
Alternative C3 Complement System: Lipids and Atherosclerosis

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Characteristics of the FH Patient Population
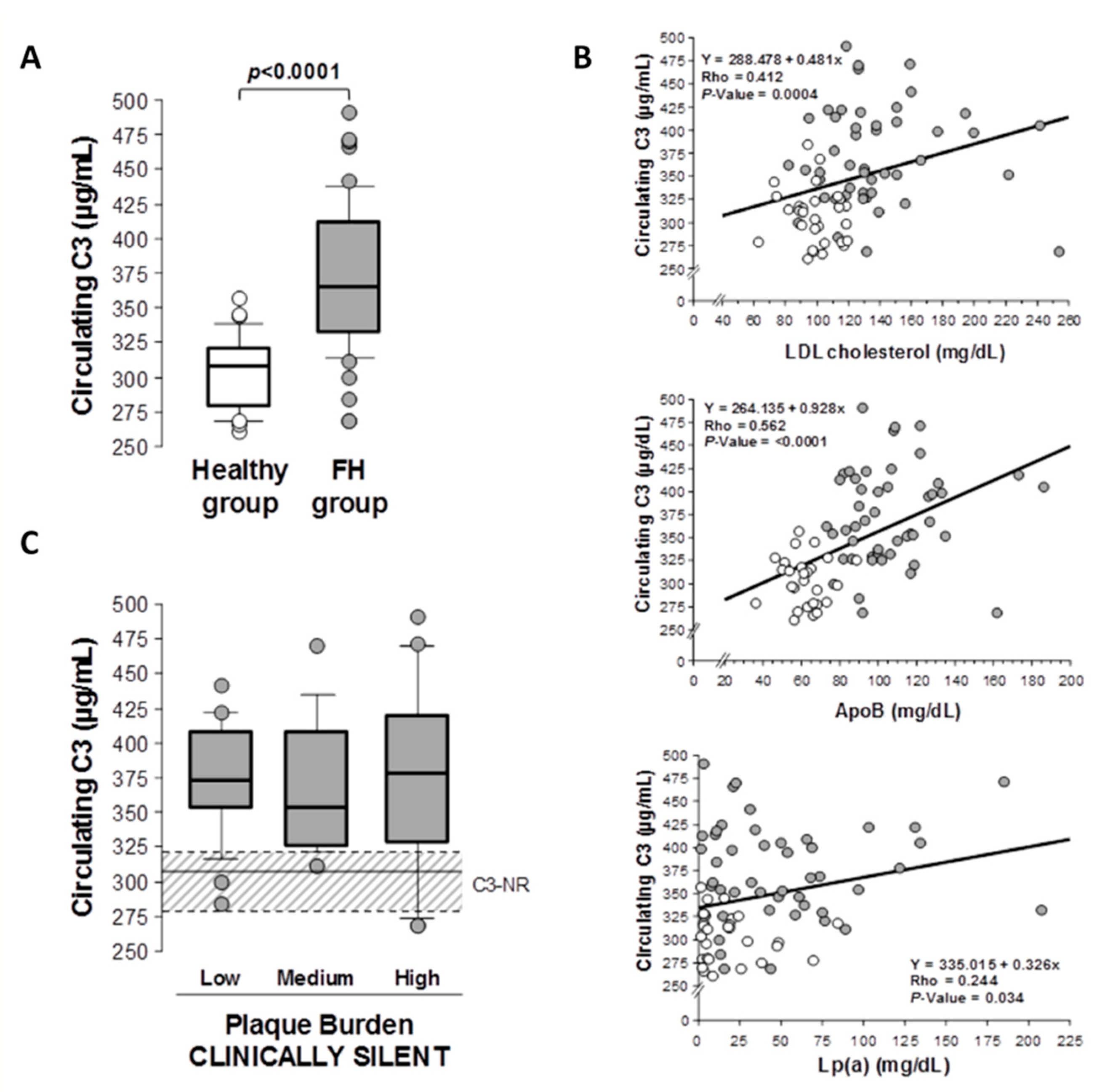
2.2. C3 Complement in Patients with Hypercholesterolemia and Subclinical Atherosclerosis
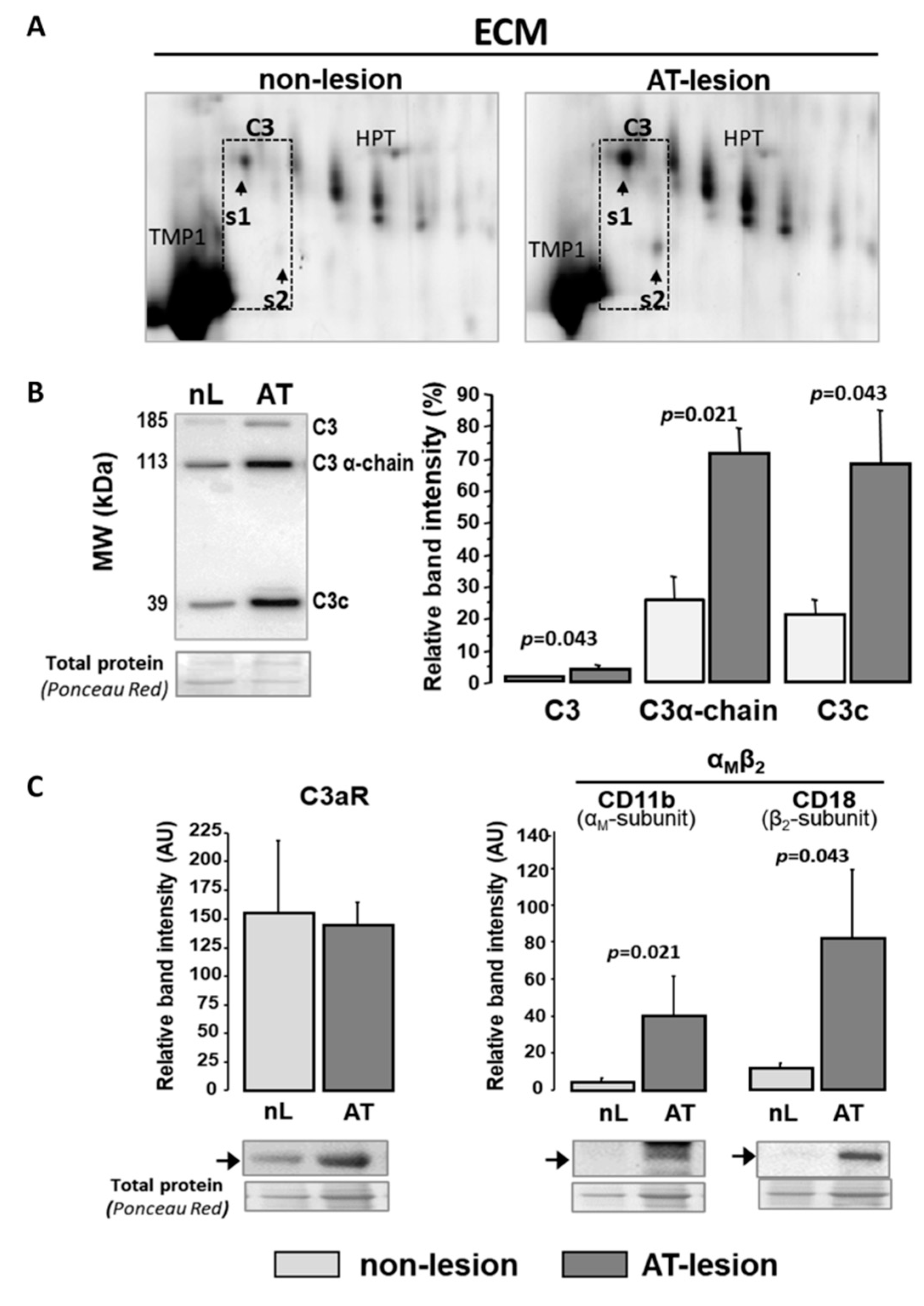
2.3. C3 Alternative System Components in Human Advanced Atherosclerotic Lesions
2.4. C3 Alternative Pathway Components Expression in Vascular Wall Resident Cells
2.5. Exogenous C3 Proteolytic Products, AgLDL and VSMC Function
3. Discussion
4. Materials and Methods
4.1. Human Samples
4.1.1. Subjects with Familial Hypercholesterolemia and Healthy Volunteers
4.1.2. Aortas and Coronary Arteries
4.1.3. VSMC Culture and LDL Preparation
4.2. Tissue Processing and Extraction of ECM Proteins
4.3. 2D Electrophoresis/Mass Spectrometry Analysis
4.4. Western Blot and ELISA Assays
4.5. RNA Extraction and Real-Time PCR Analysis
4.6. Cell Adhesion and Wound-Healing Assays
4.7. Confocal Focal Microscopy
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ridker, P.M. LDL cholesterol: Controversies and future therapeutic directions. Lancet 2014, 384, 607–617. [Google Scholar] [CrossRef]
- Vogt, A. The genetics of familial hypercholesterolemia and emerging therapies. Appl. Clin. Genet. 2015, 8, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.S.; Goldstein, J.L. Familial hypercholesterolemia: Defective binding of lipoproteins to cultured fibroblasts associated with impaired regulation of 3-hydroxy-3-methylglutaryl coenzyme a reductase activity. Proc. Natl. Acad. Sci. USA 1974, 71, 788–792. [Google Scholar] [CrossRef] [Green Version]
- De Isla, L.P.; Alonso, R.; Watts, G.F.; Mata, N.; Cerezo, A.S.; Muñiz, O.; Fuentes, F.; Diaz-Diaz, J.L.; de Andrés, R.; Zambón, D.; et al. Attainment of LDL-Cholesterol treatment goals in patients with familial hypercholesterolemia. J. Am. Coll. Cardiol. 2016, 67, 1278–1285. [Google Scholar] [CrossRef]
- Neefjes, L.A.; Kate, G.-J.R.T.; Alexia, R.; Nieman, K.; Galema-Boers, A.J.; Langendonk, J.G.; Weustink, A.C.; Mollet, N.R.; Sijbrands, E.J.; Krestin, G.P.; et al. Accelerated subclinical coronary atherosclerosis in patients with familial hypercholesterolemia. Atherosclerosis 2011, 219, 721–727. [Google Scholar] [CrossRef] [Green Version]
- Suades, R.; Padro, T.; Alonso, R.; López-Miranda, J.; Mata, P.; Badimon, L. Circulating CD45+/CD3+ lymphocyte-derived microparticles map lipid-rich atherosclerotic plaques in familial hypercholesterolaemia patients. Thromb. Haemost. 2014, 111, 111–121. [Google Scholar] [CrossRef]
