
KRAS and EGFR Mutations Differentially Alter ABC Drug Transporter Expression in Cisplatin-Resistant Non-Small Cell Lung Cancer

 ,
, 
 ,
, 
Abstract
1. Introduction
2. Results
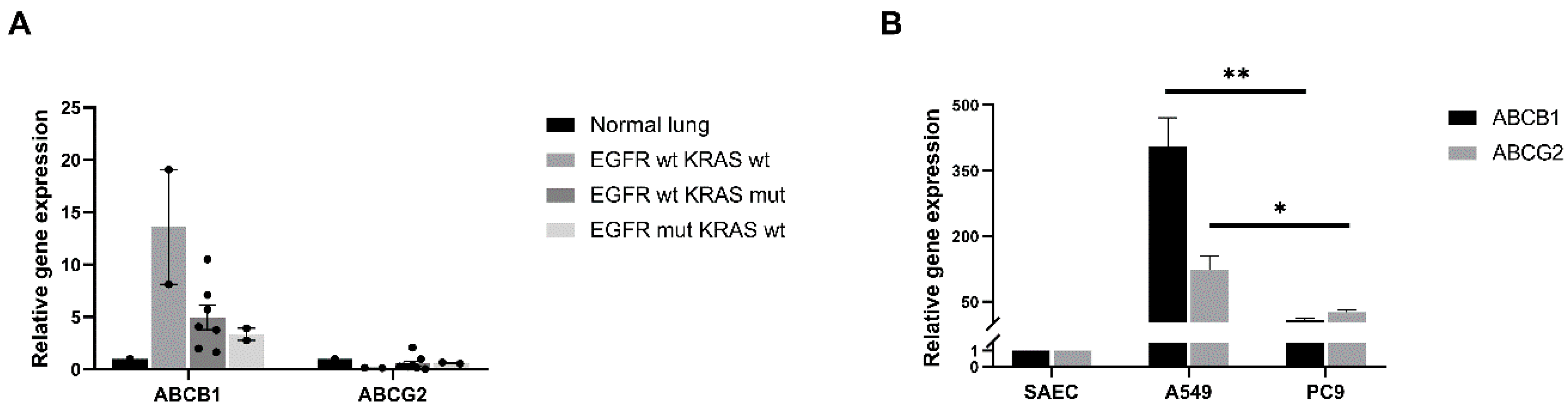
