
MicroRNA as a Prognostic and Diagnostic Marker in T-Cell Acute Lymphoblastic Leukemia


Abstract
:1. Introduction
2. microRNA in T-ALL
2.1. The Influence of miRNA on the Development of T-ALL
2.2. The Impact of miRNA on the Response to Treatment
2.3. The Most Common miRNAs in T-ALL and Their Prognostic Potential
2.3.1. miR-17-92 Cluster
2.3.2. miR-106a-363 Cluster
2.3.3. miR-221-222 Cluster
2.3.4. miR-16
2.3.5. miR-21
2.3.6. miR-26b
2.3.7. miR-30
2.3.8. miR-100
2.3.9. miR-124a
2.3.10. miR-125b and miR-99a
2.3.11. miR-128a
2.3.12. miR-142-3p
2.3.13. miR-146b-5p
2.3.14. miR-150
2.3.15. miR-193b-3p
2.3.16. miR-196b
2.3.17. miR-223
2.3.18. miR-326
2.3.19. miR-451
2.3.20. miR-664
2.3.21. mir-708
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 3′UTR | 3′untranslated region |
| ALL | acute lymphoblastic leukemia |
| AML | Acute myeloid leukemia |
| BAALC | brain and acute lymphoblastic leukemia |
| BCP-ALL | B-cell precursor acute lymphoblastic leukemia |
| CKD6 | cyclin dependent kinase 6 |
| DFR | disease free of recurrence |
| DFS | disease-free survival |
| DR | downregulation |
| EGIL | The European Group for the Immunologic Classification |
| ETP-ALL | Early T-cell precursor ALL |
| GRα | glucocorticoid receptor alpha |
| GSI | γ-secretase inhibitors |
| JAK | Janus kinase |
| LBL | lymphoblastic lymphoma |
| miRNA | MicroRNA |
| MMP9 | metalloproteinase 9 |
| MRD | minimal residual disease |
| OE | overexpression |
| OS | overall survival |
| PBMNC | peripheral blood mononuclear cells |
| PDGF | Platelet Derived Growth Factor |
| RB1 | retinoblastoma protein |
| RFS | relapse-free survival |
| shRNA | short hairpin RNA |
| STK39 | serine/threonine kinase 39 |
| T-ALL | T-cell acute lymphoblastic leukemia |
| TLX | T-cell leukemia homeobox 1 |
| TLX3 | homeobox 3 |
References
- Madhusoodhan, P.P.; Carroll, W.L.; Bhatla, T. Progress and Prospects in Pediatric Leukemia. Curr. Probl. Pediatr. Adolesc. Health Care 2016, 46, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Raetz, E.A.; Teachey, D.T. T-cell acute lymphoblastic leukemia. Hematology (USA) 2016, 2016, 580–588. [Google Scholar] [CrossRef] [Green Version]
- Hefazi, M.; Litzow, M.R. Acute Lymphocytic Leukemias (K Ballen and M Keng, Section Editors) Recent Advances in the Biology and Treatment of T Cell Acute Lymphoblastic Leukemia. Curr. Hematol. Malign Rep. 2018, 13, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Follini, E.; Marchesini, M.; Roti, G. Molecular Sciences Strategies to Overcome Resistance Mechanisms in T-Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2019, 20, 3021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, A.; Sireci, A.; Colovai, A.; Pinkney, K.; Sulis, M.; Bhagat, G.; Alobeid, B. Early T-cell precursor leukemia/lymphoma in adults and children. Leuk. Res. 2013, 37, 1027–1034. [Google Scholar] [CrossRef]