- Escate, R.; Mata, P.; Cepeda, J.M.; Padreó, T.; Badimon, L. miR-505-3p controls chemokine receptor up-regulation in macrophages: Role in familial hypercholesterolemia. FASEB J. 2017, 32, 601–612. [Google Scholar] [CrossRef]
- Bahrami, A.; Liberale, L.; Reiner, Ž. Inflammatory biomarkers for cardiovascular risk stratification in familial hy-percholesterolemia. Rev. Physiol. Biochem. Pharmacol. 2020, 177, 25–52. [Google Scholar]
- Holven, K.B.; Narverud, I.; Lindvig, H.W.; Halvorsen, B.; Langslet, G.; Nenseter, M.S.; Ulven, S.M.; Ose, L.; Aukrust, P.; Retterstøl, K. Subjects with familial hypercholesterolemia are characterized by an inflammatory phenotype despite long-term intensive cholesterol lowering treatment. Atherosclerosis 2014, 233, 561–567. [Google Scholar] [CrossRef]
- Real, J.T.; Martinez-Hervas, S.; Garcia-Garcia, A.-B.; Civera, M.; Pallardo, F.V.; Ascaso, J.F.; Vina, J.R.; Chaves, F.J.; Carmena, R.; Garcia-Garcia, A.-B. Circulating mononuclear cells nuclear factor-kappa B activity, plasma xanthine oxidase, and low grade inflammatory markers in adult patients with familial hypercholesterolaemia. Eur. J. Clin. Investig. 2010, 40, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement system Part I—Molecular mechanisms of activation and regulation. Front. Immunol. 2015, 6, 262. [Google Scholar] [CrossRef] [Green Version]
- Merle, N.S.; Noe, R.; Halbwachs-Mecarelli, L.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement system part II: Role in immunity. Front. Immunol. 2015, 6, 257. [Google Scholar] [CrossRef] [Green Version]
- Poston, R.N. Atherosclerosis: Integration of its pathogenesis as a self-perpetuating propagating inflammation: A review. Cardiovasc. Endocrinol. Metab. 2019, 8, 51–61. [Google Scholar] [CrossRef]
- Martin-Ventura, J.L.; Martinez-Lopez, D.; Roldan-Montero, R.; Gomez-Guerrero, C.; Blanco-Colio, L.M. Role of complement system in pathological remodeling of the vascular wall. Mol. Immunol. 2019, 114, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Vlaicu, S.I.; Tatomir, A.; Rus, V.; Mekala, A.P.; Mircea, P.A.; Niculescu, F.; Rus, H. The role of complement activation in atherogenesis: The first 40 years. Immunol. Res. 2016, 64, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Speidl, W.S.; Kastl, S.P.; Huber, K.; Wojta, J. Complement in atherosclerosis: Friend or foe? J. Thromb. Haemost. 2010, 9, 428–440. [Google Scholar] [CrossRef] [PubMed]
- Wezel, A.; De Vries, M.R.; Lagraauw, H.M.; Foks, A.C.; Kuiper, J.; Quax, P.H.; Bot, I. Complement factor C5a induces atherosclerotic plaque disruptions. J. Cell. Mol. Med. 2014, 18, 2020–2030. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.K.; Holm, J.; Kral, J.G. accumulation of igg and complement factor c3 in human arterial endothelium and atherosclerotic lesions. Acta Pathol. Microbiol. Scand. Ser. A Pathol. 1984, 92, 429–435. [Google Scholar] [CrossRef]
