The p24 Complex Contributes to Specify Arf1 for COPI Coat Selection

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
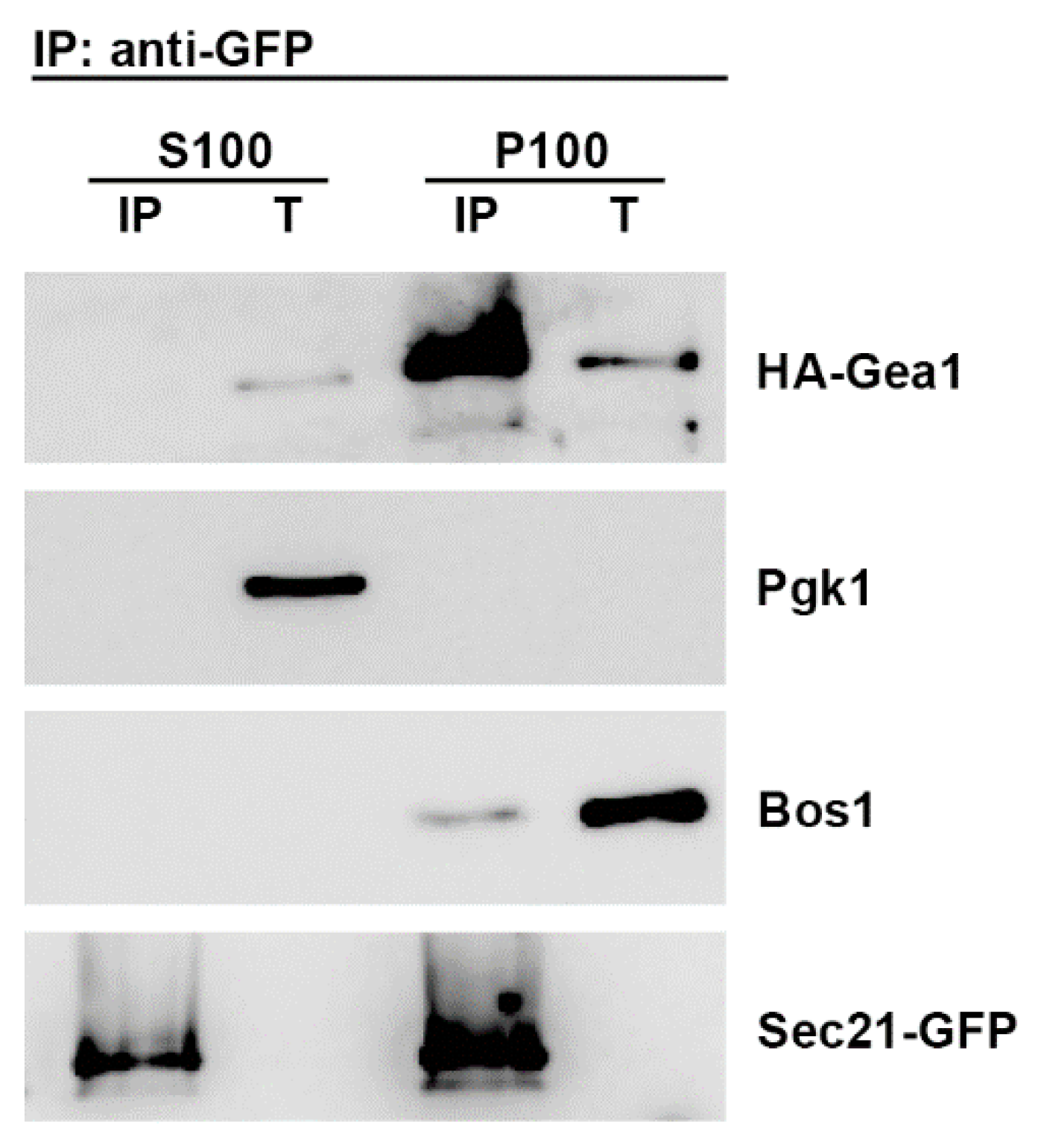
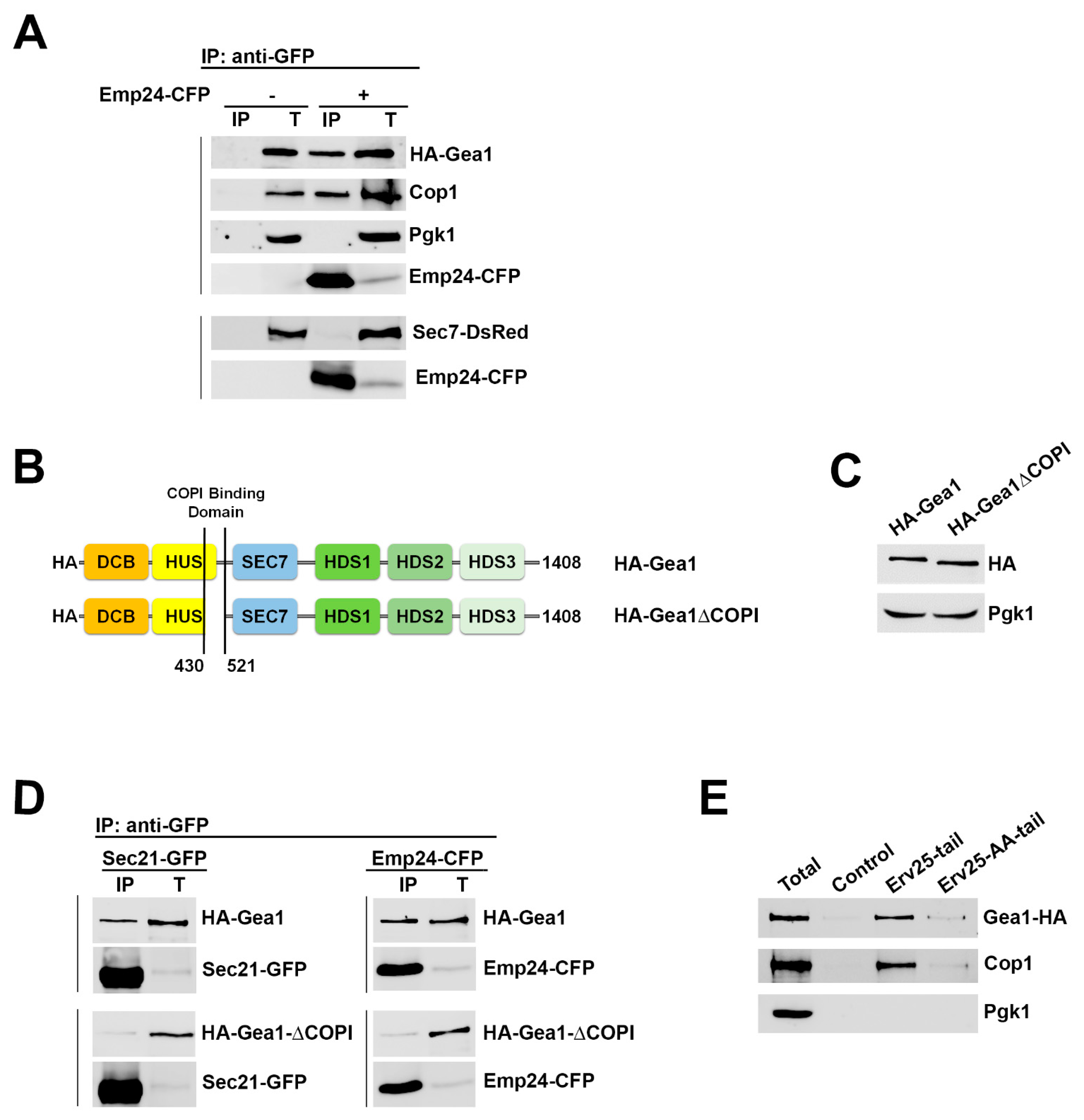
2.1. The p24 Complex Specifically Interacts with the ArfGEF Gea1 Through COPI Coat Binding
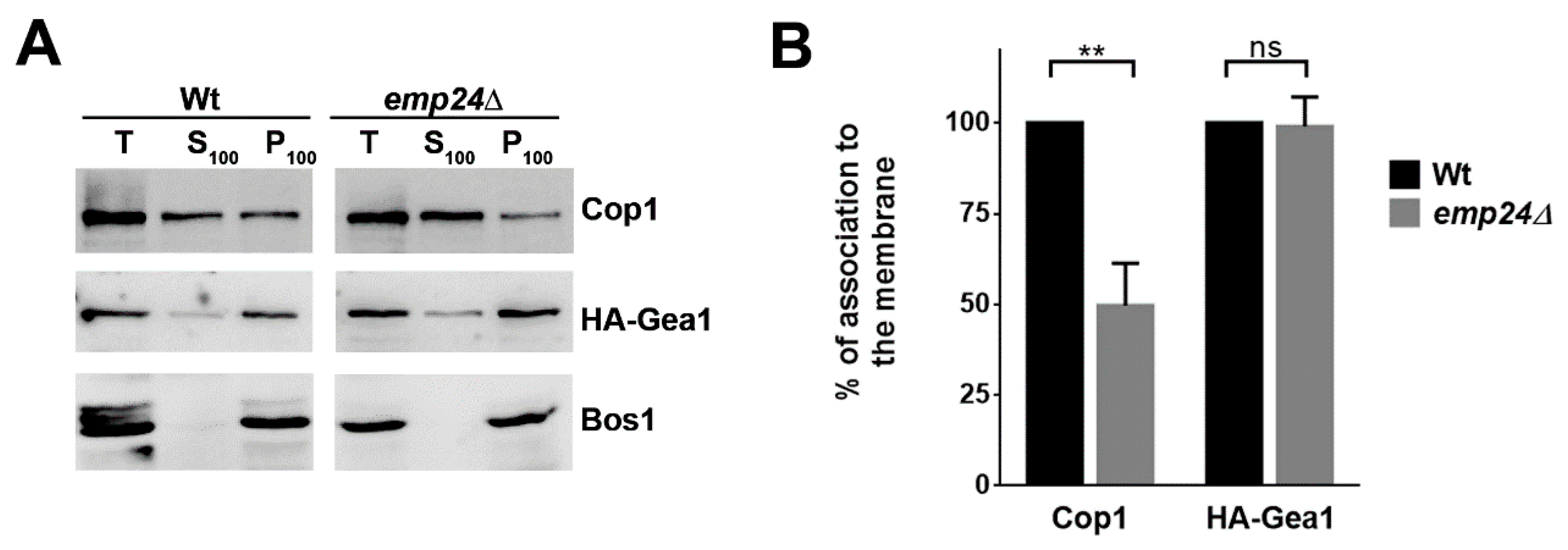
2.2. The Specific Association of the Arf1GEF Gea1 with the COPI Coat is Stabilized by the p24 Complex
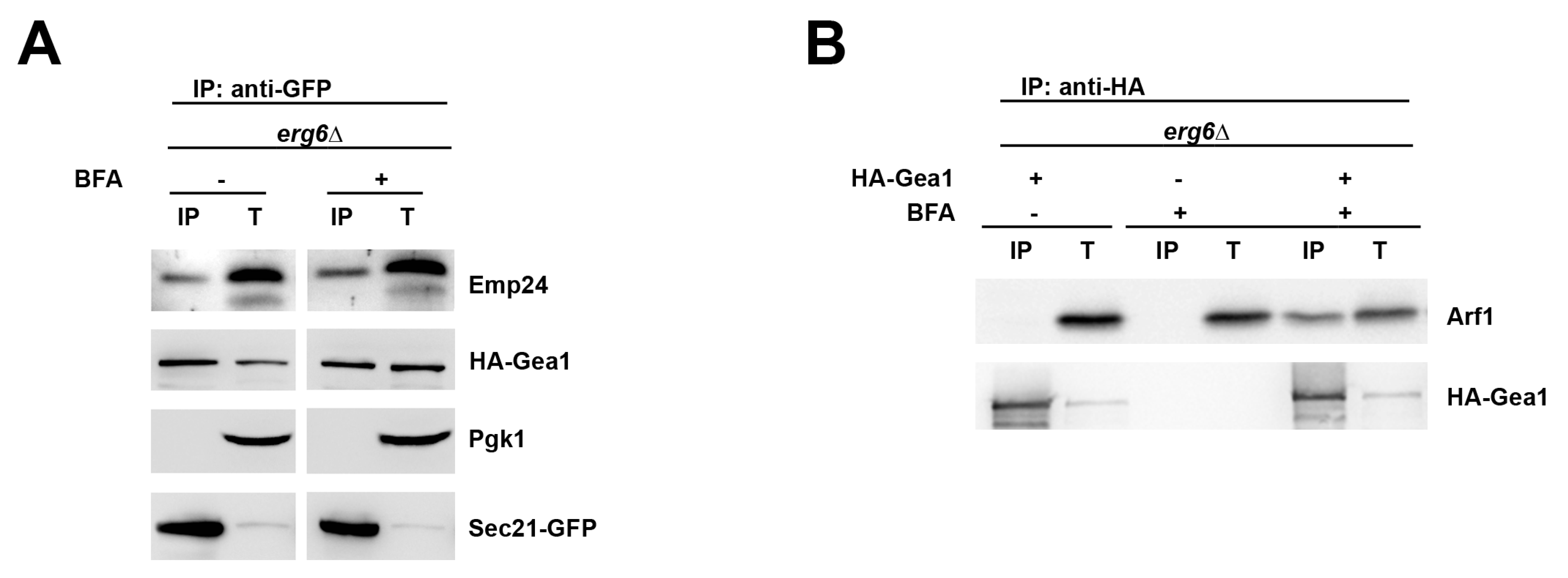
2.3. The p24 Complex Contributes to Specify Arf1 Activation for COPI Binding
3. Discussion
4. Materials and Methods
4.1. Media and Growth Conditions
4.2. Yeast Strains and Plasmids
4.3. Plasmid Construction
4.4. Differential Fractionation
4.5. Native Coimmunoprecipitation
4.6. BFA Treatment
4.7. In Vitro Pull-Down Assay
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GEF | Guanosine Exchange Factor |
| GAP | GTPase Activating Protein |
| COPI | Coat Protein Complex I |
| ARF | ADP-Ribosylation Factor |
| BFA | Brefeldin A |
References
- Barlowe, C.K.; Miller, E.A. Secretory protein biogenesis and traffic in the early secretory pathway. Genetics 2013, 193, 383–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glick, B.S.; Nakano, A. Membrane traffic within the Golgi apparatus. Annu. Rev. Cell Dev. Biol. 2009, 25, 113–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodonova, S.O.; Diestelkoetter-Bachert, P.; von Appen, A.; Hagen, W.J.; Beck, R.; Beck, M.; Wieland, F.; Briggs, J.A. VESICULAR TRANSPORT. A structure of the COPI coat and the role of coat proteins in membrane vesicle assembly. Science 2015, 349, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Letourneur, F.; Gaynor, E.C.; Hennecke, S.; Démollière, C.; Duden, R.; Emr, S.D.; Riezman, H.; Cosson, P. Coatomer is essential for retrieval of dilysine-tagged proteins to the endoplasmic reticulum. Cell 1994, 79, 1199–1207. [Google Scholar] [CrossRef]
