Drought Stress Tolerance and Photosynthetic Activity of Alloplasmic Lines T. dicoccum x T. aestivum

 ,
, 

Abstract
1. Introduction
2. Results
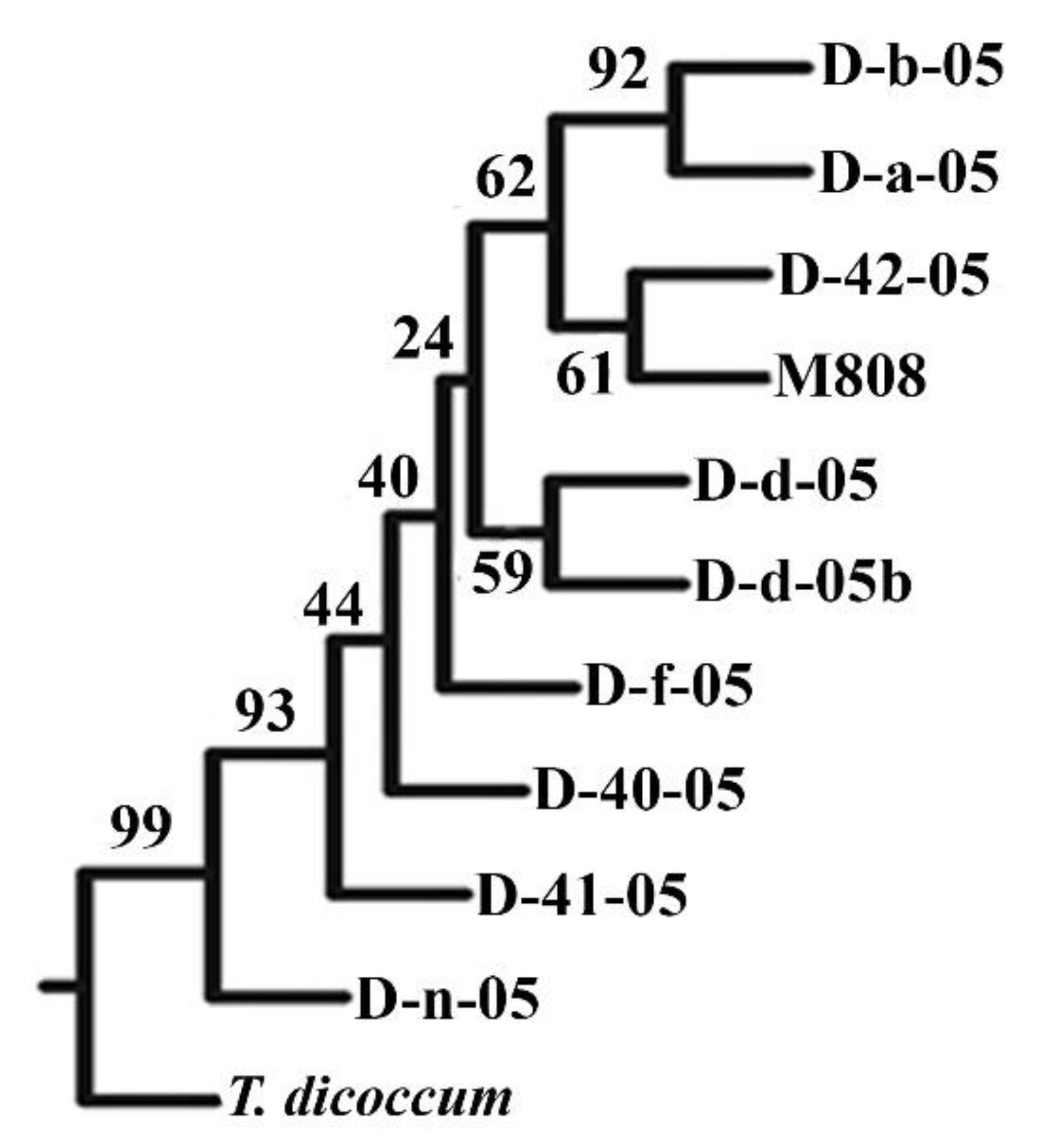
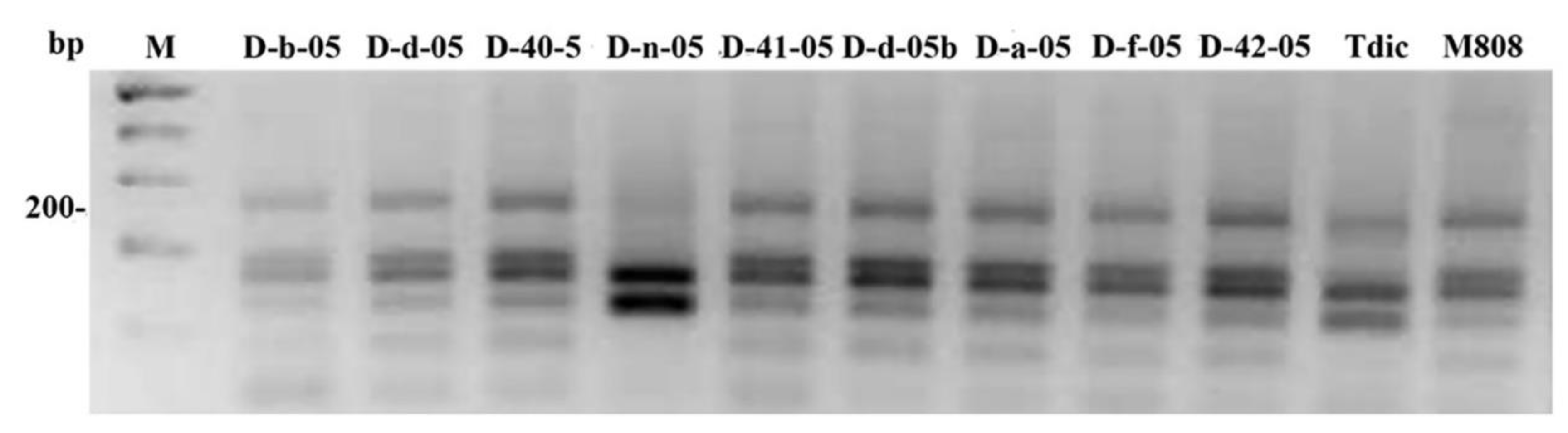
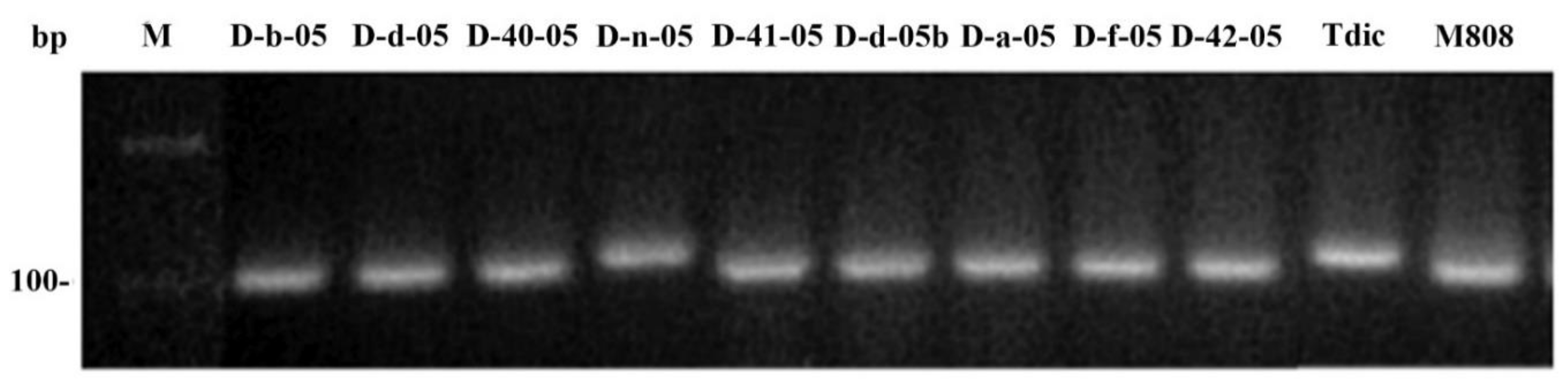
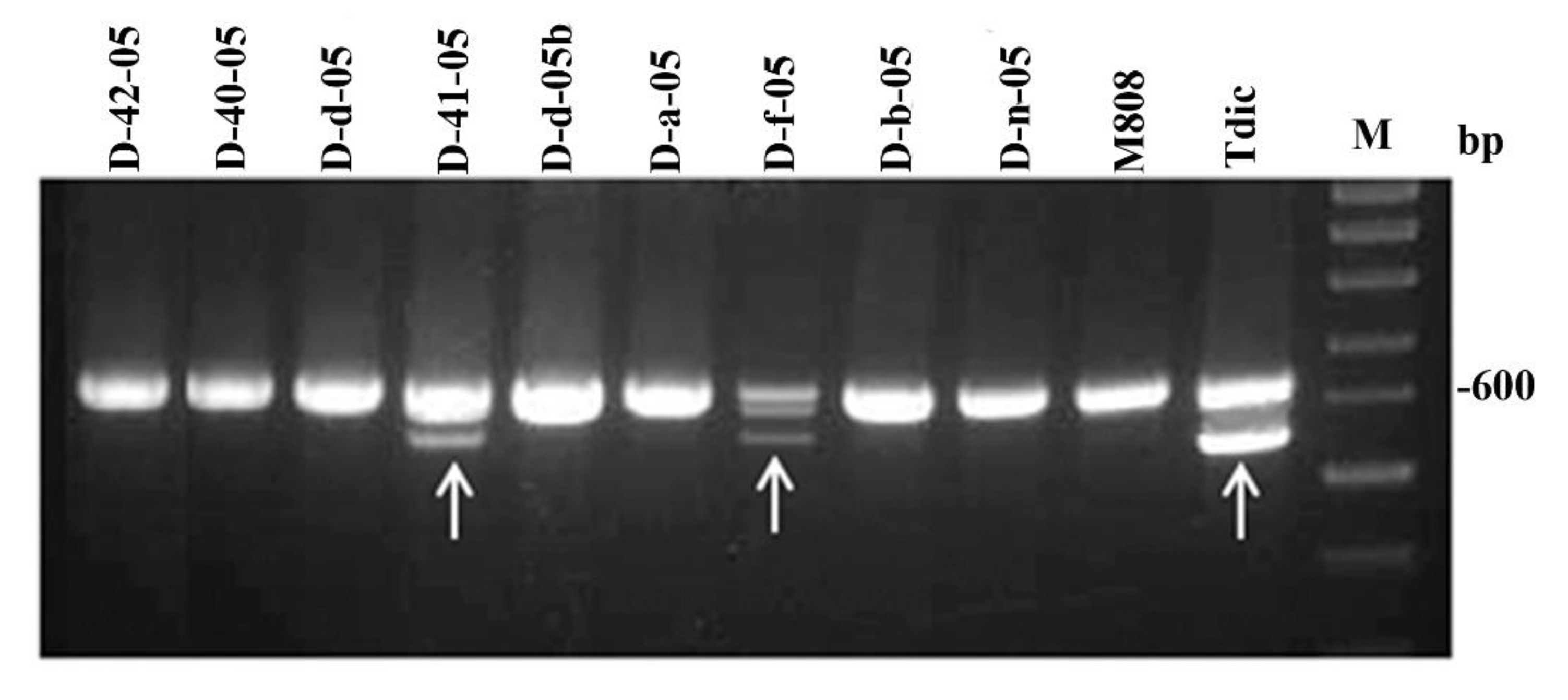
2.1. Genotyping of Allolines by Nuclear and Mitochondrial Markers
2.2. Study of Drought Tolerance and Photosynthetic Activity of Allolines
2.2.1. Growth of Seedlings
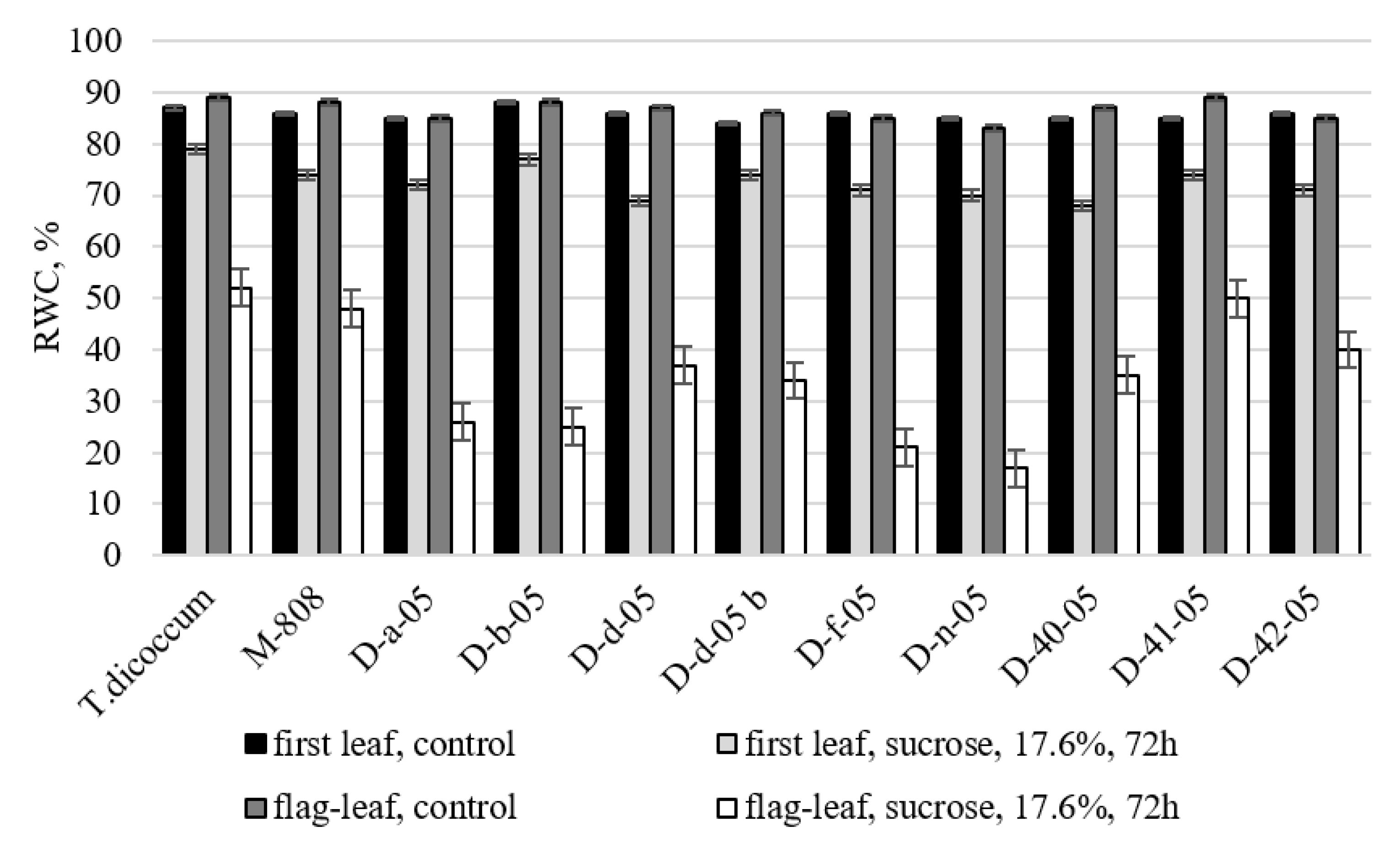
2.2.2. Relative Water Content in the First and Flag Leaves
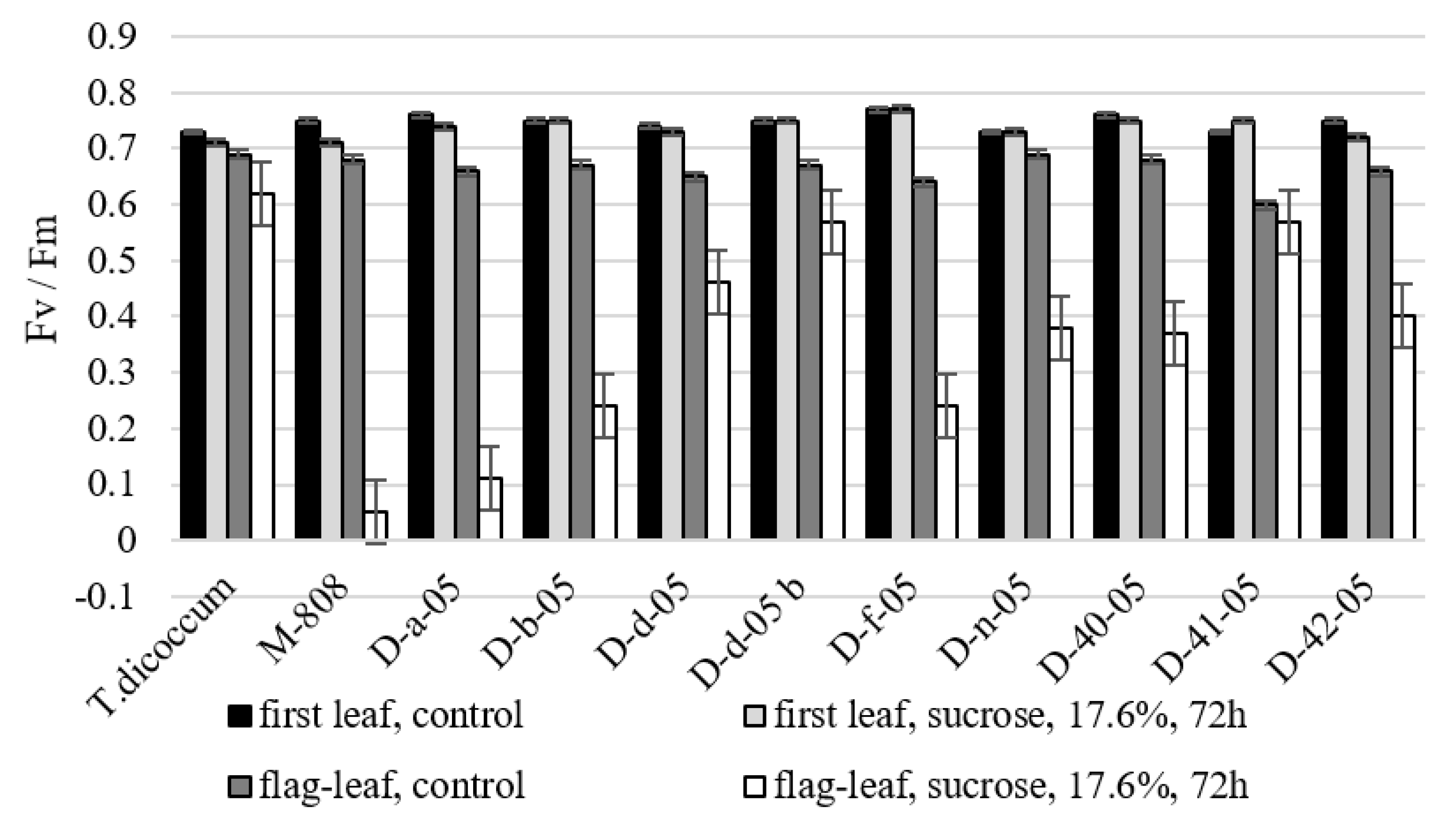
2.2.3. Quantum Efficiency of Photochemical Energy Scattering
2.2.4. Electron Transport Rate Through PSII
2.2.5. Changing of the Quantum Yield of Energy Dissipated in PSII
2.3. Identification of Gene Regulating Drought Tolerance in Alloplasmic Lines
3. Discussion
4. Materials and Methods
4.1. DNA Isolation and Analysis of SSR-Markers
4.2. Mitochondrial Genome Analysis
4.3. Analysis of the Drought Tolerance Regulator Gene Dreb-1
4.4. Analysis of Physiological Parameters of Drought Tolerance
4.5. Analysis of Photosynthetic Parameters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate Trends and Global Crop Production Since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, M.; Soolanayakanahally, R.; Ogawa, S.; Uga, Y.; Selvaraj, M.G.; Kagale, S. Drought Response in Wheat: Key Genes and Regulatory Mechanisms Controlling Root System Architecture and Transpiration Efficiency. Front. Chem. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Website ESS. Statistical Yearbooks−FAO World Food and Agriculture Statistical Yearbook. Available online: http://www.fao.org/economic/ess/ess-publications/ess-yearbook/ru/#.XrWdRGgzY2w (accessed on 20 May 2019).
