Characterization and Differentiation of the Tumor Microenvironment (TME) of Orthotopic and Subcutaneously Grown Head and Neck Squamous Cell Carcinoma (HNSCC) in Immunocompetent Mice

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Squamous Cell Carcinoma of the Head and Neck
1.2. Tumor Microenvironment and Immunosuppressive Mechanisms
1.3. Immunotherapy Supports an Antitumoral Microenvironment
1.4. Tumor Location Is Reported to Affect the Tumor Microenvironment
1.5. The C3H/HeN-SCC VII Model Represents an Immunocompetent HNSCC System
2. Results
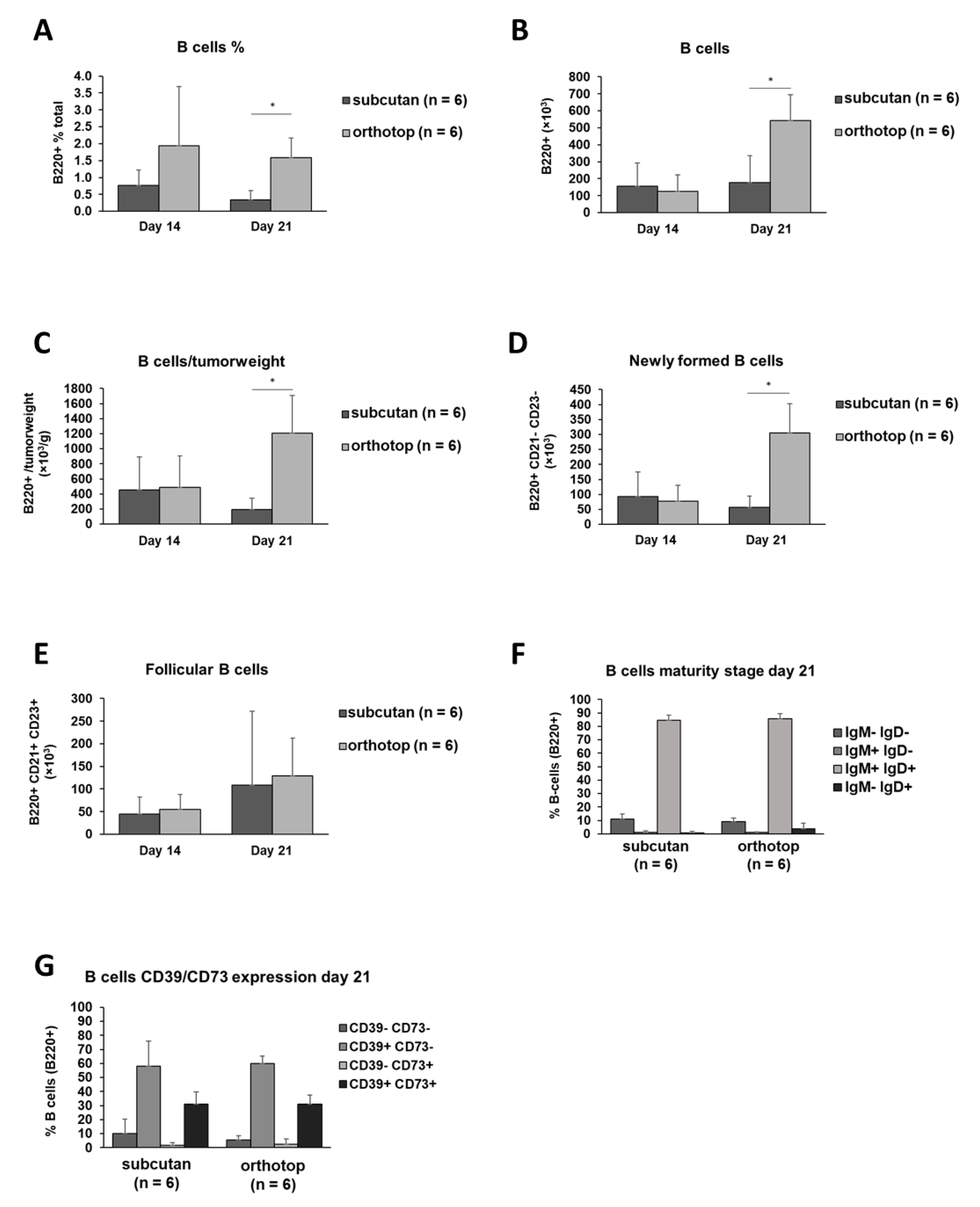
2.1. Increased B Cell and CD4+ T Cell Numbers in the Orthotopic TME
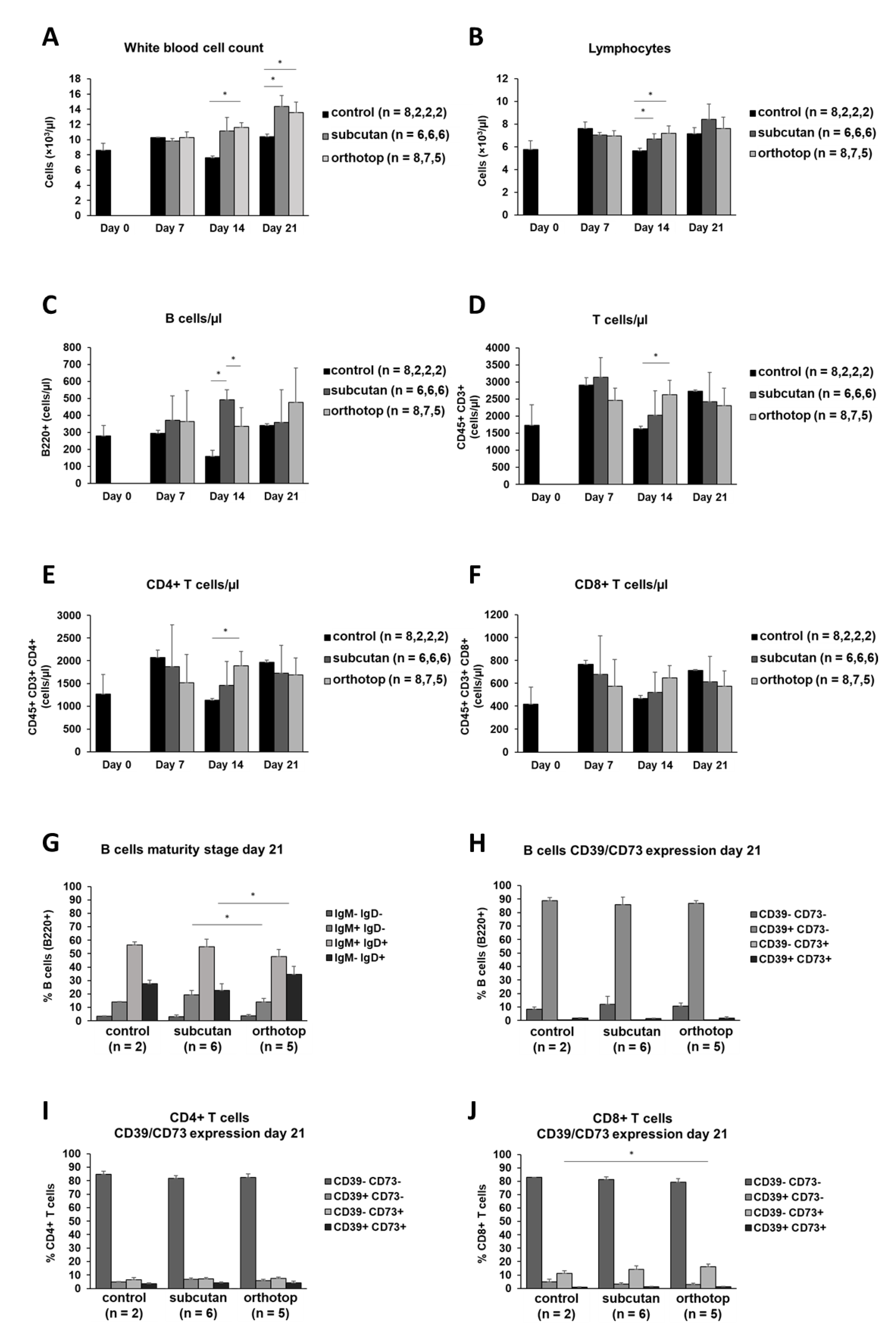
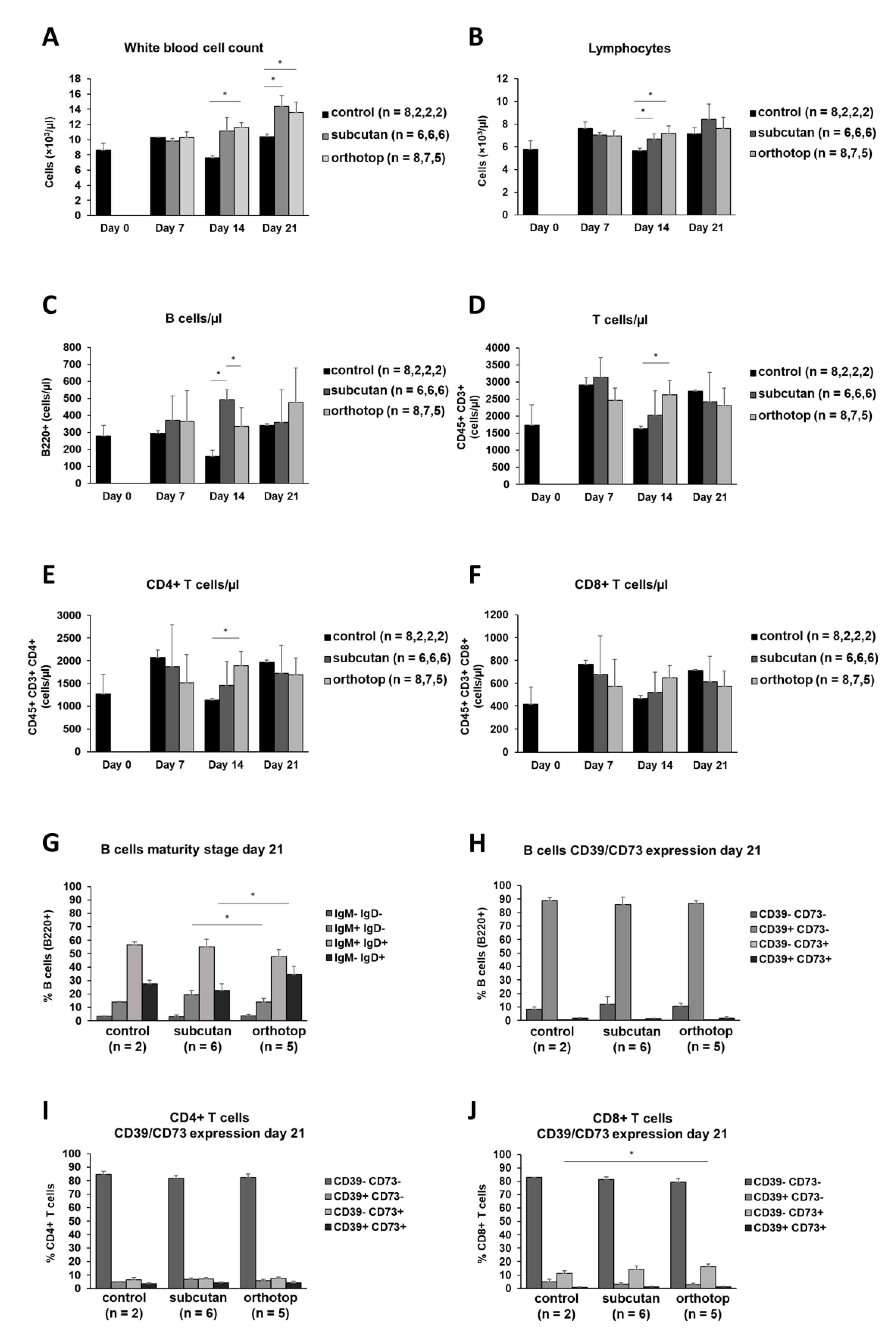
2.2. Tumor Formation Leads to an Increase in the Number of Leukocytes in the Blood of Tumor-Bearing Mice
2.3. Splenic B and T Cells of Both Tumor Models Did Not Show Any Significant Differences
2.4. The Immunogenic Composition of Inguinal Lymph Nodes Is Identical in Both Tumor Models and Is Not Affected by Tumor Formation
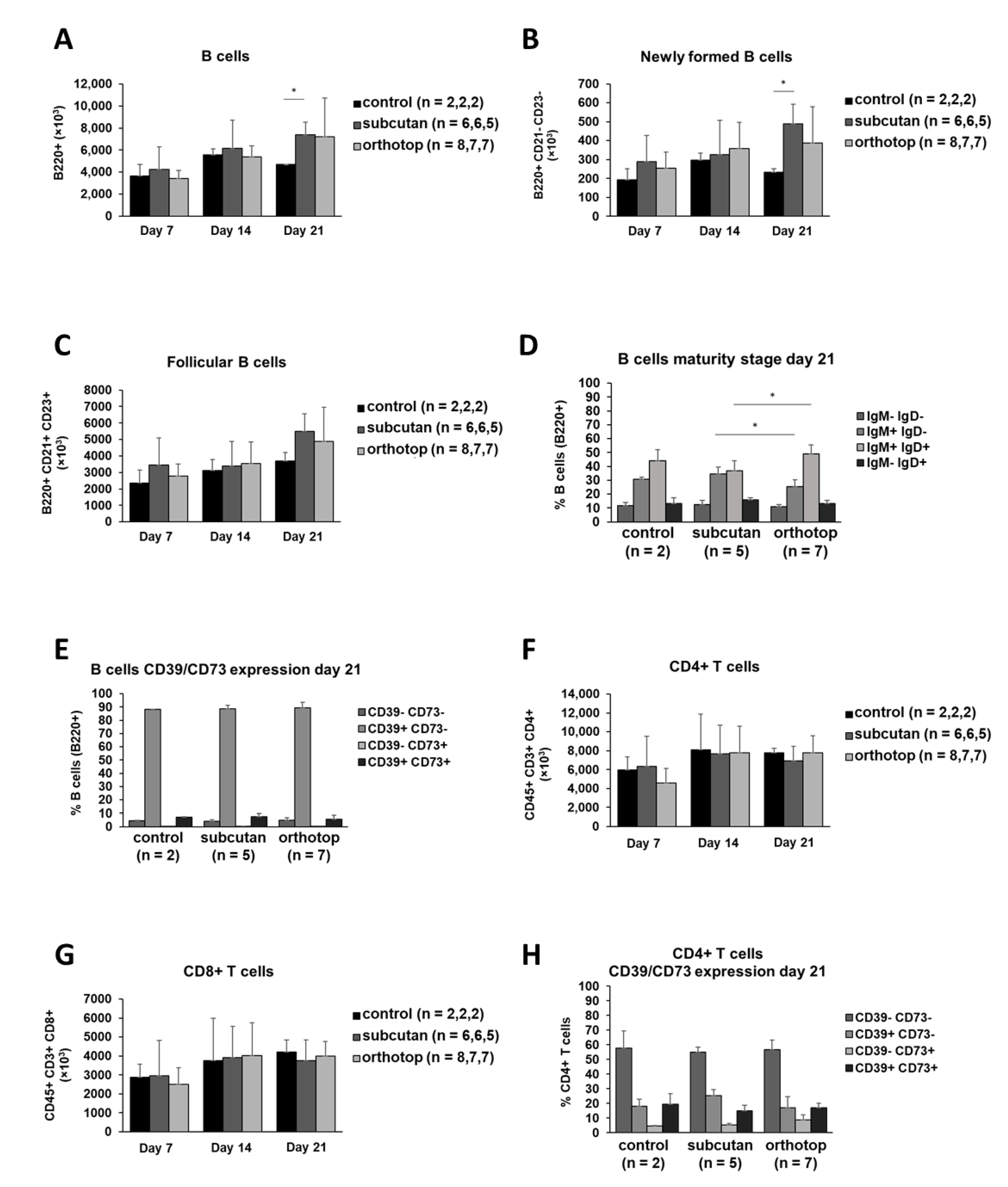
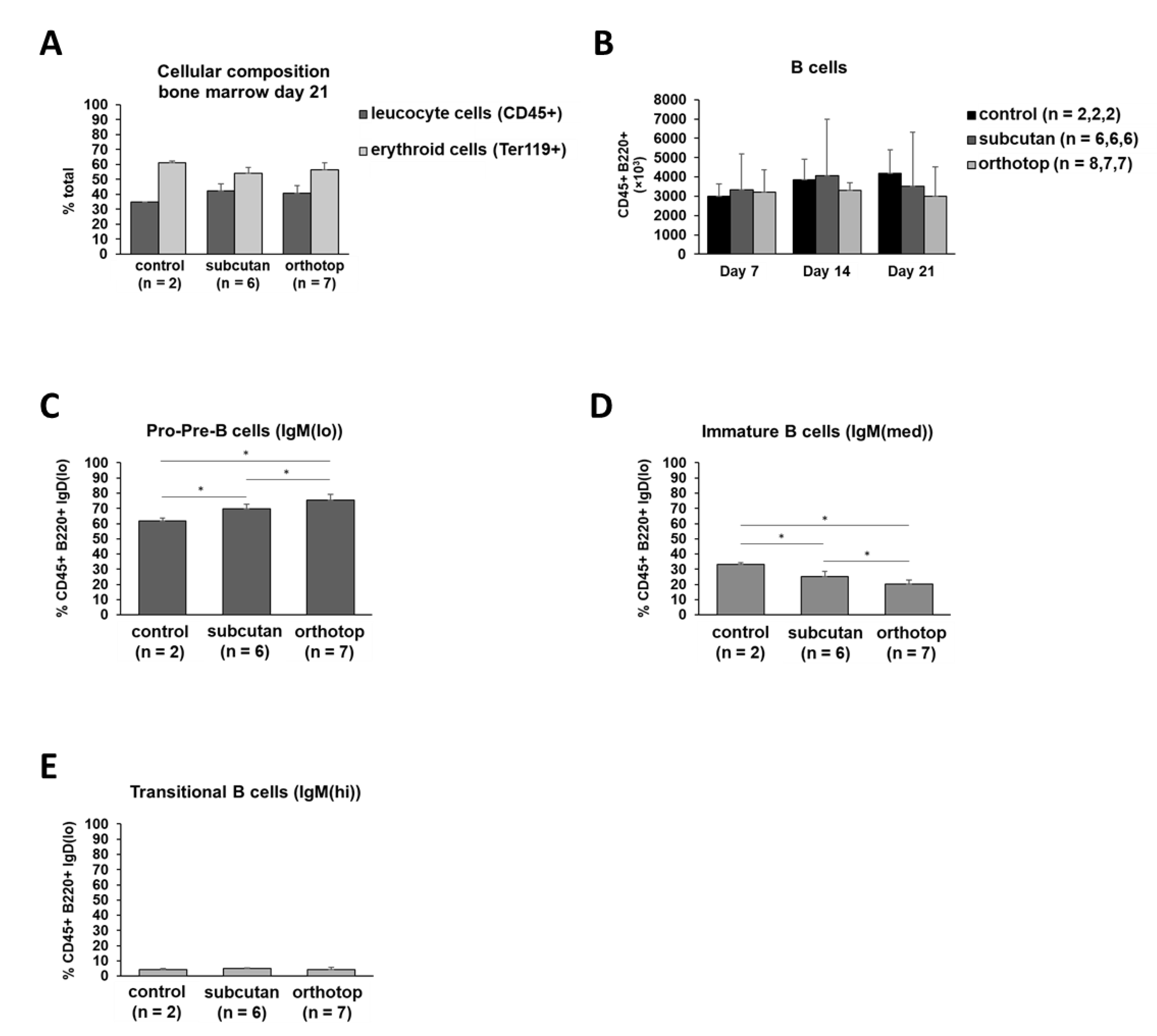
2.5. Tumor Formation Leads to Higher B Cell Proliferation and B Cell Mobilization in Bone Marrow
2.6. Tumor Formation and Location Has No Effect on B Cell and T Cell Populations in the Thymus
3. Discussion
3.1. The SCC VII-C3H/HeN System Is a Reliable Model to Study HNSCC in Immunocompetent Mice
3.2. A Larger Amount of B Cells, CD4+ T Cells, Macrophages, Dendritic Cells, and Granulocytes Creates a More Active TME in Orthotopic Grown Tumors, Accompanied by a Reduced Tumor Volume
3.3. An Increase in the White Blood Cell Count in Peripheral Blood Indicates a Systemic Inflammatory Reaction
3.4. B Cell and T Cell Populations in Spleen, Lymph Nodes, and Thymus Were Not Affected by Tumor Induction
3.5. The SCC VII-C3H/HeN Mouse Model Reflects a Situation Comparable to HNSCC Patients
4. Materials and Methods
4.1. Cell Culture
4.2. Animal Model
4.3. Tumor Preparation
4.4. Spleen, Lymph Node, and Thymus Preparation
4.5. Antibodies and Reagents
4.6. Surface Staining
