CaSR-Mediated hBMSCs Activity Modulation: Additional Coupling Mechanism in Bone Remodeling Compartment


 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
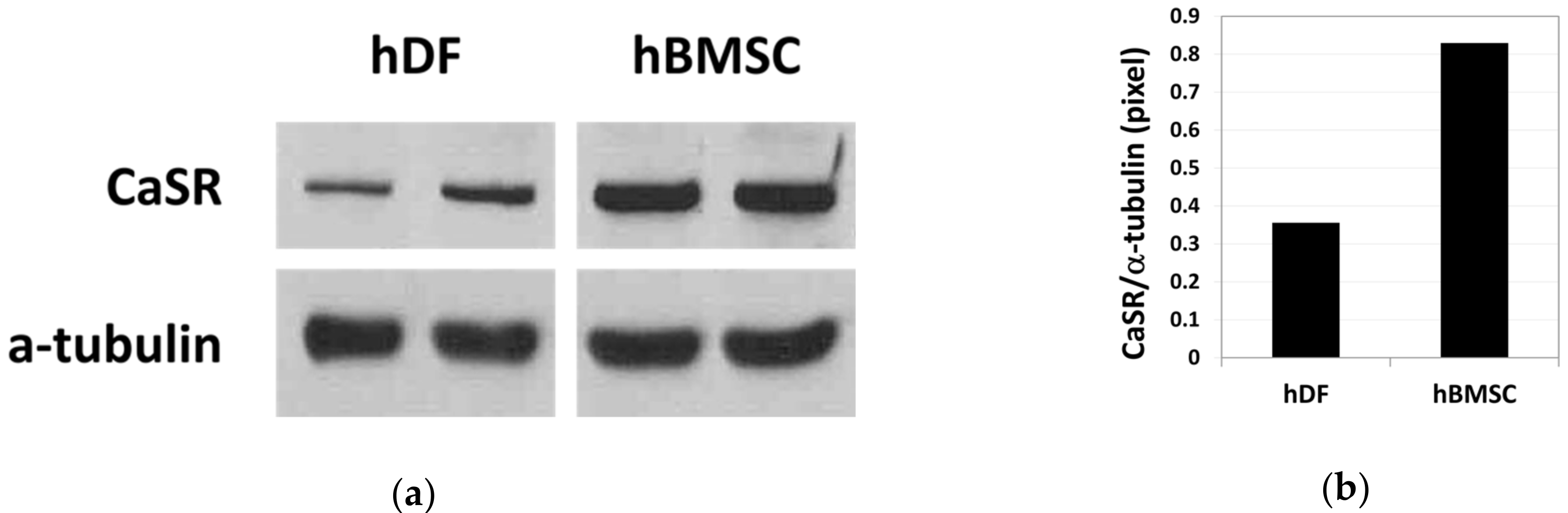
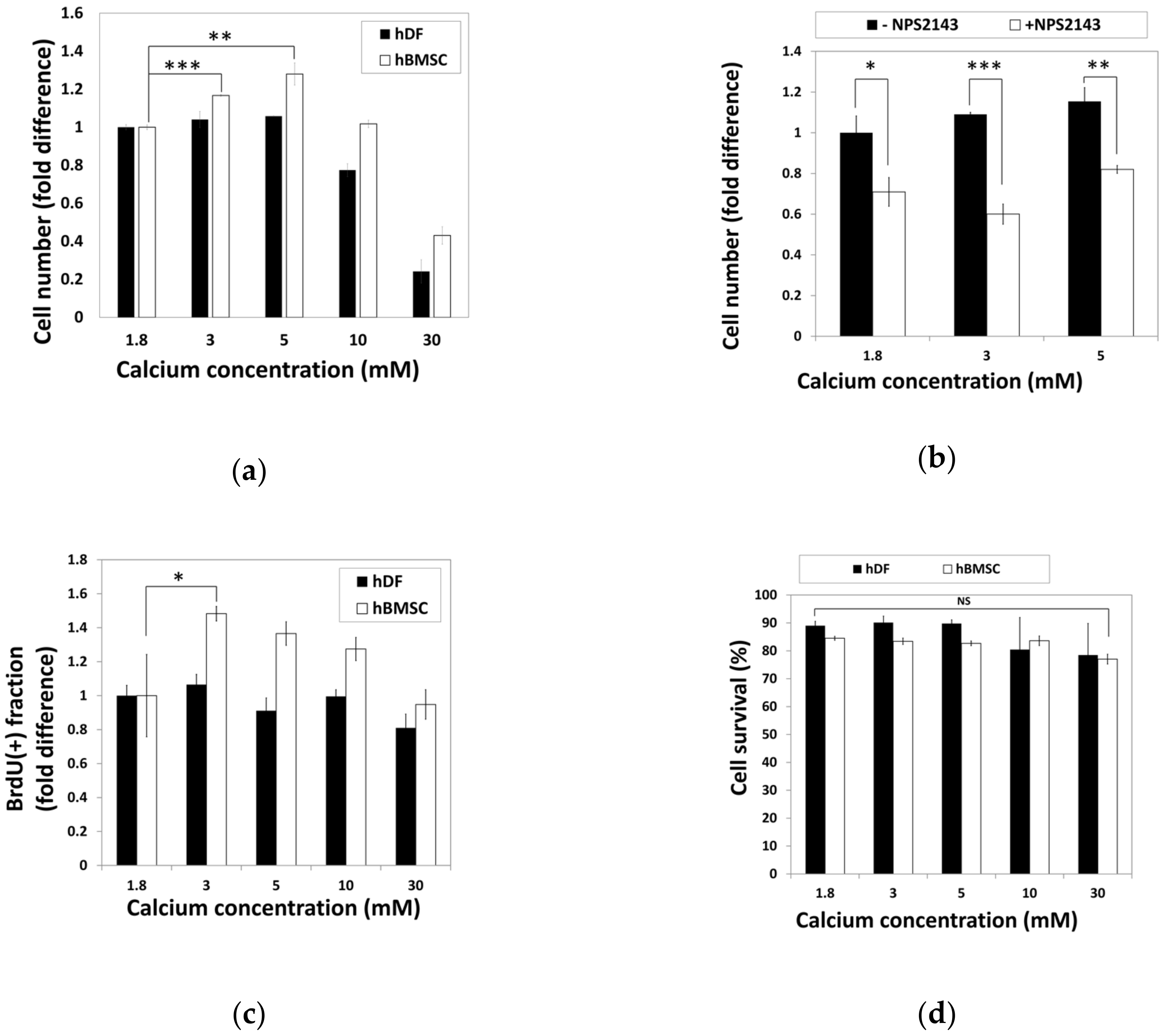
2.1. Increased Proliferation of Human Bone Marrow Stromal Cells (hBMSCs) in Moderate-High Calcium Concentration Is Mediated by Calcium-Sensing Receptor (CaSR)
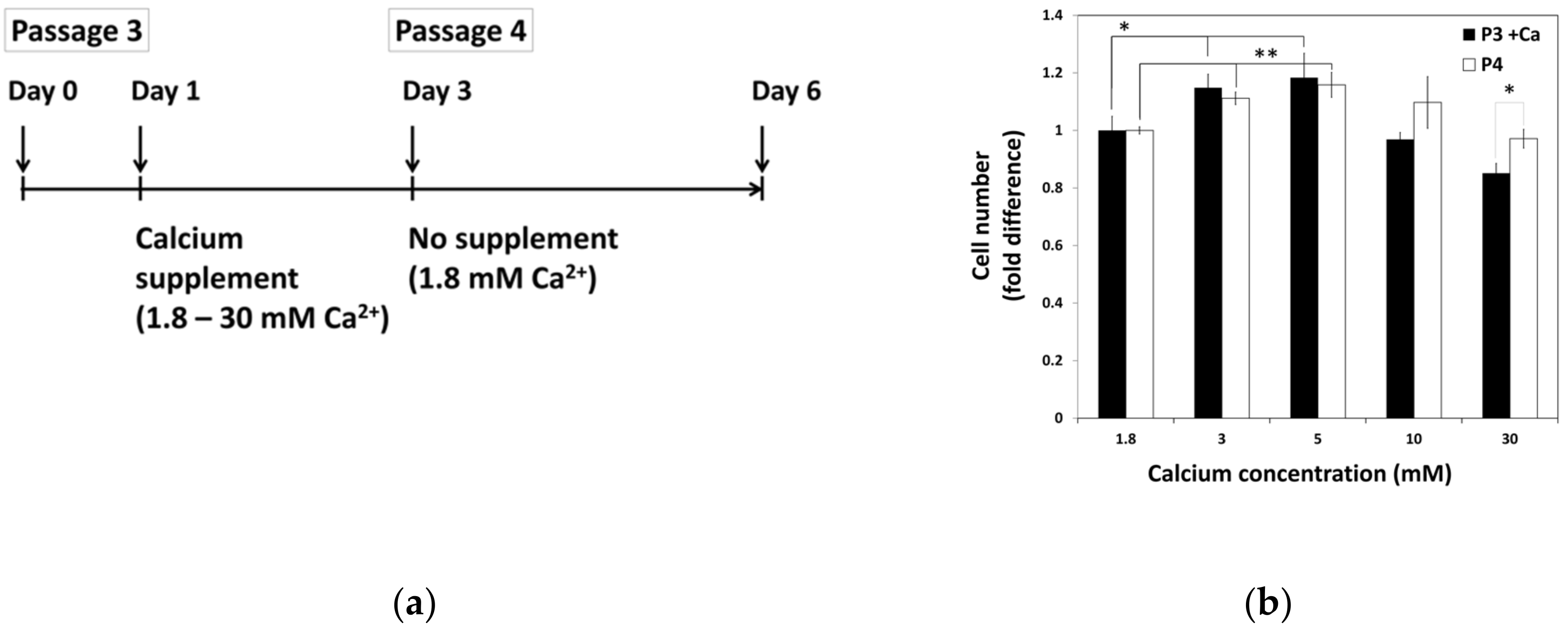
2.2. The Lasting Effect of Moderate-High Calcium Concentration on Cell Proliferation of hBMSCs
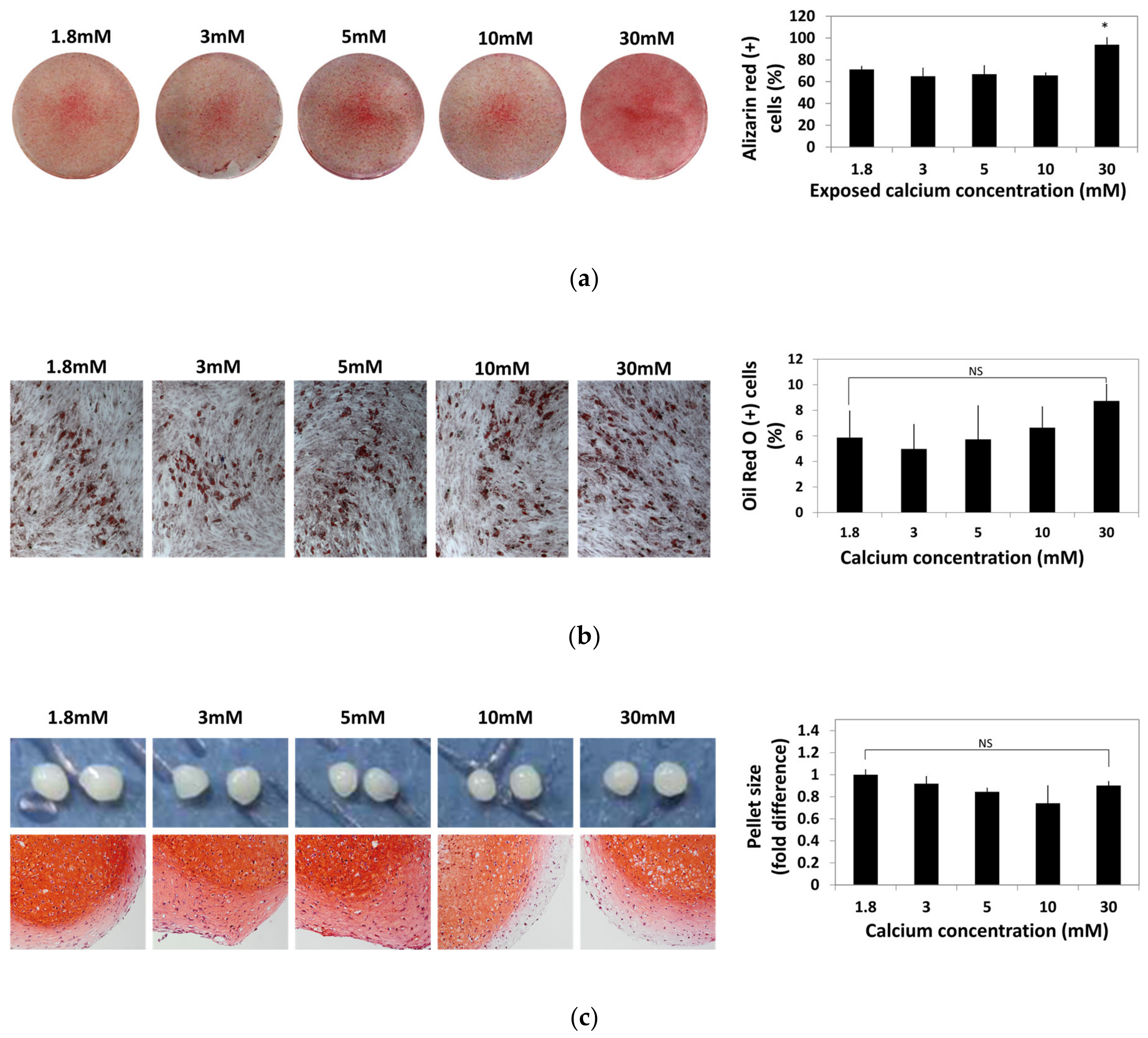
2.3. Influence of Various Calcium Concentrations on Multi-Lineage Differentiation Potentials of hBMSCs
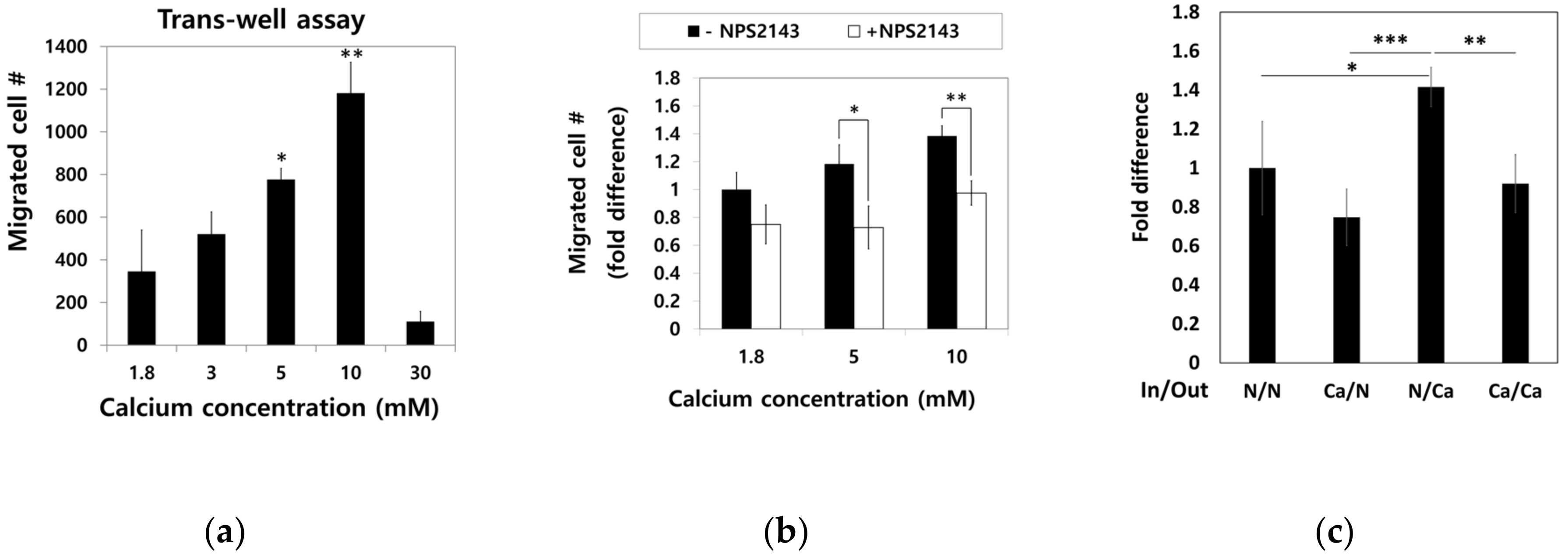
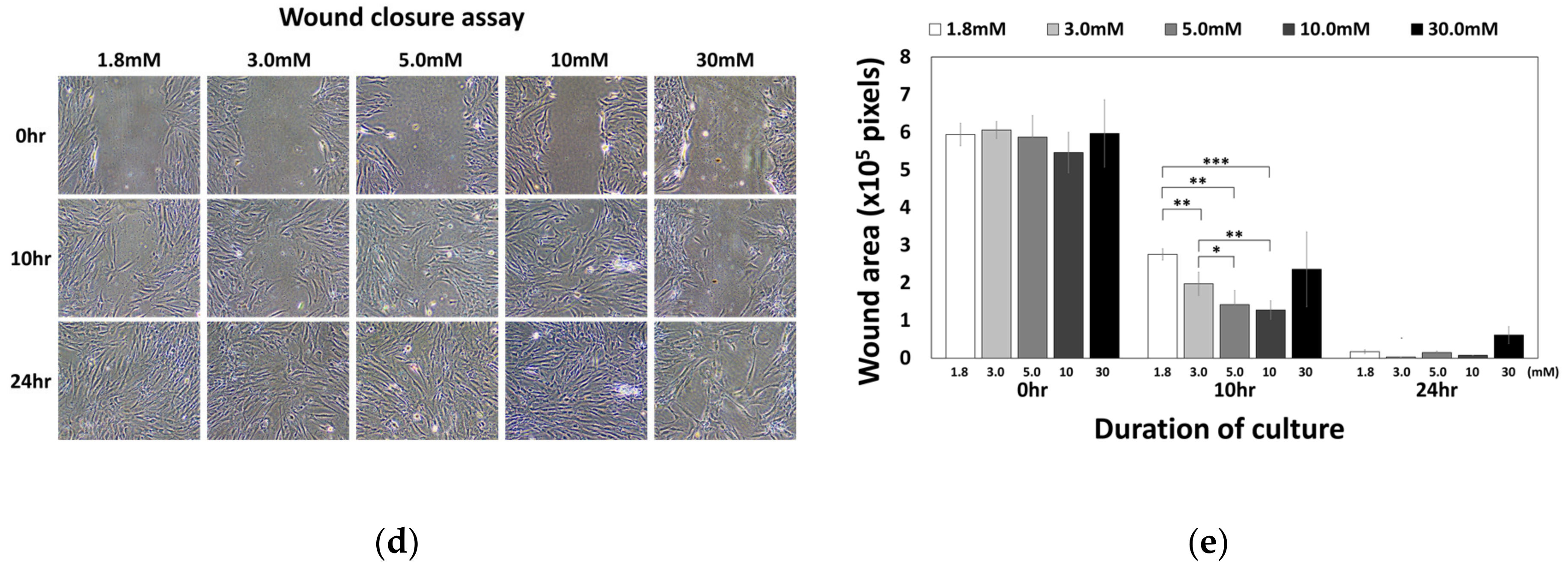
2.4. Moderate-High Calcium Concentration Induced Chemotactic Migration of hBMSCs toward Higher Calcium Concentration Gradient through CaSR
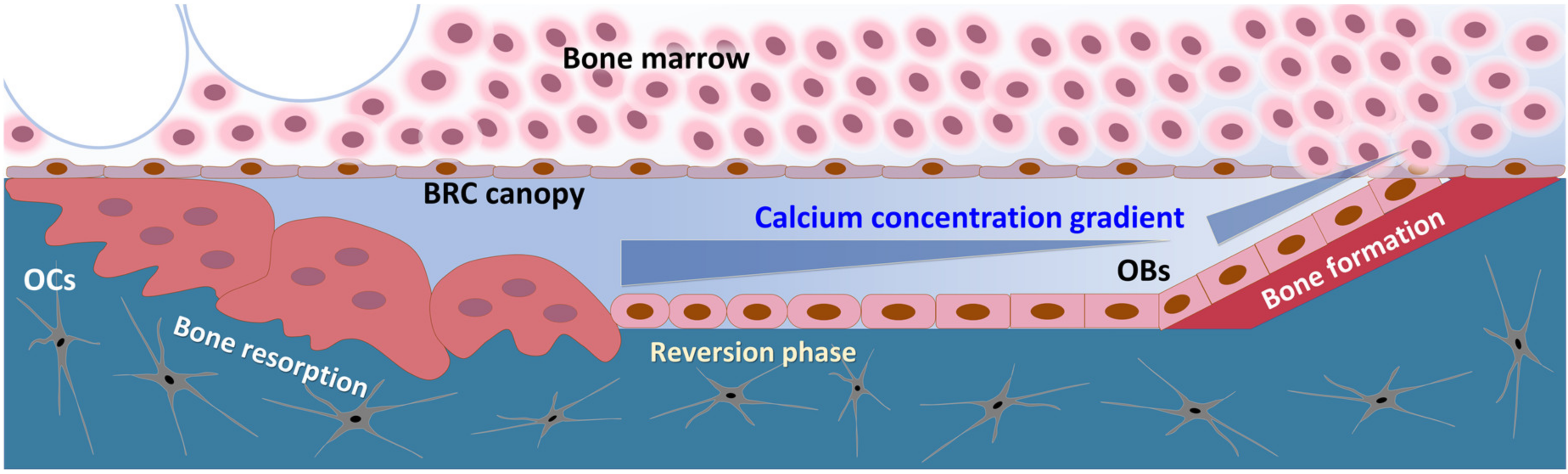
3. Discussion
4. Materials and Methods
4.1. Primary Culture of Bone Marrow Stromal Cells (BMSCs) from Human Bone Marrow Aspirate
4.2. Primary Culture of Human Dermal Fibroblasts (hDFs) from Human Foreskin Tissue
4.3. Western Blot Analysis
4.4. Calcium Chloride and NPS2143 Hydrochloride Treatment
4.5. MTT (Thiazolyl Blue Tetrazolium Bromide) Assay
4.6. BrdU (5-Bromo-2′-Deoxy-Uridine) Incorporation Assay
4.7. Propidium Iodide-Incorporated Apoptotic Assay
4.8. In Vitro Multi-Lineage Differentiation Induction
4.9. Migration Assay
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BM | Bone marrow |
| BRC | Bone remodeling compartment |
| Ca2+i | Intracellular calcium |