- Schafleitner, R.; Rosales, R.O.G.; Gaudin, A.; Aliaga, C.A.A.; Martinez, G.N.; Marca, L.R.T.; Bolivar, L.A.; Delgado, F.M.; Simon, R.; Bonierbale, M. Capturing candidate drought tolerance traits in two native Andean potato clones by transcription profiling of field grown plants under water stress. Plant Physiol. Biochem. 2007, 45, 673–690. [Google Scholar] [CrossRef] [PubMed]
- Arriagada, O.; Mora, F.; Quitral, Y.; Del Pozo, A. Identification of QTL underlying agronomic, morphological and physiological traits in barley under rainfed conditions using SNP markers. Acta Sci. Agron. 2017, 39, 321–329. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Manes, Y.; Gomez, H.F.; Puhl, L.; Reynolds, M.; Braun, H.J.; Trethowan, R. Genetic yield gains of the CIMMYT international semi-arid wheat yield trials from 1994 to 2010. Crop Sci. 2012, 52, 1543–1552. [Google Scholar] [CrossRef]
- Ashraf, M. Exploitation of genetic variation for improvement of salt tolerance in spring wheat. In Prospects for Saline Agriculture; Ahmad, R., Malik, K.A., Eds.; Kluwer Academic Publishers: Dordrecht, Netherlands, 2002; pp. 113–121. [Google Scholar]
- Colmer, T.D.; Flowers, T.J.; Munns, R. Use of wild relatives to improve salt tolerance in wheat. J. Exp. Bot. 2006, 57, 1059–1078. [Google Scholar] [CrossRef]
- Shymkevich, A.M.; Lukhanina, N.V.; Golovenko, I.M.; Davydenko, O.G. Analysis of splitting frequencies by morphological and SSR6 locks in hybrid combinations of substituted barley lines. Russ. J. Genet. 2007, 1, 209–216. [Google Scholar]
- Orlov, P.A. The Interaction of Nuclear and Cytoplasmic Genes in the Determination of Plant Development; Unipak: Minsk, Belarus, 2001. [Google Scholar]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Annal. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Kholodova, V.P.; Bormotova, T.S.; Semenov, O.G.; Dmitrieva, G.A.; Kuznetsov, V.V. Physiological mechanisms of adaptation of alloplasmic wheat hybrids to soil drought. Russ. J. Plant Physiol. 2007, 54, 480–486. [Google Scholar] [CrossRef]