- Béthune, J.; Wieland, F.T. Assembly of COPI and COPII Vesicular Coat Proteins on Membranes. Annu. Rev. Biophys. 2018, 47, 63–83. [Google Scholar] [CrossRef]
- Peyroche, A.; Courbeyrette, R.; Rambourg, A.; Jackson, C.L. The ARF exchange factors Gea1p and Gea2p regulate Golgi structure and function in yeast. J. Cell Sci. 2001, 114, 2241–2253. [Google Scholar] [CrossRef] [Green Version]
- Peyroche, A.; Paris, S.; Jackson, C.L. Nucleotide exchange on ARF mediated by yeast Gea1 protein. Nature 1996, 384, 479–481. [Google Scholar] [CrossRef]
- Spang, A.; Herrmann, J.M.; Hamamoto, S.; Schekman, R. The ADP ribosylation factor-nucleotide exchange factors Gea1p and Gea2p have overlapping, but not redundant functions in retrograde transport from the Golgi to the endoplasmic reticulum. Mol. Biol. Cell 2001, 12, 1035–1045. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, M.A.; Fromme, J.C. Regulation of Arf activation occurs via distinct mechanisms at early and late Golgi compartments. Mol. Biol. Cell 2017, 28, 3660–3671. [Google Scholar] [CrossRef]
- Wright, J.; Kahn, R.A.; Sztul, E. Regulating the large Sec7 ARF guanine nucleotide exchange factors: The when, where and how of activation. Cell Mol. Life Sci. 2014, 71, 3419–3438. [Google Scholar] [CrossRef] [Green Version]
- Suda, Y.; Kurokawa, K.; Nakano, A. Regulation of ER-Golgi Transport Dynamics by GTPases in Budding Yeast. Front. Cell Dev. Biol. 2017, 5, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-González, M.; Bravo-Plaza, I.; de Los Ríos, V.; Pinar, M.; Pantazopoulou, A.; Peñalva, M.A. COPI localizes to the early Golgi in Aspergillus nidulans. Fungal Genet. Biol. 2019, 123, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Franzusoff, A.; Redding, K.; Crosby, J.; Fuller, R.S.; Schekman, R. Localization of components involved in protein transport and processing through the yeast Golgi apparatus. J. Cell Biol. 1991, 112, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Richardson, B.C.; McDonold, C.M.; Fromme, J.C. The Sec7 Arf-GEF is recruited to the trans-Golgi network by positive feedback. Dev. Cell 2012, 22, 799–810. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.W.; Nakayama, K. Guanine nucleotide-exchange factors for arf GTPases: Their diverse functions in membrane traffic. J. Biochem. 2004, 136, 761–767. [Google Scholar] [CrossRef]
- Deng, Y.; Golinelli-Cohen, M.P.; Smirnova, E.; Jackson, C.L. A COPI coat subunit interacts directly with an early-Golgi localized Arf exchange factor. EMBO Rep. 2009, 10, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Jackson, C.L. GEF-effector interactions. Cell Logist. 2014, 4, e943616. [Google Scholar] [CrossRef]
- Strating, J.R.; Martens, G.J. The p24 family and selective transport processes at the ER-Golgi interface. Biol. Cell 2009, 101, 495–509. [Google Scholar] [CrossRef]
- Belden, W.J.; Barlowe, C. Distinct roles for the cytoplasmic tail sequences of Emp24p and Erv25p in transport between the endoplasmic reticulum and Golgi complex. J. Biol. Chem. 2001, 276, 43040–43048. [Google Scholar] [CrossRef] [Green Version]
- Pastor-Cantizano, N.; Montesinos, J.C.; Bernat-Silvestre, C.; Marcote, M.J.; Aniento, F. p24 family proteins: Key players in the regulation of trafficking along the secretory pathway. Protoplasma 2016, 253, 967–985. [Google Scholar] [CrossRef]
- Béthune, J.; Kol, M.; Hoffmann, J.; Reckmann, I.; Brügger, B.; Wieland, F. Coatomer, the coat protein of COPI transport vesicles, discriminates endoplasmic reticulum residents from p24 proteins. Mol. Cell Biol. 2006, 26, 8011–8021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzioch, M.; Henthorn, D.C.; Herrmann, J.M.; Wilson, R.; Thomas, D.Y.; Bergeron, J.J.; Solari, R.C.; Rowley, A. Erp1p and Erp2p, partners for Emp24p and Erv25p in a yeast p24 complex. Mol. Biol. Cell 1999, 10, 1923–1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gommel, D.; Orci, L.; Emig, E.M.; Hannah, M.J.; Ravazzola, M.; Nickel, W.; Helms, J.B.; Wieland, F.T.; Sohn, K. p24 and p23, the major transmembrane