| Ca2+o | Extracellular calcium |
| CaSR | Calcium-sensing receptor |
| GAGs | Glycosaminoglycans |
| hBMSCs | Human bone marrow stromal cells |
| hDFs | Human dermal fibroblasts |
| HSCs | Hematopoietic stem cells |
| IL | Interleukin |
| OB | Osteoblast |
| OC | Osteoclast |
| OPG | Osteoprotegerin |
| OPN | Osteopontin |
| PFA | Paraformadehyde |
| PI | Propidium iodide |
| RANKL | Receptor activator of nuclear factor kB ligand |
| TGF-β | Transforming growth factor-beta |
| TNF-α | Tumor necrosis factor-alpha |
References
- Brown, E.M. Clinical lessons from the calcium-sensing receptor. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 122–133. [Google Scholar] [CrossRef]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium signaling: Dynamics, homeostasis and remodeling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef]
- Hofer, A.M.; Gerbino, A.; Caroppo, R.; Curci, S. The extracellular calcium-sensing receptor and cell–cell signaling in epithelia. Cell Calcium 2004, 35, 297–306. [Google Scholar] [CrossRef]
- Magno, A.L.; Ward, B.K.; Ratajczak, T. The calcium-sensing receptor: A molecular perspective. Endocr. Rev. 2011, 32, 3–30. [Google Scholar] [CrossRef]
- Brown, E.M.; Gamba, G.; Riccardi, D.; Lombardi, M.; Butters, R.; Kifor, O.; Sun, A.; Heduger, M.A.; Lytton, J.; Hebert, S.C. Cloning and characterization of an extracellular Ca2+-sensing receptor from bovine parathyroid. Nature 1993, 366, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Riccardi, D.; Hall, A.E.; Chattopadhyay, N.; Xu, J.Z.; Brown, E.M.; Hebert, S.C. Localization of the extracellular Ca2+/polyvalent cation-sensing protein in rat kidney. Am. J. Physiol. 