- Hodaei, M.; Rahimmalek, M.; Arzani, A. Plasmon analysis in wheat alloplasmic lines using morphological and chloroplast microsatellite markers. Plant Syst. Evol. 2014, 300, 1137–1145. [Google Scholar] [CrossRef]
- Shymkevich, A.M.; Makarov, V.N.; Golovenko, I.M.; Davydenko, O.G. Functional state of photosynthetic apparatus in alloplasmic lines of barley. Ecolog. Genet. 2006, 2, 3742. [Google Scholar]
- Akhtar, M.; Jaiswal, A.; Taj, G.; Jaiswal, J.P.; Qureshi, M.I.; Singh, N.K. DREB1/CBF transcription factors: Their structure, function and role in abiotic stress tolerance in plants. J. Genet. 2012, 91, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Terletskaya, N.V.; Kurmanbayeva, M.S. Change of leaf anatomical parameters of different species of wheat seedlings under conditions of drought and salt stress. Pac. J. Bot. 2017, 49, 857–865. [Google Scholar] [CrossRef]
- Terletskaya, N.; Zobova, N.; Stupko, V.; Shuyskaya, E. Growth and photosynthetic reactions of different species of wheat seedlings under drought and saltstress. Periodicum. Biologorum. 2017, 119, 37–45. [Google Scholar] [CrossRef]
- El-Shehawi, A.M.; Fahmi, A.I.; Sayed, S.M.; Elseehy, M.M. Genetic Fingerprinting of Wheat and Its Progenitors by Mitochondrial Gene orf256. Biomolecules 2012, 2, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Noyszewski, A.K. Mitochondrial Sequence Diversity among Alloplasmic and Euplasmic Triticum Species. Ph.D. Thesis, North Dakota State University, Fargo, North Dakota, 2013. [Google Scholar]
- Wei, H.; Zhou, B.; Zhang, F.; Tu, Y.; Hu, Y.; Zhang, B.; Zhai, Q. Profiling and identification of small rDNA-derived RNAs and their potential biological functions. PLoS ONE 2013, 8, e56842. [Google Scholar] [CrossRef]
- Veselov, D.S. Growth by Extension and Water Metabolism in Water Deficit Conditions. Ph.D. Thesis, SEI Bashkir. State University, Ufa, Russia, 2009. [Google Scholar]