- Asnafi, V.; Bond, J.; Graux, C.; Lhermitte, L.; Lara, D.; Cluzeau, T.; Leguay, T.; Cieslak, A.; Trinquand, A.; Pastoret, C.; et al. Early response–based therapy stratification improves survival in adult early thymic precursor acute lymphoblastic leukemia: A Group for research on adult acute lymphoblastic leukemia study. J. Clin. Oncol. 2017, 35, 2683–2691. [Google Scholar] [CrossRef] [Green Version]
- Noronha, E.P.; Marques, L.V.C.; Andrade, F.G.; Sardou-Cezar, I.; Dos Santos-Bueno, F.V.; Da Paz Zampier, C.; Terra-Granado, E.; Pombo-de-Oliveira, M.S. T-lymphoid/myeloid mixed phenotype acute leukemia and early T-cell precursor lymphoblastic leukemia similarities with NOTCH1 mutation as a good prognostic factor. Cancer Manag. Res. 2019, 11, 3933–3943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiaretti, S.; Zini, G.; Bassan, R. Diagnosis and Subclassification of Acute Lymphoblastic Leukemia. Cit. Mediterr. J. Hematol. Infect. Dis. 2014, 6, 2014073. [Google Scholar] [CrossRef] [Green Version]
- Coustan-Smith, E.; Mullighan, C.G.; Onciu, M.; Behm, F.G.; Raimondi, S.C.; Pei, D.; Cheng, C.; Su, X.; Rubnitz, J.E.; Basso, G.; et al. Early T-cell precursor leukaemia: A subtype of very high-risk acute lymphoblastic leukaemia. Lancet Oncol. 2009, 10, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Jain, N.; Lamb, A.V.; O’Brien, S.; Ravandi, F.; Konopleva, M.; Jabbour, E.; Zuo, Z.; Jorgensen, J.; Lin, P.; Pierce, S.; et al. Early T-cell precursor acute lymphoblastic leukemia/lymphoma (ETP-ALL/LBL) in adolescents and adults: A high-risk subtype. Blood 2016, 127, 1863–1869. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Li, W. Genomics and pharmacogenomics of pediatric acute lymphoblastic leukemia. Crit. Rev. Oncol. Hematol. 2018, 126, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Pui, C.H.; Carroll, W.L.; Meshinchi, S.; Arceci, R.J. Biology, risk stratification, and therapy of pediatric acute leukemias: An update. J. Clin. Oncol. 2011, 29, 551–565. [Google Scholar] [CrossRef] [Green Version]
- Carroll, W.L.; Raetz, E.A. Clinical and laboratory biology of childhood acute lymphoblastic leukemia. J. Pediatr. 2012, 160, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Belver, L.; Ferrando, A. The genetics and mechanisms of T cell acute lymphoblastic leukaemia. Nat. Rev. Cancer 2016, 16, 494–507. [Google Scholar] [CrossRef]
- Liu, X.-L.; Liu, H.-Q.; Li, J.; Mao, C.-Y.; He, J.-T.; Zhao, X. Role of epigenetic in leukemia: From mechanism to therapy. Chem. Biol. Interact. 2020, 317, 108963. [Google Scholar] [CrossRef]
- Memari, F.; Joneidi, Z.; Taheri, B.; Aval, S.F.; Roointan, A.; Zarghami, N. Epigenetics and Epi-miRNAs: Potential markers/therapeutics in leukemia. Biomed. Pharmacother. 2018, 106, 1668–1677. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Heikkinen, L.; Wang, C.; Yang, Y.; Sun, H.; Wong, G. Trends in the development of miRNA bioinformatics tools. Brief. Bioinform. 2019, 20, 1836–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicente, C.; Schwab, C.; Broux, M.; Geerdens, E.; Degryse, S.; Demeyer, S.; Lahortiga, I.; Elliott, A.; Chilton, L.; La Starza, R.; et al. Targeted sequencing identifies associations between IL7R-JAK mutations and epigenetic modulators in T-cell acute lymphoblastic leukemia. Haematologica 2015, 100, 1301–1310. [Google Scholar] [CrossRef] [Green Version]
