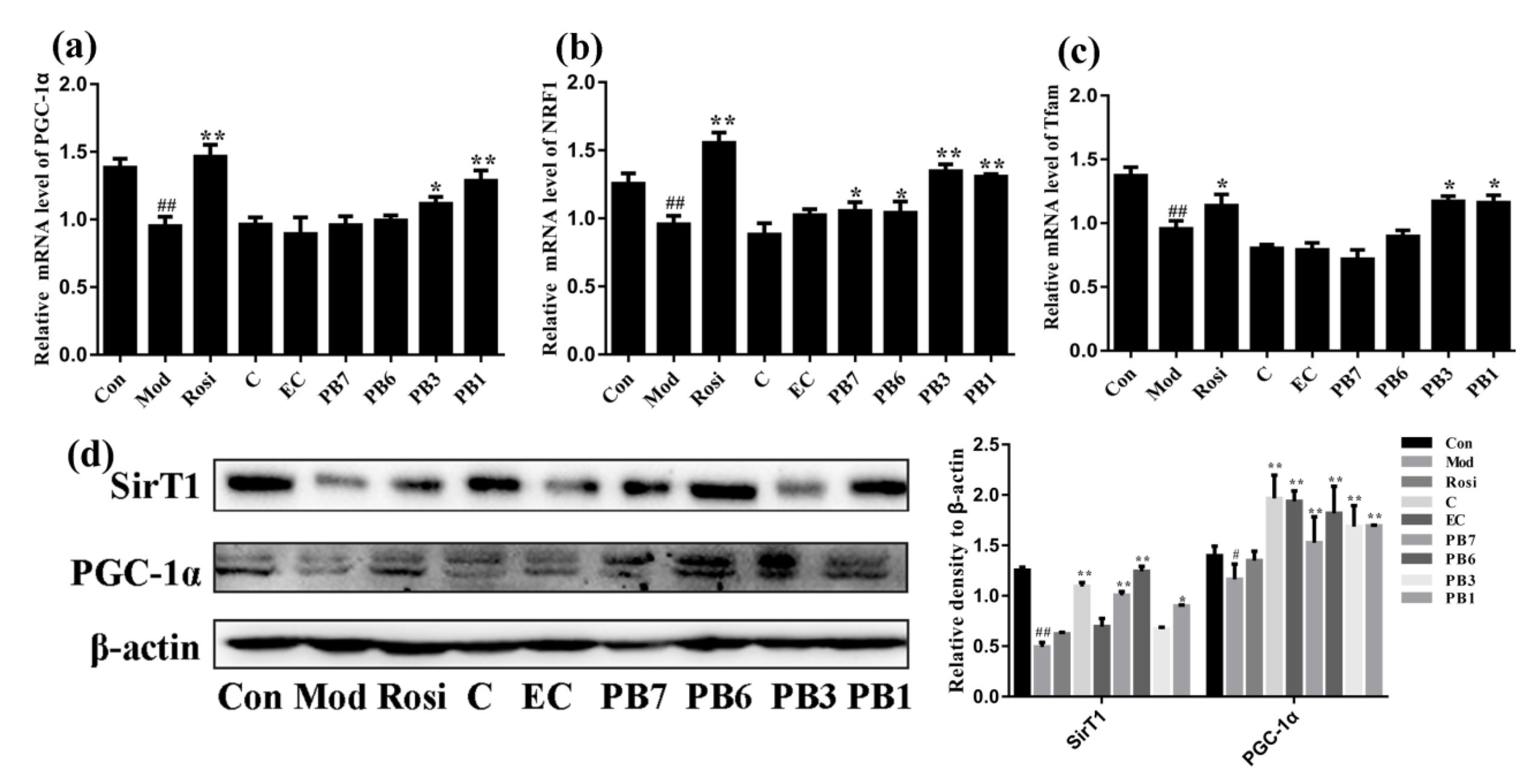
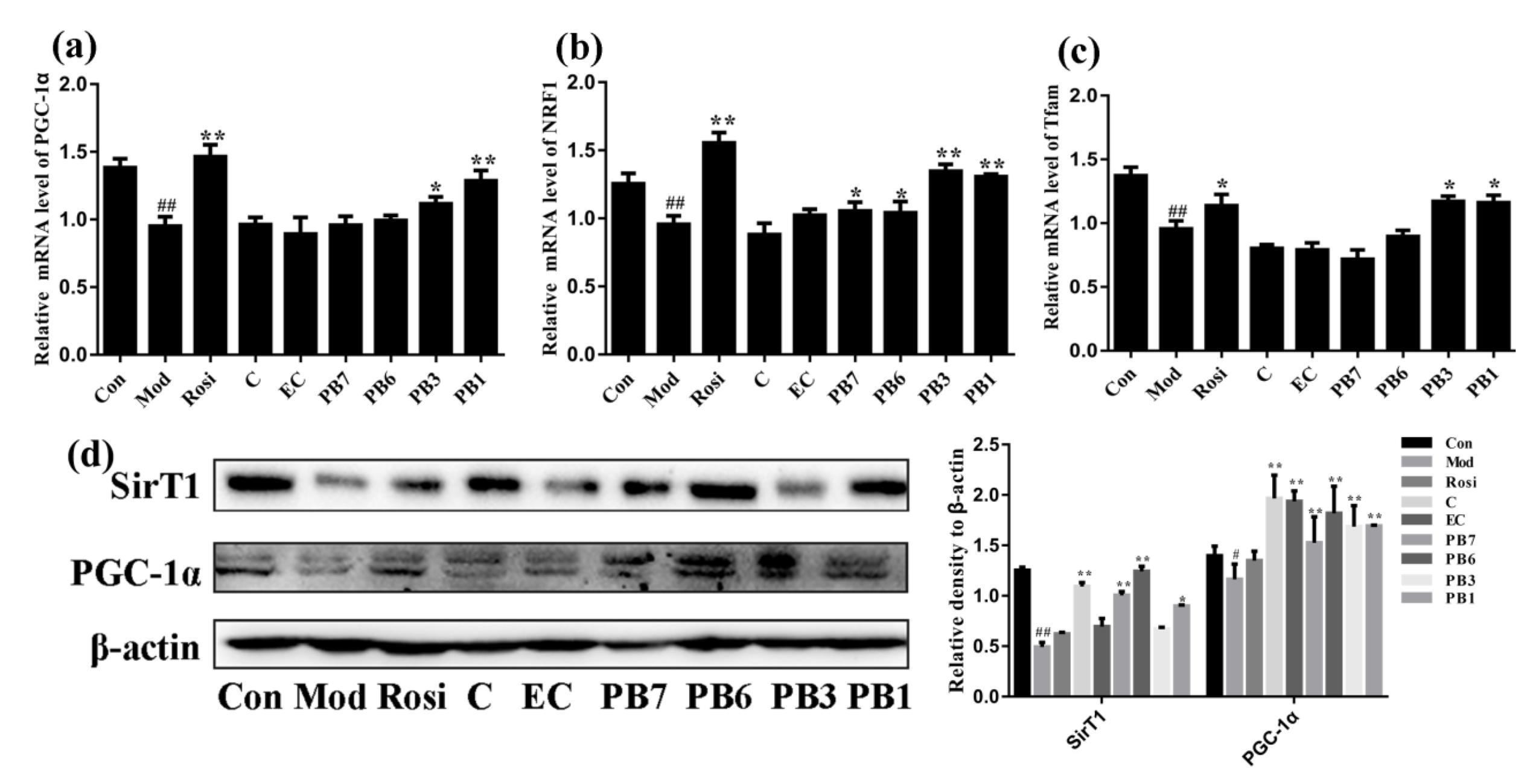
2.9. Effects of Flavan-3-Ols on Expression of Mitochondrial Biogenesis-Related Proteins in Insulin-Resistant 3T3-L1 Adipocytes
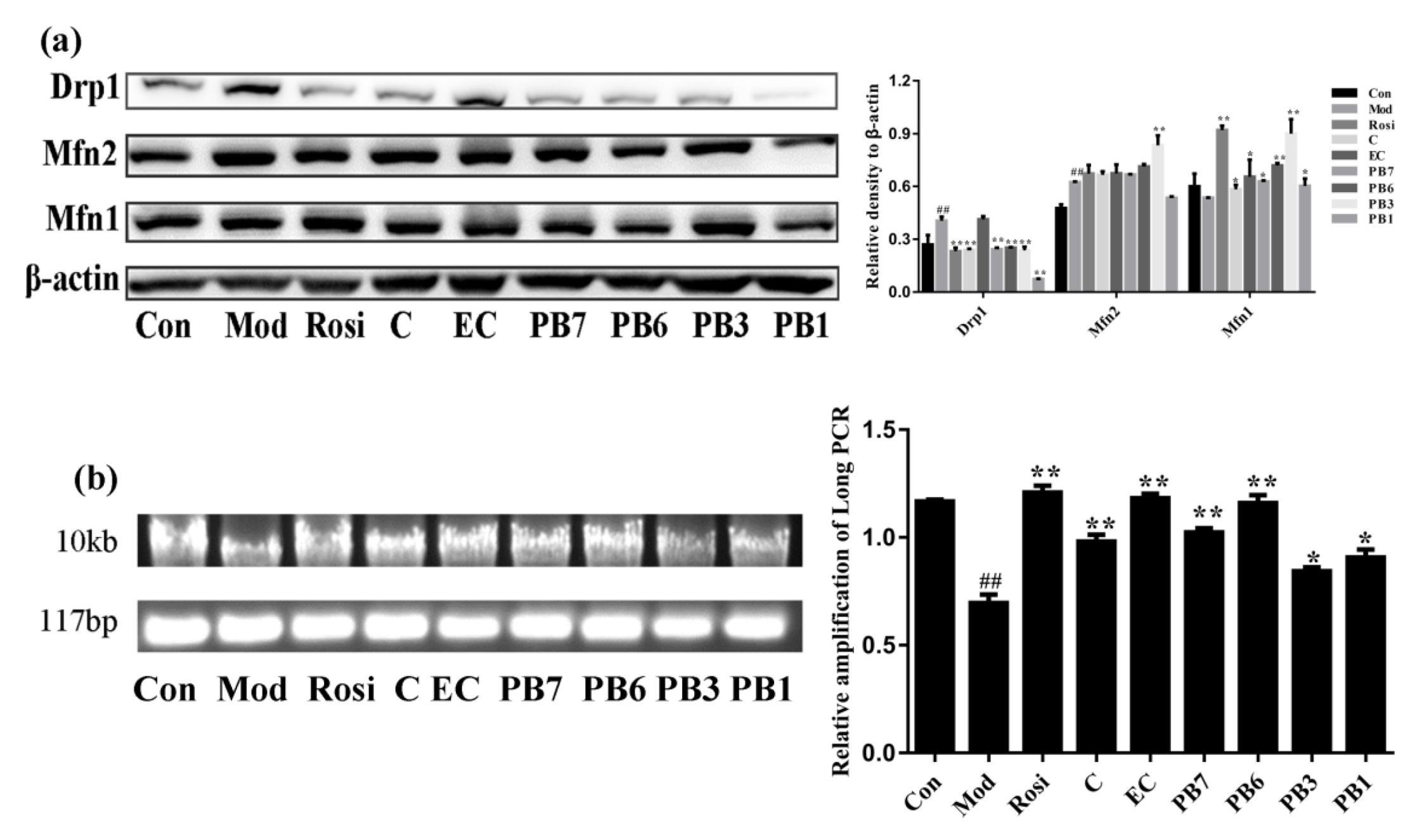
2.10. Effects of Flavan-3-Ols on Mitochondrial Dynamics and Mtdna Damage in Insulin-Resistant 3T3-L1 Adipocytes
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation of Enriched Proanthocyanidins
4.3. Spectrophotometric Analysis
4.4. HPLC Analysis
4.5. UPLC-Triple-TOF/MS
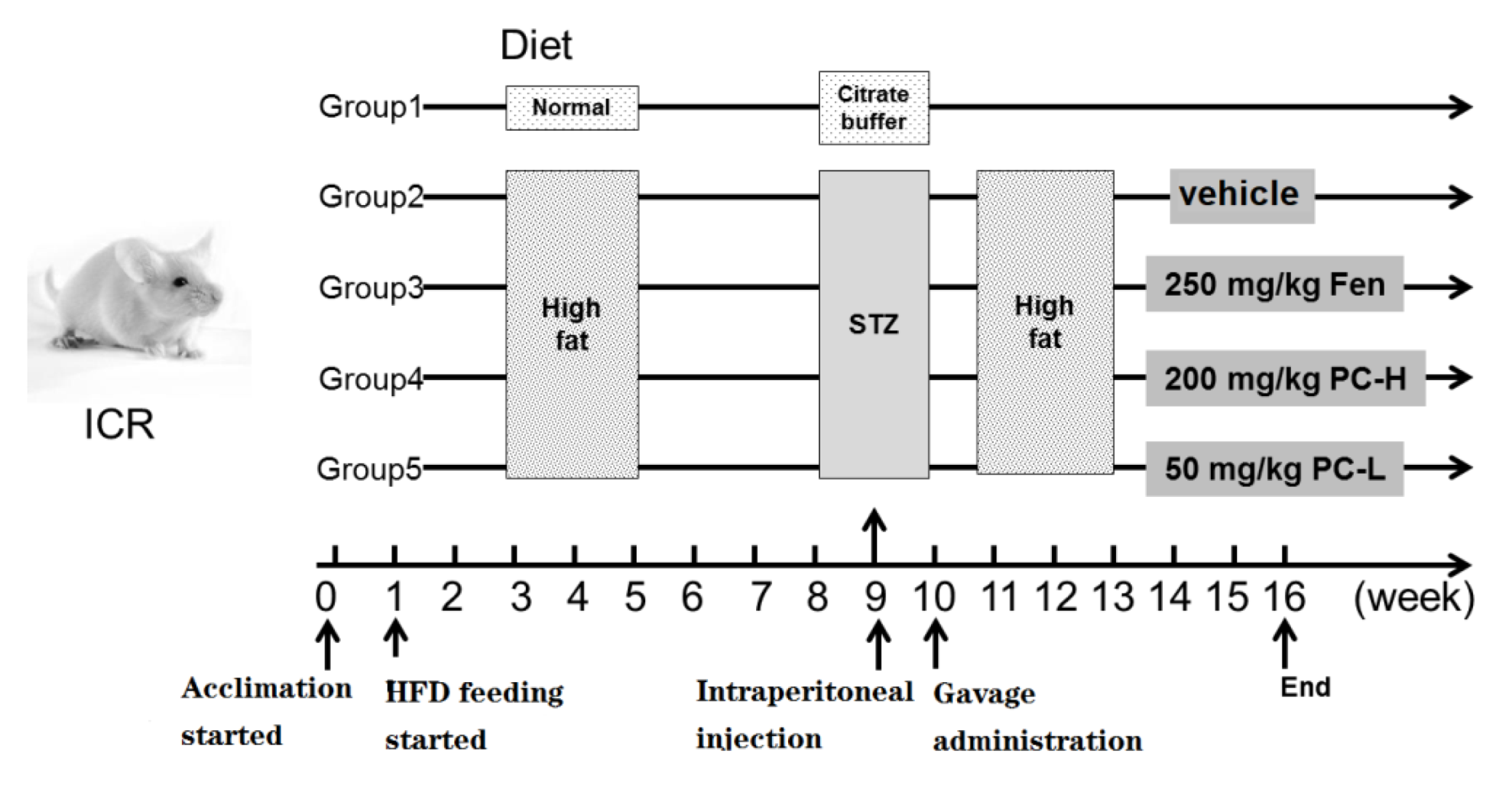
4.6. Animals and Treatments
4.7. Biochemical Analysis of Blood Samples
4.8. Western Blot Analysis
4.9. Cell Culture and Treatment
4.10. MTT Assay
4.11. Measurement of Cellular Lipid Accumulation and Lipolysis
4.12. Glucose Uptake Assay
4.13. Measurement of Mitochondrial Functions
4.14. RNA Isolation and Reverse Transcription PCR
4.15. DNA Isolation and Long PCR
4.16. Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| T2DM | Type 2 diabetes mellitus |
| TG | Triglycerides |
| TC | Total cholesterol |
| HDL-C | High-density lipoprotein-cholesterol |
| LDL-C | Low-density lipoprotein-cholesterol |
| ALT | Alanine aminotransferase |
| AST | Aspartate aminotransferase |
| FAS | Fatty acid synthase |
| ACC | Acetyl-CoA carboxylase |
| ATGL | Adipocyte-triglyceride lipase |
| CPT1A | Carnitine palmitoyltransferase1 |
| AMPK | Adenosine 5’–monophosphate (AMP)-activated protein kinase |
| PPARγ | Peroxisome proliferator activated receptor gamma |
| C/EBPα | CCAAT/enhancer binding protein alpha |
| NRF1 | Nuclear respiratory factor 1 |
| PGC-1α | Peroxisome proliferator-activa-ted receptor gamma coactivator 1-alpha |
| Tfam | Mitochondrial transcription factor |
References
- Association, A.D. Diagnosis and classification of diabetes mellitus. Diabetes Care 2012, 35 (Suppl. 1), S64–S71. [Google Scholar] [CrossRef] [Green Version]
- Guariguata, L.; Whiting, D.R.; Hambleton, I.; Beagley, J.; Linnenkamp, U.; Shaw, J.E. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabetes Res. Clin. Pract. 2014, 103, 137–149. [Google Scholar] [CrossRef]
- Hackett, R.A.; Steptoe, A. Type 2 diabetes mellitus and psychological stress—a modifiable risk factor. Nat. Rev. Endocrinol. 2017, 13, 547–560. [Google Scholar] [CrossRef]
- Niu, J.; Pi, Z.F.; Yue, H.; Yang, H.; Wang, Y.; Yu, Q.; Liu, S.Y. Effect of 20(S)-ginsenoside Rg3 on streptozotocin-induced experimental type 2 diabetic rats: A urinary metabonomics study by rapid-resolution liquid chromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 2012, 26, 2683–2689. [Google Scholar] [CrossRef]
