Selective Activation of ZAK β Expression by 3-Hydroxy-2-Phenylchromone Inhibits Human Osteosarcoma Cells and Triggers Apoptosis via JNK Activation

 , ,
, ,  and
and 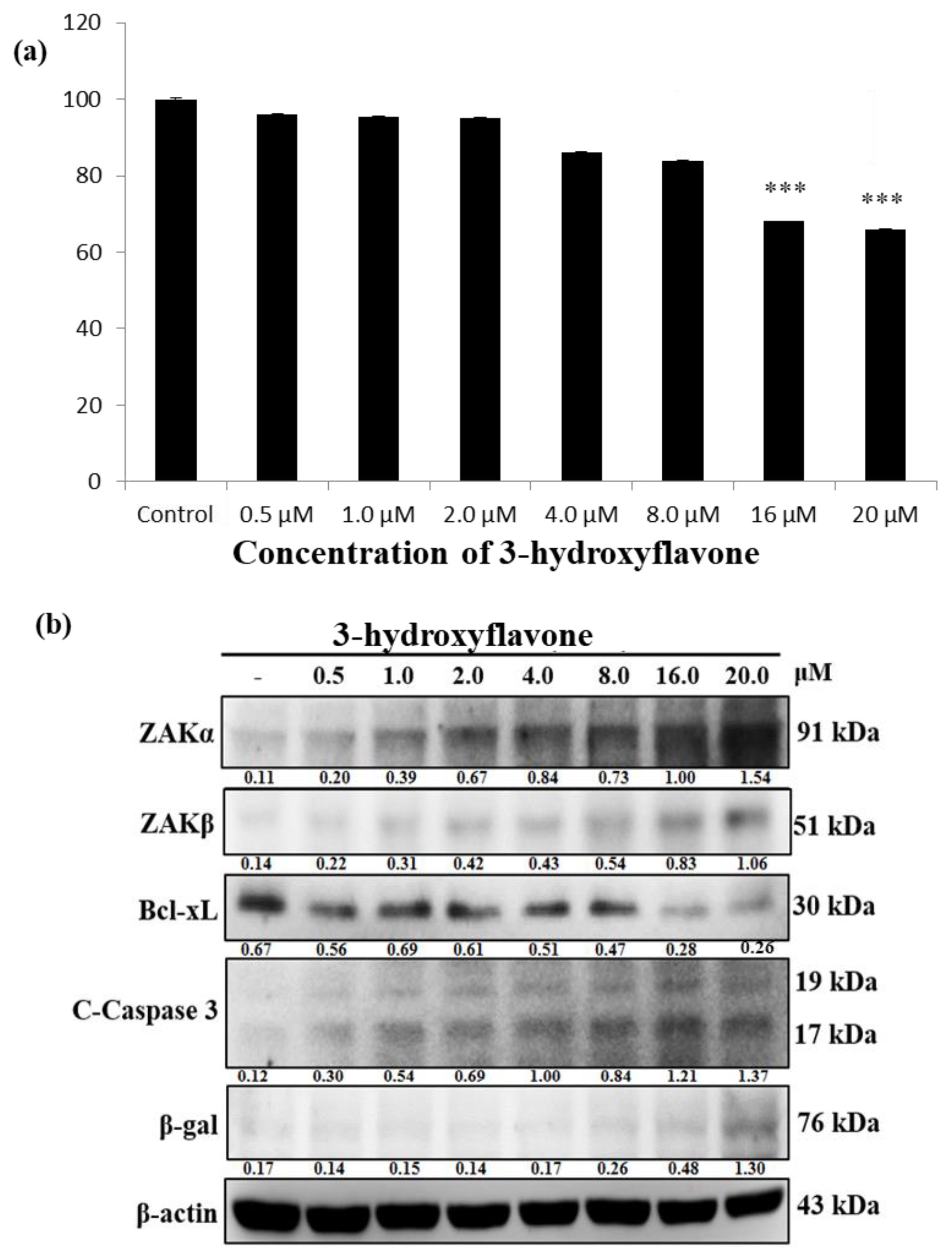{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. 3-HF Reduces the Viability of Human Osteosarcoma Cells, which Correlates with Simultaneous Upregulation in ZAKα, β Levels and Cleaved Caspase Levels
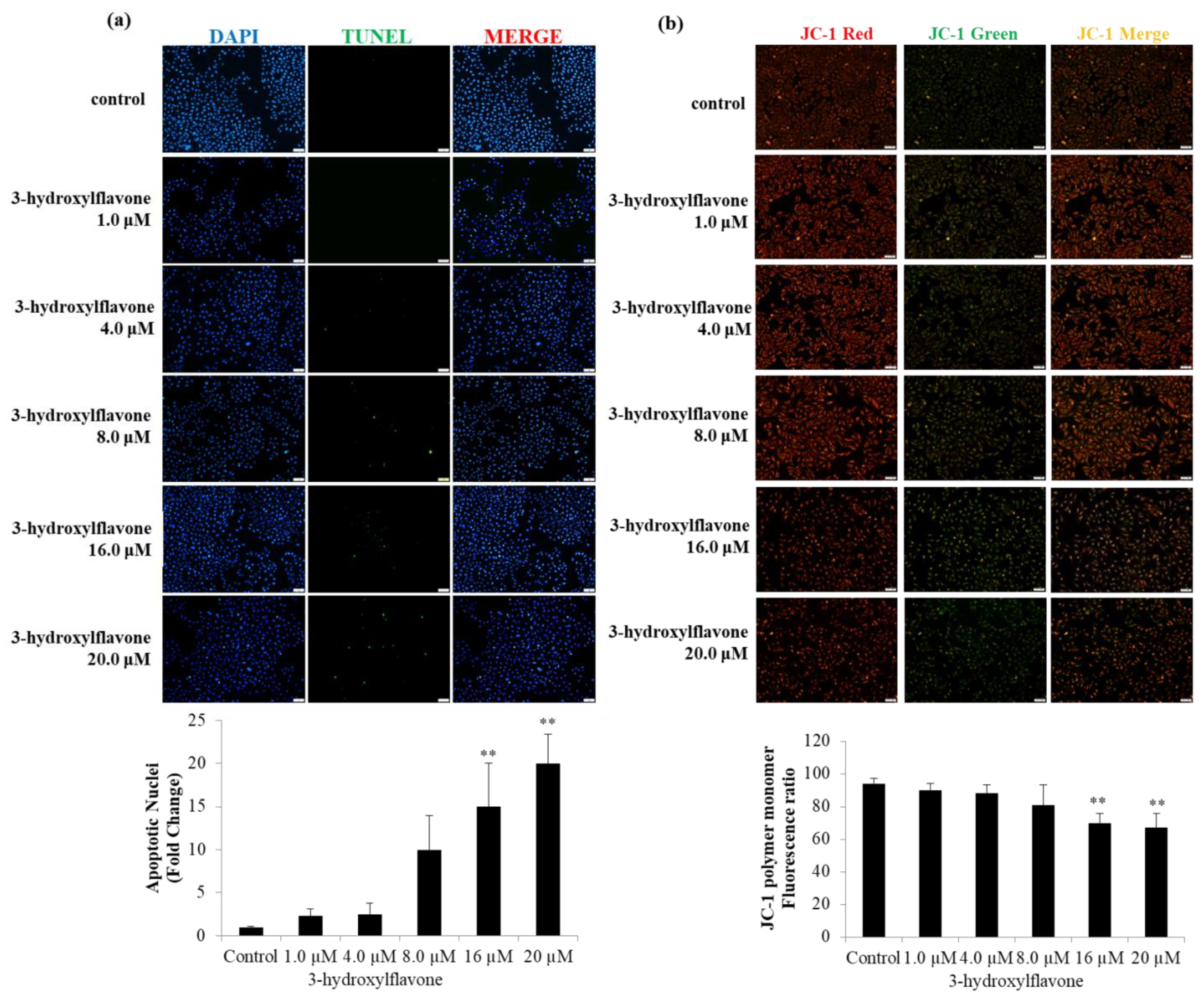
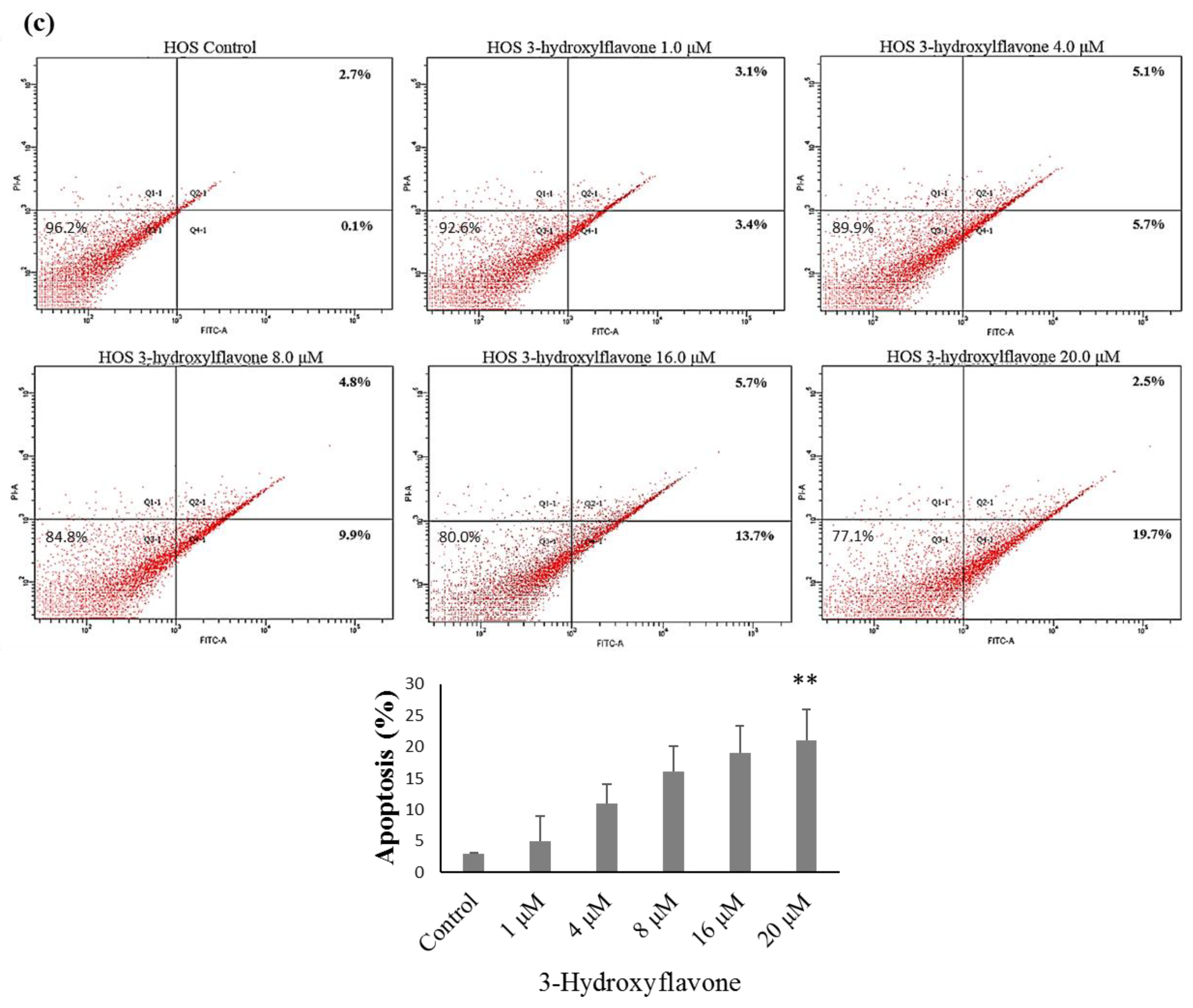
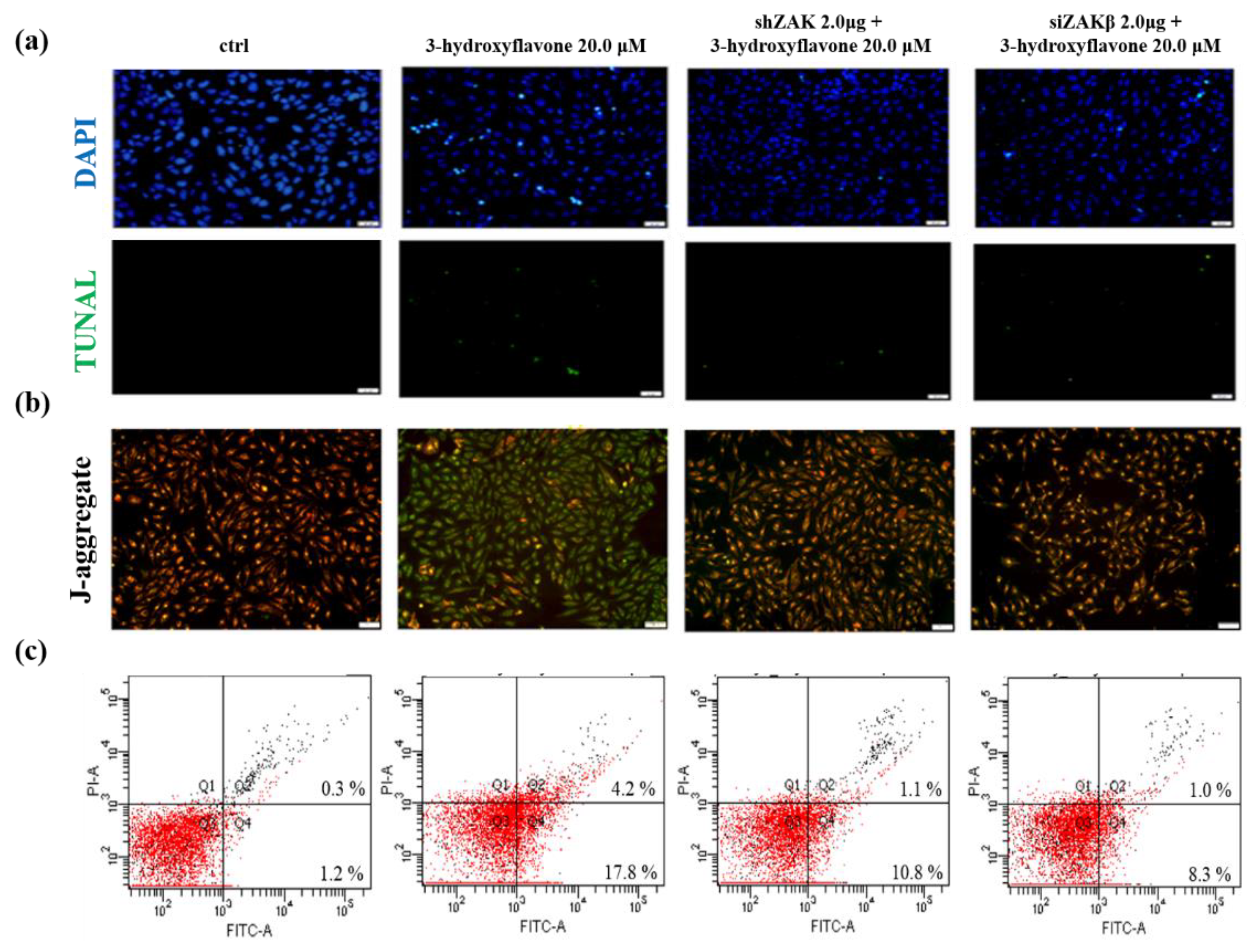
2.2. 3-HF Can Trigger Cell Apoptosis and Decrease Mitochondrial Membrane Potential
2.3. 3-HF Can Upregulate ZAK Expression to Induce Cell Apoptosis in Human OS Cells
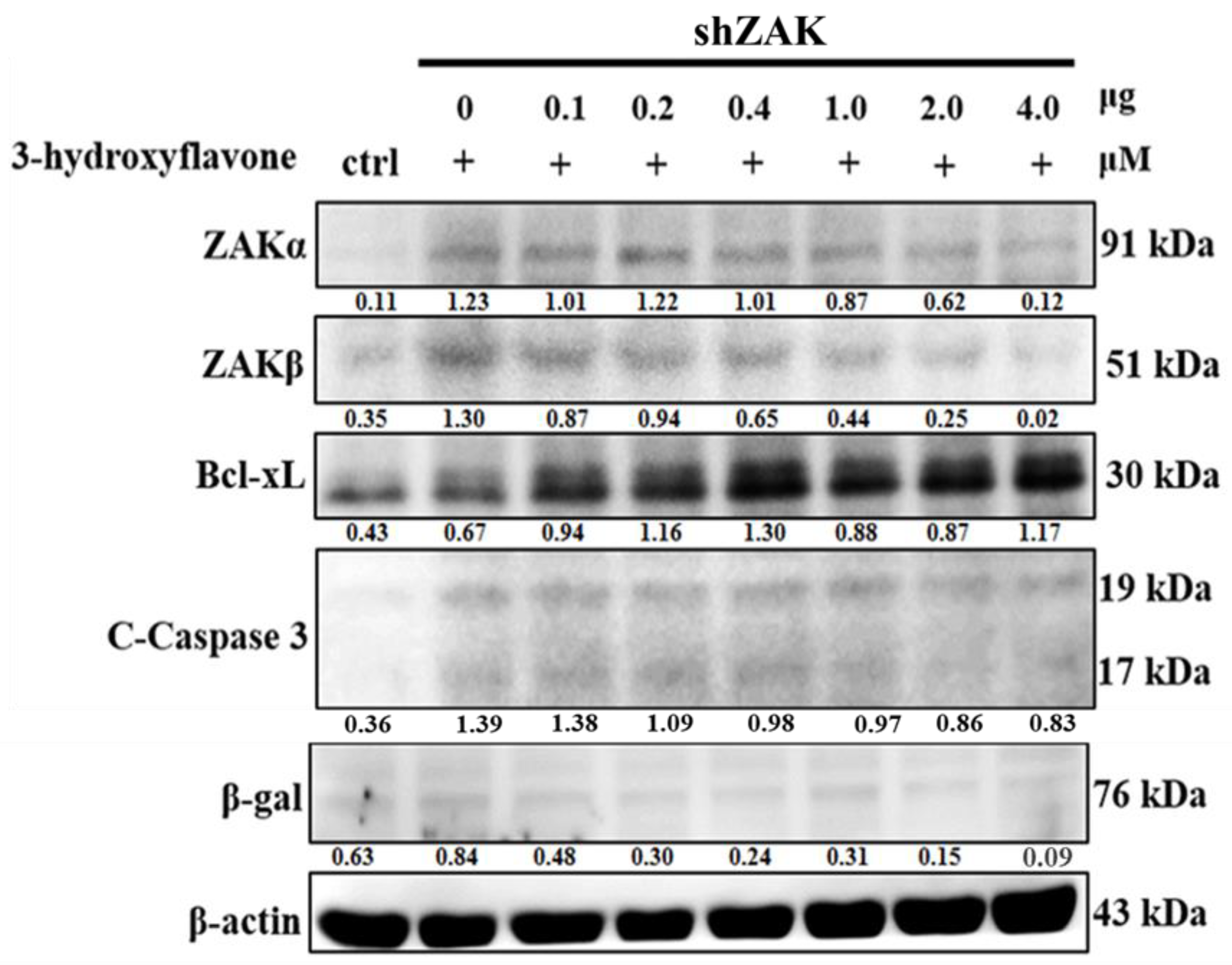
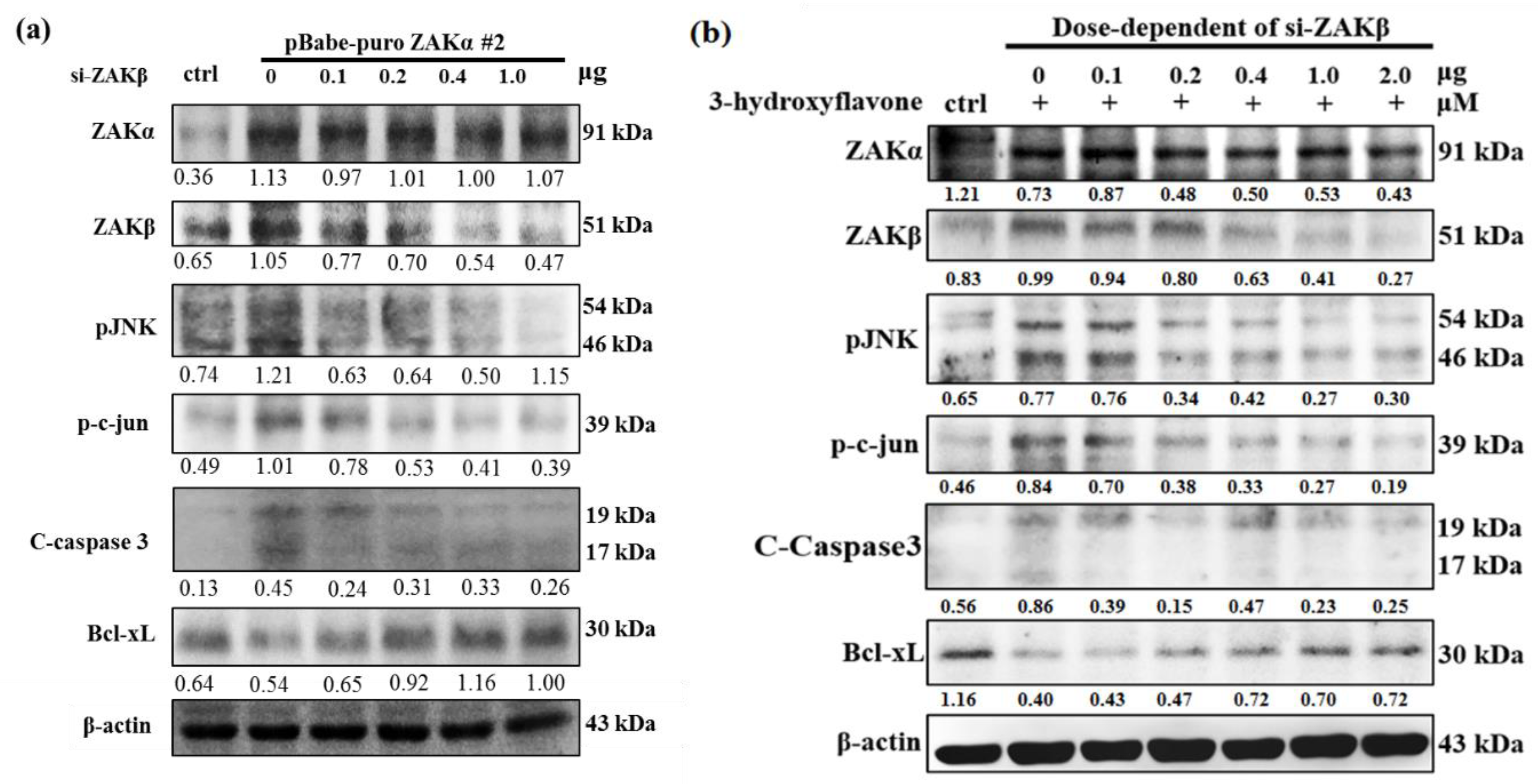
2.4. Cell Apoptosis Induced by 3-HF is Due to ZAKβ Overexpression not ZAKα in Human OS Cells
2.5. 3-HF Can Induce Cell Apoptosis by ZAKβ in Human OS Cells
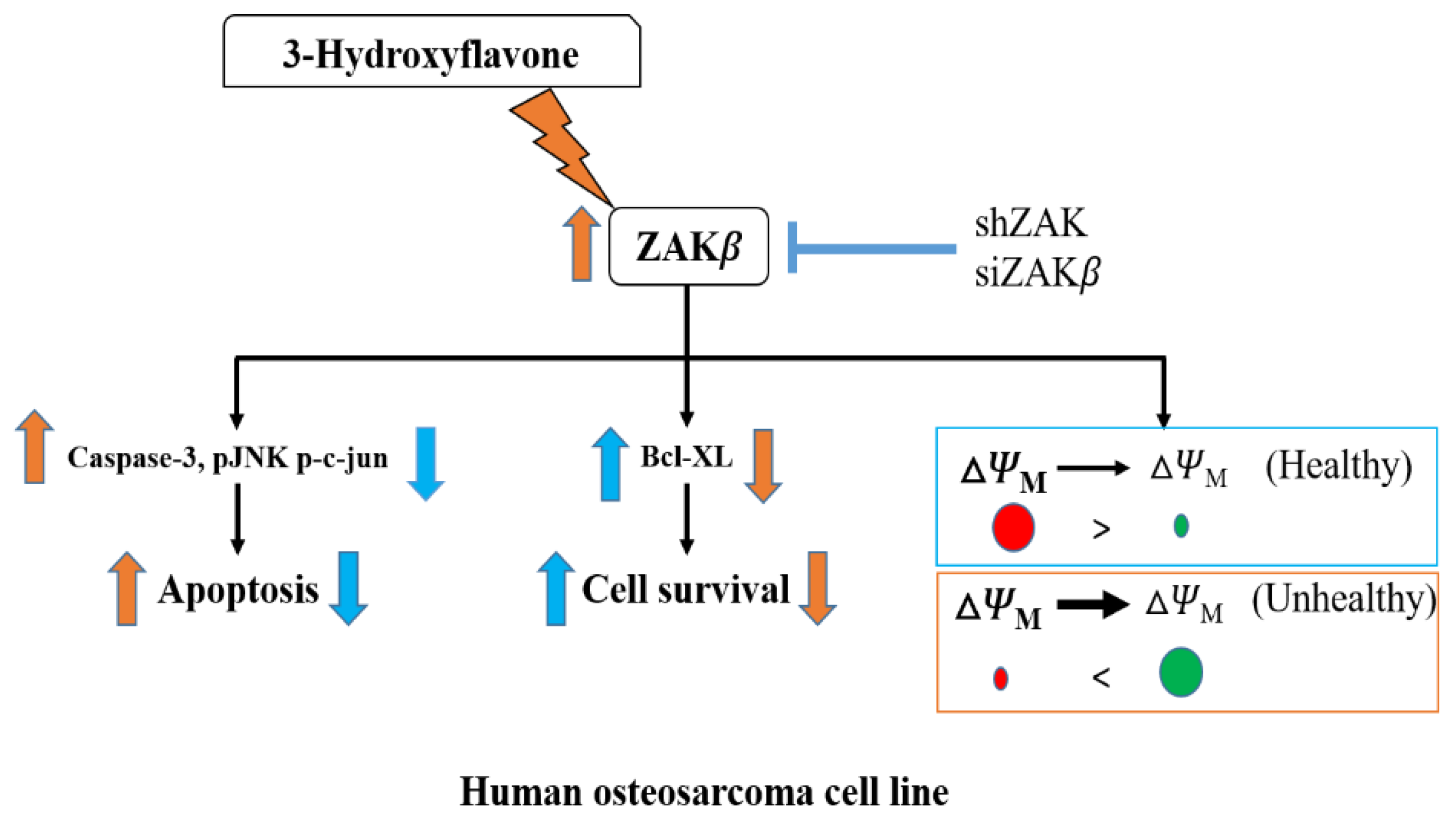
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Whole Cell Extraction
4.3. MTT Assay
4.4. Transient Transfection
4.5. siRNA Transfection
4.6. Western Blot
4.7. TUNEL
4.8. JC-1 Staining
4.9. Detection of Cell Apoptosis Using Flow Cytometry