- Drobna, M.; Szarzyńska-Zawadzka, B.; Daca-Roszak, P.; Kosmalska, M.; Jaksik, R.; Witt, M.; Dawidowska, M. Identification of endogenous control mirnas for RT-qPCR in T-cell acute lymphoblastic leukemia. Int. J. Mol. Sci. 2018, 19, 2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawidowska, M.; Jaksik, R.; Drobna, M.; Szarzyńska-Zawadzka, B.; Kosmalska, M.; Sędek, Ł.; Machowska, L.; Lalik, A.; Lejman, M.; Ussowicz, M.; et al. Comprehensive Investigation of miRNome Identifies Novel Candidate miRNA-mRNA Interactions Implicated in T-Cell Acute Lymphoblastic Leukemia. Neoplasia (USA) 2019, 21, 294–310. [Google Scholar] [CrossRef] [PubMed]
- Coskun, E.; von der Heide, E.K.; Schlee, C.; Kühnl, A.; Gökbuget, N.; Hoelzer, D.; Hofmann, W.K.; Thiel, E.; Baldus, C.D. The role of microRNA-196a and microRNA-196b as ERG regulators in acute myeloid leukemia and acute T-lymphoblastic leukemia. Leuk. Res. 2011, 35, 208–213. [Google Scholar] [CrossRef]
- Schotte, D.; Chau, J.C.K.; Sylvester, G.; Liu, G.; Chen, C.; van der Velden, V.H.J.; Broekhuis, M.J.C.; Peters, T.C.J.M.; Pieters, R.; den Boer, M.L. Identification of new microRNA genes and aberrant microRNA profiles in childhood acute lymphoblastic leukemia. Leukemia 2009, 23, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Coskun, E.; Neumann, M.; Schlee, C.; Liebertz, F.; Heesch, S.; Goekbuget, N.; Hoelzer, D.; Baldus, C.D. MicroRNA profiling reveals aberrant microRNA expression in adult ETP-ALL and functional studies implicate a role for miR-222 in acute leukemia. Leuk. Res. 2013, 37, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Correia, N.C.; Barata, J.T. MicroRNAs and their involvement in T-ALL: A brief overview. Adv. Biol. Regul. 2019, 74, 100650. [Google Scholar] [CrossRef]
- Drobna, M.; Szarzyńska-Zawadzka, B.; Dawidowska, M. T-cell acute lymphoblastic leukemia from miRNA perspective: Basic concepts, experimental approaches, and potential biomarkers. Blood Rev. 2018, 32, 457–472. [Google Scholar] [CrossRef]
- Wallaert, A.; Van Loocke, W.; Hernandez, L.; Taghon, T.; Speleman, F.; Van Vlierberghe, P. Comprehensive miRNA expression profiling in human T-cell acute lymphoblastic leukemia by small RNA-sequencing. Sci. Rep. 2017, 7, 7901. [Google Scholar] [CrossRef] [Green Version]
- Nemes, K.; Csóka, M.; Nagy, N.; Márk, Á.; Váradi, Z.; Dankó, T.; Kovács, G.; Kopper, L.; Sebestyén, A. Expression of Certain Leukemia/Lymphoma Related microRNAs and its Correlation with Prognosis in Childhood Acute Lymphoblastic Leukemia. Pathol. Oncol. Res. 2015, 21, 597–604. [Google Scholar] [CrossRef]