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATP | adenosine-triphosphate |
| Breg | regulatory B cells |
| CD | cluster of differentiation |
| DOAJ | Directory of open access journals |
| FOXP3 | forkhead-box-protein P3 |
| HNC | head and neck cancer |
| HNSCC | head and neck squamous cell carcinoma |
| HPV | human papillomavirus |
| mAb | monoclonal antibody |
| MDPI | Multidisciplinary Digital Publishing Institute |
| NK cells | natural-killer cells |
| SD | standard deviation |
| TAM | tumor associated macrophages |
| TIL | tumor-infiltrating lymphocytes |
| TLR-4 | toll-like-receptor-4 |
| TME | tumor micoenvironment |
| Treg | regulatory T cells |
| VEGF | vascular endothelial growth factor |
| WBC | white blood cell count |
References
- Ghanizada, M.; Jakobsen, K.K.; Grønhøj, C.; von Buchwald, C. The effects of checkpoint inhibition on head and neck squamous cell carcinoma: A systematic review. Oral Oncol. 2019, 90, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Warnakulasuriya, S. Global epidemiology of oral and oropharyngeal cancer. Oral Oncol. 2009, 45, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Marur, S.; Forastiere, A.A. Head and neck cancer: Changing epidemiology, diagnosis, and treatment. In Proceedings of the Mayo Clinic Proceedings; Elsevier: Amsterdam, The Netherlands, 2008; Volume 83, pp. 489–501. [Google Scholar]
- Freiser, M.E.; Serafini, P.; Weed, D.T. The immune system and head and neck squamous cell carcinoma: From carcinogenesis to new therapeutic opportunities. Immunol. Res. 2013, 57, 52–69. [Google Scholar] [CrossRef]
- Schwartz, M.; Zhang, Y.; Rosenblatt, J.D. B cell regulation of the anti-tumor response and role in carcinogenesis. J. Immunother. Cancer 2016, 4, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferris, R.L.; Blumenschein, G.; Fayette, J.; Guigay, J.; Colevas, A.D.; Licitra, L.; Harrington, K.; Kasper, S.; Vokes, E.E.; Even, C.; et al. Nivolumab for recurrent squamous-cell carcinoma of the head and neck. N. Engl. J. Med. 2016, 375, 1856–1867. [Google Scholar] [CrossRef]