4.10. Antibodies and Reagents
4.11. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| OS | Osteosarcoma |
| ZAK | Zipper sterile-alpha-motif kinase |
| ∆ΨM | Mitochondrial membrane potential |
References
- Bielack, S.; Carrle, D.; Casali, P.G. Osteosarcoma: ESMO clinical recommendations for diagnosis, treatment and follow-up. Ann. Oncol. 2009, 20, 137–139. [Google Scholar] [CrossRef]
- Wu, P.K.; Chen, W.M.; Chen, C.F.; Lee, O.K.; Haung, C.K.; Chen, T.H. Primary Osteogenic Sarcoma with Pulmonary Metastasis: Clinical Results and Prognostic Factors in 91 Patients. Jpn. J. Clin. Oncol. 2009, 39, 514–522. [Google Scholar] [CrossRef] [Green Version]
- Ottaviani, G.; Jaffe, N. The epidemiology of osteosarcoma. Cancer Treat. Res. 2009, 152, 3–13. [Google Scholar] [PubMed]
- Wang, B.; Xu, M.; Zheng, K.; Yu, X. Effect of Unplanned Therapy on the Prognosis of Patients with Extremity Osteosarcoma. Sci. Rep. 2016, 6, 38783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, C.Y.; Tseng, Y.S.; Chen, M.C.; Hsu, H.H.; Yang, J.J.; Tu, C.C.; Lin, Y.M.; Viswanadha, V.P.; Kuo, W.W.; Huang, C.Y. Doxorubicin induces ZAKalpha overexpression with a subsequent enhancement of apoptosis and attenuation of survivability in human osteosarcoma cells. Environ. Toxicol. 