- Ge, X.; Xu, C.; Liu, Y.; Zhu, K.; Zeng, H.; Su, J.; Huang, J.; Ji, Y.; Tan, Y.; Hou, Y. Complement activation in the arteries of patients with severe atherosclerosis. Int. J. Clin. Exp. Pathol. 2018, 11, 1–9. [Google Scholar]
- Buono, C.; Come, C.E.; Witztum, J.L.; Maguire, G.F.; Connelly, P.W.; Carroll, M.; Lichtman, A.H. Influence of C3 deficiency on atherosclerosis. Circulation 2002, 105, 3025–3031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Persson, L.; Borén, J.; Robertson, A.-K.L.; Wallenius, V.; Hansson, G.K.; Pekna, M. Lack of complement factor C3, but not factor B, increases hyperlipidemia and atherosclerosis in apolipoprotein E−/− low-density lipoprotein receptor−/− mice. Arter. Thromb. Vasc. Biol. 2004, 24, 1062–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Nanda, V.; DiRenzo, D.; Ye, J.; Xiao, S.; Kojima, Y.; Howe, K.L.; Jarr, K.-U.; Flores, A.M.; Tsantilas, P.; et al. Clonally expanding smooth muscle cells promote atherosclerosis by escaping efferocytosis and activating the complement cascade. Proc. Natl. Acad. Sci. USA 2020, 117, 15818–15826. [Google Scholar] [CrossRef] [PubMed]
- Verdeguer, F.; Kubicek, M.; Pla, D.; Vila-Caballer, M.; Civeira, F.; Castro, C.; Vinué, Á.; Pocoví, M.; Calvete, J.J.; Andrés, V. Complement regulation in murine and human hypercholesterolemia and role in the control of macrophage and smooth muscle cell proliferation. Cardiovasc. Res. 2007, 76, 340–350. [Google Scholar] [CrossRef] [Green Version]
- Van Wijk, D.F.; Sjouke, B.; Figueroa, A.; Emami, H.; van der Valk, F.M.; MacNabb, M.H.; Hemphill, L.C.; Schulte, D.M.; Koopman, M.G.; Lobatto, M.E.; et al. Nonpharmacological lipoprotein apheresis reduces arterial inflammation in familial hypercholesterolemia. J. Am. Coll. Cardiol. 2014, 64, 1418–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toutouzas, K.; Skoumas, J.; Koutagiar, I.; Benetos, G.; Pianou, N.; Georgakopoulos, A.; Galanakos, S.; Antonopoulos, A.; Drakopoulou, M.; Oikonomou, E.K.; et al. Vascular inflammation and metabolic activity in hematopoietic organs and liver in familial combined hyperlipidemia and heterozygous familial hypercholesterolemia. J. Clin. Lipidol. 2018, 12, 33–43. [Google Scholar] [CrossRef]
- Sampietro, T.; Bigazzi, F.; Rossi, G.; Pino, B.D.; Puntoni, M.R.; Sbrana, F.; Chella, E.; Bionda, A. Upregulation of the immune system in primary hypercholesterolaemia: Effect of atorvastatin therapy. J. Intern. Med. 2005, 257, 523–530. [Google Scholar] [CrossRef] [PubMed]