- Lewis, G.F.; Carpentier, A.; Adeli, K.; Giacca, A. Disordered fat storage and mobilization in the pathogenesis of insulin resistance and type 2 diabetes. Endocr. Rev. 2002, 23, 201–229. [Google Scholar] [CrossRef] [PubMed]
- Savage, D.B.; Petersen, K.F.; Shulman, G.I. Disordered lipid metabolism and the pathogenesis of insulin resistance. Physiol. Rev. 2007, 87, 507–520. [Google Scholar] [CrossRef] [PubMed]
- Cham, S.; Koslik, H.J.; Golomb, B.A. Mood, personality, and behavior changes during treatment with statins: A case series. Drug Saf. Case Rep. 2016, 3, 1–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedro-Botet, J.; Millan Nunez-Cortes, J.; Chillaron, J.J.; Flores-Le Roux, J.A.; Rius, J. Severity of statin-induced adverse effects on muscle and associated conditions: Data from the DAMA study. Expert Opin. Drug Saf. 2016, 15, 1583–1587. [Google Scholar] [CrossRef]
- Yan, F.; Wang, Q.; Xu, C.; Cao, M.; Zhou, X.; Wang, T.; Yu, C.; Jing, F.; Chen, W.; Gao, L.; et al. Peroxisome proliferator-activated receptor alpha activation induces hepatic steatosis, suggesting an adverse effect. PLoS ONE 2014, 9, e99245. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Guo, J.X.; Meng, Q.L.; Cui, X.Y. Physiological responses of krishum (Iris lactea Pall. var. Chinensis Koidz) to neutral and alkaline salts. J. Agron. Crop. Sci. 2008, 194, 429–437. [Google Scholar] [CrossRef]
- Zhou, T.R.; Ge, G.T.; Jia, Y.S.; Hou, M.L.; Wang, W.; Nuo, M.; Ba, D.L. The effect advantage natural grassland on mixed grass group of silage quality. Grassl. Prataculture 2015, 27, 19–26. [Google Scholar]
- Tie, F.F.; Luan, G.X.; Zhou, W.N.; Wang, Z.H.; Shi, X.B.; Li, G.; Wang, H.L. Effects of the oligostilbenes from Iris lactea Pall. var. Chinensis Koidz on the adipocytes differentiation of 3T3-L1 cells. Pharmazie 2018, 73, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Sun, G.; Wang, H.; Yue, Q.; Tang, H.; Wu, Q. Dynamic observation of the radiosensitive effect of Irisquinone on rabbit VX2 lung transplant tumors by using fluorine-18-deoxyglucose positron emission tomography/computed tomography. Nucl. Med. Commun. 2013, 34, 220–228. [Google Scholar] [CrossRef]
- Zhu, W.H.; Sun, W.K.; Yu, Y.C. The impact of radiosensitizer irisquinone on lung metastasis in H22-bearing mice. Jiangsu Med. J. 2008, 34, 176–178. [Google Scholar]
- Wang, H.L.; Mao, Y.; Liang, J.; Wang, Y.; Li, N.; Liu, W.Q. Irisquinone protects against radiation-induced lung injury in Rats. Chin. J. Exp. Trad. Med. Formulae 2016, 22, 151–159. [Google Scholar]
- Bi, J.T.; Pan, X.; Huang, P.P.; Ma, F.; Guan, X.Q. Isolation of endophytic fungi from Iris lactea var. chinensis and its microbial inhibition activity. J. Anhui Agri. Sci. 2012, 40, 15651–15654. [Google Scholar]