2018, 33, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Lin, L.; Li, X.-P.; Yu, Y.-X.; Zhang, G.-L.; Chen, W.-J. All-optical switchings of 3-hydroxyflavone in different solvents. Chin. Phys. B 2008, 17, 1461. [Google Scholar]
- Lang, Q.; Zhang, H.; Li, J.; Xie, F.; Zhang, Y.; Wan, B.; Yu, L. 3-Hydroxyflavone inhibits endogenous Aurora B and induces growth inhibition of cancer cell line. Mol. Biol. Rep. 2010, 37, 1577–1583. [Google Scholar] [CrossRef]
- Lu, K.H.; Chen, P.N.; Hsieh, Y.H.; Lin, C.Y.; Cheng, F.Y.; Chiu, P.C.; Chu, S.C.; Hsieh, Y.S. 3-Hydroxyflavone inhibits human osteosarcoma U2OS and 143B cells metastasis by affecting EMT and repressing u-PA/MMP-2 via FAK-Src to MEK/ERK and RhoA/MLC2 pathways and reduces 143B tumor growth in vivo. Food Chem. Toxicol. 2016, 97, 177–186. [Google Scholar] [CrossRef]
- Uhm, S.J.; Gupta, M.K.; Das, Z.C.; Kim, N.H.; Lee, H.T. 3-Hydroxyflavone improves the in vitro development of cloned porcine embryos by inhibiting ROS production. Cell. Reprogramming 2011, 13, 441–449. [Google Scholar] [CrossRef]