- De Isla, L.P.; Alonso, R.; De Diego, J.J.G.; Muñiz-Grijalvo, O.; Díaz-Díaz, J.L.; Zambón, D.; Miramontes, J.P.; Fuentes, F.; De Andrés, R.; Werenitzky, J.; et al. Coronary plaque burden, plaque characterization and their prognostic implications in familial hypercholesterolemia: A computed tomographic angiography study. Atherosclerosis 2021, 317, 52–58. [Google Scholar] [CrossRef]
- Subirana, M.T.; Juan-Babot, J.O.; Puig, T.; Lucena, J.; Rico, A.; Salguero, M.; Borondo, J.C.; Ordóñez, J.; Arimany, J.; Vázquez, R.; et al. Specific characteristics of sudden death in a mediterranean spanish population. Am. J. Cardiol. 2011, 107, 622–627. [Google Scholar] [CrossRef] [PubMed]
- García-Arguinzonis, M.; Padró, T.; Lugano, R.; Llorente-Cortes, V.; Badimon, L. Low-Density lipoproteins induce heat shock protein 27 dephosphorylation, oligomerization, and subcellular relocalization in human vascular smooth muscle cells. Arter. Thromb. Vasc. Biol. 2010, 30, 1212–1219. [Google Scholar] [CrossRef] [Green Version]
- Lugano, R.; Peña, E.; Casani, L.; Badimon, L.; Padró, T. UPA promotes lipid-loaded vascular smooth muscle cell migration through LRP-1. Cardiovasc. Res. 2013, 100, 262–271. [Google Scholar] [CrossRef] [Green Version]
- Padró, T.; Peña, E.; García-Arguinzonis, M.; Llorente-Cortes, V.; Badimon, L. Low-density lipoproteins impair migration of human coronary vascular smooth muscle cells and induce changes in the proteomic profile of myosin light chain. Cardiovasc. Res. 2007, 77, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Engström, G.; Hedblad, B.; Janzon, L.; Lindgärde, F. Complement C3 and C4 in plasma and incidence of myocardial infarction and stroke: A population-based cohort study. Eur. J. Cardiovasc. Prev. Rehabil. 2007, 14, 392–397. [Google Scholar] [CrossRef]
- Van Greevenbroek, M.M.; Arts, I.C.; Van Der Kallen, C.J.; Geijselaers, S.L.; Feskens, E.J.; Jansen, E.H.; Schalkwijk, C.G.; Stehouwer, C.D.; Hertle, E. Distinct associations of complement C3a and its precursor C3 with atherosclerosis and cardiovascular disease. Thromb. Haemost. 2014, 111, 1102–1111. [Google Scholar] [CrossRef]
- Imanaka-Yoshida, K. Extracellular Matrix Remodeling in Vascular Development and Disease. In Etiology and Morphogenesis of Congenital Heart Disease; Nakanishi, T., Markwald, R.R., Baldwin, H.S., Keller, B.B., Srivastava, D., Yamagishi, H., Eds.; Springer: Tokyo, Japan, 2016; Chapter 29. [Google Scholar]
- Oksjoki, R.; Kovanen, P.T.; Pentikäinen, M.O. Role of complement activation in atherosclerosis. Curr. Opin. Lipidol. 2003, 14, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, S.; Asare, Y.; Grommes, J.; Soehnlein, O.; Lutgens, E.; Shagdarsuren, G.; Togtokh, A.; Jacobs, M.J.; Fischer, J.W.; Bernhagen, J.; et al. High expression of C5L2 correlates with high proinflammatory cytokine expression in advanced human atherosclerotic plaques. Am. J. Pathol. 2014, 184, 2123–2133. [Google Scholar] [CrossRef]
