Transcriptional Profiling of the Probiotic Escherichia coli Nissle 1917 Strain under Simulated Microgravity

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
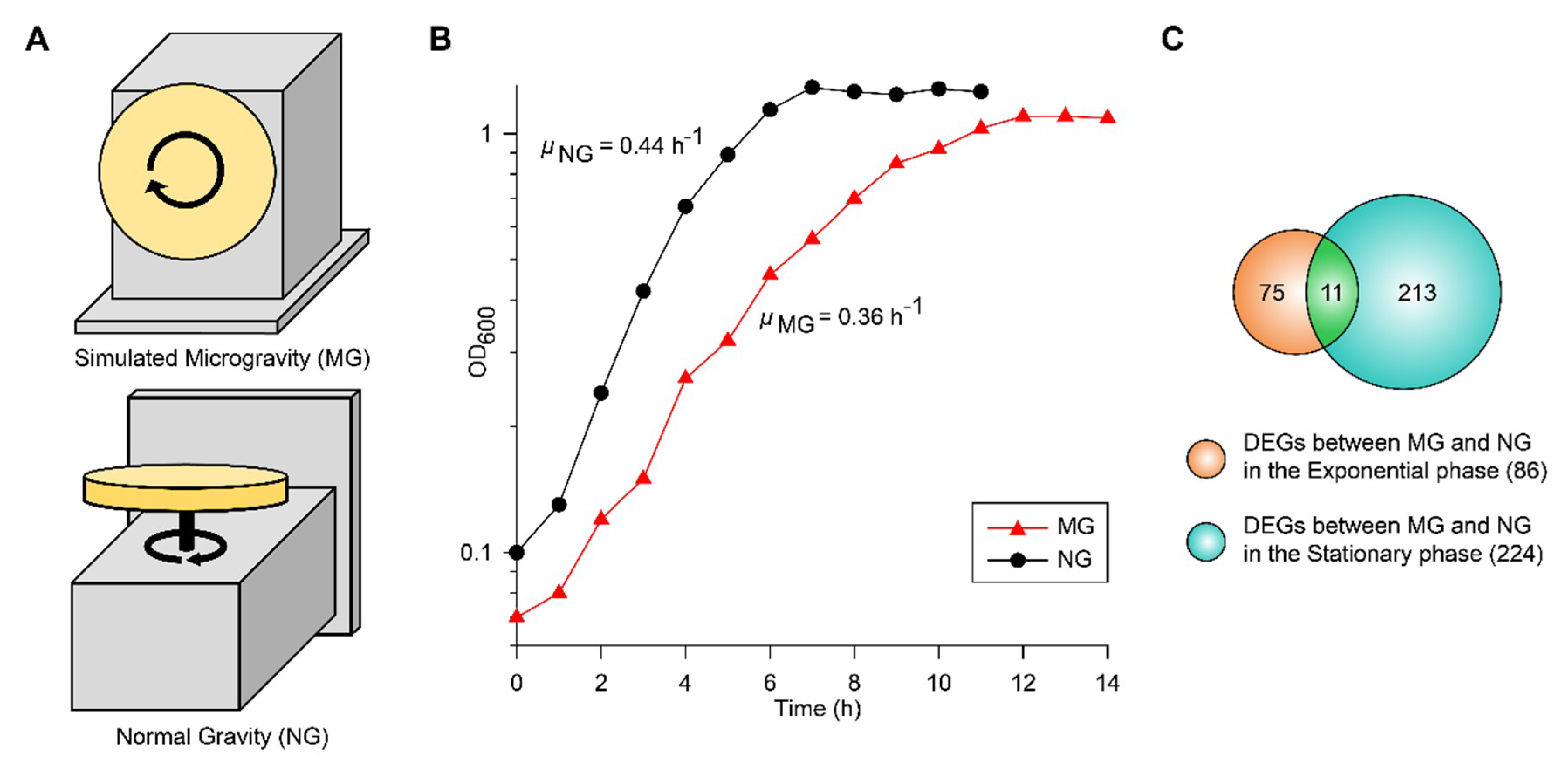
2.1. Growth and Transcriptomic Profiling of EcN under Simulated MG and NG
2.2. Effect of MG on EcN Biofilm Formation
2.3. Effect of MG on EcN Stress Responses
2.4. Effect of MG on EcN Metal Ion Transport/Utilization
2.5. Effect of MG on EcN Carbon Metabolism
3. Conclusions
4. Materials and Methods
4.1. Overall Work Flow of This Study
4.2. Bacterial Strain and Growth Conditions
4.3. Growth of EcN in a Clinostat
4.4. RNA Extraction, cDNA Synthesis, and Sequencing
4.5. Read Processing, Alignment, and Quantification
4.6. Differential Expression and Functional Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Garrett-Bakelman, F.E.; Darshi, M.; Green, S.J.; Gur, R.C.; Lin, L.; Macias, B.R.; McKenna, M.J.; Meydan, C.; Mishra, T.; Nasrini, J.; et al. The NASA Twins Study: A multidimensional analysis of a year-long human spaceflight. Science 2019, 364, eaau8650. [Google Scholar] [PubMed]
- Voorhies, A.A.; Mark Ott, C.; Mehta, S.; Pierson, D.L.; Crucian, B.E.; Feiveson, A.; Oubre, C.M.; Torralba, M.; Moncera, K.; Zhang, Y.; et al. Study of the impact of long-duration space missions at the International Space Station on the astronaut microbiome. Sci. Rep. 2019, 9, 9911. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.W.; Ott, C.M.; Quick, L.; Davis, R.; Honer zu Bentrup, K.; Crabbe, A.; Richter, E.; Sarker, S.; Barrila, J.; Porwollik, S.; et al. Media ion composition controls regulatory and virulence response of Salmonella in spaceflight. PLoS ONE 2008, 3, e3923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, M.D.; Fajardo-Cavazos, P.; Nicholson, W.L. Comparison of Bacillus subtilis transcriptome profiles from two separate missions to the International Space Station. NPJ Microgravity 2019, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Crabbe, A.; Schurr, M.J.; Monsieurs, P.; Morici, L.; Schurr, J.; Wilson, J.W.; Ott, C.M.; Tsaprailis, G.; Pierson, D.L.; Stefanyshyn-Piper, H.; et al. Transcriptional and proteomic responses of Pseudomonas aeruginosa PAO1 to spaceflight conditions involve Hfq regulation and reveal a role for oxygen. Appl. Environ. Microbiol. 2011, 77, 1221–1230. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, J.A.; Abogunde, O.; Thomas, K.; Lawal, A.; Nguyen, Y.U.; Sodipe, A.; Jejelowo, O. Spaceflight and modeled microgravity effects on microbial growth and virulence. Appl. Microbiol. Biotechnol. 2010, 85, 885–891. [Google Scholar] [CrossRef] [Green Version]
- Arunasri, K.; Adil, M.; Venu Charan, K.; Suvro, C.; Himabindu Reddy, S.; Shivaji, S. Effect of simulated microgravity on E. coli K12 MG1655 growth and gene expression. PLoS ONE 2013, 8, e57860. [Google Scholar] [CrossRef] [Green Version]