- Liu, T.C.; Huang, C.J.; Chu, Y.C.; Wei, C.C.; Chou, C.C.; Chou, M.Y.; Chou, C.K.; Yang, J.J. Cloning and expression of ZAK, a mixed lineage kinase-like protein containing a leucine-zipper and a sterile-alpha motif. Biochem. Biophys. Res. Commun. 2000, 274, 811–816. [Google Scholar] [CrossRef]
- Gotoh, I.; Adachi, M.; Nishida, E. Identification and characterization of a novel MAP kinase kinase kinase, MLTK. J. Biol. Chem. 2001, 276, 4276–4286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jandhyala, D.M.; Ahluwalia, A.; Obrig, T.; Thorpe, C.M. ZAK: A MAP3Kinase that transduces Shiga toxin- and ricin-induced proinflammatory cytokine expression. Cell. Microbiol. 2008, 10, 1468–1477. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.J. Mixed lineage kinase ZAK utilizing MKK7 and not MKK4 to activate the c-Jun N-terminal kinase and playing a role in the cell arrest. Biochem. Biophys. Res. Commun. 2002, 297, 105–110. [Google Scholar] [CrossRef]
- Fu, C.Y.; Kuo, W.W.; Ho, T.J.; Wen, S.Y.; Lin, L.C.; Tseng, Y.S.; Hung, H.C.; Viswanadha, V.P.; Yang, J.J.; Huang, C.Y. ZAKbeta antagonizes and ameliorates the cardiac hypertrophic and apoptotic effects induced by ZAKalpha. Cell Biochem. Funct. 