2.1. Differential ABCB1 and ABCG2 mRNA Levels Correlate with Driver Mutations in Primary Human Lung AC Samples
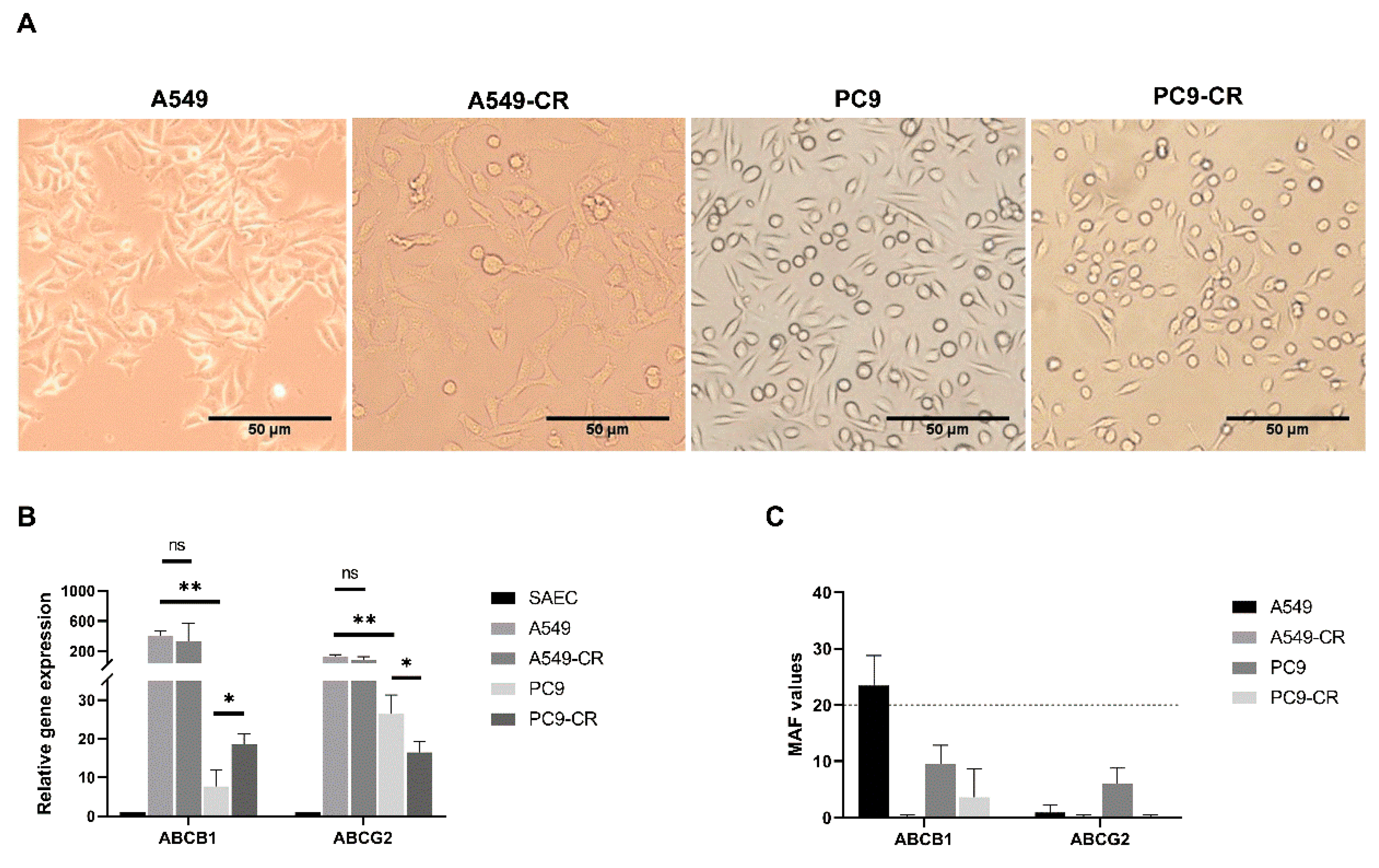
2.2. Cisplatin Resistance Alters ABC Transporter Expression and Function
