Protein Arginine Methyltransferase 1 (PRMT1) Selective Inhibitor, TC-E 5003, Has Anti-Inflammatory Properties in TLR4 Signaling

 , and
, and 
Abstract
:1. Introduction
2. Results
2.1. The Effects of TC-E on NO Production
2.2. The Effects of TC-E on Pro-Inflammatory Gene Expression
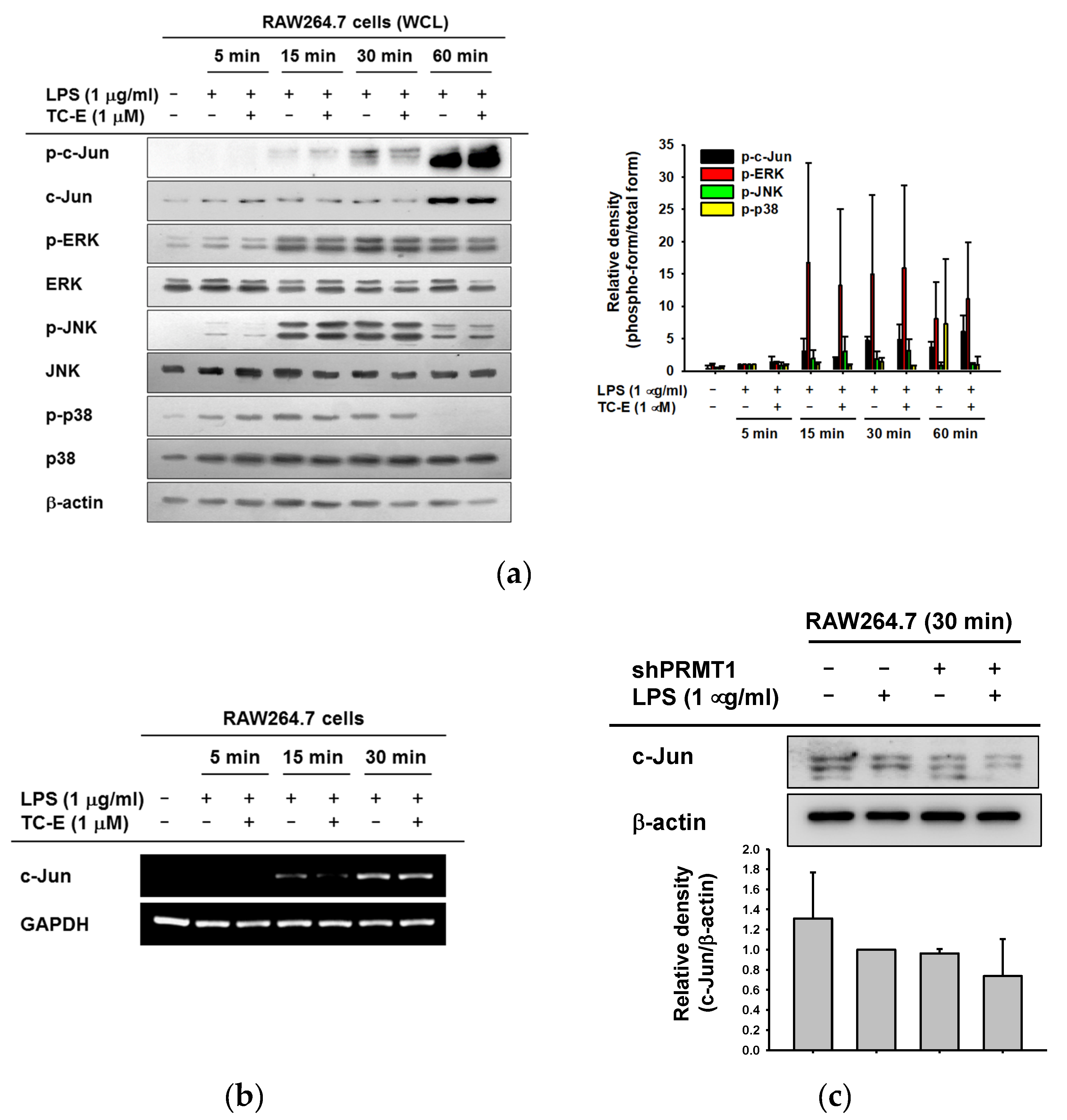
2.3. The Regulatory Effects of TC-E on the AP-1 Signaling Pathway
2.4. The Regulatory Effects of TC-E on the NF-κB Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. NO Production and Griess Assay
4.4. Cell Viability Assay
4.5. Preparation of mRNA and Semiquantitative PCR
4.6. Plasmid Transfection
4.7. Preparation of Nuclear Fraction
4.8. Preparation of Whole Cell Lysates
4.9. Western Blotting Analysis
4.10. Lentivirus-Mediated Knockdown (shRNA)
4.11. Statical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADMA | Asymmetric dimethylarginine |
| AdoMet | S-Adenosylmethionine |
| AP-1 | Activator protein 1 |
| CETSA | Cellular thermal shift assay |
| CSK | C-terminal Src kinase |
| DAMPs | Damage-associated molecular patterns |
| EDTA | Ethylenediaminetetraacetic acid |
| EGTA | Ethyleneglycotetraacetic acid |
| ERK | Extracellular signal-regulated kinase |
| HIF | Hypoxia-inducible transcription factor |
| ICAM | Intracellular adhesion molecule |
| IL | Interleukin |
| IRF | Interferon regulatory factor |
| JNK | c-Jun N-terminal kinase |
| MAPKs | Mitogen-activated protein kinases |
| MIP | Macrophage inflammatory protein |
| MMA | Monomethylated arginine |
| NF-kB | Nuclear factor kB |
| NO | Nitric oxide |
| NOS | Nitric oxide synthase |
| PAMPs | Pathogen-associated molecular patterns |
| PKC | Protein kinase C |
| PPAR | Peroxisome proliferator-activator receptor |
| PRMT | Protein arginine methyltransferase |
| PTP | Protein tyrosine phosphatase |
| RTK | Receptor tyrosine kinase |
| SDMA | Symmetric dimethylarginine |
| Sp1 | Specificity protein 1 |
| TNF | Tumor necrosis factor |
| TRAF | TNF receptor associated factor |
References
- Kim, H.G.; Choi, S.; Lee, J.; Hong, Y.H.; Jeong, D.; Yoon, K.; Yoon, D.H.; Sung, G.-H.; Lee, S.; Hong, S. Src is a prime target inhibited by celtis choseniana methanol extract in its anti-inflammatory action. Evid. Based Complementary Altern. Med. 2018, 2018, 3909038. [Google Scholar] [CrossRef]
- Nowarski, R.; Gagliani, N.; Huber, S.; Flavell, R.A. Innate immune cells in inflammation and cancer. Cancer Immunol. Res. 2013, 1, 77–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Al-Kuhlani, M.; Johnston, S.C.; Ojcius, D.M.; Chou, J.; Dean, D. Transcription factor complex AP-1 mediates inflammation initiated by C hlamydia pneumoniae infection. Cell. Microbiol. 2013, 15, 779–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.Y.; Kim, J.; Kim, E.; Kim, S.H.; Seo, D.B.; Kim, J.H.; Shin, S.S.; Cho, J.Y. AKT-targeted anti-inflammatory activity of Panax ginseng calyx ethanolic extract. J. Ginseng Res. 2018, 42, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Yi, Y.S.; Son, Y.J.; Han, S.Y.; Kim, D.H.; Nam, G.; Hossain, M.A.; Kim, J.H.; Park, J.; Cho, J.Y. BIOGF1K, a compound K-rich fraction of ginseng, plays an antiinflammatory role by targeting an activator protein-1 signaling pathway in RAW264.7 macrophage-like cells. J. Ginseng Res. 2018, 42, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Jeong, D.; Nam, G.; Yi, Y.