2016, 34, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Christe, M.; Jin, N.; Wang, X.; Gould, K.E.; Iversen, P.W.; Yu, X.; Lorenz, J.N.; Kadambi, V.; Zuckerman, S.H.; Bloem, L.J. Transgenic mice with cardiac-specific over-expression of MLK7 have increased mortality when exposed to chronic beta-adrenergic stimulation. J. Mol. Cell. Cardiol. 2004, 37, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.Y.; Tseng, Y.S.; Chen, M.C.; Hsu, H.H.; Yang, J.J.; Tu, C.C.; Lin, Y.M.; Viswanadha, V.P.; Ding, K.; Kuo, W.W.; et al. Overexpression of ZAKbeta in human osteosarcoma cells enhances ZAKalpha expression, resulting in a synergistic apoptotic effect. Cell Biochem. Funct. 2018, 36, 176–182. [Google Scholar] [CrossRef]
- Worch, J.; Matthay, K.K.; Neuhaus, J.; Goldsby, R.; DuBois, S.G. Osteosarcoma in children 5 years of age or younger at initial diagnosis. Pediatr. Blood Cancer 2010, 55, 285–289. [Google Scholar] [CrossRef]
- Taran, S.J.; Taran, R.; Malipatil, N.B. Pediatric Osteosarcoma: An Updated Review. Indian J. Med. Paediatr. Oncol. 2017, 38, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Buondonno, I.; Gazzano, E.; Jean, S.R.; Audrito, V.; Kopecka, J.; Fanelli, M.; Salaroglio, I.C.; Costamagna, C.; Roato, I.; Mungo, E.; et al. Mitochondria-Targeted Doxorubicin: A New Therapeutic Strategy against Doxorubicin-Resistant Osteosarcoma. Mol. Cancer Ther. 2016, 15, 2640–2652. [Google Scholar] [CrossRef] [Green Version]