- Agirre, X.; Vilas-Zornoza, A.; Jiménez-Velasco, A.; Martin-Subero, J.I.; Cordeu, L.; Gárate, L.; José-Eneriz, E.S.; Abizanda, G.; Rodríguez-Otero, P.; Fortes, P.; et al. Epigenetic silencing of the tumor suppressor microRNA Hsa-miR-124a regulates CDK6 expression and confers a poor prognosis in acute lymphoblastic leukemia. Cancer Res. 2009, 69, 4443–4453. [Google Scholar] [CrossRef] [Green Version]
- Li, X.J.; Luo, X.Q.; Han, B.W.; Duan, F.T.; Wei, P.P.; Chen, Y.Q. MicroRNA-100/99a, deregulated in acute lymphoblastic leukaemia, suppress proliferation and promote apoptosis by regulating the FKBP51 and IGF1R/mTOR signalling pathways. Br. J. Cancer 2013, 109, 2189–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, M.; Zhang, Q.; Xia, M.; Hu, F.; Ma, Z.; Chen, Z.; Guo, A.Y. Differential Co-expression and Regulatory Network Analysis Uncover the Relapse Factor and Mechanism of T Cell Acute Leukemia. Mol. Ther. Nucleic Acids 2018, 12, 184–194. [Google Scholar] [CrossRef] [Green Version]
- Mesrian Tanha, H.; Mojtabavi Naeini, M.; Rahgozar, S.; Moafi, A.; Honardoost, M.A. Integrative computational in-depth analysis of dysregulated miRNA-mRNA interactions in drug-resistant pediatric acute lymphoblastic leukemia cells: An attempt to obtain new potential gene-miRNA pathways involved in response to treatment. Tumor Biol. 2016, 37, 7861–7872. [Google Scholar] [CrossRef]
- Mei, Y.; Gao, C.; Wang, K.; Cui, L.; Li, W.; Zhao, X.; Liu, F.; Wu, M.; Deng, G.; Ding, W.; et al. Effect of microRNA-210 on prognosis and response to chemotherapeutic drugs in pediatric acute lymphoblastic leukemia. Cancer Sci. 2014, 105, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Zhang, W.; Lei, B.; He, A.; Ye, L.; Li, X.; Dong, X. MicroRNA-101 regulates T-cell acute lymphoblastic leukemia progression and chemotherapeutic sensitivity by targeting Notch1. Oncol. Rep. 2016, 36, 2511–2516. [Google Scholar] [CrossRef] [Green Version]
- Mavrakis, K.J.; Van Der Meulen, J.; Wolfe, A.L.; Liu, X.; Mets, E.; Taghon, T.; Khan, A.A.; Setti, M.; Rondou, P.; Vandenberghe, P.; et al. A cooperative microRNA-tumor suppressor gene network in acute T-cell lymphoblastic leukemia (T-ALL). In Proceedings of the 11th Annual Meeting of the Belgian Society of Human Genetics (BeSHG), Louvain-La-Neuve, Belgium, 4 March 2011; Volume 43, pp. 673–678. [Google Scholar]
- Rawoof, A.; Swaminathan, G.; Tiwari, S.; Nair, R.A.; Dinesh Kumar, L. LeukmiR: A database for miRNAs and their targets in acute lymphoblastic leukemia. Database 2020, 2020, baz151. [Google Scholar] [CrossRef]
- Ravegnini, G.; Cargnin, S.; Sammarini, G.; Zanotti, F.; Bermejo, J.L.; Hrelia, P.; Terrazzino, S.; Angelini, S. Prognostic role of miR-221 and miR-222 expression in cancer patients: A systematic review and meta-analysis. Cancers 2019, 11, 970. [Google Scholar] [CrossRef] [Green Version]