- Aroca, R.; Porcel, R.; Ruis-Lozano, J.M. Regulation of Root Water Uptake under Drought Stress Conditions. J. Exp. Bot. 2011, 63, 43–57. [Google Scholar] [CrossRef]
- Lugojan, C.; Ciulca, S. Evaluation of relative water content in winter wheat. J. Hortic. Fores. Biotechnol. 2011, 15, 173–177. [Google Scholar]
- Liu, Y.; Fiskum, G.; Schubert, D. Generation of reactive oxygen species by the mitochondrial electron transport chain. J. Neurochem. 2002, 80, 780–787. [Google Scholar] [CrossRef]
- Ramos, M.L.G.; Parsons, R.; Sprent, J.I.; Games, E.K. Effect of water stress on nitrogen fixation and nodule structure of common bean. Pesq. Agropec. Brasil. 2003, 38, 339–347. [Google Scholar] [CrossRef]
- Efeoglu, B.; Ekmekci, Y.; Cicek, N. Physiological responses of three maize cultivars to drought stress and recovery. J. Bot. S. Afr. 2009, 75, 34–42. [Google Scholar] [CrossRef]
- Dalal, V.K.; Tripathy, B.C. Water-stress induced downsizing of light-harvesting antenna complex protects developing rice seedlings from photo-oxidative damage. Sci. Rep. 2018, 8, 5955. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence–a practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Guidi, L.; Piccolo, E.L.; Landi, M. Chlorophyll Fluorescence, Photoinhibition and Abiotic Stress: Does it Make Any Difference the Fact to Be a C3 or C4 Species? Front. Plant Sci. 2019, 10, 174. [Google Scholar] [CrossRef]
- Ralph, P.J.; Gademann, R. Rapid light curves: A powerful tool to assess photosynthetic activity. Aquat. Bot. 2005, 82, 222–237. [Google Scholar] [CrossRef]
- Horton, P.; Ruban, A.V. Molecular design of the photosystem II light-harvesting antenna: Photosynthesis and photoprotection. J. Exp. Bot. 2005, 56, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.V.; Johnson, M.P.; Duffy, C.D.P. The photoprotective molecular switch in the photosystem II antenna. Biochim. Biophys. Acta 2012, 1817, 167–181. [Google Scholar]