- Mavrogenis, A.F.; Angelini, A.; Sakellariou, V.I.; Skarpidi, E.; Ruggieri, P.; Papagelopoulos, P.J. Osteosarcoma invasion of the inferior vena cava and right atrium. J. Surg. Orthop. Adv. 2012, 21, 107–112. [Google Scholar] [CrossRef]
- Munajat, I.; Zulmi, W.; Norazman, M.Z.; Wan Faisham, W.I. Tumour volume and lung metastasis in patients with osteosarcoma. J. Orthop. Surg. 2008, 16, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, Z.; Li, Y.; Xia, J.; Li, D.; Li, H.; Ren, M.; Liao, Y.; Yu, S.; Chen, Y.; et al. Cell apoptosis, autophagy and necroptosis in osteosarcoma treatment. Oncotarget 2016, 7, 44763–44778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Xu, Y.; Lin, N.; Jiang, S.; Wang, Y.; Ye, Z. High-dose methotrexate (HD-MTX) used as an adjunct with other chemotherapeutics for the treatment of osteosarcoma. Cell Biochem. Biophys. 2015, 71, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Ni, J.; Huang, J. Molecular mechanisms of chemoresistance in osteosarcoma (Review). Oncol. Lett. 2014, 7, 1352–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conner, C.S. Mitoxantrone: A replacement for doxorubicin? Drug Intell. Clin. Pharm. 1984, 18, 479–480. [Google Scholar] [CrossRef]