- Gimenes-Teixeira, H.L.; Lucena-Araujo, A.R.; dos Santos, G.A.; Zanette, D.L.; Scheucher, P.S.; Oliveira, L.C.; Dalmazzo, L.F.; Silva-Júnior, W.A.; Falcão, R.P.; Rego, E.M. Increased expression of miR-221 is associated with shorter overall survival in T-cell acute lymphoid leukemia. Exp. Hematol. Oncol. 2013, 2, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.W.; Li, H.; Zhang, Z.P.; Zhuo, F.; Li, Z.X. Expression and Clinical Significance of MiR-146a and MiR-221 in Childhood Acute T Lymphoblastic Leukemia. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2020, 28, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Xing, C.; Chen, C.; Lin, S.; Dong, P.; Yu, F. miR-15a and miR-16-1 inhibit the proliferation of leukemic cells by down-regulating WT1 protein level. J. Exp. Clin. Cancer Res. 2011, 30, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aqeilan, R.I.; Calin, G.A.; Croce, C.M. MiR-15a and miR-16-1 in cancer: Discovery, function and future perspectives. Cell Death Differ. 2010, 17, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, X.; Li, X.; Shen, Y.; Miao, J.; Liu, H.; Li, G.; Wang, Z. MiR-16 regulates mouse peritoneal macrophage polarization and affects T-cell activation. J. Cell. Mol. Med. 2016, 20, 1898–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, Y.; Li, J.; Zan, L.; Wang, J.; Wang, G.; Ning, Y. Micro-RNA-16 expression in paraffin-embedded specimen correlates with overall survival of T-lymphoblastic lymphoma/leukemia. Hum. Pathol. 2013, 44, 1011–1016. [Google Scholar] [CrossRef]
- Junker, F.; Chabloz, A.; Koch, U.; Radtke, F. Dicer1 imparts essential survival cues in Notch-driven T-ALL via miR-21-mediated tumor suppressor Pdcd4 repression. Blood 2015, 126, 993–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, A.; Yunes, J.A.; Cardoso, B.A.; Martins, L.R.; Jotta, P.Y.; Abecasis, M.; Nowill, A.E.; Leslie, N.R.; Cardoso, A.A.; Barata, J.T. PTEN posttranslational inactivation and hyperactivation of the PI3K/Akt pathway sustain primary T cell leukemia viability. J. Clin. Investig. 2008, 118, 3762–3774. [Google Scholar] [CrossRef] [Green Version]
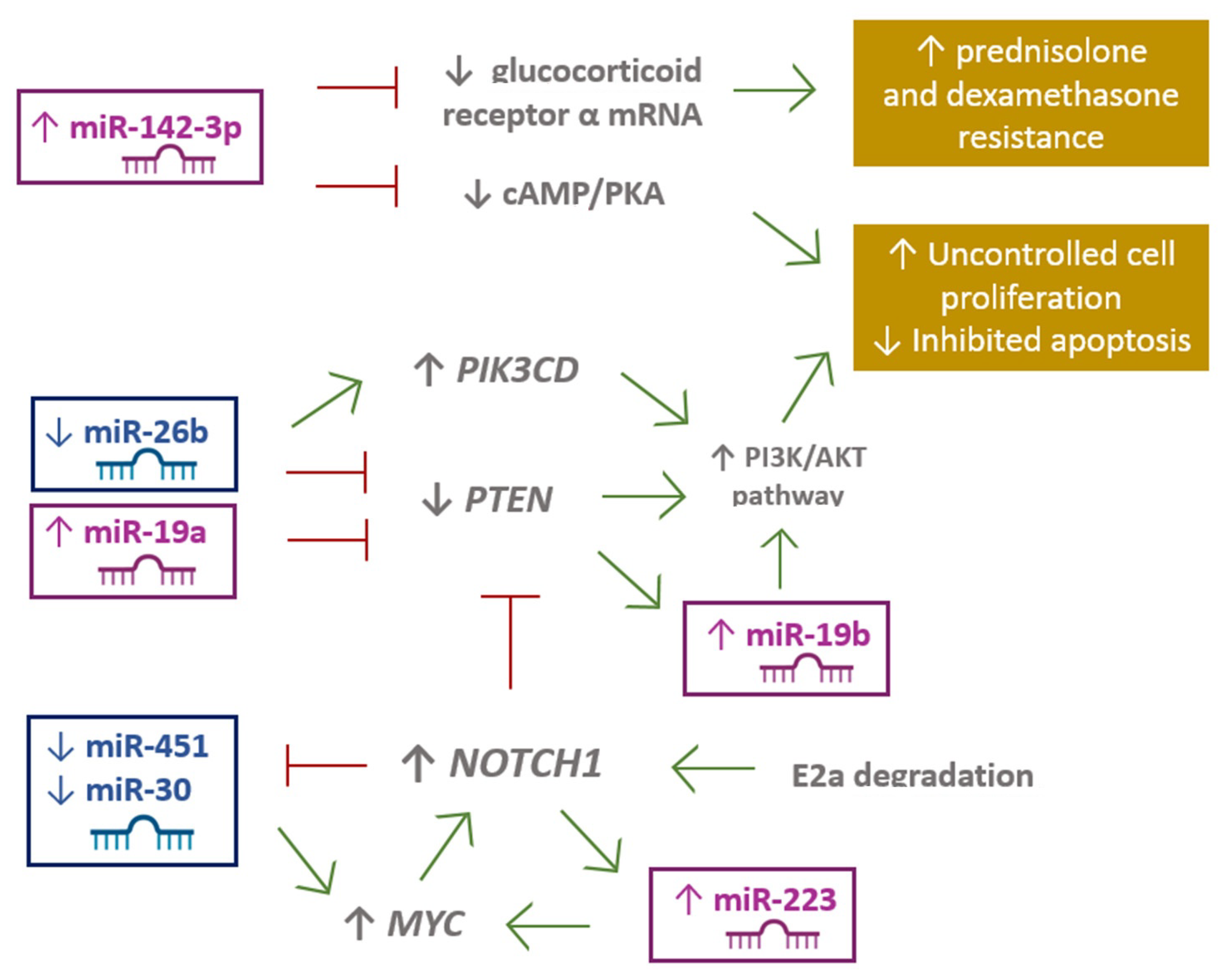
- Yuan, T.; Yang, Y.; Chen, J.; Li, W.; Li, W.; Zhang, Q.; Mi, Y.; Goswami, R.S.; You, J.Q.; Lin, D.; et al. Regulation of PI3K signaling in T cell acute lymphoblastic leukemia: A novel PTEN/Ikaros/miR-26b mechanism reveals a critical targetable role for PIK3CD HHS Public Access. Leukemia 2017, 31, 2355–2364. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Yang, X.; Hu, S.; Kang, M.; Chen, J.; Fang, Y. A genetic variant in miR-100 is a protective factor of childhood acute lymphoblastic leukemia. Cancer Med. 2019, 8, 2553–2560. [Google Scholar] [CrossRef]
- Hassan, N.M.; Refaat, L.A.; Ismail, G.N.; Abdellateif, M.; Fadel, S.A.; AbdelAziz, R.S. Diagnostic, prognostic and predictive values of miR-100 and miR-210 in pediatric acute lymphoblastic Leukemia. Hematology (UK) 2020, 25, 405–413. [Google Scholar] [CrossRef]
- Renou, L.; Boelle, P.-Y.; Deswarte, C.; Spicuglia, S.; Benyoucef, A.; Calvo, J.; Uzan, B.; Belhocine, M.; Cieslak, A.; Landman-Parker, J.; et al. Homeobox protein TLX3 activates miR-125b expression to promote T-cell acute lymphoblastic leukemia. Blood Adv. 2017, 1, 733–747. [Google Scholar] [CrossRef] [Green Version]