1998, 274, F611–F622. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, N.; Cheng, I.; Rogers, K.; Riccardi, D.; Hall, A.; Diaz, R.; Hebert, S.C.; Soybel, D.I.; Brown, E.M. Identification and localization of extracellular Ca2+-sensing receptor in rat intestine. Am. J. Physiol. 1998, 274, G122–G130. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.; Tu, C.; Chen, T.H.; Komuves, L.; Oda, Y.; Pratt, S.A.; Miller, S.; Shoback, D. Expression and signal transduction of calcium-sensing receptors in cartilage and bone. Endocrinology 1999, 140, 5883–5893. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.; Conner, D.A.; Pollak, M.R.; Ladd, D.J.; Kifor, O.; Warren, H.B.; Brown, E.M.; Seidman, J.G.; Seidman, C.E. A mouse model of human familial hypocalciuric hypercalcemia and neonatal severe hyperparathyroidism. Nat. Genet. 1995, 11, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Yamauchi, M.; Sugimoto, T.; Chauhan, D.; Anderson, K.C.; Brown, E.M.; Chihara, K. The extracellular calcium (Ca2+o)-sensing receptor is expressed in myeloma cells and modulates cell proliferation. Biochem. Biophys. Res. Commun. 2002, 299, 532–538. [Google Scholar] [CrossRef]
- McNeil, S.E.; Hobson, S.A.; Nipper, V.; Rodland, K.D. Functional calcium-sensing receptors in rat fibroblasts are required for activation of SRC kinase and mitogen-activated protein kinase in response to extracellular calcium. J. Biol. Chem. 1998, 273, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Godwin, S.L.; Soltoff, S.P. Extracellular calcium and platelet-derived growth factor promote receptor-mediated chemotaxis in osteoblasts through different signaling pathways. J. Biol. Chem. 1997, 272, 11307–11312. [Google Scholar] [CrossRef] [PubMed]