- Tucker, D.L.; Ott, C.M.; Huff, S.; Fofanov, Y.; Pierson, D.L.; Willson, R.C.; Fox, G.E. Characterization of Escherichia coli MG1655 grown in a low-shear modeled microgravity environment. BMC Microbiol. 2007, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Vukanti, R.; Mintz, E.; Leff, L. Changes in Gene Expression of E. coli under Conditions of Modeled Reduced Gravity. Microgravity Sci. Technol. 2008, 20, 41–57. [Google Scholar] [CrossRef]
- Wilson, J.W.; Ott, C.M.; Ramamurthy, R.; Porwollik, S.; McClelland, M.; Pierson, D.L.; Nickerson, C.A. Low-Shear modeled microgravity alters the Salmonella enterica serovar typhimurium stress response in an RpoS-independent manner. Appl. Environ. Microbiol. 2002, 68, 5408–5416. [Google Scholar] [CrossRef] [Green Version]
- Nickerson, C.A.; Ott, C.M.; Wilson, J.W.; Ramamurthy, R.; Pierson, D.L. Microbial responses to microgravity and other low-shear environments. Microbiol. Mol. Biol. Rev. 2004, 68, 345–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crabbe, A.; Pycke, B.; Van Houdt, R.; Monsieurs, P.; Nickerson, C.; Leys, N.; Cornelis, P. Response of Pseudomonas aeruginosa PAO1 to low shear modelled microgravity involves AlgU regulation. Environ. Microbiol. 2010, 12, 1545–1564. [Google Scholar] [PubMed]
- Nickerson, C.A.; Ott, C.M.; Wilson, J.W.; Ramamurthy, R.; LeBlanc, C.L.; Honer zu Bentrup, K.; Hammond, T.; Pierson, D.L. Low-shear modeled microgravity: A global environmental regulatory signal affecting bacterial gene expression, physiology, and pathogenesis. J. Microbiol. Methods 2003, 54, 1–11. [Google Scholar] [CrossRef]
- Guzy, C.; Paclik, D.; Schirbel, A.; Sonnenborn, U.; Wiedenmann, B.; Sturm, A. The probiotic Escherichia coli strain Nissle 1917 induces gammadelta T cell apoptosis via caspase- and FasL-dependent pathways. Int. Immunol. 2008, 20, 829–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnenborn, U. Escherichia coli strain Nissle 1917-from bench to bedside and back: History of a special Escherichia coli strain with probiotic properties. FEMS Microbiol. Lett. 2016, 363, fnw212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ukena, S.N.; Singh, A.; Dringenberg, U.; Engelhardt, R.; Seidler, U.; Hansen, W.; Bleich, A.; Bruder, D.; Franzke, A.; Rogler, G.; et al. Probiotic Escherichia coli Nissle 1917 inhibits leaky gut by enhancing mucosal integrity. PLoS ONE 2007, 2, e1308. [Google Scholar] [CrossRef] [PubMed]
- Wehkamp, J.; Harder, J.; Wehkamp, K.; Wehkamp-von Meissner, B.; Schlee, M.; Enders, C.; Sonnenborn, U.; Nuding, S.; Bengmark, S.; Fellermann, K.; et al. NF-kappaB- and AP-1-mediated induction of human beta defensin-2 in intestinal epithelial cells by Escherichia coli Nissle 1917: A novel effect of a probiotic bacterium. Infect. Immun. 2004, 72, 5750–5758. [Google Scholar] [CrossRef] [Green Version]
- Jacobi, C.A.; Malfertheiner, P. Escherichia coli Nissle 1917 (Mutaflor): New insights into an old probiotic bacterium. Dig. Dis. 2011, 29, 600–607. [Google Scholar] [CrossRef]