- Carvalho De Oliveira, J.; Scrideli, C.A.; Brassesco, M.S.; Gois Morales, A.; Pezuk, J.A.; De Paula Queiroz, R.; Yunes, A.; Brandalise, S.R.; Gonzaga Tone, L. Differential MiRNA expression in childhood acute lymphoblastic leukemia and association with clinical and biological features. Leuk. Res. 2011, 36, 293–298. [Google Scholar] [CrossRef]
- Lv, M.; Zhang, X.; Jia, H.; Li, D.; Zhang, B.; Zhang, H.; Hong, M.; Jiang, T.; Jiang, Q.; Lu, J.; et al. An oncogenic role of miR-142-3p in human T-cell acute lymphoblastic leukemia (T-ALL) by targeting glucocorticoid receptor-α and cAMP/PKA pathways. Leukemia 2012, 26, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Tu, Z.; Xiong, J.; Xiao, R.; Shao, L.; Yang, X.; Zhou, L.; Yuan, W.; Wang, M.; Yin, Q.; Wu, Y.; et al. Loss of miR-146b-5p promotes T cell acute lymphoblastic leukemia migration and invasion via the IL-17A pathway. J. Cell. Biochem. 2019, 120, 5936–5948. [Google Scholar] [CrossRef]
- Correia, N.C.; Fragoso, R.; Carvalho, T.; Enguita, F.J.; Barata, J.T. MiR-146b negatively regulates migration and delays progression of T-cell acute lymphoblastic leukemia. Sci. Rep. 2016, 6, 31894. [Google Scholar] [CrossRef]
- Mets, E.; Van Der Meulen, J.; Van Peer, G.; Boice, M.; Mestdagh, P.; Van De Walle, I.; Lammens, T.; Goossens, S.; De Moerloose, B.; Benoit, Y.; et al. MicroRNA-193b-3p acts as a tumor suppressor by targeting the MYB oncogene in T-cell acute lymphoblastic leukemia. Leukemia 2015, 29, 798–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schotte, D.; Lange-Turenhout, E.A.M.; Stumpel, D.J.P.M.; Stam, R.W.; Buijs-Gladdines, J.G.C.A.M.; Meijerink, J.P.P.; Pieters, R.; den Boer, M.L. Expression of miR-196b is not exclusively MLL-driven but is especially linked to activation of HOXA genes in pediatric acute lymphoblastic leukemia. Haematologica 2010, 95, 1675–1682. [Google Scholar] [CrossRef]
- Mansour, M.R.; Sanda, T.; Lawton, L.N.; Li, X.; Kreslavsky, T.; Novina, C.D.; Brand, M.; Gutierrez, A.; Kelliher, M.A.; Jamieson, C.H.M.; et al. The TAL1 complex targets the FBXW7 tumor suppressor by activating miR-223 in human T cell acute lymphoblastic leukemia. J. Exp. Med. 2013, 210, 1545–1557. [Google Scholar] [CrossRef] [PubMed]
- Gusscott, S.; Kuchenbauer, F.; Humphries, R.K.; Weng, A.P. Notch-mediated repression of miR-223 contributes to IGF1R regulation in T-ALL. Leuk. Res. 2012, 36, 905–911. [Google Scholar] [CrossRef]
- Sadat Ghodousi, E.; Aberuyi, N.; Rahgozar, S. Simultaneous changes in expression levels of BAALC and miR-326: A novel prognostic biomarker for childhood ALL. Jpn. J. Clin. Oncol. 2020, 50, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Wu, S. miR-451: A Novel Biomarker and Potential Therapeutic Target for Cancer. Onco Targets Ther. 2019, 12, 11069–11082. [Google Scholar] [CrossRef] [Green Version]