proteins of COPI-coated transport vesicles, form hetero-oligomeric complexes and cycle between the organelles of the early secretory pathway. FEBS Lett. 1999, 447, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Sohn, K.; Orci, L.; Ravazzola, M.; Amherdt, M.; Bremser, M.; Lottspeich, F.; Fiedler, K.; Helms, J.B.; Wieland, F.T. A major transmembrane protein of Golgi-derived COPI-coated vesicles involved in coatomer binding. J. Cell Biol. 1996, 135, 1239–1248. [Google Scholar] [CrossRef]
- Muñiz, M.; Riezman, H. Trafficking of glycosylphosphatidylinositol anchored proteins from the endoplasmic reticulum to the cell surface. J. Lipid Res. 2016, 57, 352–360. [Google Scholar] [CrossRef] [Green Version]
- Muñiz, M.; Nuoffer, C.; Hauri, H.P.; Riezman, H. The Emp24 complex recruits a specific cargo molecule into endoplasmic reticulum-derived vesicles. J. Cell Biol. 2000, 148, 925–930. [Google Scholar] [CrossRef] [Green Version]
- Aguilera-Romero, A.; Kaminska, J.; Spang, A.; Riezman, H.; Muñiz, M. The yeast p24 complex is required for the formation of COPI retrograde transport vesicles from the Golgi apparatus. J. Cell Biol. 2008, 180, 713–720. [Google Scholar] [CrossRef] [Green Version]
- Bremser, M.; Nickel, W.; Schweikert, M.; Ravazzola, M.; Amherdt, M.; Hughes, C.A.; Söllner, T.H.; Rothman, J.E.; Wieland, F.T. Coupling of coat assembly and vesicle budding to packaging of putative cargo receptors. Cell 1999, 96, 495–506. [Google Scholar] [CrossRef] [Green Version]
- Casanova, J.E. Regulation of Arf activation: The Sec7 family of guanine nucleotide exchange factors. Traffic 2007, 8, 1476–1485. [Google Scholar] [CrossRef]
- Mitrovic, S.; Ben-Tekaya, H.; Koegler, E.; Gruenberg, J.; Hauri, H.P. The cargo receptors Surf4, endoplasmic reticulum-Golgi intermediate compartment (ERGIC)-53, and p25 are required to maintain the architecture of ERGIC and Golgi. Mol. Biol. Cell 2008, 19, 1976–1990. [Google Scholar] [CrossRef] [Green Version]