- Huijser, R. Desktop RPM: New small size microgravity simulator for the bioscience laboratory. Fokker Space 2000, 1. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Keseler, I.M.; Collado-Vides, J.; Gama-Castro, S.; Ingraham, J.; Paley, S.; Paulsen, I.T.; Peralta-Gil, M.; Karp, P.D. EcoCyc: A comprehensive database resource for Escherichia coli. Nucleic Acids Res. 2005, 33, D334–D337. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]
- Bak, G.; Lee, J.; Suk, S.; Kim, D.; Young Lee, J.; Kim, K.S.; Choi, B.S.; Lee, Y. Identification of novel sRNAs involved in biofilm formation, motility, and fimbriae formation in Escherichia coli. Sci. Rep. 2015, 5, 15287. [Google Scholar] [CrossRef] [Green Version]
- Van Houdt, R.; Michiels, C.W. Role of bacterial cell surface structures in Escherichia coli biofilm formation. Res. Microbiol. 2005, 156, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Khadke, S.K.; Lee, J.-H.; Woo, J.-T.; Lee, J. Inhibitory Effects of Honokiol and Magnolol on Biofilm Formation by Acinetobacter baumannii. Biotechnol. Bioproc. E 2019, 24, 359–365. [Google Scholar] [CrossRef]
- Zea, L.; Prasad, N.; Levy, S.E.; Stodieck, L.; Jones, A.; Shrestha, S.; Klaus, D. A Molecular Genetic Basis Explaining Altered Bacterial Behavior in Space. PLoS ONE 2016, 11, e0164359. [Google Scholar] [CrossRef]
- Benoit, M.R.; Klaus, D.M. Microgravity, bacteria, and the influence of motility. Adv. Space Res. 2007, 39, 1225–1232. [Google Scholar] [CrossRef]
- Kim, W.; Tengra, F.K.; Young, Z.; Shong, J.; Marchand, N.; Chan, H.K.; Pangule, R.C.; Parra, M.; Dordick, J.S.; Plawsky, J.L.; et al. Spaceflight promotes biofilm formation by Pseudomonas aeruginosa. PLoS ONE 2013, 8, e62437. [Google Scholar] [CrossRef] [Green Version]
- Lynch, S.V.; Mukundakrishnan, K.; Benoit, M.R.; Ayyaswamy, P.S.; Matin, A. Escherichia coli biofilms formed under low-shear modeled microgravity in a ground-based system. Appl. Environ. Microbiol. 2006, 72, 7701–7710. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yan, Y.; Rong, D.; Wang, J.; Wang, H.; Liu, Z.; Wang, J.; Yang, R.; Han, Y. Increased biofilm formation ability in Klebsiella pneumoniae after short-term exposure to a simulated microgravity environment. Microbiologyopen 2016, 5, 793–801. [Google Scholar] [CrossRef]
- Alexeeva, S.; Hellingwerf, K.J.; Teixeira de Mattos, M.J. Quantitative assessment of oxygen availability: Perceived aerobiosis and its effect on flux distribution in the respiratory chain of Escherichia coli. J. Bacteriol. 2002, 184, 1402–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorachek-Warren, M.K.; Carty, S.M.; Lin, S.; Cotter, R.J.; Raetz, C.R. An Escherichia coli mutant lacking the cold shock-induced palmitoleoyltransferase of lipid A biosynthesis: Absence of unsaturated acyl chains and antibiotic hypersensitivity at 12 degrees C. J. Biol. Chem. 2002, 277, 14186–14193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, M.L.; Chorell, E.; Taylor, J.D.; Åden, J.; Götheson, A.; Li, F.; Koch, M.; Sefer, L.; Matthews, S.J.; Wittung-Stafshede, P.; et al. The bacterial curli system possesses a potent and selective inhibitor of amyloid formation. Mol. Cell 2015, 57, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lasaro, M.A.; Salinger, N.; Zhang, J.; Wang, Y.; Zhong, Z.; Goulian, M.; Zhu, J. F1C fimbriae play an important role in biofilm formation and intestinal colonization by the Escherichia coli commensal strain Nissle 1917. Appl. Environ. Microbiol. 2009, 75, 246–251. [Google Scholar] [CrossRef] [Green Version]