- Whitman, S.P.; Maharry, K.; Radmacher, M.D.; Becker, H.; Mrózek, K.; Margeson, D.; Holland, K.B.; Wu, Y.Z.; Schwind, S.; Metzeler, K.H.; et al. FLT3 internal tandem duplication associates with adverse outcome and gene- and microRNA-expression signatures in patients 60 years of age or older with primary cytogenetically normal acute myeloid leukemia: A cancer and leukemia group B study. Blood 2010, 116, 3622–3626. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Wang, J.; Yue, M.; Cai, X.; Wang, T.; Wu, C.; Su, H.; Wang, Y.; Han, M.; Zhang, Y.; et al. Direct Phosphorylation and Stabilization of MYC by Aurora B Kinase Promote T-cell Leukemogenesis. Cancer Cell 2020, 37, 200–215.e5. [Google Scholar] [CrossRef]
- Huang, A.; Zhao, H.; Quan, Y.; Jin, R.; Feng, B.; Zheng, M. E2A Predicts Prognosis of Colorectal Cancer Patients and Regulates Cancer Cell Growth by Targeting miR-320a. PLoS ONE 2014, 9, e85201. [Google Scholar] [CrossRef]
- Li, X.; Sanda, T.; Thomas Look, A.; Novina, C.D.; von Boehmer, H. Repression of tumor suppressor miR-451 is essential for NOTCH1-induced oncogenesis in T-ALL. J. Exp. Med. 2011, 208, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Tao, P.; Feng, J.; Li, Q.; Liu, W.E.I.; Yang, L.I.N.; Zhao, X.; Ni, H.; Xia, P. Expression of miR-664 and miR-184 on proliferation, apoptosis and migration of osteosarcoma cells. Oncol. Lett. 2019, 17, 1791–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, B.W.; Feng, D.D.; Li, Z.G.; Luo, X.Q.; Zhang, H.; Li, X.J.; Zhang, X.J.; Zheng, L.L.; Zeng, C.W.; Lin, K.Y.; et al. A set of miRNAs that involve in the pathways of drug resistance and leukemic stem-cell differentiation is associated with the risk of relapse and glucocorticoid response in childhood ALL. Hum. Mol. Genet. 2011, 20, 4903–4915. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Wang, W.-T.; Fang, K.; Chen, Z.-H.; Sun, Y.-M.; Han, C.; Sun, L.-Y.; Luo, X.-Q.; Chen, Y.-Q. MIR-708 promotes phagocytosis to eradicate T-ALL cells by targeting CD47. Mol. Cancer 2018, 17, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
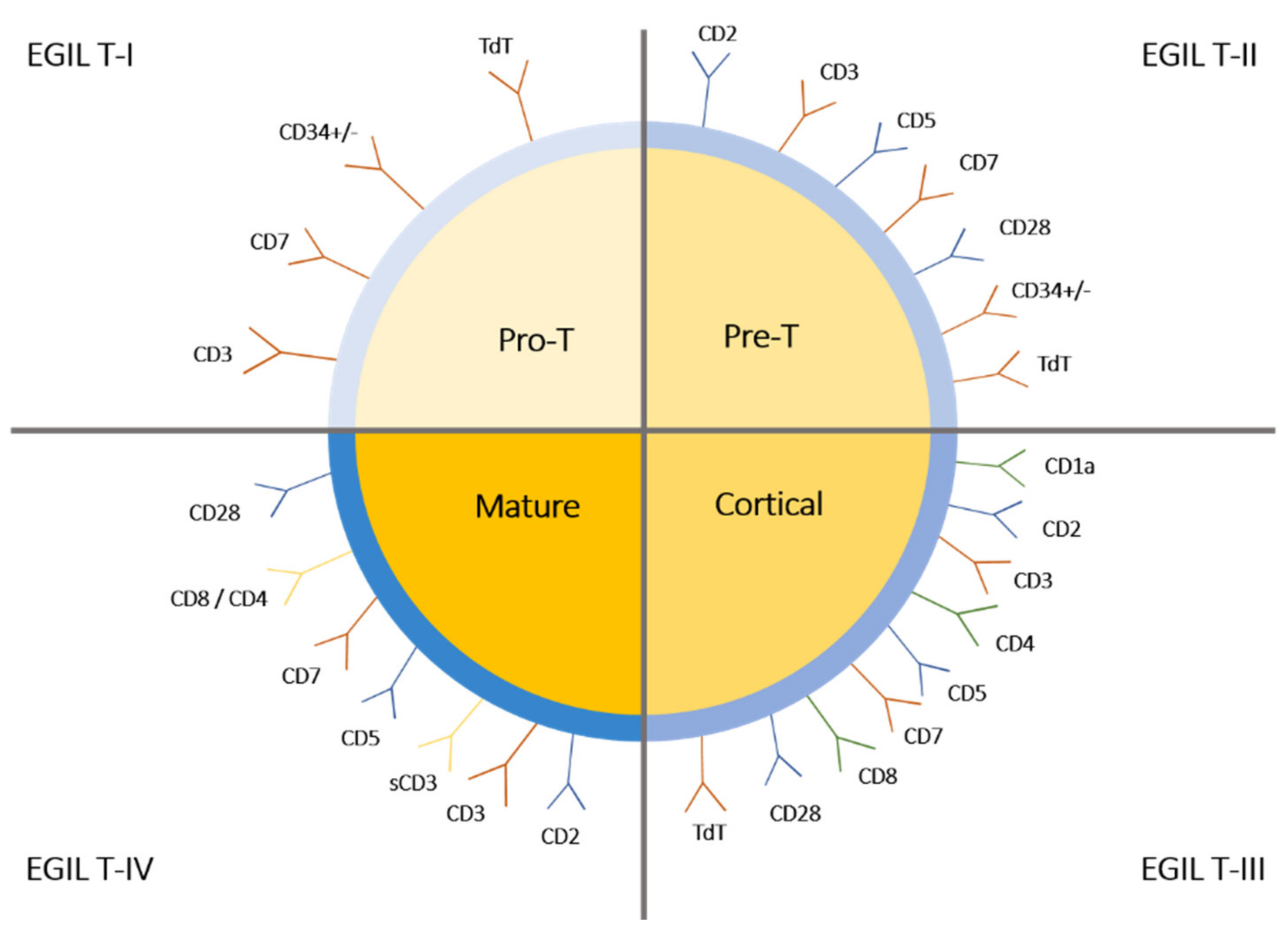
| Genes overexpressed in ETP-ALL | CD44, CD34, KIT, GATA2, CEPBA, SPI1, ID2, MYB |
| Genes underexpressed in ETP-ALL | CD1, CD3, CD4, CD8, RAG1, NOTCH3, PTCRA, LEF1, TCF12, LAT, LCK, TCF7, ZAP70 |
| Myeloid or stem cell markers | CD117, CD34, HLA-DR, CD13, CD33, CD11b, CD65 |
| Immature phenotype of T cells in ETP-ALL | Lack of CD1a and CD8, weak CD5(dim) expression |
| miRNA | Locus | OE/DR | Subtype | Target | Outcome | Reference |
|---|---|---|---|---|---|---|
| miR-16 | 13q14.2 | OE | T-ALL | No data | Higher one year OS | [43] |
| miR-17-5p | 13q31 | DR | T-ALL | Upregulation of lysosomal proteins | Poor prognosis Daunorubicin resistance | [32] |
| miR-26a | 3p22.2 | OE | T-ALL | PHF6, PTEN | Oncogenic role in T-ALL | [35] |
| miR-30a-3p | 6q13 | OE | T-ALL | STAG2, PSME3, RAD1, PSMD11, PSMD7, PSMD3, MDM2, CDC27, CDK1, CCNA2 | Suppression of apoptosis by impair cell cycle arrest in G1 phase (STAG2, PSME3, RAD1) | [32] |