- Peyroche, A.; Antonny, B.; Robineau, S.; Acker, J.; Cherfils, J.; Jackson, C.L. Brefeldin A acts to stabilize an abortive ARF-GDP-Sec7 domain protein complex: Involvement of specific residues of the Sec7 domain. Mol. Cell 1999, 3, 275–285. [Google Scholar] [CrossRef]
- Mossessova, E.; Corpina, R.A.; Goldberg, J. Crystal structure of ARF1*Sec7 complexed with Brefeldin A and its implications for the guanine nucleotide exchange mechanism. Mol. Cell 2003, 12, 1403–1411. [Google Scholar] [CrossRef]
- Renault, L.; Guibert, B.; Cherfils, J. Structural snapshots of the mechanism and inhibition of a guanine nucleotide exchange factor. Nature 2003, 426, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J. Decoding of sorting signals by coatomer through a GTPase switch in the COPI coat complex. Cell 2000, 100, 671–679. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.M.; Poon, P.P.; Singer, R.A.; Johnston, G.C.; Spang, A. The ArfGAP Glo3 is required for the generation of COPI vesicles. Mol. Biol. Cell 2004, 15, 4064–4072. [Google Scholar] [CrossRef] [Green Version]
- Arakel, E.C.; Huranova, M.; Estrada, A.F.; Rau, E.M.; Spang, A.; Schwappach, B. Dissection of GTPase-activating proteins reveals functional asymmetry in the COPI coat of budding yeast. J. Cell Sci. 2019, 132, jcs232124. [Google Scholar] [CrossRef] [Green Version]
- Richardson, B.C.; Fromme, J.C. Autoregulation of Sec7 Arf-GEF activity and localization by positive feedback. Small GTPases 2012, 3, 240–243. [Google Scholar] [CrossRef] [Green Version]
- Fromme, J.C. Membrane trafficking: Licensing a cargo receptor for ER export. Curr. Biol. 2015, 25, R67–R68. [Google Scholar] [CrossRef] [Green Version]
- McDonold, C.M.; Fromme, J.C. Four GTPases differentially regulate the Sec7 Arf-GEF to direct traffic at the trans-golgi network. Dev. Cell 2014, 30, 759–767. [Google Scholar] [CrossRef] [Green Version]
- Tojima, T.; Suda, Y.; Ishii, M.; Kurokawa, K.; Nakano, A. Spatiotemporal dissection of the trans-Golgi network in budding yeast. J. Cell Sci. 2019, 132, jcs231159. [Google Scholar] [CrossRef] [Green Version]