- Sperdouli, I.; Moustakas, M. Spatio-temporal heterogeneity in Arabidopsis thaliana leaves under drought stress. Plant Biol. 2012, 14, 118–128. [Google Scholar] [CrossRef]
- Liberatore, K.L.; Dukowic-schulze, S.; Miller, M.E.; Chen, C.; Kianian, S.F. The role of mitochondria in plant development and stress tolerance. Free Radic. Biol. Med. 2016, 100, 238–256. [Google Scholar] [CrossRef]
- Trubacheeva, N.V.; Efremova, T.; Badaeva, E.D.; Kravtsova, L.A.; Belova, L.I.; Devyatkina, E.P.; Pershina, L. Production of alloplasmic and euplasmic wheat-barley ditelosomic substitution lines 7H1Lmar(7D) and analysis of the 18S/5S mitochondrial repeat in these lines. Russ. J. Genet. 2009, 45, 1627–1633. [Google Scholar] [CrossRef]
- Middleton, C.P.; Senerchia, N.; Stein, N.; Akhunov, E.D. Sequencing of Chloroplast Genomes from Wheat, Barley, Rye and Their Relatives Provides a Detailed Insight into the Evolution of the Triticeae Tribe. PLoS ONE 2014, 9, 85761. [Google Scholar] [CrossRef] [PubMed]
- Awada, M.; Fahmyb, R.M.; Mosaa, K.A.; Helmyd, M.; El-Fekya, F.A. Identification of effective DNA barcodes for Triticum plants through chloroplast genome-wide analysis. Comput. Biol. Chem. 2017, 71, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Pellegrineschi, A.; Reynolds, M.; Pacheco, M.; Brito, R.M.; Almeraya, R.; Yamaguchi-Shinozaki, K.; Hoisington, D.A. Stress-induced expression in wheat of the Arabidopsis thaliana DREB1A gene delays water stress symptoms under greenhouse conditions. Genome 2004, 47, 493–500. [Google Scholar] [CrossRef]
- Pierre, C.C.; Crossa, J.J.; Bonnett, D.; Yamaguchi-Shinozaki, K.; Reynolds, M.P. Phenotyping transgenic wheat for drought resistance. J. Exp. Bot. 2012, 63, 1799–1808. [Google Scholar] [CrossRef]