- Ibáñez-Vea, M.; Zuazo, M.; Gato, M.; Arasanz, H.; Fernández-Hinojal, G.; Escors, D.; Kochan, G. Myeloid-Derived Suppressor Cells in the Tumor Microenvironment: Current Knowledge and Future Perspectives. Arch. Immunol. Ther. Exp. (Warsz.) 2018, 66, 113–123. [Google Scholar] [CrossRef]
- Schupp, J.; Krebs, F.K.; Zimmer, N.; Trzeciak, E.; Schuppan, D.; Tuettenberg, A. Targeting myeloid cells in the tumor sustaining microenvironment. Cell. Immunol. 2019, 343, 103713. [Google Scholar] [CrossRef]
- Jeske, S.S.; Brand, M.; Ziebart, A.; Laban, S.; Doescher, J.; Greve, J.; Jackson, E.K.; Hoffmann, T.K.; Brunner, C.; Schuler, P.J. Adenosine-producing regulatory B cells in head and neck cancer. Cancer Immunol. Immunother. 2020, 69, 1205–1216. [Google Scholar] [CrossRef] [Green Version]
- Kim, S. Animal models of cancer in the head and neck region. Clin. Exp. Otorhinolaryngol. 2009, 2, 55–60. [Google Scholar] [CrossRef]
- Vahle, A.K.; Kerem, A.; Öztürk, E.; Bankfalvi, A.; Lang, S.; Brandau, S. Optimization of an orthotopic murine model of head and neck squamous cell carcinoma in fully immunocompetent mice - Role of toll-like-receptor 4 expressed on host cells. Cancer Lett. 2012, 317, 199–206. [Google Scholar] [CrossRef]
- O’Malley, B.W.; Cope, K.A.; Johnson, C.S.; Schwartz, M.R. A new immunocompetent murine model for oral cancer. Arch. Otolaryngol.-Head Neck Surg. 1997, 123, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Cannon, T.Y.; Guttridge, D.; Dahlman, J.; George, J.R.; Lai, V.; Shores, C.; Bůžková, P.; Couch, M.E. The effect of altered toll-like receptor 4 signaling on cancer cachexia. Arch. Otolaryngol. Head Neck Surg. 2007, 133, 1263–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, K.K.; Rayner, P.A.; Lamx, K.N. Modification of the effects of continuous low dose rate irradiation by concurrent chemotherapy infusion. Int. J. Radiat. Oncol. Biol. Phys. 1984, 10, 1473–1478. [Google Scholar] [CrossRef]