- Davidson, N.; Camburn, T.; Keary, I.; Houghton, D. Substituting Doxorubicin with nonpegylated liposomal Doxorubicin for the treatment of early breast cancer: Results of a retrospective study. Int. J. Breast Cancer 2014, 2014, 984067. [Google Scholar] [CrossRef] [PubMed]
- Vin, H.; Ching, G.; Ojeda, S.S.; Adelmann, C.H.; Chitsazzadeh, V.; Dwyer, D.W.; Ma, H.; Ehrenreiter, K.; Baccarini, M.; Ruggieri, R.; et al. Sorafenib suppresses JNK-dependent apoptosis through inhibition of ZAK. Mol. Cancer Ther. 2014, 13, 221–229. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-Y.; Lee, S.-D. Possible pathophysiology of heart failure in obesity: Cardiac apoptosis. BioMedicine 2012, 2, 36–40. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, C.-Y.; Lay, I.-S.; Shibu, M.A.; Tseng, Y.-S.; Kuo, W.-W.; Yang, J.-J.; Wang, T.-F.; Mahalakshmi, B.; Yeh, Y.-L.; Huang, C.-Y. Selective Activation of ZAK β Expression by 3-Hydroxy-2-Phenylchromone Inhibits Human Osteosarcoma Cells and Triggers Apoptosis via JNK Activation. Int. J. Mol. Sci. 2020, 21, 3366. https://doi.org/10.3390/ijms21093366