- Nzakizwanayo, J.; Kumar, S.; Ogilvie, L.A.; Patel, B.A.; Dedi, C.; Macfarlane, W.M.; Jones, B.V. Disruption of Escherichia coli Nissle 1917 K5 Capsule Biosynthesis, through Loss of Distinct kfi genes, Modulates Interaction with Intestinal Epithelial Cells and Impact on Cell Health. PLoS ONE 2015, 10, e0120430. [Google Scholar] [CrossRef] [Green Version]
- Ott, C.M.; Chouker, A. Stress challenges and Immunity in Space: From Mechanisms to Monitoring and Preventive Strategies; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011; pp. 203–227. [Google Scholar]
- Kim, H.W.; Rhee, M.S. Influence of Low-Shear Modeled Microgravity on Heat Resistance, Membrane Fatty Acid Composition, and Heat Stress-Related Gene Expression in Escherichia coli O157:H7 ATCC 35150, ATCC 43889, ATCC 43890, and ATCC 43895. Appl. Environ. Microbiol. 2016, 82, 2893–2901. [Google Scholar] [CrossRef] [Green Version]
- Phadtare, S.; Alsina, J.; Inouye, M. Cold-shock response and cold-shock proteins. Curr. Opin. Microbiol. 1999, 2, 175–180. [Google Scholar] [CrossRef]
- Malki, A.; Le, H.T.; Milles, S.; Kern, R.; Caldas, T.; Abdallah, J.; Richarme, G. Solubilization of protein aggregates by the acid stress chaperones HdeA and HdeB. J. Biol. Chem. 2008, 283, 13679–13687. [Google Scholar] [CrossRef] [Green Version]
- Castanie-Cornet, M.P.; Cam, K.; Bastiat, B.; Cros, A.; Bordes, P.; Gutierrez, C. Acid stress response in Escherichia coli: Mechanism of regulation of gadA transcription by RcsB and GadE. Nucleic Acids Res. 2010, 38, 3546–3554. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Kolmakova-Partensky, L.; Miller, C. A bacterial arginine-agmatine exchange transporter involved in extreme acid resistance. J. Biol. Chem. 2007, 282, 176–182. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.W.; Matin, A.; Rhee, M.S. Microgravity Alters the Physiological Characteristics of Escherichia coli O157:H7 ATCC 35150, ATCC 43889, and ATCC 43895 under Different Nutrient Conditions. Appl. Environ. Microbiol. 2014, 80, 2270–2278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rensing, C.; Grass, G. Escherichia coli mechanisms of copper homeostasis in a changing environment. FEMS Microbiol. Rev. 2003, 27, 197–213. [Google Scholar] [CrossRef] [Green Version]