- Méry, B.; Rancoule, C.; Guy, J.B.; Espenel, S.; Wozny, A.S.; Battiston-Montagne, P.; Ardail, D.; Beuve, M.; Alphonse, G.; Rodriguez-Lafrasse, C.; et al. Preclinical models in HNSCC: A comprehensive review. Oral Oncol. 2017, 65, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.P.; Thomas, G.R. Animal models for the study of squamous cell carcinoma of the upper aerodigestive tract: A historical perspective with review of their utility and limitations. Part A. Chemically-inducedde novo cancer, syngeneic animal models of HNSCC, animal models of transplanted xenogeneic human tumors. Int. J. Cancer 2006, 118, 2111–2122. [Google Scholar] [CrossRef] [PubMed]
- Khurana, D.; Martin, E.A.; Kasperbauer, J.L.; O’Malley, B.W.; Salomao, D.R.; Chen, L.; Strome, S.E. Characterization of a spontaneously arising murine squamous cell carcinoma (SCC VII) as a prerequisite for head and neck cancer immunotherapy. Head Neck 2001, 23, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Ohta, A.; Gorelik, E.; Prasad, S.J.; Ronchese, F.; Lukashev, D.; Wong, M.K.K.; Huang, X.; Caldwell, S.; Liu, K.; Smith, P.; et al. A2A adenosine receptor protects tumors from antitumor T cells. Proc. Natl. Acad. Sci. USA 2006, 103, 13132–13137. [Google Scholar] [CrossRef] [Green Version]
- Nelson, B.H. CD20 + B Cells: The Other Tumor-Infiltrating Lymphocytes. J. Immunol. 2010, 185, 4977–4982. [Google Scholar] [CrossRef] [Green Version]
- Carragher, D.M.; Rangel-Moreno, J.; Randall, T.D. Ectopic lymphoid tissues and local immunity. Semin. Immunol. 2008, 20, 26–42. [Google Scholar] [CrossRef] [Green Version]
- DiLillo, D.J.; Yanaba, K.; Tedder, T.F. B Cells Are Required for Optimal CD4 + and CD8 + T Cell Tumor Immunity: Therapeutic B Cell Depletion Enhances B16 Melanoma Growth in Mice. J. Immunol. 2010, 184, 4006–4016. [Google Scholar] [CrossRef] [Green Version]
- Badoual, C.; Hans, S.; Merillon, N.; Van Ryswick, C.; Ravel, P.; Benhamouda, N.; Levionnois, E.; Nizard, M.; Si-Mohamed, A.; Besnier, N.; et al. PD-1-expressing tumor-infiltrating T cells are a favorable prognostic biomarker in HPV-Associated head and neck cancer. Cancer Res. 2013, 73, 128–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badoual, C.; Hans, S.; Rodriguez, J.; Peyrard, S.; Klein, C.; Agueznay, N.E.H.; Mosseri, V.; Laccourreye, O.; Bruneval, P.; Fridman, W.H.; et al. Prognostic value of tumor-infiltrating CD4+ T-cell subpopulations in head