- Hertle, E.; Van Greevenbroek, M.; Arts, I.; Van Der Kallen, C.; Feskens, E.; Schalkwijk, C.; Stehouwer, C. Complement activation products C5a and sC5b-9 are associated with low-grade inflammation and endothelial dysfunction, but not with atherosclerosis in a cross-sectional analysis: The CODAM study. Int. J. Cardiol. 2014, 174, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Martínez-López, D.; Roldan-Montero, R.; García-Marqués, F.; Nuñez, E.; Jorge, I.; Camafeita, E.; Minguez, P.; de Cordoba, S.R.; López-Melgar, B.; Lara-Pezzi, E.; et al. Complement C5 protein as a marker of subclinical atherosclerosis. J. Am. Coll. Cardiol. 2020, 75, 1926–1941. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Becker, L.; Pritchard, D.K.; Gharib, S.A.; Wijsman, E.M.; Bammler, T.K.; Beyer, R.P.; Vaisar, T.; Oram, J.F.; Heinecke, J.W. Cholesterol accumulation regulates expression of macrophage proteins implicated in proteolysis and complement activation. Arter. Thromb. Vasc. Biol. 2012, 32, 2910–2918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camejo, G.; Hurt-Camejo, E.; Wiklund, O.; Bondjers, G. Association of apo B lipoproteins with arterial proteoglycans: Pathological significance and molecular basis. Atherosclerosis 1998, 139, 205–222. [Google Scholar] [CrossRef]
- Williams, K.J. Arterial wall chondroitin sulfate proteoglycans: Diverse molecules with distinct roles in lipoprotein retention and atherogenesis. Curr. Opin. Lipidol. 2001, 12, 477–487. [Google Scholar] [CrossRef]
- Shankman, L.S.; Gomez, D.; Cherepanova, O.A.; Salmon, M.; Alencar, G.F.; Haskins, R.M.; Swiatlowska, P.; Newman, A.A.C.; Greene, E.S.; Straub, A.C. KLF4-dependent phenotypic modulation of smooth muscle cells has a key role in atherosclerotic plaque pathogenesis. Nat. Med. 2015, 21, 628–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chappell, J.; Harman, J.L.; Narasimhan, V.M.; Yu, H.; Foote, K.; Simons, B.D.; Bennett, M.R.; Jorgensen, H.F. Extensive proliferation of a subset of differentiated, yet plastic, medial vascular smooth muscle cells contributes to neointimal formation in mouse injury and atherosclerosis models. Circ. Res. 2016, 119, 1313–1323. [Google Scholar] [CrossRef]
- Frey, A.; Ertl, G.; Angermann, C.E.; Hofmann, U.; Störk, S.; Frantz, S. Complement C3c as a biomarker in heart failure. Mediat. Inflamm. 2013, 2013, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grande, M.A.; Belstrøm, D.; Damgaard, C.; Holmstrup, P.; Thangaraj, S.S.; Nielsen, C.H.; Palarasah, Y. Complement split product C3c in saliva as biomarker for periodontitis and response to periodontal treatment. J. Periodontal Res. 2021, 56, 27–33. [Google Scholar] [CrossRef]