- Mentaverri, R.; Yano, S.; Chattopadhyay, N.; Petit, L.; Kifor, O.; Kamel, S.; Terwilliger, E.F.; Brazier, M.; Brown, E.M. The calcium sensing receptor is directly involved in both osteoclast differentiation and apoptosis. FASEB J. 2006, 20, 2562–2564. [Google Scholar] [CrossRef] [PubMed]
- Hadjidakis, D.J.; Androulakis, I.I. Bone remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef]
- Silver, I.A.; Murrills, R.J.; Etherington, D.J. Microelectrode studies on the acid microenvironment beneath adherent macrophages and osteoclasts. Exp. Cell Res. 1988, 175, 266–276. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Chattopadhyay, N.; Kifor, O.; Butters JR, R.R.; Sugimoto, T.; Brown, E.M. Mouse Osteoblastic Cell Line (MC3T3-E1) Expresses Extracellular Calcium (Ca2+o)–Sensing Receptor and Its Agonists Stimulate Chemotaxis and Proliferation of MC3T3-E1 Cells. J. Bone Miner. Res. 1998, 13, 1530–1538. [Google Scholar] [CrossRef]
- Dvorak, M.M.; Siddiqua, A.; Ward, D.T.; Carter, D.H.; Dallas, S.L.; Nemeth, E.F.; Riccardi, D. Physiological changes in extracellular calcium concentration directly control osteoblast function in the absence of calciotropic hormones. Proc. Natl. Acad. Sci. USA 2004, 101, 5140–5145. [Google Scholar] [CrossRef]
- Yamauchi, M.; Yamaguchi, T.; Kaji, H.; Sugimoto, T.; Chihara, K. Involvement of calcium-sensing receptor in osteoblastic differentiation of mouse MC3T3-E1 cells. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E608–E616. [Google Scholar] [CrossRef]