- Caster, A.H.; Sztul, E.; Kahn, R.A. A role for cargo in Arf-dependent adaptor recruitment. J. Biol. Chem. 2013, 288, 14788–14804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canagarajah, B.J.; Ren, X.; Bonifacino, J.S.; Hurley, J.H. The clathrin adaptor complexes as a paradigm for membrane-associated allostery. Protein Sci. 2013, 22, 517–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, L.L.; Fromme, J.C. Extensive GTPase crosstalk regulates Golgi trafficking and maturation. Curr. Opin. Cell Biol. 2020, 65, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pantazopoulou, A.; Glick, B.S. A Kinetic View of Membrane Traffic Pathways Can Transcend the Classical View of Golgi Compartments. Front. Cell Dev. Biol. 2019, 7, 153. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Cai, H.; Park, S.K.; Menon, S.; Jackson, C.L.; Ferro-Novick, S. Trs65p, a subunit of the Ypt1p GEF TRAPPII, interacts with the Arf1p exchange factor Gea2p to facilitate COPI-mediated vesicle traffic. Mol. Biol. Cell 2011, 22, 3634–3644. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Source |
|---|---|---|
| MMY1156 | MATa SEC21-GFP::HIS3 ura3 leu2 his3 lys2 ade2 trp1 | This study |
| BY4742 | MATα ura3 leu2 his3 lys2 | Euroscarf |
| Y06829 | MATa gea1Δ::kanMx ura3 leu2 his3 met15 | Euroscarf |
| MMY835 | MATa gea1Δ::kanMx emp24Δ::HmBx ura3 leu2 his3 met15 | This study |
| MMY1157 | MATa SEC21-GFP::HIS3 emp24Δ::kanMx ura3 leu2 his3 lys2 trp1 | This study |
| RSY1818 | MATα ura3 leu2 his3 lys2 ade2 | A. Spang |
| MMY914 | MATα emp24Δ::kanMx leu2.3,112 ura3-52 his3Δ200 | This study |
| APY022 | MATα gea2Δ::HIS3 gea1-6 leu2 his3 lys2 ade2 ura3 | A. Spang |
| MMY816 | MATα gea1-6 gea2Δ::HIS3 emp24Δ::HmBx ura3 leu2 his3 ade2 lys2 | This study |
| MMY1320 | MATa sec7-4 trp1 ura3 leu2 his3 ade2 | This study |
| MMY1321 | MATα emp24Δ::kanMx sec7-4 ura3 leu2 his3 ade2 | This study |