-S.; Yoon, D.H.; Kim, T.W.; Park, Y.C.; Hwang, H.; Rhee, M.H.; Hong, S. AP-1 pathway-targeted inhibition of inflammatory responses in LPS-treated macrophages and EtOH/HCl-treated stomach by Archidendron clypearia methanol extract. J. Ethnopharmacol. 2013, 146, 637–644. [Google Scholar] [CrossRef]
- Lawrence, T. The nuclear factor NF-κB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Straub, R.H.; Schradin, C. Chronic inflammatory systemic diseases: An evolutionary trade-off between acutely beneficial but chronically harmful programs. Evol. Med. Public Health 2016, 2016, 37–51. [Google Scholar] [CrossRef] [Green Version]
- Ahuja, A.; Kim, J.H.; Kim, J.H.; Yi, Y.S.; Cho, J.Y. Functional role of ginseng-derived compounds in cancer. J. Ginseng Res. 2018, 42, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, P.; Tian, E.; Zhang, H. Arginine methylation modulates autophagic degradation of PGL granules in C. elegans. Mol. Cell 2013, 52, 421–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedford, M.T.; Clarke, S.G. Protein arginine methylation in mammals: Who, what, and why. Mol. Cell 2009, 33, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bachand, F. Protein arginine methyltransferases: From unicellular eukaryotes to humans. Eukaryot. Cell 2007, 6, 889–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, R.S.; Richard, S. Arginine methylation: The coming of age. Mol. Cell 2017, 65, 8–24. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Yoo, B.C.; Yang, W.S.; Kim, E.; Hong, S.; Cho, J.Y. The role of protein arginine methyltransferases in inflammatory responses. Mediat. Inflamm. 2016, 2016, 4028353. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Mundade, R.; Lange, K.; Lu, T. Protein arginine methylation of non-histone proteins and its role in diseases. Cell Cycle 2014, 13, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-I.; Park, M.-J.; Choi, J.-H.; Kim, I.-S.; Han, H.-J.; Yoon, K.-C.; Park, S.-W.; Lee, M.-Y.; Oh, K.-S.; Park, S.-H. PRMT1 and PRMT4 regulate oxidative stress-induced retinal pigment epithelial cell damage in SIRT1-dependent and SIRT1-independent manners. Oxidative Med. Cell. Longev. 2015, 2015, 617919. [Google Scholar] [CrossRef] [Green Version]
- Hata, K.; Yanase, N.; Sudo, K.; Kiyonari, H.; Mukumoto, Y.; Mizuguchi, J.; Yokosuka, T. Differential regulation of T-cell dependent and T-cell independent antibody responses through arginine methyltransferase PRMT 1 in vivo. FEBS Lett. 2016, 590, 1200–1210. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Sun, Q.; Bao, R.; Roth, M.; Zhong, B.; Lan, X.; Tian, J.; He, Q.; Li, D.; Sun, J. Specific regulation of PRMT1 expression by PIAS1 and RKIP in BEAS-2B epithelia cells and HFL-1 fibroblasts in lung inflammation. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Brooks, W.H.; Le Dantec, C.; Pers, J.-O.; Youinou, P.; Renaudineau, Y. Epigenetics and autoimmunity. J. Autoimmun. 2010, 34, J207–J219. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, K.; Clifford, R.; Patel, J.; Shaw, D.; Dowden, J.; Knox, A. The role of protein arginine N-Methyltransferase 1 (PRMT1) in inflammatory gene expression of airway smooth muscle cells in asthma, D58. In Airways Hyperresponsiveness: Novel Mechanisms And Targets; American Thoracic Society: New York, NY, USA, 2018; p. A7264. [Google Scholar]
- Liu, H.; Jiang, J.; Zhao, L. Protein arginine methyltransferase-1 deficiency restrains depression-like behavior of mice by inhibiting inflammation and oxidative stress via Nrf-2. Biochem. Biophys. Res. Commun. 2019, 518, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Li, J.; Li, P.; Ye, Q.; Xu, H.; Wu, X.; Xu, Y. Protein arginine methyltransferase 1 (PRMT1) represses MHC II transcription in macrophages by methylating CIITA. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tikhanovich, I.; Zhao, J.; Olson, J.; Adams, A.; Taylor, R.; Bridges, B.; Marshall, L.; Roberts, B.; Weinman, S.A. Protein arginine methyltransferase 1 modulates innate immune responses through regulation of peroxisome proliferator-activated receptor γ-dependent macrophage differentiation. J. Biol. Chem. 2017, 292, 6882–6894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Qian, K.; Yan, C.; He, M.; Jassim, B.A.; Ivanov, I.; Zheng, Y.G. Discovery of decamidine as a new and potent PRMT1 inhibitor. MedChemComm 2017, 8, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Ouyang, Y.; Ma, H.; Cong, H.; Zhuang, C.; Lok, W.-T.; Wang, Z.; Zhu, X.; Sun, Y.; Hong, W. Design and synthesis of novel PRMT1 inhibitors and investigation of their binding preferences using molecular modelling. Bioorganic Med. Chem. Lett. 2017, 27, 4635–4642. [Google Scholar] [CrossRef] [PubMed]
- Bissinger, E.-M.; Heinke, R.; Spannhoff, A.; Eberlin, A.; Metzger, E.; Cura, V.; Hassenboehler, P.; Cavarelli, J.; Schüle, R.; Bedford, M.T. Acyl derivatives of p-aminosulfonamides and dapsone as new inhibitors of the arginine methyltransferase hPRMT1. Bioorganic Med. Chem. 2011, 19, 3717–3731. [Google Scholar] [CrossRef]
- Sun, Q.; Liu, L.; Mandal, J.; Molino, A.; Stolz, D.; Tamm, M.; Lu, S.; Roth, M. PDGF-BB induces PRMT1 expression through ERK1/2 dependent STAT1 activation and regulates remodeling in primary human lung fibroblasts. Cell. Signal. 2016, 28, 307–315. [Google Scholar] [CrossRef]
- Xia, L.; Zhang, H.-X.; Xing, M.-L.; Xu, Y.-B.; Li, P.; Huang, L.-K.; Bai, J.; Tian, Z.; Zhao, Z.-D. Knockdown of prmt1 suppresses il-1β-induced cartilage degradation and inflammatory responses in human chondrocytes through gli1-mediated hedgehog signaling pathway. Mol. Cell. Biochem. 2018, 438, 17–24. [Google Scholar] [CrossRef]