- House, M.G.; Kohlmeier, L.; Chattopadhyay, N.; Kifor, O.; Yamaguchi, T.; Leboff, M.S.; Glowacki, J.; Brown, E.M. Expression of an extracellular calcium-sensing receptor in human and mouse bone marrow cells. J. Bone Miner. Res. 1997, 12, 1959–1970. [Google Scholar] [CrossRef]
- Nemeth, E.F.; Delmar, E.G.; Heaton, W.L.; Miller, M.A.; Lambert, L.D.; Conklin, R.L.; Gowen, M.; Gleason, J.G.; Bhatnagar, P.K.; Fox, J. Calcilytic compounds: Potent and selective Ca2+ receptor antagonists that stimulate secretion of parathyroid hormone. J. Pharmacol. Exp. Ther. 2001, 299, 323–331. [Google Scholar]
- Adams, G.B.; Chabner, K.T.; Alley, I.R.; Olson, D.P.; Szczepiorkowski, Z.M.; Poznansky, M.C.; Kos, C.H.; Pollak, M.R.; Brown, E.M.; Scadden, D.T. Stem cell engraftment at the endosteal niche is specified by the calcium-sensing receptor. Nature 2006, 439, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Lam, B.S.; Cunningham, C.; Adams, G.B. Pharmacologic modulation of the calcium-sensing receptor enhances hematopoietic stem cell lodgment in the adult bone marrow. Blood 2011, 117, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Lee, J.; Lee, E.A.; Kwon, Y.S.; Lee, E.; Ahn, W.; Jiang, M.H.; Kim, J.C.; Son, Y. A new role of substance P as an injury-inducible messenger for mobilization of CD29(+) stromal-like cells. Nat. Med. 2009, 15, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Barradas, A.M.C.; Fernandes, H.A.M.; Groen, N.; Chai, Y.C.; Schrooten, J.; van de Peppel, J.; van Leeuven, J.P.T.M.; van Blitterswijk, C.A.; de Boer, J. A calcium-induced signaling cascade leading to osteogenic differentiation of human bone marrow-derived mesenchymal stromal cells. Biomaterials 2012, 33, 3205–3215. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, S.; Hu, Z.; Chen, D.; Wang, J.; Li, Z.; Li, Z.; Cui, H.; Dai, G.; Liu, L.; et al. Aberrant upregulation of CaSR promotes pathological new bone formation in anlkylosing spondylitis. EMBO Mol. Med. 2020, 12, e12109. [Google Scholar] [CrossRef] [PubMed]
- Smajilovic, S.; Hansen, J.L.; Christoffersen, T.E.; Lewin, E.; Sheikh, S.P.; Terwilliger, E.F.; Brown, E.M.; Haunso, S.; Tfelt-Hansen, J. Extracellular calcium sensing in rat aortic vascular smooth muscle cells. Biochem. Biophys. Res. Commun. 2006, 348, 1215–1223. [Google Scholar] [CrossRef]
- Molostvov, G.; Fletcher, S.; Bland, R.; Zehnder, D. Extracellular calcium-sensing receptor mediated signaling is involved in human vascular smooth muscle cell proliferation and apoptosis. Cell. Physiol. Biochem. 2008, 22, 413–422. [Google Scholar] [CrossRef]
- Boudot, C.; Saidak, Z.; Boulanouar, A.K.; Petit, L.; Gouilleux, F.; Massy, Z.; Brazier, M.; Mentaverri, R.; Kamel, S. Implication of the calcium sensing receptor and the Phosphoinositide 3-kinase/Akt pathway in the extracellular calcium-mediated migration of RAW 264.7 osteoclast precursor cells. Bone 2010, 46, 1416–1423. [Google Scholar] [CrossRef]
- Aguirre, A.; González, A.; Planell, J.A.; Engel, E. Extracellular calcium modulates in vitro bone marrow-derived Flk-1+ CD34+ progenitor cell chemotaxis and differentiation through a calcium-sensing receptor. Biochem. Biophys. Res. Commun. 2010, 393, 156–161. [Google Scholar] [CrossRef]