- Zhang, F.G.; Li, D.H.; Qi, J.; Liu, C.X. In vitro anticancer effects of pallasone A and its induced apoptosis on leukemic K562 cells. Chin. Pharm. J. 2010, 22, 1716–1719. [Google Scholar]
- Jiang, X.G.; Hou, D.Y.; Weng, X.; Wang, C.Y. Process optimization for ultrasonic extraction of lavonoids and determination of antioxidation effect in Iris lactea Pall.Var. chinensis Koidz. Mod. Agric. Sci. Technol. 2014, 2, 301–303. [Google Scholar]
- Meng, Y.; Qin, M.; Qi, B.; Xie, G. Four new C -glycosylflavones from the leaves of Iris lactea Pall. var. Chinensis (Fisch.) Koidz. Phytochem. Lett. 2017, 22, 33–38. [Google Scholar] [CrossRef]
- Shen, W.; Qin, M.; Shu, P.; Zhang, C. Two new C-glycosylflavones from the leaves of Iris lactea var. Chinensis. Chin. Chem. Lett. 2008, 19, 821–824. [Google Scholar] [CrossRef]
- He, F.; Pan, Q.H.; Shi, Y.; Duan, C.Q. Biosynthesis and genetic regulation of proanthocyanidins in plants. Molecules 2008, 13, 2674–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvado, M.J.; Casanova, E.; Fernandez-Iglesias, A.; Arola, L.; Blade, C. Roles of proanthocyanidin rich extracts in obesity. Food Funct. 2015, 6, 1053–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, S.L.; Shaw, O.M. Progress in the understanding of the pathology of allergic asthma and the potential of fruit proanthocyanidins as modulators of airway inflammation. Food Funct. 2017, 8, 4315–4324. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Sturzenbaum, S.R. Proanthocyanidins of natural origin: Molecular mechanisms and implications for lipid disorder and aging-associated diseases. Adv. Nutr. 2019, 10, 464–478. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Han, X.; Li, R.; Zhao, W.; Bai, B.; Yan, C.; Dong, X. Anti-atherosclerosis of oligomeric proanthocyanidins from Rhodiola rosea on rat model via hypolipemic, antioxidant, anti-inflammatory activities together with regulation of endothelial function. Phytomedicine 2018, 51, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Ouyang, J.; Wang, X.; Ma, X.; Suo, Y.; Wang, H. Separation and purification of four flavan-3-ols from Iris lactea Pall. var. Chinensis (Fisch.) Koidz by high-speed counter-current chromatography with flow-rate gradient. J. Liq. Chromatogr. Relat. Technol. 2015, 38, 1486–1493. [Google Scholar] [CrossRef]
- Lv, H.; Yuan, Z.; Wang, X.; Wang, Z.; Suo, Y.; Wang, H. Rapid separation of three proanthocyanidin dimers from Iris lactea Pall. var. Chinensis (Fisch.) Koidz by high-speed counter-current chromatography with continuous sample load and double-pump balancing mode. Phytochem. Anal. 2015, 26, 444–453. [Google Scholar] [CrossRef]