- Intayoung, P.; Limtrakul, P.; Yodkeeree, S. Antiinflammatory activities of crebanine by inhibition of NF-κB and AP-1 activation through suppressing MAPKs and Akt signaling in LPS-induced RAW264. 7 macrophages. Biol. Pharm. Bull. 2016, 39, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Hossen, M.J.; Hong, Y.D.; Baek, K.S.; Yoo, S.; Hong, Y.H.; Kim, J.H.; Lee, J.O.; Kim, D.; Park, J.; Cho, J.Y. In vitro antioxidative and anti-inflammatory effects of the compound K-rich fraction BIOGF1K, prepared from Panax ginseng. J. Ginseng Res. 2017, 41, 43–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karin, M. The regulation of AP-1 activity by mitogen-activated protein kinases. J. Biol. Chem. 1995, 270, 16483–16486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, C.; Frazier-Jessen, M.; Rawat, R.; Nordan, R.; Brown, R. Evaluation of methods for transient transfection of a murine macrophage cell line, RAW 264.7. Biotechniques 1999, 27, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Cheung, S.T.; Shakibakho, S.; So, E.Y.; Mui, A.L. Transfecting RAW264. 7 cells with a luciferase reporter gene. JoVE 2015, e52807. [Google Scholar] [CrossRef] [Green Version]
- Yao, Q.; Liu, J.; Zhang, Z.; Li, F.; Zhang, C.; Lai, B.; Xiao, L.; Wang, N. Peroxisome proliferator-activated receptor γ (PPARγ) induces the gene expression of integrin α(V)β(5) to promote macrophage M2 polarization. J. Biol. Chem. 2018, 293, 16572–16582. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; O’Neil, M.; Vittal, A.; Weinman, S.A.; Tikhanovich, I. PRMT1-dependent macrophage IL-6 production is required for alcohol-induced HCC progression. Gene Expr. 2019, 19, 137–150. [Google Scholar] [CrossRef]
- Lafleur, V.N.; Richard, S.; Richard, D.E. Transcriptional repression of hypoxia-inducible factor-1 (HIF-1) by the protein arginine methyltransferase PRMT1. Mol. Biol. Cell 2014, 25, 925–935. [Google Scholar] [CrossRef]
- Davies, C.C.; Chakraborty, A.; Diefenbacher, M.E.; Skehel, M.; Behrens, A. Arginine methylation of the c-Jun coactivator RACO-1 is required for c-Jun/AP-1 activation. EMBO J. 2013, 32, 1556–1567. [Google Scholar] [CrossRef]
- Byeon, S.E.; Yi, Y.-S.; Oh, J.; Yoo, B.C.; Hong, S.; Cho, J.Y. The role of Src kinase in macrophage-mediated inflammatory responses. Mediat. Inflamm. 2012, 2012. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.S.; Yoo, B.C.; Yang, W.S.; Han, S.Y.; Jeong, D.; Song, J.M.; Kim, K.H.; Aravinthan, A.; Kim, J.H.; Kim, J.-H. Src is the primary target of aripiprazole, an atypical antipsychotic drug, in its anti-tumor action. Oncotarget 2018, 9, 5979. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.M.; Resnick, R.J.; Shalloway, D. A phosphotyrosine displacement mechanism for activation of Src by PTPα. EMBO J. 2000, 19, 964–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roskoski, R., Jr. Src kinase regulation by phosphorylation and dephosphorylation. Biochem. Biophys. Res. Commun. 2005, 331, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Roux, B. Locking the active conformation of c-Src kinase through the phosphorylation of the activation loop. J. Mol. Biol. 2014, 426, 423–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, S.J.; Parsons, J.T. Src family kinases, key regulators of signal transduction. Oncogene 2004, 23, 7906–7909. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Gong, P.; Hyun, S.W.; Wang, K.Z.; Cates, E.A.; Perkins, D.; Bannerman, D.D.; Puché, A.C.; Toshchakov, V.Y.; Fang, S. TRAF6 couples TLR4 signaling to Src family kinase activation and opening of the paracellular pathway in human lung microvascular endothelia. J. Biol. Chem. 2012. [Google Scholar] [CrossRef] [Green Version]