- Lee, M.N.; Hwang, H.S.; Oh, S.H.; Roshanzadeh, A.; Kim, J.W.; Song, J.H.; Kim, E.S.; Koh, J.T. Elevated extracellular calcium ions promote proliferation and migration of mesenchymal stem cells vial increasing osteopontin expression. Exp. Mol. Med. 2018, 50, 142. [Google Scholar] [CrossRef]
- Pacifici, R. Estrogen, cytokines, and pathogenesis of postmenopausal osteoporosis. J. Bone Miner. Res. 1996, 11, 1043–1051. [Google Scholar] [CrossRef] [PubMed]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.L.; Sondergaard, T.E.; Skorzynska, K.E.; Dagnaes-Hansen, F.; Plesner, T.L.; Hauge, E.M.; Plesner, T.; Delaisse, J.M. A physical mechanism for coupling bone resorption and formation in adult human bone. Am. J. Pathol. 2009, 174, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Satel, F.; Chabadel, A.; Zhao, Y.; Lafage-Proust, M.H.; Clezardin, P.; Jurdic, P.; Bonnelye, E. Transmigration: A new property of mature multinucleated osteoclasts. J. Bone Miner. Res. 2006, 21, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.J.; Sims, N.A. Osteoclast-derived activity in the coupling of bone formation to resorption. Trends Mol. Med. 2005, 11, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Parfit, A.M. The mechanism of coupling: A role for the vasculature. Bone 2000, 26, 319–323. [Google Scholar] [CrossRef]
- Jensen, P.R.; Andersen, T.L.; Soe, K.; Hauge, E.M.; Bollerslev, J.; Amling, M.; Barvencik, F.; Delaisse, J.M. Premature loss of bone remodeling compartment canopies is associated with deficient bone formation: A study of healthy individuals and patients with Cushing’s syndrome. J. Bone Miner. Res. 2012, 27, 770–780. [Google Scholar] [CrossRef]