- Goldknopf, I.L.; Sheta, E.A.; Bryson, J.; Folsom, B.; Wilson, C.; Duty, J.; Yen, A.A.; Appel, S.H. Complement C3c and related protein biomarkers in amyotrophic lateral sclerosis and Parkinson’s disease. Biochem. Biophys. Res. Commun. 2006, 342, 1034–1039. [Google Scholar] [CrossRef]
- Pagano, M.B.; Zhou, H.-F.; Ennis, T.L.; Wu, X.; Lambris, J.D.; Atkinson, J.P.; Thompson, R.W.; Hourcade, D.E.; Pham, C.T. Complement-dependent neutrophil recruitment is critical for the development of elastase-induced abdominal aortic aneurysm. Circulation 2009, 119, 1805–1813. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Fontaine, C.; Theveneau, E.; Tzekou, A.; Tada, M.; Woods, M.; Page, K.M.; Parsons, M.; Lambris, J.D.; Mayor, R. Complement Fragment C3a Controls Mutual Cell Attraction during Collective Cell Migration. Dev. Cell 2011, 21, 1026–1037. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Zou, Q.; Ding, X.; Shi, D.; Zhu, X.; Hu, W.; Liu, L.; Zhou, H. Complement component C3a plays a critical role in endothelial activation and leukocyte recruitment into the brain. J. Neuroinflamm. 2016, 13, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Öörni, K.; Pentikäinen, M.O.; Ala-Korpela, M.; Kovanen, P.T. Aggregation, fusion, and vesicle formation of modified low density lipoprotein particles: Molecular mechanisms and effects on matrix interactions. J. Lipid Res. 2000, 41, 1703–1714. [Google Scholar] [CrossRef]
- Badimon, L.; Martinez-Gonzalez, J.; Llorente-Cortes, V.; Rodriguez, C.; Padro, T. Cell biology and lipoproteins in atherosclerosis. Curr. Mol. Med. 2006, 6, 439–456. [Google Scholar] [CrossRef]
- Padró, T.; Lugano, R.; García-Arguinzonis, M.; Badimon, L. LDL-Induced impairment of human vascular smooth muscle cells repair function is reversed by HMG-CoA reductase inhibition. PLoS ONE 2012, 7, e38935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugano, R.; Pena, E.; Badimon, L.; Padro, T. Aggregated low-density lipoprotein induce impairment of the cytoskeleton dynamics through urokinase-type plasminogen activator/urokinase-type plasminogen activator receptor in human vascular smooth muscle cell. J. Thromb. Haemost. 2012, 10, 2158–2167. [Google Scholar] [CrossRef]