- Park, G.; Kim, E.; Son, Y.-J.; Yoon, D.H.; Sung, G.-H.; Aravinthan, A.; Park, Y.C.; Kim, J.-H.; Cho, J.Y. Anti-inflammatory effect of torilidis fructus ethanol extract through inhibition of Src. Pharm. Biol. 2017, 55, 2074–2082. [Google Scholar] [CrossRef]
- Sun, Q.; Liu, L.; Roth, M.; Tian, J.; He, Q.; Zhong, B.; Bao, R.; Lan, X.; Jiang, C.; Sun, J. PRMT1 upregulated by epithelial proinflammatory cytokines participates in COX2 expression in fibroblasts and chronic antigen-induced pulmonary inflammation. J. Immunol. 2015, 195, 298–306. [Google Scholar] [CrossRef]
- Li, T.; Kong, A.-N.T.; Ma, Z.; Liu, H.; Liu, P.; Xiao, Y.; Jiang, X.; Wang, L. Protein arginine methyltransferase 1 may be involved in pregnane x receptor-activated overexpression of multidrug resistance 1 gene during acquired multidrug resistant. Oncotarget 2016, 7, 20236. [Google Scholar] [CrossRef] [Green Version]
- Boykin, D.W. Antimicrobial activity of the DNA minor groove binders furamidine and analogs. J. Braz. Chem. Soc. 2002, 13, 763–771. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Qian, K.; Ho, M.-C.; Zheng, Y.G. Small molecule inhibitors of protein arginine methyltransferases. Expert Opin. Investig. Drugs 2016, 25, 335–358. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Liu, S.-M.; Sun, C.-W.; Bedford, M.T.; Zhao, X. Protein Arginine Methyltransferase 1 (PRMT1) Dampens Self-Renewal and Promotes Differentiation of Hematopoietic Stem Cells in Mouse Models; American Society of Hematology: Washington, DC, USA, 2019. [Google Scholar]
- Bayen, S.; Saini, S.; Gaur, P.; Duraisamy, A.J.; Sharma, A.K.; Pal, K.; Vats, P.; Singh, S.B. PRMT1 promotes hyperglycemia in a FoxO1-dependent manner, affecting glucose metabolism, during hypobaric hypoxia exposure, in rat model. Endocrine 2018, 59, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.Y.; Ng, S.Y.; Toepp, S.L.; Ljubicic, V. Protein arginine methyltransferase expression and activity during myogenesis. Biosci. Rep. 2018, 38, BSR20171533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, H.-B.; Sui, G.-G.; Lu, X.-Y.; Sun, Z.-L. Elevated levels of ADMA are associated with lower DDAH2 and higher PRMT1 in LPS-induced endometritis rats. Inflammation 2018, 41, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.Y.; Hong, J.T.; Han, S.B.; Park, Y.H.; Son, D.J. Effect of Ixeris dentata Nakai extract on nitric oxide production and prostaglandin E2 generation in LPS-stimulated RAW264. 7 Cells. Immune Netw. 2015, 15, 325–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-O.; Yang, W.S.; Park, J.G.; Jeong, D.; Kim, H.G.; Yoon, K.D.; Aravinthan, A.; Kim, J.-H.; Kim, E.; Cho, J.Y. Src and Syk contribute to the anti-inflammatory activities of Achyranthes aspera ethanolic extract. J. Ethnopharmacol. 2017, 206, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Byeon, S.E.; Chung, J.Y.; Lee, Y.G.; Kim, B.H.; Kim, K.H.; Cho, J.Y. In vitro and in vivo anti-inflammatory effects of taheebo, a water extract from the inner bark of Tabebuia avellanedae. J. Ethnopharmacol. 2008, 119, 145–152. [Google Scholar] [CrossRef]
