Versican G1 Fragment Establishes a Strongly Stabilized Interaction with Hyaluronan-Rich Expanding Matrix during Oocyte Maturation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Versican Fragments Containing the Neo-Epitope DPEAAE in OCCs Expanded in Vivo
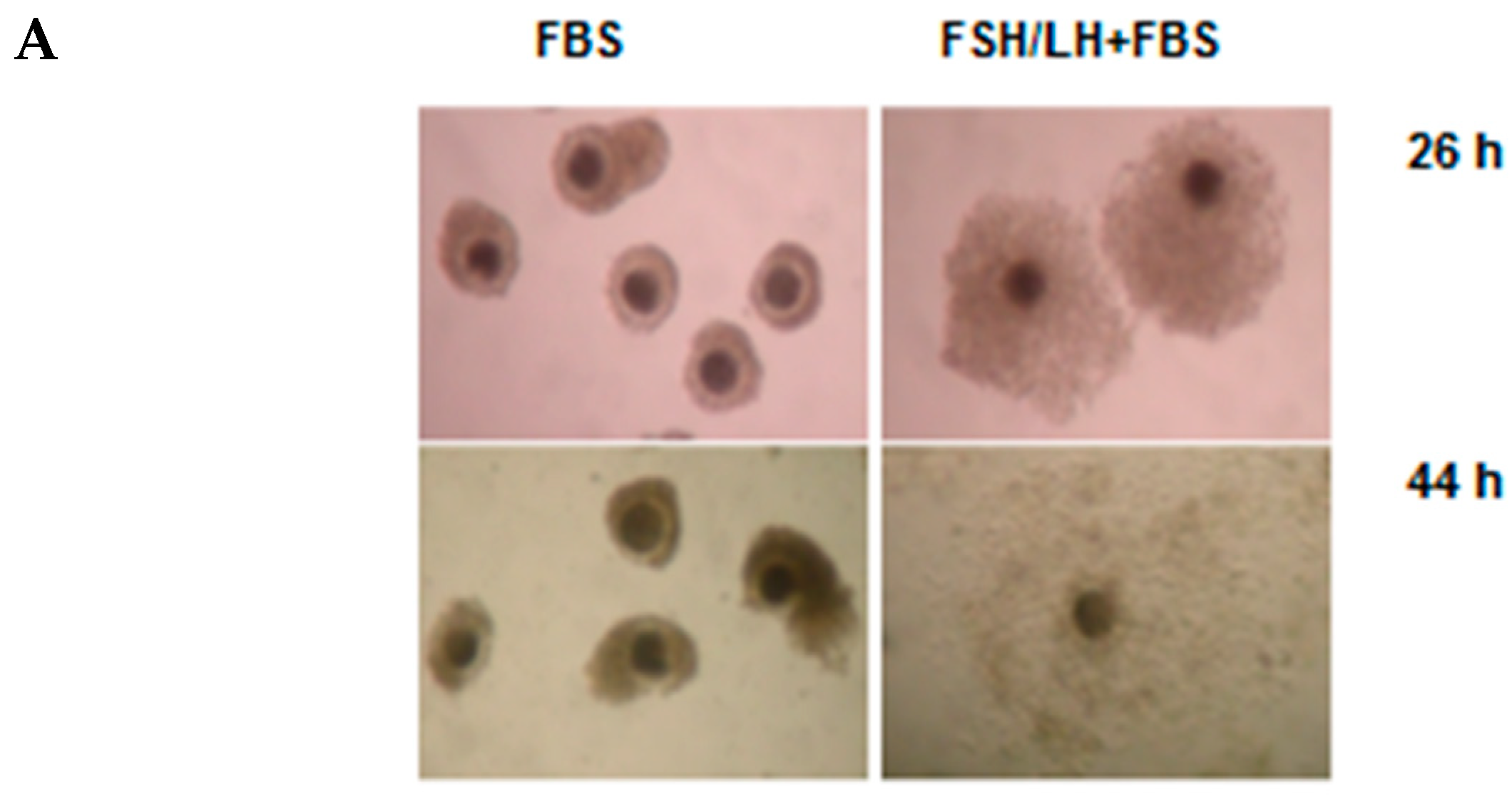
2.2. Versican Fragments Produced by Gonadotropin-Stimulated OCCs Cultured in Vitro
2.3. Temporal Pattern of Versican Cleavage during in Vitro Expansion
2.4. Versican Fragments Produced by Mural Granulosa Cells after in Vitro Gonadotropin Stimulation
3. Discussion
4. Materials and Methods
4.1. Isolation and Culture of Oocyte-Cumulus Complexes and Mural Granulosa Cells
4.2. RNA Isolation
4.3. Real-Time (RT)-PCR
4.4. Western Blot Analysis
4.5. Immunofluorescence Analysis
4.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ECM | Extracellular matrix |
| HA | Hyaluronan |
| OCC | Oocyte-cumulus complex |
| FSH | Follicole-stimulating hormone |
| LH | Luteinizing hormone |
| Hyal | Hyaluronidase from Streptomyces hyaluronlyticus |
| ChABC | Chondroitinase ABC |
| V1 | Versican 1 |
| VG1 | G1-domain from Versican 1 |
| ADAMTS | A disintegrin and metalloproteinase with trombospondin like repeats |
| FF | Follicular fluid |
| PVP | Polyvinylpyrolidone |
| IαI | Inter-alpha-trypsin inhibitor |
| HCs | Heavy Chains from Inter-alpha.trypsin inhibitor |
| TNFAIP6 | Tumor necrosis factor alpha-induced protein 6 |
| PTX3 | Pentraxin 3 |
References
- Richards, J.S.; Russell, D.L.; Ochsner, S.; Hsieh, M.; Doyle, K.H.; Falender, A.E.; Lo, Y.K.; Sharma, S.C. Novel signaling pathways that control ovarian follicular development, ovulation, and luteinization. Recent Prog. Horm. Res. 2002, 57, 195–220. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.L.; Salustri, A. Extracellular matrix of the cumulus-oocyte complex. Semin. Reprod. Med. 2006, 24, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Nagyova, E. The biological role of hyaluronan-rich oocyte-cumulus extracellular matrix in female reproduction. Int. J. Mol. Sci. 2018, 19, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camaioni, A.; Salustri, A.; Yanagishita, M.; Hascall, V.C. Proteoglycans and proteins in the extracellular matrix of mouse cumulus cell-oocyte complexes. Arch. Biochem. Biophys. 1996, 325, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Nagyova, E.; Camaioni, A.; Prochazka, R.; Salustri, A. Covalent transfer of heavy chains of inter-alpha-trypsin inhibitor family proteins to hyaluronan in in vivo and in vitro expanded porcine oocyte-cumulus complexes. Biol. Reprod. 2004, 71, 1838–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagyova, E.; Camaioni, A.; Prochazka, R.; Day, A.J.; Salustri, A. Synthesis of tumor necrosis factor alpha-induced protein 6 in porcine preovulatory follicles: A study with A38 antibody. Biol. Reprod. 2008, 78, 903–909. [Google Scholar] [CrossRef] [Green Version]