- Angin, Y.; Beauloye, C.; Horman, S.; Bertrand, L. Regulation of carbohydrate metabolism, lipid metabolism, and protein metabolism by AMPK. Exp. Suppl. 2016, 107, 23–43. [Google Scholar] [CrossRef]
- Wu, M.; Liu, D.; Zeng, R.; Xian, T.; Lu, Y.; Zeng, G.; Sun, Z.; Huang, B.; Huang, Q. Epigallocatechin-3-gallate inhibits adipogenesis through down-regulation of PPARgamma and FAS expression mediated by PI3K-AKT signaling in 3T3-L1 cells. Eur. J. Pharmacol. 2017, 795, 134–142. [Google Scholar] [CrossRef]
- Moseti, D.; Regassa, A.; Kim, W.K. Molecular regulation of adipogenesis and potential anti-adipogenic bioactive molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.A.; Wei, Y.; Sowers, J.R. Role of mitochondrial dysfunction in insulin resistance. Circ Res. 2008, 102, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Kizhakekuttu, T.J.; Wang, J.L.; Dharmashankar1, K.; Ying, R.; Gutterman, D.D.; Vita, J.A.; Widlansky, M.E. Adverse alterations in mitochondrial function contribute to type 2 diabetes induced endothelial dysfunction in human. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2531–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Tang, B.L. Sirt1 and the mitochondria. Mol. Cells 2016, 39, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Chan, D.C. Mitochondrial dynamics--fusion, fission, movement, and mitophagy--in neurodegenerative diseases. Hum. Mol. Genet. 2009, 18, R169–R176. [Google Scholar] [CrossRef]
- Ding, Y.; Dai, X.; Jiang, Y.; Zhang, Z.; Bao, L.; Li, Y.; Zhang, F.; Ma, X.; Cai, X.; Jing, L.; et al. Grape seed proanthocyanidin extracts alleviate oxidative stress and ER stress in skeletal muscle of low-dose streptozotocin- and high-carbohydrate/high-fat diet-induced diabetic rats. Mol. Nutr. Food Res. 2013, 57, 365–369. [Google Scholar] [CrossRef]
- Li, X.; Sui, Y.; Li, S.; Xie, B.; Sun, Z. A-type procyanidins from litchi pericarp ameliorate hyperglycaemia by regulating hepatic and muscle glucose metabolism in streptozotocin (STZ)-induced diabetic mice fed with high fat diet. J. Funct. Foods 2016, 27, 711–722. [Google Scholar] [CrossRef]
- Quesada, H.; del Bas, J.M.; Pajuelo, D.; Diaz, S.; Fernandez-Larrea, J.; Pinent, M.; Arola, L.; Salvado, M.J.; Blade, C. Grape seed proanthocyanidins correct dyslipidemia associated with a high-fat diet in rats and repress genes controlling lipogenesis and VLDL assembling in liver. Int. J. Obes. (Lond) 2009, 33, 1007–1012. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, K.; Viswanad, B.; Asrat, L.; Kaul, C.L.; Ramarao, P. Combination of high-fat diet-fed and low-dose streptozotocin-treated rat: A model for type 2 diabetes and pharmacological screening. Pharmacol. Res. 2005, 52, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liao, G.; Yang, G.; Lu, Y.; Du, X.; Liu, J.; Li, L.; Wang, C.; Li, L.; Ren, Y.; et al. High-fat diet combined with low-dose streptozotocin injections induces metabolic syndrome in Macaca mulatta. Endocrine 2015, 49, 659–668. [Google Scholar] [CrossRef]