and neck cancers. Clin. Cancer Res. 2006, 12, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Chikamatsu, K.; Albers, A.; Stanson, J.; Kwok, W.W.; Appella, E.; Whiteside, T.L.; DeLeo, A.B. p53110-124-specific human CD4+ T-helper cells enhance in vitro generation and antitumor function of tumor-reactive CD8+ T cells. Cancer Res. 2003, 63, 3675–3681. [Google Scholar] [PubMed]
- Punt, S.; Dronkers, E.A.C.; Welters, M.J.P.; Goedemans, R.; Koljenović, S.; Bloemena, E.; Snijders, P.J.F.; Gorter, A.; van der Burg, S.H.; de Jong, R.J.B.; et al. A beneficial tumor microenvironment in oropharyngeal squamous cell carcinoma is characterized by a high T cell and low IL-17+ cell frequency. Cancer Immunol. Immunother. 2016, 65, 393–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, C.; Strauss, L.; Wang, Y.; Szczepanski, M.J.; Lang, S.; Johnson, J.T.; Whiteside, T.L. T regulatory type 1 cells in squamous cell carcinoma of the head and neck: Mechanisms of suppression and expansion in advanced disease. Clin. Cancer Res. 2008, 14, 3706–3715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobie, J.J.; Shah, P.R.; Yang, L.; Rebhahn, J.A.; Fowell, D.J.; Mosmann, T.R. T Regulatory and Primed Uncommitted CD4 T Cells Express CD73, Which Suppresses Effector CD4 T Cells by Converting 5′-Adenosine Monophosphate to Adenosine. J. Immunol. 2006, 177, 6780–6786. [Google Scholar] [CrossRef]
- Tanaka, Y.; Eda, H.; Tanaka, T.; Udagawa, T.; Ishikawa, T.; Horii, I.; Ishitsuka, H.; Kataoka, T.; Taguchi, T. Experimental cancer cachexia induced by transplantable colon 26 adenocarcinoma in mice. Cancer Res. 1990, 50, 2290–2295. [Google Scholar]
- Flier, J.S.; Underhill, L.H.; Dvorak, H.F. Tumors: Wounds That Do Not Heal. N. Engl. J. Med. 1986, 315, 1650–1659. [Google Scholar] [CrossRef]
- van de Veen, W.; Stanic, B.; Wirz, O.F.; Jansen, K.; Globinska, A.; Akdis, M. Role of regulatory B cells in immune tolerance to allergens and beyond. J. Allergy Clin. Immunol. 2016, 138, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, S.; Bhattacharya-Chatterjee, M.; O’Malley, B.W.; Chatterjee, S.K. Inhibition of NK Cell Activity through TGF-β1 by Down-Regulation of NKG2D in a Murine Model of Head and Neck Cancer. J. Immunol. 2005, 175, 5541–5550. [Google Scholar] [CrossRef]