- Noor, S.; Ali, S.; Ali, G.M. Comparative Study of Transgenic (DREB1A) and Non-ransgenic Wheat Lines on Relative Water Content, Sugar, Proline and Chlorophyll under Drought and Salt Stresses. Sarhad. J. Agric. 2018, 34, 986–993. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Baho, M.; Lopato, S.; Langridge, P. The TaDREB3 transgene transferred by conventional crossings to different genetic backgrounds of bread wheat improves drought tolerance. Plant Biotechnol. J. 2016, 14, 313–322. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Zhumalin, A.; Serikbay, D.; Botayeva, M.; Otemisova, A.; Absattarova, A.; Sereda, G.; Sereda, S.; Shvidchenko, V.; Turbekova, A.; et al. Expression Level of the DREB2-Type Gene, Identified with Amplifluor SNP Markers, Correlates with Performance, and Tolerance to Dehydration in Bread Wheat Cultivars from Northern Kazakhstan. Front. Plant Sci. 2016, 7, 1736. [Google Scholar] [CrossRef]
- Zotova, L.; Kurishbayev, A.; Jatayev, S.; Khassanova, G.; Zhubatkanov, A.; Serikbay, D.; Sereda, S.; Sereda, T.; Shvidchenko, V.; Lopato, S.; et al. Genes Encoding Transcription Factors TaDREB5 and TaNFYC-A7 Are Differentially Expressed in Leaves of Bread Wheat in Response to Drought, Dehydration and ABA. Front. Plant Sci. 2018, 9, 1441. [Google Scholar] [CrossRef]
- Hsieh, T.-H.; Lee, J.-T.; Yang, P.-T.; Chiu, L.-H.; Charng, Y.-Y.; Wang, Y.-C.; Chan, M.-T. Heterology expression of the Arabidopsis C-repeat/dehydration response element binding factor, gene confers elevated tolerance to chilling and oxidative stresses in transgenic tomato. Plant Physiol. 2002, 129, 1086–1094. [Google Scholar] [CrossRef]