- King, A.; Selak, M.A.; Gottlieb, E. Succinate dehydrogenase and fumarate hydratase: Linking mitochondrial dysfunction and cancer. Oncogene 2006, 25, 4675–4682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauchene, N.A.; Mettert, E.L.; Moore, L.J.; Keles, S.; Willey, E.R.; Kiley, P.J. O2 availability impacts iron homeostasis in Escherichia coli. Proc. Natl. Acad. Sci. USA 2017, 114, 12261–12266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miethke, M. Molecular strategies of microbial iron assimilation: From high-affinity complexes to cofactor assembly systems. Metallomics 2013, 5, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.W.; Kim, D.; Szubin, R.; Palsson, B.O. Genome-wide Reconstruction of OxyR and SoxRS Transcriptional Regulatory Networks under Oxidative Stress in Escherichia coli K-12 MG1655. Cell Rep. 2015, 12, 1289–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Yu, Y.; Leary, J.A. Mechanism and kinetics of metalloenzyme phosphomannose isomerase: Measurement of dissociation constants and effect of zinc binding using ESI-FTICR mass spectrometry. Anal. Chem. 2005, 77, 5596–5603. [Google Scholar] [CrossRef]
- Wolf, G.; Schmidt, W. Zinc and glutamate dehydrogenase in putative glutamatergic brain structures. Acta Histochem. 1983, 72, 15–23. [Google Scholar] [CrossRef]
- Noll, M.; Petrukhin, K.; Lutsenko, S. Identification of a novel transcription regulator from Proteus mirabilis, PMTR, revealed a possible role of YJAI protein in balancing zinc in Escherichia coli. J. Biol. Chem. 1998, 273, 21393–21401. [Google Scholar] [CrossRef] [Green Version]
- Koh, E.-I.; Hung, C.S.; Henderson, J.P. The Yersiniabactin-Associated ATP Binding Cassette Proteins YbtP and YbtQ Enhance Escherichia coli Fitness during High-Titer Cystitis. Infect. Immun. 2016, 84, 1312–1319. [Google Scholar] [CrossRef] [Green Version]
- Behnsen, J.; Liu, J.; Valeri, M.; Hoover, E.; Tjokrosurjo, J.; Montaldo, N.P.; Treacy-Abarca, S.; Garibay, O.; Gilston, B.A.; Edwards, R.A. Probiotic Escherichia coli Nissle 1917 Uses Zinc Transporters and the Siderophore Yersiniabactin to Acquire Zinc in the Inflamed Gut and Outcompete Salmonella Typhimurium. FASEB J. 2017, 31, 622–626. [Google Scholar]
- Hasona, A.; Self, W.T.; Ray, R.M.; Shanmugam, K.T. Molybdate-dependent transcription of hyc and nar operons of Escherichia coli requires MoeA protein and ModE-molybdate. FEMS Microbiol. Lett. 1998, 169, 111–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thome, R.; Gust, A.; Toci, R.; Mendel, R.; Bittner, F.; Magalon, A.; Walburger, A. A sulfurtransferase is essential for activity of formate dehydrogenases in Escherichia coli. J. Biol. Chem. 2012, 287, 4671–4678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gisselberg, J.E.; Dellibovi-Ragheb, T.A.; Matthews, K.A.; Bosch, G.; Prigge, S.T. The suf iron-sulfur cluster synthesis pathway is required for apicoplast maintenance in malaria parasites. PLoS Pathog. 2013, 9, e1003655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, S.W.; Kim, D.; Latif, H.; O’Brien, E.J.; Szubin, R.; Palsson, B.O. Deciphering Fur transcriptional regulatory network highlights its complex role beyond iron metabolism in Escherichia coli. Nat. Commun. 2014, 5, 4910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pomposiello, P.J.; Demple, B. Global adjustment of microbial physiology during free radical stress. Adv. Microb. Physiol. 2002, 46, 319–341. [Google Scholar]