- Mata, N.; Alonso, R.; Badimón, L.; Padró, T.; Fuentes, F.; Muñiz, O.; Perez-Jiménez, F.; López-Miranda, J.; Díaz, J.L.; Vidal, J.I.; et al. Clinical characteristics and evaluation of LDL-cholesterol treatment of the Spanish Familial Hypercholesterolemia Longitudinal Cohort Study (SAFEHEART). Lipids Health Dis. 2011, 10, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmann, B. Harmonisation of Medico-Legal Autopsy Rules. Int. J. Leg. Med. 1999, 113, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Basso, C.; Burke, M.; Fornes, P.; Gallagher, P.J.; De Gouveia, R.H.; Sheppard, M.; Thiene, G.; Van Der Wal, A.; On Behalf of the Association for European Cardiovascular Pathology. Guidelines for autopsy investigation of sudden cardiac death. Virchows Archiv 2007, 452, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Guyton, J.R.; Klemp, K.F.; Mims, M.P. Altered ultrastructural morphology of self-aggregated low density lipoproteins: Coalescence of lipid domains forming droplets and vesicles. J. Lipid. Res. 1991, 32, 953–962. [Google Scholar] [CrossRef]
- Llorente-Cortés, V.; Martínez-González, J.; Badimon, L. Esterified cholesterol accumulation induced by aggregated LDL uptake in human vascular smooth muscle cells is reduced by HMG-CoA reductase inhibitors. Arter. Thromb. Vasc. Biol. 1998, 18, 738–746. [Google Scholar] [CrossRef] [Green Version]
- Llorente-Cortés, V.; Otero-Viñas, M.; Badimon, L. Differential role of heparan sulfate proteoglycans on aggregated LDL uptake in human vascular smooth muscle cells and mouse embryonic fibroblasts. Arter. Thromb. Vasc. Biol. 2002, 22, 1905–1911. [Google Scholar] [CrossRef] [Green Version]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Escate, R.; Padro, T.; Badimon, L. LDL accelerates monocyte to macrophage differentiation: Effects on adhesion and anoikis. Atherosclerosis 2016, 246, 177–186. [Google Scholar] [CrossRef]
- Didangelos, A.; Yin, X.; Mandal, K.; Saje, A.; Smith, A.; Xu, Q.; Jahangiri, M.; Mayr, M. Extracellular matrix composition and remodeling in human abdominal aortic aneurysms: A proteomics approach. Mol. Cell. Proteom. 2011, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubedo, J.; Padró, T.; García-Arguinzonis, M.; Vilahur, G.; Miñambres, I.; Pou, J.M.; Ybarra, J.; Badimon, L. A novel truncated form of apolipoprotein A-I transported by dense LDL is increased in diabetic patients. J. Lipid Res. 2015, 56, 1762–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llorente-Cortés, V.; Otero-Viñas, M.; Camino-López, S.; Llampayas, O.; Badimon, L. Aggregated low-density lipoprotein uptake induces membrane tissue factor procoagulant activity and microparticle release in human vascular smooth muscle Cells. Circulation 2004, 110, 452–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Familial Hyperchlesterolemia n = 49 | Healthy Subjects n = 28 | |
|---|---|---|
| Demographic Characteristics; mean ± SD | ||
| Female/male, n | 18/31 | 16/12 |