- McArthur, M.E.; Irving-Rodgers, H.F.; Byers, S.; Rodgers, R.J. Identification and immunolocalization of decorin, versican, perlecan, nidogen, and chondroitin sulfate proteoglycans in bovine small-antral ovarian follicles. Biol. Reprod. 2000, 63, 913–924. [Google Scholar] [CrossRef] [Green Version]
- Russell, D.L.; Doyle, K.M.; Ochsner, S.A.; Sandy, J.D.; Richards, J.S. Processing and localization of ADAMTS-1 and proteolytic cleavage of versican during cumulus matrix expansion and ovulation. J. Biol. Chem. 2003, 278, 42330–42339. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, R.J.; Irving-Rodgers, H.F. Formation of the ovarian follicular antrum and follicular fluid. Biol. Reprod. 2010, 82, 1021–1029. [Google Scholar] [CrossRef] [Green Version]
- Foulcer, S.J.; Day, A.J.; Apte, S.S. Isolation and purification of versican and analysis of versican proteolysis. Methods Mol. Biol. 2015, 1229, 587–604. [Google Scholar] [CrossRef] [Green Version]
- Ito, K.; Shinomura, T.; Zako, M.; Ujita, M.; Kimata, K. Multiple forms of mouse PG-M, a large chondroitin sulfate proteoglycan generated by alternative splicing. J. Biol. Chem. 1995, 270, 958–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmalfeldt, M.; Dours-Zihmermann, M.T.; Winterhalter, K.H.; Zimmermann, D.R. Versican V2 is a major extracellular matrix component of the mature bovine brain. J. Biol. Chem. 1998, 273, 15758–15764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kresse, H.; Schonherr, E. Proteoglycans of the extracellular matrix and growth control. J. Cell. Physiol. 2001, 189, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Wight, T.N. Versican: A versatile extracellular matrix proteoglycan in cell biology. Curr. Opin. Cell Biol. 2002, 14, 617–623. [Google Scholar] [CrossRef]
- Lemire, J.M.; Merrilees, M.J.; Braun, K.R.; Wight, T.N. Overexpression of the V3 variant of versican alters arterial smooth muscle cell adhesion, migration, and proliferation in vitro. J. Cell. Physiol. 2002, 190, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Sandy, J.D.; Westling, J.; Kenagy, R.D.; Iruela-Arispe, M.L.; Verscharen, C.; Rodriguez-Mazaneque, J.C.; Zimmermann, D.R.; Lemire, J.M.; Fischer, J.W.; Wight, T.N.; et al. Versican V1 proteolysis in human aorta in vivo occurs at the Glu441-Ala442 bond, a site that is cleaved by recombinant ADAMTS-1 and ADAMTS-4. J. Biol. Chem. 2001, 276, 13372–13378. [Google Scholar] [CrossRef] [Green Version]