- Wendisch, V.F. Amino Acid Biosynthesis–Pathways, Regulation and Metabolic Engineering; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007; Volume 5, pp. 198–218. [Google Scholar]
- André, G.; Haudecoeur, E.; Monot, M.; Ohtani, K.; Shimizu, T.; Dupuy, B.; Martin-Verstraete, I. Global regulation of gene expression in response to cysteine availability in Clostridium perfringens. BMC Microbiol. 2010, 10, 204–208. [Google Scholar] [CrossRef] [Green Version]
- Fontecave, M.; Py, B.; Ollagnier de Choudens, S.; Barras, F. From Iron and Cysteine to Iron-Sulfur Clusters: The Biogenesis Protein Machineries. EcoSal Plus 2008, 3. [Google Scholar] [CrossRef]
- Kawano, Y.; Suzuki, K.; Ohtsu, I. Current understanding of sulfur assimilation metabolism to biosynthesize L-cysteine and recent progress of its fermentative overproduction in microorganisms. Appl. Microbiol. Biotechnol. 2018, 102, 8203–8211. [Google Scholar] [CrossRef]
- Ostrowski, J.; Kredich, N.M. In vitro interactions of CysB protein with the cysJIH promoter of Salmonella typhimurium: Inhibitory effects of sulfide. J. Bacteriol. 1990, 172, 779–785. [Google Scholar] [CrossRef] [Green Version]
- McPherson, A. Effects of a microgravity environment on the crystallization of biological macromolecules. Microgravity Sci. Technol. 1993, 6, 101–109. [Google Scholar] [PubMed]
- Shimizu, K. Regulation Systems of Bacteria such as Escherichia coli in Response to Nutrient Limitation and Environmental Stresses. Metabolites 2013, 4, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Schreiber, K.; Appel, J.; Makowka, A.; Fähnrich, B.; Roettger, M.; Hajirezaei, M.R.; Sönnichsen, F.D.; Schönheit, P.; Martin, W.F.; et al. The Entner-Doudoroff pathway is an overlooked glycolytic route in cyanobacteria and plants. Proc. Natl. Acad. Sci. USA 2016, 113, 5441–5446. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Mailloux, R.J.; Puiseux-Dao, S.; Appanna, V.D. Oxidative Stress Evokes a Metabolic Adaptation That Favors Increased NADPH Synthesis and Decreased NADH Production in Pseudomonas fluorescens. J. Bacteriol. 2007, 189, 6665–6675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasylenko, T.M.; Ahn, W.S.; Stephanopoulos, G. The oxidative pentose phosphate pathway is the primary source of NADPH for lipid overproduction from glucose in Yarrowia lipolytica. Metab. Eng. 2015, 30, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Voigt, B.; Hoi, L.T.; Jürgen, B.; Albrecht, D.; Ehrenreich, A.; Veith, B.; Evers, S.; Maurer, K.H.; Hecker, M.; Schweder, T. The glucose and nitrogen starvation response of Bacillus licheniformis. Proteomics 2007, 7, 413–423. [Google Scholar] [CrossRef]
- Bolton, J.L.; Dunlap, T. Formation and Biological Targets of Quinones: Cytotoxic versus Cytoprotective Effects. Chem. Res. Toxicol. 2017, 30, 13–37. [Google Scholar] [CrossRef]