- Matsui, M.; Kishida, T.; Nakano, H.; Yoshimoto, K.; Shin-Ya, M.; Shimada, T.; Nakai, S.; Imanishi, J.; Yoshimoto, T.; Hisa, Y.; et al. Interleukin-27 activates natural killer cells and suppresses NK-resistant head and neck squamous cell carcinoma through inducing antibody-dependent cellular cytotoxicity. Cancer Res. 2009, 69, 2523–2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egenolf, D.D.; Rafferty, P.; Brosnan, K.; Walker, M.; Jordan, J.; Makropoulos, D.; Kavalkovich, K.; Watson, S.; Johns, L.; Volk, A.; et al. Development of a murine model of lymph node metastases suitable for immunotoxicity studies. J. Pharmacol. Toxicol. Methods 2011, 63, 236–249. [Google Scholar] [CrossRef]
- Schreiber, R.D.; Old, L.J.; Smyth, M.J. Cancer immunoediting: Integrating immunity’s roles in cancer suppression and promotion. Science 2011, 331, 1565–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuss, I.; Hathaway, B.; Ferris, R.L.; Gooding, W.; Whiteside, T.L. Decreased absolute counts of T lymphocyte subsets and their relation to disease in squamous cell carcinoma of the head and neck. Clin. Cancer Res. 2004, 10, 3755–3762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gastman, B.R.; Yin, X.M.; Johnson, D.E.; Wieckowski, E.; Wang, G.Q.; Watkins, S.C.; Rabinowich, H. Tumor-induced apoptosis of T cells: Amplification by a mitochondrial cascade. Cancer Res. 2000, 60, 6811–6817. [Google Scholar] [PubMed]
- Szczepanski, M.J.; Czystowska, M.; Szajnik, M.; Harasymczuk, M.; Boyiadzis, M.; Kruk-Zagajewska, A.; Szyfter, W.; Zeromski, J.; Whiteside, T.L. Triggering of toll-like receptor 4 expressed on human head and neck squamous cell carcinoma promotes tumor development and protects the tumor from immune attack. Cancer Res. 2009, 69, 3105–3113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varilla, V.; Atienza, J.; Dasanu, C.A. Immune alterations and immunotherapy prospects in head and neck cancer. Expert Opin. Biol. Ther. 2013, 13, 1241–1256. [Google Scholar] [CrossRef]