- Andersen, T.L.; Hauge, E.M.; Rolighed, L.; Bollerslev, J.; Kjaersgaard-Andersen, P.; Delaisse, J.M. Correlation between absence of bone remodeling compartment canopies, reversal phase arrest, and deficient bone formation in post-menopausal osteoporosis. Am. J. Pathol. 2014, 184, 1142–1151. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, H.; Lee, J.; Jang, S.; Lee, J.; Oh, T.I.; Son, Y.; Lee, E. CaSR-Mediated hBMSCs Activity Modulation: Additional Coupling Mechanism in Bone Remodeling Compartment. Int. J. Mol. Sci. 2021, 22, 325. https://doi.org/10.3390/ijms22010325
Cho H, Lee J, Jang S, Lee J, Oh TI, Son Y, Lee E. CaSR-Mediated hBMSCs Activity Modulation: Additional Coupling Mechanism in Bone Remodeling Compartment. International Journal of Molecular Sciences. 2021; 22(1):325. https://doi.org/10.3390/ijms22010325
Chicago/Turabian StyleCho, Hyunji, Jisoo Lee, Seoyoung Jang, Jungsun Lee, Tong In Oh, Youngsook Son, and EunAh Lee. 2021. "CaSR-Mediated hBMSCs Activity Modulation: Additional Coupling Mechanism in Bone Remodeling Compartment" International Journal of Molecular Sciences 22, no. 1: 325. https://doi.org/10.3390/ijms22010325
APA StyleCho, H., Lee, J., Jang, S., Lee, J., Oh, T. I., Son, Y., & Lee, E. (2021). CaSR-Mediated hBMSCs Activity Modulation: Additional Coupling Mechanism in Bone Remodeling Compartment. International Journal of Molecular Sciences, 22(1), 325. https://doi.org/10.3390/ijms22010325