- Komiya, C.; Tsuchiya, K.; Shiba, K.; Miyachi, Y.; Furuke, S.; Shimazu, N.; Yamaguchi, S.; Kanno, K.; Ogawa, Y. Ipragliflozin improves hepatic steatosis in obese mice and liver dysfunction in type 2 diabetic patients irrespective of body weight reduction. PLoS ONE 2016, 11, e0151511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, J.; Guan, K.L.; Kim, J. AMPK and autophagy in glucose/glycogen metabolism. Mol. Aspects Med. 2015, 46, 46–62. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.; Kahn, C.R. Transplantation of adipose tissue and stem cells: Role in metabolism and disease. Nat. Rev. Endocrinol. 2010, 6, 195–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.Z.; Ma, R.L.; Zhang, J.Y.; He, J.; Ma, J.L.; Pang, H.R.; Mu, L.T.; Ding, Y.S.; Guo, H.; Zhang, M.; et al. Association of insulin resistance with glucose and lipid metabolism: Ethnic heterogeneity in far western China. Mediators Inflamm. 2016, 2016, e3825037. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A. Insulin resistance, lipotoxicity, type 2 diabetes and atherosclerosis: The missing links. The Claude Bernard Lecture 2009. Diabetologia 2010, 53, 1270–1287. [Google Scholar] [CrossRef] [Green Version]
- Choo, H.J.; Kim, J.H.; Kwon, O.B.; Lee, C.S.; Mun, J.Y.; Han, S.S.; Yoon, Y.S.; Yoon, G.; Choi, K.M.; Ko, Y.G. Mitochondria are impaired in the adipocytes of type 2 diabetic mice. Diabetologia 2006, 49, 784–791. [Google Scholar] [CrossRef] [Green Version]
- Suwanjang, W.; Wu, K.L.H.; Prachayasittikul, S.; Chetsawang, B.; Charngkaew, K. Mitochondrial dynamics impairment in dexamethasone-treated neuronal cells. Neurochem. Res. 2019, 44, 1567–1581. [Google Scholar] [CrossRef]
- Chan, D.C. Mitochondrial fusion and fission in mammals. Annu. Rev. Cell Dev. Biol. 2006, 22, 79–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belenguer, P.; Pellegrini, L. The dynamin GTPase OPA1: More than mitochondria? Biochim. Biophys. Acta. 2013, 1833, 176–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, S.L. Mitochondrial dynamics--mitochondrial fission and fusion in human diseases. N. Engl. J. Med. 2013, 369, 2236–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, G.; Wang, Y.; Wang, Z.; Zhou, W.; Hu, N.; Li, G.; Wang, H. Flavonoid glycosides from fenugreek seeds regulate glycolipid metabolism by improving mitochondrial function in 3T3-L1 adipocytes in vitro. J. Agric. Food Chem. 2018, 66, 3169–3178. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.C.; Huang, C.C.; Lin, W.T.; Hsieh, C.C.; Huang, S.Y.; Lin, S.J.; Yang, S.C. Lycopene supplementation attenuated xanthine oxidase and myeloperoxidase activities in skeletal muscle tissues of rats after exhaustive exercise. Br. J. Nutr. 2005, 94, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ames, B.N. Reducing mitochondrial decay with mitochondrial nutrients to delay and treat cognitive dysfunction, Alzheimer’s disease, and Parkinson’s disease. Nutr. Neurosci. 2005, 8, 67–89. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Talalay, P. Induction of phase 2 genes by sulforaphane protects retinal pigment epithelial cells against photooxidative damage. Proc. Natl. Acad. Sci. USA 2004, 101, 10446–10451. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Hao, J.; Tian, C.; Ren, J.; Yang, L.; Li, X.; Luo, C.; Cotma, C.W.; Liu, J. A combination of nutriments improves mitochondrial biogenesis and function in skeletal muscle of type 2 diabetic Goto-Kakizaki rats. PLoS ONE 2008, 3, e2328. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Liu, K.; Tian, C.; Yang, L.; Li, X.; Ren, J.; Packer, L.; Cotman, C.W.; Liu, J. R-alpha-lipoic acid and acetyl-L-carnitine complementarily promote mitochondrial biogenesis in murine 3T3-L1 adipocytes. Diabetologia 2008, 51, 165–174. [Google Scholar] [CrossRef]
- Prince, P.S. (-) Epicatechin prevents alterations in lysosomal glycohydrolases, cathepsins and reduces myocardial infarct size in isoproterenol-induced myocardial infarcted rats. Eur. J. Pharmacol. 2013, 706, 63–69. [Google Scholar] [CrossRef]
- Rowley, T.J.; Bitner, B.F.; Ray, J.D.; Lathen, D.R.; Smithson, A.T.; Dallon, B.W.; Plowman, C.J.; Bikman, B.T.; Hansen, J.M.; Dorenkott, M.R.; et al. Monomeric cocoa catechins enhance beta-cell function by increasing mitochondrial respiration. J. Nutr. Biochem. 2017, 49, 30–41. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Savickas, A.; Vetchy, D.; Masteikova, R.; Kasauskas, A.; Bernatoniene, J. Direct effects of (-)-epicatechin and procyanidin B2 on the respiration of rat heart mitochondria. Biomed. Res. Int. 2015, 2015, e232836. [Google Scholar] [CrossRef] [Green Version]
- Butler, L.G.; Price, M.L.; Brotherton, J.E. Vanillin assay for proanthocyanidins (condensed tannins): Modification of the solvent for estimation of the degree of polymerization. J. Agric. Food Chem. 1982, 30, 1087–1089. [Google Scholar] [CrossRef]