- Yilmaz, T.; Gedikoglu, G.; Çelik, A.; Önerci, M.; Turan, E. Prognostic significance of Langerhans cell infiltration in cancer of the larynx. Otolaryngol. Head Neck Surg. 2005, 132, 309–316. [Google Scholar] [CrossRef]
- Albuquerque, R.L.C.; Miguel, M.C.C.; Costa, A.L.L.; Souza, L.B. Correlation of c-erbB-2 and S-100 expression with the malignancy grading and anatomical site in oral squamous cell carcinoma. Int. J. Exp. Pathol. 2003, 84, 259–265. [Google Scholar] [CrossRef]
- Valladeau, J.; Saeland, S. Cutaneous dendritic cells. Semin. Immunol. 2005, 17, 273–283. [Google Scholar] [CrossRef]
- Coussens, L.M.; Tinkle, C.L.; Hanahan, D.; Werb, Z. MMP-9 supplied by bone marrow-derived cells contributes to skin carcinogenesis. Cell 2000, 103, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Salven, P.; Heikkilä, P.; Anttonen, A.; Kajanti, M.; Joensuu, H. Vascular endothelial growth factor in squamous cell head and neck carcinoma: Expression and prognostic significance. Mod. Pathol. 1997, 10, 1128–1133. [Google Scholar] [PubMed]
- Pretscher, D.; Distel, L.V.; Grabenbauer, G.G.; Wittlinger, M.; Buettner, M.; Niedobitek, G. Distribution of immune cells in head and neck cancer: CD8+T-cells and CD20+B-cells in metastatic lymph nodes are associated with favourable outcome in patients with oro- and hypopharyngeal carcinoma. BMC Cancer 2009, 9, 292. [Google Scholar] [CrossRef] [Green Version]
- Distel, L.V.; Fickenscher, R.; Dietel, K.; Hung, A.; Iro, H.; Zenk, J.; Nkenke, E.; Büttner, M.; Niedobitek, G.; Grabenbauer, G.G. Tumour infiltrating lymphocytes in squamous cell carcinoma of the oro- and hypopharynx: Prognostic impact may depend on type of treatment and stage of disease. Oral Oncol. 2009, 45, e167–e174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L.; Li, J.; Mo, H.Y.; Qiu, F.; Zheng, L.M.; Qian, C.N.; Zeng, Y.X. Different subsets of tumor infiltrating lymphocytes correlate with NPC progression in different ways. Mol. Cancer 2010, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Gunderson, A.J.; Kaneda, M.M.; Tsujikawa, T.; Nguyen, A.V.; Affara, N.I.; Ruffell, B.; Gorjestani, S.; Liudahl, S.M.; Truit, M.; Olson, P.; et al. Bruton tyrosine kinase–Dependent immune cell cross-talk drives pancreas cancer. Cancer Discov. 