- Choi, E.; Kim, E.; Kim, J.H.; Yoon, K.; Kim, S.; Lee, J.; Cho, J.Y. AKT1-targeted proapoptotic activity of compound K in human breast cancer cells. J. Ginseng Res. 2019, 43, 692–698. [Google Scholar] [CrossRef]
- Baek, K.-S.; Yi, Y.-S.; Son, Y.-J.; Yoo, S.; Sung, N.Y.; Kim, Y.; Hong, S.; Aravinthan, A.; Kim, J.-H.; Cho, J.Y. In vitro and in vivo anti-inflammatory activities of Korean Red Ginseng-derived components. J. Ginseng Res. 2016, 40, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Sung, N.Y.; Kim, M.-Y.; Cho, J.Y. Scutellarein reduces inflammatory responses by inhibiting Src kinase activity. Korean J. Physiol. Pharmacol. 2015, 19, 441–449. [Google Scholar] [CrossRef] [Green Version]
- Burnette, W.N. “Western blotting”: Electrophoretic transfer of proteins from sodium dodecyl sulfate-polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein A. Anal. Biochem. 1981, 112, 195–203. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences | |
|---|---|---|
| TNF-α | F | 5’-TTGACCTCAGCGCTGAGTTG-3’ |
| R | 5’-CCTGTAGCCCACGTCGTAGC-3’ | |
| IL-1β | F | 5’-CAGGATGAGGACATGAGCACC-3’ |
| R | 5’-CTCTGCAGACTCAAACTCCAC-3’ | |
| IL-6 | F | 5’-GTACTCCAGAAGACCAGAGG-3’ |
| R | 5’-TGCTGGTGACAACCACGGCC-3’ | |
| iNOS | F | 5‘-CCCTTCCGAAGTTTCTGGCAGCAGC-3` |
| R | 5‘-GGCTGTCAGAGCCTCGTGGCTTTGG-3` | |
| COX-2 | F | 5′-CACTACATCCTGACCCACTT-3′ |
| R | 5′-ATGCTCCTGCTTGAGTATGT-3′ | |
| c-Jun | F | 5′- ACGACCTTCTACGACGATGC-3′ |
| R | 5′- CCAGGTTCAAGGTCATGCTC-3′ | |
| GAPDH | F | 5’-CACTCACGGCAAATTCAACGGCAC-3’ |
| R | 5’-GACTCCACGACATACTCAGCAC-3’ |
| Gene | Sequences |
|---|---|
| shScrambled | TCCTAAGGTTAAGTCGCCCTCG |
| shPRMT1 | CATGATGCAGTTCGCGGCCTCGG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.; Jang, J.; Park, J.G.; Kim, K.-H.; Yoon, K.; Yoo, B.C.; Cho, J.Y. Protein Arginine Methyltransferase 1 (PRMT1) Selective Inhibitor, TC-E 5003, Has Anti-Inflammatory Properties in TLR4 Signaling. Int. J. Mol. Sci. 2020, 21, 3058. https://doi.org/10.3390/ijms21093058
Kim E, Jang J, Park JG, Kim K-H, Yoon K, Yoo BC, Cho JY. Protein Arginine Methyltransferase 1 (PRMT1) Selective Inhibitor, TC-E 5003, Has Anti-Inflammatory Properties in TLR4 Signaling. International Journal of Molecular Sciences. 2020; 21(9):3058. https://doi.org/10.3390/ijms21093058
Chicago/Turabian StyleKim, Eunji, Jiwon Jang, Jae Gwang Park, Kyung-Hee Kim, Keejung Yoon, Byong Chul Yoo, and Jae Youl Cho. 2020. "Protein Arginine Methyltransferase 1 (PRMT1) Selective Inhibitor, TC-E 5003, Has Anti-Inflammatory Properties in TLR4 Signaling" International Journal of Molecular Sciences 21, no. 9: 3058. https://doi.org/10.3390/ijms21093058