- Guaita, M.; Bosso, A. Polyphenolic characterization of grape skins and seeds of four Italian red cultivars at harvest and after fermentative maceration. Foods 2019, 8, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; Yao, K.; Jia, D.; Fan, H.; Liao, X.; Shi, B. Determination of total catechins in tea extracts by HPLC and spectrophotometry. Nat. Prod. Res. 2009, 23, 93–100. [Google Scholar] [CrossRef]
- Gilbert, E.R.; Fu, Z.; Liu, D. Development of a nongenetic mouse model of type 2 diabetes. Exp. Diabetes Res. 2011, 2011, e416254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Chen, D.; Xie, Q.; Yang, Y.; Shen, W. Nebivolol stimulates mitochondrial biogenesis in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2013, 438, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Hao, J.; Feng, Z.; Tian, C.; Chen, W.; Packer, L.; Shi, X.; Zang, W.; Liu, J. Lipoamide or lipoic acid stimulates mitochondrial biogenesis in 3T3-L1 adipocytes via the endothelial NO synthase-cGMP-protein kinase G signalling pathway. Br. J. Pharmacol. 2011, 162, 1213–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, W.; Li, X.; Hull, L.; Xiao, M. Measuring radiation-induced DNA damage in Cryptococcus neoformans and Saccharomyces cerevisiae using long range quantitative PCR. PLoS ONE 2018, 13, e0207071. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Category | tz (min) | M + (m/z) | MS/MS (m/z) |
|---|---|---|---|---|
| 1 | 3,4-Dihydroxybenzoic acid | 4.48 | 153.0218 | 65/91/109 |
| 2 | procyanidin B1 | 6.40 | 577.1349 | 125/245/289/407/425/451 |
| 3 | 3’-O-(1-hydroxy-6-oxo-2-cyclohexene-1-carboxylic acid ester) of procyanidin B1 | 6.63 | 715.1663 | 289/419/425/571 |
| 4 | 6-[(1S)-3-methoxy-3-oxo-1-(2,4,5-trihydroxyphenyl)propyl]catechin | 6.68 | 499.1233 | 245/289/345/389 |
| 5 | procyanidin B3 | 6.87 | 577.1342 | 125/245/289/407/425/451 |
| 6 | catechin | 7.29 | 289.0720 | 109/125/203/245 |
| 7 | fisetinidol (4α,8)-catechin | 7.92 | 577.1342 | 125/245/289/407/435 |
| 8 | epiafzelechin-(4β- > 8)-epicatechin | 8.66 | 561.1394 | 125/245/289/407/435 |
| 9 | epicatechin | 9.37 | 289.0722 | 109/125/203/245 |
| 10 | procyanidin B7 | 10.61 | 577.1345 | 125/245/289/407/425/451 |
| 11 | procyanidin A1 | 11.01 | 591.1136 | 125/289/407/465 |
| 12 | norathyriol | 12.35 | 259.0257 | 159/191/231 |
| 13 | proanthocyanidin A1 | 12.82 | 575.1186 | 125/289/407/449 |
| 14 | procyanidin B6 | 13.14 | 577.1185 | 125/289/407/449 |
| 15 | (2’S,3’R)-9-(5’,6’-dihydroxy-2’-hydroxymethyl-2’,3’-dihydrobenzo[b]furan-3-yloxy)-6H-dibenzo[b,d]pyran-6-oe | 14.93 | 575.1185 | 191/258/299/355 |
| 16 | hopeaphenol | 19.91 | 905.2597 | 265/358/451/717/811 |
| 17 | isohopeaphenol | 20.49 | 905.2605 | 265/359/451/717/811 |
| 18 | n-butyl pro-lithospermate | 21.11 | 413.1239 | 145/218/233/367 |