- Savitch, L.V.; Allard, G.; Seki, M.; Robert, L.S.; Tinker, N.A.; Hüner, N.P.A.; Shinozaki, K.; Singh, J. The effect of overexpression of two Brassica CBF/DREB1-like transcription factors on photosynthetic capacity and freezing tolerance in Brassica napus. Plant Cell Physiol. 2005, 46, 1525–1539. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Kwon, C.W.; Choi, D.W.; Song, S.I.; Kim, J.K. Expression of barley HvCBF4 enhances tolerance to abiotic stress in transgenic rice. Plant Biotechnol. J. 2007, 5, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Jing, R.; Wang, C.; Chen, J.; Mao, X.; Chang, X.; Jia, J. Dreb1 genes in wheat (Triticum aestivum L.): Development of functional markers and gene mapping based on SNPs. Mol. Breed. 2009, 23, 13–22. [Google Scholar] [CrossRef]
- Kiseleva, A.A.; Shcherban, A.B.; Leonova, I.N.; Frenkel, Z.; Salina, E.A. Identification of new heading date determinants in wheat 5B chromosome. BMC Plant Biol. 2016, 16, 8. [Google Scholar] [CrossRef] [PubMed]
- Roeder, M.S.; Korzun, V.; Wendehake, K.; Plaschke, J.; Tixier, M.H.; Leroy, P.; Ganal, M.W. A microsatellite map of wheat. Genetics 1998, 149, 2007–2023. [Google Scholar]
- Felsenstein, J. PHYLIP, Phylogeny Inference Package (Version 3.5c). Department of Genetics, University of Washington: Seattle. WA, USA, 1995. [Google Scholar]
- Diagnostics stability of plant resistance to stress. Methodical Guidance; Udovenko, G.V., Ed.; VIR: Leningrad, Russia, 1988; p. 89. [Google Scholar]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis in Vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species, Line | Root, cm | Leaf, cm | ||||
|---|---|---|---|---|---|---|
| Control | Stress | % to control | Control | Stress | % to control | |
| T. aestivum | 8.22 ± 1.53 | 6.86 ± 0.81 | 83 | 15.42 ± 1.15 | 13.3 ± 1.04 | 86 |
| T. dicoccum | 7.03 ± 0.65 | 6.58 ± 0.85 | 94 | 16.92 ± 1.10 | 13.99 ± 1.21 | 83 |
| D-a-05 | 7.46 ± 0.61 | 4.32 ± 0.76 ad* | 58 | 14.74 ± 1.11 | 11.83 ± 1.06 | 80 |
| D-b-05 | 7.81 ± 1.66 | 7.04 ± 0.96 | 90 | 14.66 ± 1.42 | 14.02 ± 1.28 | 95 |
| D-f-05 | 7.43 ± 0.84 | 4.71 ± 0.62 a* | 63 | 17.60 ± 1.19 | 11.10 ± 1.11 * | 63 |