- Zheng, L.; Cardaci, S.; Jerby, L.; MacKenzie, E.D.; Sciacovelli, M.; Johnson, T.I.; Gaude, E.; King, A.; Leach, J.D.G.; Edrada-Ebel, R.; et al. Fumarate induces redox-dependent senescence by modifying glutathione metabolism. Nat. Commun. 2015, 6, 6001. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.-F.; Zhang, Y.Y.; Di He, M.; Li, C.Y.; Zhou, C.X.; Hong, P.Z.; Qian, Z.-J. Antioxidant Peptide Purified from Enzymatic Hydrolysates of Isochrysis Zhanjiangensis and Its Protective Effect against Ethanol Induced Oxidative Stress of HepG2 Cells. Biotechnol. Bioproc. E 2019, 24, 308–317. [Google Scholar] [CrossRef]
- Li, S.J.; Cronan, J.E., Jr. The gene encoding the biotin carboxylase subunit of Escherichia coli acetyl-CoA carboxylase. J. Biol. Chem. 1992, 267, 855–863. [Google Scholar]
- Wu, Y.; Janetopoulos, C. Systematic analysis of γ-aminobutyric acid (GABA) metabolism and function in the social amoeba Dictyostelium discoideum. J. Biol. Chem. 2013, 288, 15280–15290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, P.; Chen, K.; Huang, X.; Wang, X.; Ren, Q. Production of γ-aminobutyric acid in Escherichia coli by engineering MSG pathway. Prep. Biochem. Biotechnol. 2018, 48, 906–913. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus Tag | Fold Change (Exp) | Fold Change (Sta) | Gene Function | Gene | Function Category |
|---|---|---|---|---|---|
| ECOLIN_01835 | −3.51 | −2.48 | hypothetical protein | ecpC | Biofilm formation |
| ECOLIN_01840 | −3.57 | −4.46 | hypothetical protein | ecpB | Biofilm formation |
| ECOLIN_01845 | −3.56 | −7.36 | fimbrial protein | ecpA | Biofilm formation |
| ECOLIN_01850 | −2.76 | −3.78 | LuxR family transcriptional regulator | ecpR | Biofilm formation |
| ECOLIN_19660 | 2.38 | −2.24 | cold-shock protein | cspA | Stress resistance |
| ECOLIN_19450 | −2.36 | −2.27 | glutamate decarboxylase | gadA | Stress resistance |
| ECOLIN_09330 | 2.27 | −2.30 | L-cystine transporter tcyP | tcyP | Metal ion utilization |
| ECOLIN_09975 | −2.82 | −5.68 | zinc ABC transporter substrate-binding protein | znuA | Metal ion utilization |
| ECOLIN_10835 | −2.43 | −15.62 | zinc/cadmium-binding protein | zinT | Metal ion utilization |
| ECOLIN_02515 | 2.50 | −2.17 | cytochrome o ubiquinol oxidase subunit II | cyoA | Uncategorized |
| ECOLIN_01860 | −3.45 | −11.11 | 50 S ribosomal protein L31 type B | - | Uncategorized |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yim, J.; Cho, S.W.; Kim, B.; Park, S.; Han, Y.H.; Seo, S.W. Transcriptional Profiling of the Probiotic Escherichia coli Nissle 1917 Strain under Simulated Microgravity. Int. J. Mol. Sci. 2020, 21, 2666. https://doi.org/10.3390/ijms21082666
Yim J, Cho SW, Kim B, Park S, Han YH, Seo SW. Transcriptional Profiling of the Probiotic Escherichia coli Nissle 1917 Strain under Simulated Microgravity. International Journal of Molecular Sciences. 2020; 21(8):2666. https://doi.org/10.3390/ijms21082666
Chicago/Turabian StyleYim, Jaewoo, Sung Won Cho, Beomhee Kim, Sungwoo Park, Yong Hee Han, and Sang Woo Seo. 2020. "Transcriptional Profiling of the Probiotic Escherichia coli Nissle 1917 Strain under Simulated Microgravity" International Journal of Molecular Sciences 21, no. 8: 2666. https://doi.org/10.3390/ijms21082666