2016, 6, 270–285. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, A.; Okada, H.; Jiang, A.; Kurosaki, M.; Greenberg, S.; Clark, E.A.; Kurosaki, T. Involvement of guanosine triphosphatases and phospholipase C-γ2 in extracellular signal-regulated kinase, c-Jun NH2-terminal kinase, and p38 mitogen-activated protein kinase activation by the B cell antigen receptor. J. Exp. Med. 1998, 188, 1287–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brand, M.; Laban, S.; Theodoraki, M.-N.; Doescher, J.; Hoffmann, T.K.; Schuler, P.J.; Brunner, C. Characterization and Differentiation of the Tumor Microenvironment (TME) of Orthotopic and Subcutaneously Grown Head and Neck Squamous Cell Carcinoma (HNSCC) in Immunocompetent Mice. Int. J. Mol. Sci. 2021, 22, 247. https://doi.org/10.3390/ijms22010247
Brand M, Laban S, Theodoraki M-N, Doescher J, Hoffmann TK, Schuler PJ, Brunner C. Characterization and Differentiation of the Tumor Microenvironment (TME) of Orthotopic and Subcutaneously Grown Head and Neck Squamous Cell Carcinoma (HNSCC) in Immunocompetent Mice. International Journal of Molecular Sciences. 2021; 22(1):247. https://doi.org/10.3390/ijms22010247
Chicago/Turabian StyleBrand, Matthias, Simon Laban, Marie-Nicole Theodoraki, Johannes Doescher, Thomas K. Hoffmann, Patrick J. Schuler, and Cornelia Brunner. 2021. "Characterization and Differentiation of the Tumor Microenvironment (TME) of Orthotopic and Subcutaneously Grown Head and Neck Squamous Cell Carcinoma (HNSCC) in Immunocompetent Mice" International Journal of Molecular Sciences 22, no. 1: 247. https://doi.org/10.3390/ijms22010247
APA StyleBrand, M., Laban, S., Theodoraki, M.-N., Doescher, J., Hoffmann, T. K., Schuler, P. J., & Brunner, C. (2021). Characterization and Differentiation of the Tumor Microenvironment (TME) of Orthotopic and Subcutaneously Grown Head and Neck Squamous Cell Carcinoma (HNSCC) in Immunocompetent Mice. International Journal of Molecular Sciences, 22(1), 247. https://doi.org/10.3390/ijms22010247