| miR-100 | 11q24.1 | OE | T-ALL | No data | Vincristine resistance | [32] |
| miR-124a | 8p23.1 | DR | T-ALL | CDK6; phosphorylation of retinoblastoma | Vincristine resistance; Increased mortality and recurrence rates and the shorter OS, DFS; poor prognosis | [29] |
| miR-125b | 11q24.0 | OE | T-ALL | No data | Vincristine and daunorubicin cross-resistance | [32] |
| miR-125b-5p | 11q24.1 | OE | T-ALL | MAD4 | Bortezomib and dexamethasone resistance | [32] |
| ETS1 | Vincistrine resistance | |||||
| miR-128b | 3p22.3 | DR | T-ALL | No data | Poor prognosis and poor response to prednisone treatment on day 8 | [28] |
| miR-128-3p | 2q21.3 | DR | T-ALL | Upregulation of lysosomal proteins | Poor prognosis Daunorubicin resistance | [32] |
| miR-142-3p | 17q22 | OE | T-ALL | Decreased cAMP/PKA, GRa | Promote leukemic cell growth and glucocorticoid resistance | [51] |
| miR-146a-5p | 5q33.3 | OE | T-ALL | STAG2, PSME3, RAD1, PSMD11, PSMD7, PSMD3, MDM2, CDC27, CDK1, CCNA2 | NGS signaling (PSMD7) | [32] |
| miR-221 | Xp11.3 | OE | ETP-ALL | No data | Lower 5-year OS; poor prognosis | [26] |
| High (N-CAM; CD56) | [25] | |||||
| miR-222 | Xp11.3 | OE | ETP-ALL | Protooncogene ETS1-inhibition | Poor prognosis | [24,25] |
| miR-223-3p; miR-223-5p | Xq12 | DR | T-ALL | IGFR | Cell survival and proliferation (relapse) | [28] |
| Increases IL6/JAK/STAT | No data | [21] | ||||
| PTEN, BIM, NF1, FBXW7, IKZF1, PHF6 | Increases the level of anti-apoptotic MCL1 | |||||
| Decreases: E2F1, FOXO1, RHOB, EPB41L3 | No data | |||||
| OE | FBXW7; MYC | No data | [25,35] | |||
| miR-326 | 11q13.4 | OE | T-ALL | BAALC | Drug resistance | [48] |
| miR-363-3p | Xq26.2 | DR | ETP-ALL | High MN1 | No data | [24] |
| miR-451a | 17q11.2 | DR | NOTCH1 T-ALL | Degradation E2a | T-ALL induced by NOTCH1 | [62] |
| Overexpression of MYC | Disease progression, increased T-ALL cell proliferation | |||||
| miR-708 | 11q14.1 | OE | T-ALL | E2F1 | No data | [65] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gębarowska, K.; Mroczek, A.; Kowalczyk, J.R.; Lejman, M. MicroRNA as a Prognostic and Diagnostic Marker in T-Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2021, 22, 5317. https://doi.org/10.3390/ijms22105317
Gębarowska K, Mroczek A, Kowalczyk JR, Lejman M. MicroRNA as a Prognostic and Diagnostic Marker in T-Cell Acute Lymphoblastic Leukemia. International Journal of Molecular Sciences. 2021; 22(10):5317. https://doi.org/10.3390/ijms22105317
Chicago/Turabian StyleGębarowska, Katarzyna, Anna Mroczek, Jerzy R. Kowalczyk, and Monika Lejman. 2021. "MicroRNA as a Prognostic and Diagnostic Marker in T-Cell Acute Lymphoblastic Leukemia" International Journal of Molecular Sciences 22, no. 10: 5317. https://doi.org/10.3390/ijms22105317
APA StyleGębarowska, K., Mroczek, A., Kowalczyk, J. R., & Lejman, M. (2021). MicroRNA as a Prognostic and Diagnostic Marker in T-Cell Acute Lymphoblastic Leukemia. International Journal of Molecular Sciences, 22(10), 5317. https://doi.org/10.3390/ijms22105317