- Wight, T.N. Provisional matrix: A role for versican and hyaluronan. Matrix Biol. 2017, 60–61, 38–56. [Google Scholar] [CrossRef]
- Murasawa, Y.; Nakamura, H.; Watanabe, K.; Kanoh, H.; Koyama, E.; Fujii, S.; Kimata, K.; Zako, M.; Yoneda, M.; Isogai, Z. The Versican G1 fragment and serum-derived hyaluronan-associated proteins interact and form a complex in granulation tissue of pressure ulcers. Am. J. Pathol. 2018, 188, 432–449. [Google Scholar] [CrossRef] [Green Version]
- Toole, B.P.; Wight, T.N.; Tammi, M.I. Hyaluronan-cell interactions in cancer and vascular disease. J. Biol. Chem. 2002, 277, 4593–4596. [Google Scholar] [CrossRef] [Green Version]
- Russell, D.L.; Ochsner, S.A.; Hsieh, M.; Mulders, S.; Richards, J.S. Hormone-regulated expression and localization of versican in the rodent ovary. Endocrinology 2003, 144, 1020–1031. [Google Scholar] [CrossRef]
- Dunning, K.R.; Lane, M.; Brown, H.M.; Yeo, C.; Robker, R.L.; Russell, D.L. Altered composition of the cumulus-oocyte complex matrix during in vitro maturation of oocytes. Hum. Reprod. 2007, 22, 2842–2850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, M.; Nishibori, M.; Yamashita, Y.; Ito, J.; Mori, T.; Richards, J.S. Down-regulated expression of A disintegrin and metalloproteinase with thrombospondin-like repeats-1 by progesterone receptor antagonist is associated with impaired expansion of porcine cumulus-oocyte complexes. Endocrinology 2004, 145, 4603–4614. [Google Scholar] [CrossRef] [PubMed]
- Nagyova, E. Organization of the expanded cumulus-extracellular matrix in preovulatory follicles: A role for inter-alpha-trypsin inhibitor. Endocr. Regul. 2015, 49, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Baranova, N.S.; Foulcer, S.J.; Briggs, D.C.; Tilakaratna, V.; Enghild, J.J.; Milner, C.M.; Day, A.J.; Richter, R.P. Inter-α-inhibitor impairs TSG-6induced hyaluronan cross-linking. J. Biol. Chem. 2013, 288, 29642–29653. [Google Scholar] [CrossRef] [Green Version]
- Salustri, A.; Campagnolo, L.; Klinger, F.G.; Camaioni, A. Molecular organization and mechanical properties of the hyaluronan matrix surrounding the mammalian oocyte. Matrix Biol. 2019, 78, 11–23. [Google Scholar] [CrossRef]
- Chen, X.; Bonfiglio, R.; Banerji, S.; Jackson, D.G.; Salustri, A.; Richter, R.P. Micromechanical analysis of the hyaluronan-rich matrix surrounding the oocyte reveals a uniquely soft and elastic composition. Biophys. J. 2016, 110, 2779–2789. [Google Scholar] [CrossRef] [Green Version]