| D-n-05 | 7.65 ± 1.13 | 4.30 ± 0.91 ad* | 56 | 14.26 ± 1.30 | 11.04 ± 1.33 | 77 |
| D-d-05 | 10.64 ± 0.84 | 6.76 ± 0.78 * | 64 | 15.61 ± 1.50 | 12.74 ± 1.71 | 82 |
| D-d-05 b | 9.39 ± 1.99 | 8.17 ± 0.71 | 87 | 17.20 ± 1.61 | 14.43 ± 1.71 | 83 |
| D-40-05 | 9.67 ± 1.37 | 7.45 ± 1.12 | 77 | 16.46 ± 1.37 | 14.24 ± 1.32 | 87 |
| D-41-05 | 7.26 ± 1.07 | 6.43 ± 1.27 | 88 | 15.69 ± 1.30 | 13.10 ± 1.11 | 83 |
| D-42-05 | 9.26 ± 1.33 | 6.33 ± 0.92 | 68 | 15.91 ± 0.97 | 12.63 ± 1.37 | 79 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terletskaya, N.V.; Shcherban, A.B.; Nesterov, M.A.; Perfil’ev, R.N.; Salina, E.A.; Altayeva, N.A.; Blavachinskaya, I.V. Drought Stress Tolerance and Photosynthetic Activity of Alloplasmic Lines T. dicoccum x T. aestivum. Int. J. Mol. Sci. 2020, 21, 3356. https://doi.org/10.3390/ijms21093356
Terletskaya NV, Shcherban AB, Nesterov MA, Perfil’ev RN, Salina EA, Altayeva NA, Blavachinskaya IV. Drought Stress Tolerance and Photosynthetic Activity of Alloplasmic Lines T. dicoccum x T. aestivum. International Journal of Molecular Sciences. 2020; 21(9):3356. https://doi.org/10.3390/ijms21093356
Chicago/Turabian StyleTerletskaya, Nina V., Andrey B. Shcherban, Michail A. Nesterov, Roman N. Perfil’ev, Elena A. Salina, Nazira A. Altayeva, and Irina V. Blavachinskaya. 2020. "Drought Stress Tolerance and Photosynthetic Activity of Alloplasmic Lines T. dicoccum x T. aestivum" International Journal of Molecular Sciences 21, no. 9: 3356. https://doi.org/10.3390/ijms21093356
APA StyleTerletskaya, N. V., Shcherban, A. B., Nesterov, M. A., Perfil’ev, R. N., Salina, E. A., Altayeva, N. A., & Blavachinskaya, I. V. (2020). Drought Stress Tolerance and Photosynthetic Activity of Alloplasmic Lines T. dicoccum x T. aestivum. International Journal of Molecular Sciences, 21(9), 3356. https://doi.org/10.3390/ijms21093356