| 19 | viniferin | 21.84 | 453.1332 | 197/225/279/345/359 |
| 20 | vitisin Β | 22.55 | 905.2603 | 359/451/545/799811 |
| 21 | vitisin C | 23.43 | 905.2602 | 359/545/693/799811 |
| Target Gene | Primer Sequence | Size (bp) | Accession Numbers |
|---|---|---|---|
| PPARγ | Forward: 5’-CCTGGCAAAGCATTTCTATG-3’ | 100 | XM_017321456 |
| Reverse: 5’-TGGTGATTTGTCCGTTGTCT-3’ | |||
| C/EBPα | Forward: 5’- CGGCGGGAACGCAACAACAT -3’ | 109 | NM_001287514 |
| Reverse: 5’- GGCGGTCATTGTCACTGGTC -3’ | |||
| FABP4 | Forward: 5’-TCACCTGGAAGACAGCTCCT-3’ | 182 | XM_024406 |
| Reverse: 5’-AATCCCCATTTACGCTGAT-3’ | |||
| PGC-1α | Forward: 5’-CGGAAATCATATCCAACCAG-3’ | 243 | XM_006503779 |
| Reverse: 5’-TGAGGACCGCTAGCAAGTTTG-3’ | |||
| NRF1 | Forward: 5’-TGGTCCAGAGAGTGCTTGTG-3’ | 184 | NM_001361693 |
| Reverse: 5’-TTCCTGGGAAGGGAGAAGAT-3’ | |||
| Tfam | Forward:5’-GGAATGTGGAGCGTGCTAAAA-3’ | 118 | NM_009360 |
| Reverse:5’-TGCTGGAAAAACACTTCGGAATA-3’ | |||
| β-actin | Forward: 5’-CCTGAGGCTCTTTTCCAGCC-3’ | 110 | NM_007393 |
| Reverse:5’-TAGAGGTCTTTACGGATGTCAACGT-3’ | |||
| Long fragment | Forward: 5’-TACTAGTCCGCGAGCCTTCAAAGC-3’ | 8636 | AJ512208.1 |
| Reverse: 5’-GGGTGATCTTTGTTTGCGGGT-3’ | |||
| Short fragment | Forward: 5’-CCCAGCTACTACCATCATTCAAGT-3’ | 117 | NC_005089 |
| Reverse: 5’-GATGGTTTGGGAGATTGGTTGATG-3’ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tie, F.; Wang, J.; Liang, Y.; Zhu, S.; Wang, Z.; Li, G.; Wang, H. Proanthocyanidins Ameliorated Deficits of Lipid Metabolism in Type 2 Diabetes Mellitus Via Inhibiting Adipogenesis and Improving Mitochondrial Function. Int. J. Mol. Sci. 2020, 21, 2029. https://doi.org/10.3390/ijms21062029
Tie F, Wang J, Liang Y, Zhu S, Wang Z, Li G, Wang H. Proanthocyanidins Ameliorated Deficits of Lipid Metabolism in Type 2 Diabetes Mellitus Via Inhibiting Adipogenesis and Improving Mitochondrial Function. International Journal of Molecular Sciences. 2020; 21(6):2029. https://doi.org/10.3390/ijms21062029
Chicago/Turabian StyleTie, Fangfang, Jifei Wang, Yuexin Liang, Shujun Zhu, Zhenhua Wang, Gang Li, and Honglun Wang. 2020. "Proanthocyanidins Ameliorated Deficits of Lipid Metabolism in Type 2 Diabetes Mellitus Via Inhibiting Adipogenesis and Improving Mitochondrial Function" International Journal of Molecular Sciences 21, no. 6: 2029. https://doi.org/10.3390/ijms21062029
APA StyleTie, F., Wang, J., Liang, Y., Zhu, S., Wang, Z., Li, G., & Wang, H. (2020). Proanthocyanidins Ameliorated Deficits of Lipid Metabolism in Type 2 Diabetes Mellitus Via Inhibiting Adipogenesis and Improving Mitochondrial Function. International Journal of Molecular Sciences, 21(6), 2029. https://doi.org/10.3390/ijms21062029