- Stanton, H.; Melrose, J.; Little, C.B.; Fosang, A.J. Proteoglycan degradation by the ADAMTS family of proteinases. Biochim. Biophys. Acta 2011, 812, 1616–1629. [Google Scholar] [CrossRef] [Green Version]
- Nagyova, E. Regulation of cumulus expansion and hyaluronan synthesis in porcine oocyte-cumulus complexes during in vitro maturation. Endocr. Regul. 2012, 46, 225–235. [Google Scholar] [CrossRef]
- D’Alessandris, C.; Canipari, R.; Di Giacomo, M.; Epifano, O.; Camaioni, A.; Siracusa, G.; Salustri, A. Control of mouse cumulus cell-oocyte complex integrity before and after ovulation: Plasminogen activator synthesis and matrix degradation. Endocrinology 2001, 142, 3033–3040. [Google Scholar] [CrossRef]
- Matsumoto, K.; Shionyu, M.; Go, M.; Shimizu, K.; Shinomura, T.; Kimata, K.; Watanabe, H. Distinct interaction of versican/PG-M with hyaluronan and link protein. J. Biol. Chem. 2003, 278, 41205–41212. [Google Scholar] [CrossRef] [Green Version]
- Yanagishita, M.; Rodbard, D.; Hascall, V.C. Isolation and characterization of proteoglycans from porcine ovarian follicular fluid. J. Biol. Chem. 1979, 254, 911–920. [Google Scholar] [PubMed]
- Yanagishita, M.; Hascall, V.C. Biosynthesis of proteoglycans by rat granulosa cells cultured in vitro. J. Biol. Chem. 1979, 254, 12355–12364. [Google Scholar] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagyova, E.; Salustri, A.; Nemcova, L.; Scsukova, S.; Kalous, J.; Camaioni, A. Versican G1 Fragment Establishes a Strongly Stabilized Interaction with Hyaluronan-Rich Expanding Matrix during Oocyte Maturation. Int. J. Mol. Sci. 2020, 21, 2267. https://doi.org/10.3390/ijms21072267
Nagyova E, Salustri A, Nemcova L, Scsukova S, Kalous J, Camaioni A. Versican G1 Fragment Establishes a Strongly Stabilized Interaction with Hyaluronan-Rich Expanding Matrix during Oocyte Maturation. International Journal of Molecular Sciences. 2020; 21(7):2267. https://doi.org/10.3390/ijms21072267
Chicago/Turabian StyleNagyova, Eva, Antonietta Salustri, Lucie Nemcova, Sona Scsukova, Jaroslav Kalous, and Antonella Camaioni. 2020. "Versican G1 Fragment Establishes a Strongly Stabilized Interaction with Hyaluronan-Rich Expanding Matrix during Oocyte Maturation" International Journal of Molecular Sciences 21, no. 7: 2267. https://doi.org/10.3390/ijms21072267
APA StyleNagyova, E., Salustri, A., Nemcova, L., Scsukova, S., Kalous, J., & Camaioni, A. (2020). Versican G1 Fragment Establishes a Strongly Stabilized Interaction with Hyaluronan-Rich Expanding Matrix during Oocyte Maturation. International Journal of Molecular Sciences, 21(7), 2267. https://doi.org/10.3390/ijms21072267