| Age, years | 38.6 ± 11.3 | 24.5 ± 4.6 |
| Risk Factors; n (%) | ||
| Smokers | 14 (29) | 12 (43) |
| Hypertension | 1 (2) | 0 (0) |
| Diabetes mellitus | 0 (0) | 0 (0) |
| Dyslipidaemia | 48 (98) | 0 (0) |
| Biochemical Data, Mean ± SD | ||
| Total cholesterol, mg/dL | 282 ± 72 | 170 ± 20 |
| Triglycerides, mg/dL | 104 ± 67 | 77 ± 35 |
| HDL cholesterol, mg/dL | 47 ± 11 | 56 ± 15 |
| LDL cholesterol, mg/dL | 221 ± 78 | 99 ± 15 |
| Apo AI, mg/dL | 135 ± 20 | 139 ± 29 |
| Apo B, mg/dL | 134 ± 41 | 61 ± 10 |
| Lipoprotein(a), mg/dL | 42 ± 35 | 18 ± 21 |
| Glucose, mg/dL | 89 ± 9 | 78 ± 9 |
| C-reactive protein | 1.86 ± 2.6 | 0.73 ± 0.2 |
| Subclinical Atherosclerotic Disease; (%) | ||
| Plaque burden, % | 23.5 ± 6.3 | - |
| Calcium burden, % | 2.2 ± 2.5 | - |
| Non-calcium burden | 21.3 ± 5.3 | - |
| BAckground Medication; n (%) | ||
| Angiotensin-converting-enzyme inhibitors | 0 (0) | 0 (0) |
| Angiotensin II receptor blockers | 1 (2) | 0 (0) |
| Beta-blockers | 0 (0) | 0 (0) |
| Diuretics | 2 (4) | 0 (0) |
| Statins * | 39 (80) | 0 (0) |
| Fraction | Protein | UniProt-Code | Gene-Code | MS-Score * | Seq/Int Cov. (%) * | MW (kDa) | pI-Value | Fold-Change |
|---|---|---|---|---|---|---|---|---|
| sb-ECM | Complement Factor H | P08603 | CFH | 102 | 9.9/78.4 | 143.7 | 6.20 | ≈(1.2) |
| Sb-ECM | Complement Factor H-related protein 1 | Q03591 | CFHR1 | 120 | 28.2/95.3 | 38.8 | 8.70 | ↑(2.4) |
| Sb-ECM | Complement Factor H-related protein 5 ** | Q9BXR6 | CFHR5 | 28 | -- | 66.4 | 7.00 | ↑(3.9) |
| Lb-ECM | Complement C3 | P01024 | C3 | 83 | 9.4/76.7 | 188.6 | 6.00 | ↑(2.9) |
| Control | +iC3b | |||
|---|---|---|---|---|
| 30 min | 60 min | 120 min | ||
| −agLDL | 100.0 ± 0.0 | 93.9 ± 4.4 | 115.9 ± 4.8 * | 93.7 ± 14.0 |
| +agLDL | 100.0 ± 0.0 | 65.4 ± 1.8 | 122.0 ± 2.6 | 177.1 ± 42.3 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Arguinzonis, M.; Diaz-Riera, E.; Peña, E.; Escate, R.; Juan-Babot, O.; Mata, P.; Badimon, L.; Padro, T. Alternative C3 Complement System: Lipids and Atherosclerosis. Int. J. Mol. Sci. 2021, 22, 5122. https://doi.org/10.3390/ijms22105122
Garcia-Arguinzonis M, Diaz-Riera E, Peña E, Escate R, Juan-Babot O, Mata P, Badimon L, Padro T. Alternative C3 Complement System: Lipids and Atherosclerosis. International Journal of Molecular Sciences. 2021; 22(10):5122. https://doi.org/10.3390/ijms22105122
Chicago/Turabian StyleGarcia-Arguinzonis, Maisa, Elisa Diaz-Riera, Esther Peña, Rafael Escate, Oriol Juan-Babot, Pedro Mata, Lina Badimon, and Teresa Padro. 2021. "Alternative C3 Complement System: Lipids and Atherosclerosis" International Journal of Molecular Sciences 22, no. 10: 5122. https://doi.org/10.3390/ijms22105122