Fu C-Y, Lay I-S, Shibu MA, Tseng Y-S, Kuo W-W, Yang J-J, Wang T-F, Mahalakshmi B, Yeh Y-L, Huang C-Y. Selective Activation of ZAK β Expression by 3-Hydroxy-2-Phenylchromone Inhibits Human Osteosarcoma Cells and Triggers Apoptosis via JNK Activation. International Journal of Molecular Sciences. 2020; 21(9):3366. https://doi.org/10.3390/ijms21093366
Chicago/Turabian StyleFu, Chien-Yao, Ing-Shiow Lay, Marthandam Asokan Shibu, Yan-Shen Tseng, Wei-Wen Kuo, Jaw-Ji Yang, Tso-Fu Wang, B. Mahalakshmi, Yu-Lan Yeh, and Chih-Yang Huang. 2020. "Selective Activation of ZAK β Expression by 3-Hydroxy-2-Phenylchromone Inhibits Human Osteosarcoma Cells and Triggers Apoptosis via JNK Activation" International Journal of Molecular Sciences 21, no. 9: 3366. https://doi.org/10.3390/ijms21093366
APA StyleFu, C.-Y., Lay, I.-S., Shibu, M. A., Tseng, Y.-S., Kuo, W.-W., Yang, J.-J., Wang, T.-F., Mahalakshmi, B., Yeh, Y.-L., & Huang, C.-Y. (2020). Selective Activation of ZAK β Expression by 3-Hydroxy-2-Phenylchromone Inhibits Human Osteosarcoma Cells and Triggers Apoptosis via JNK Activation. International Journal of Molecular Sciences, 21(9), 3366. https://doi.org/10.3390/ijms21093366