| INV-ARFGFP | MATa ARF1-GFP::HIS3 ura3Δ0 leu2Δ0 his3Δ1 met15Δ0 | Invitrogen |
| MMY562 | MATα emp24Δ::KanMx4 ARF1-GFP::HIS3 ura3 leu2 his3 | This study |
| MMY1668 | MATa erg6∆::hph SEC21-GFP::HIS3 | This study |
| Number | Yeast Marker | Yeast Replication | Plasmid | Source |
|---|---|---|---|---|
| pRC4 | URA3 | 2μ | 5xHA-GEA1 | C.L. Jackson |
| pRC4-GEA1-ΔCOPI | URA3 | 2μ | pRC4-5xHA-GEA1-ΔCOPI | This study |
| RH3127 | URA3 | CEN | EMP24-CFP (YCPlac33) | G. Castillón |
| pRS415 | LEU2 | CEN | SEC7-dsRED | A. Spang |
| pYD111 | URA3 | 2μ | YEp352-5xHA GEA1 (5-611) | C.L. Jackson |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabido-Bozo, S.; Perez-Linero, A.M.; Manzano-Lopez, J.; Rodriguez-Gallardo, S.; Aguilera-Romero, A.; Cortes-Gomez, A.; Lopez, S.; Wellinger, R.E.; Muñiz, M. The p24 Complex Contributes to Specify Arf1 for COPI Coat Selection. Int. J. Mol. Sci. 2021, 22, 423. https://doi.org/10.3390/ijms22010423
Sabido-Bozo S, Perez-Linero AM, Manzano-Lopez J, Rodriguez-Gallardo S, Aguilera-Romero A, Cortes-Gomez A, Lopez S, Wellinger RE, Muñiz M. The p24 Complex Contributes to Specify Arf1 for COPI Coat Selection. International Journal of Molecular Sciences. 2021; 22(1):423. https://doi.org/10.3390/ijms22010423
Chicago/Turabian StyleSabido-Bozo, Susana, Ana Maria Perez-Linero, Javier Manzano-Lopez, Sofia Rodriguez-Gallardo, Auxiliadora Aguilera-Romero, Alejandro Cortes-Gomez, Sergio Lopez, Ralf Erik Wellinger, and Manuel Muñiz. 2021. "The p24 Complex Contributes to Specify Arf1 for COPI Coat Selection" International Journal of Molecular Sciences 22, no. 1: 423. https://doi.org/10.3390/ijms22010423
APA StyleSabido-Bozo, S., Perez-Linero, A. M., Manzano-Lopez, J., Rodriguez-Gallardo, S., Aguilera-Romero, A., Cortes-Gomez, A., Lopez, S., Wellinger, R. E., & Muñiz, M. (2021). The p24 Complex Contributes to Specify Arf1 for COPI Coat Selection. International Journal of Molecular Sciences, 22(1), 423. https://doi.org/10.3390/ijms22010423