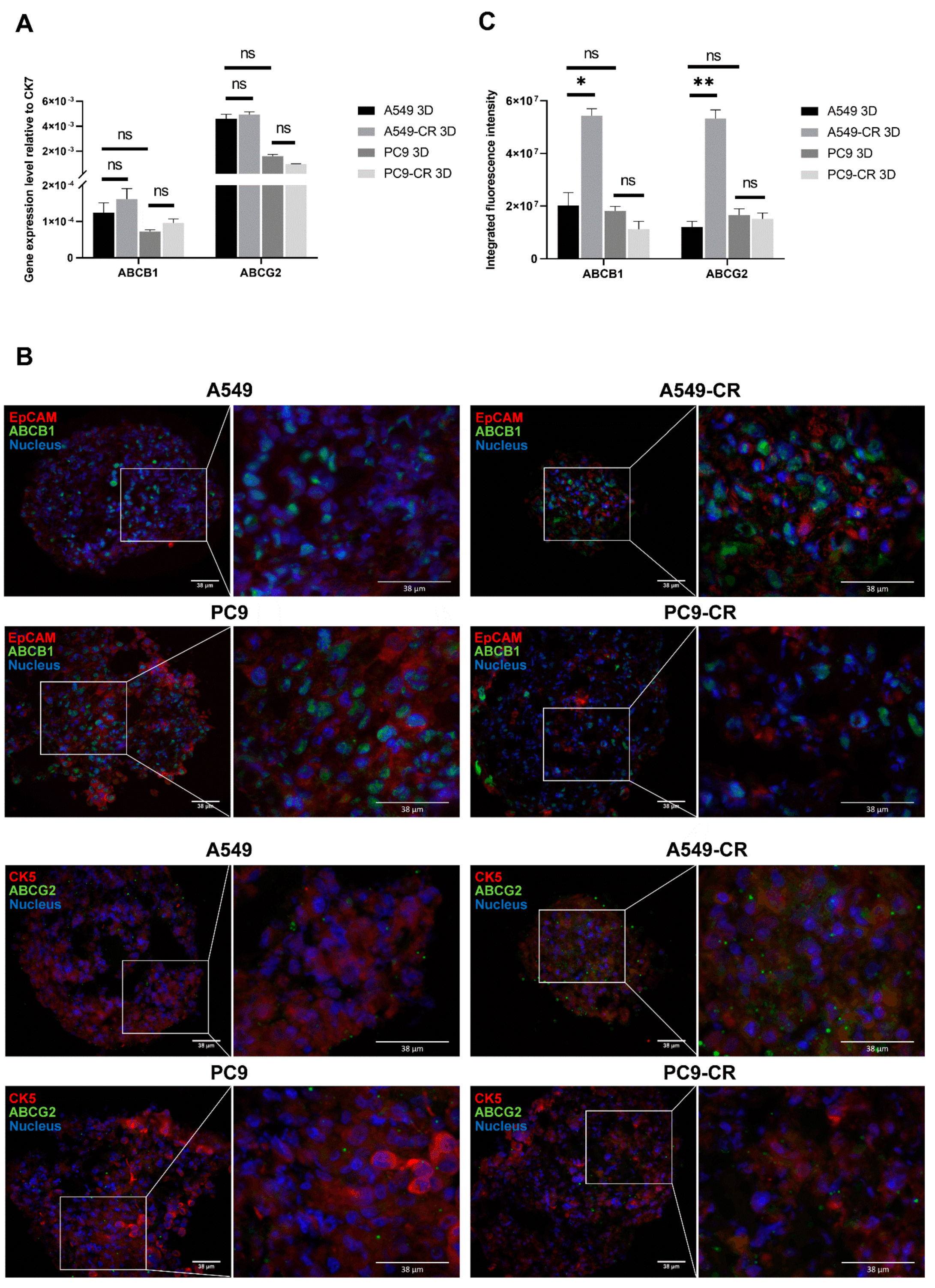
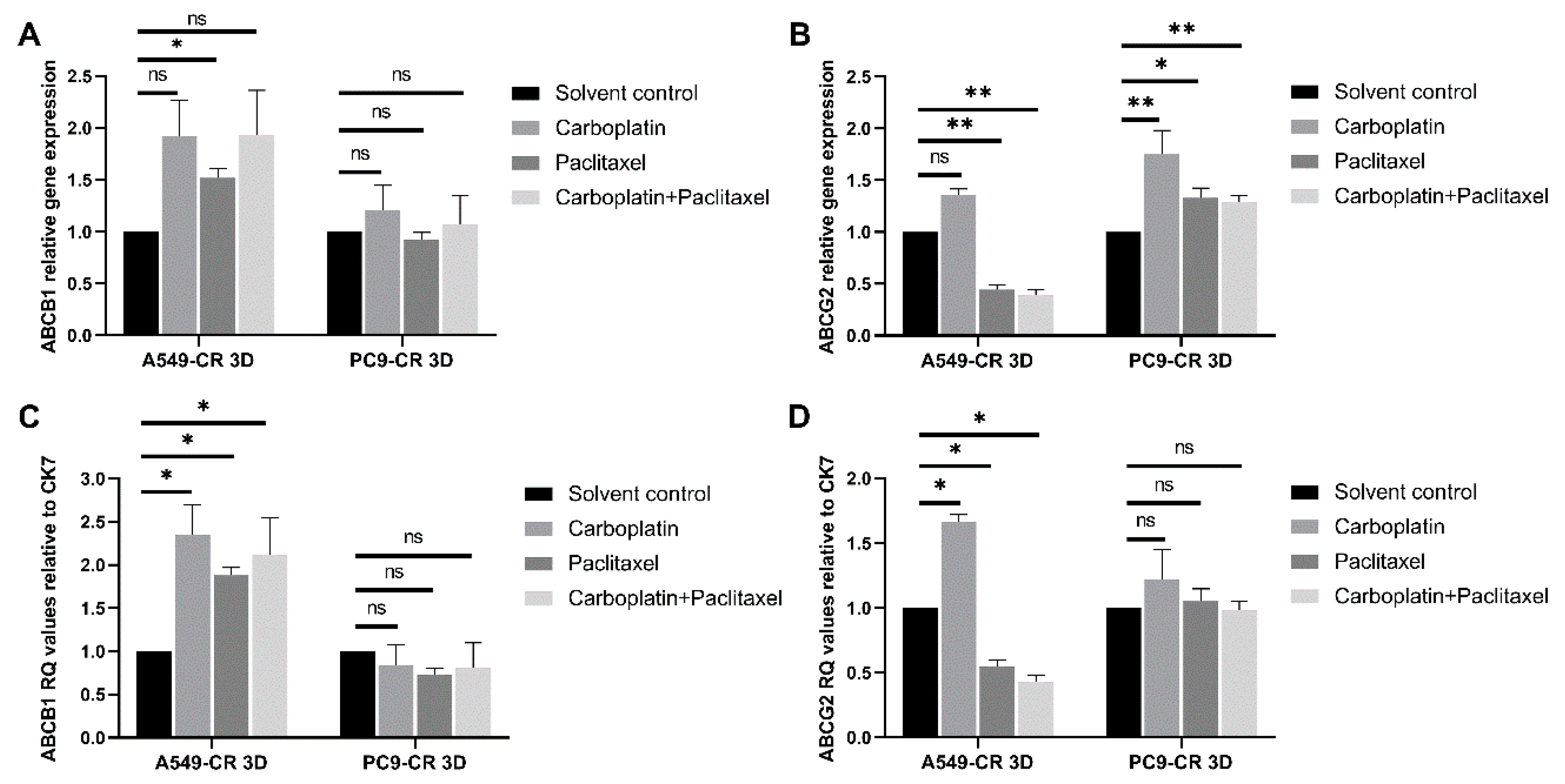
2.3. ABCB1 and ABCG2 Expression in 3D Tissue Aggregates of Cisplatin Resistant and Parental Controls of KRAS and EGFR Mutant Cell Lines
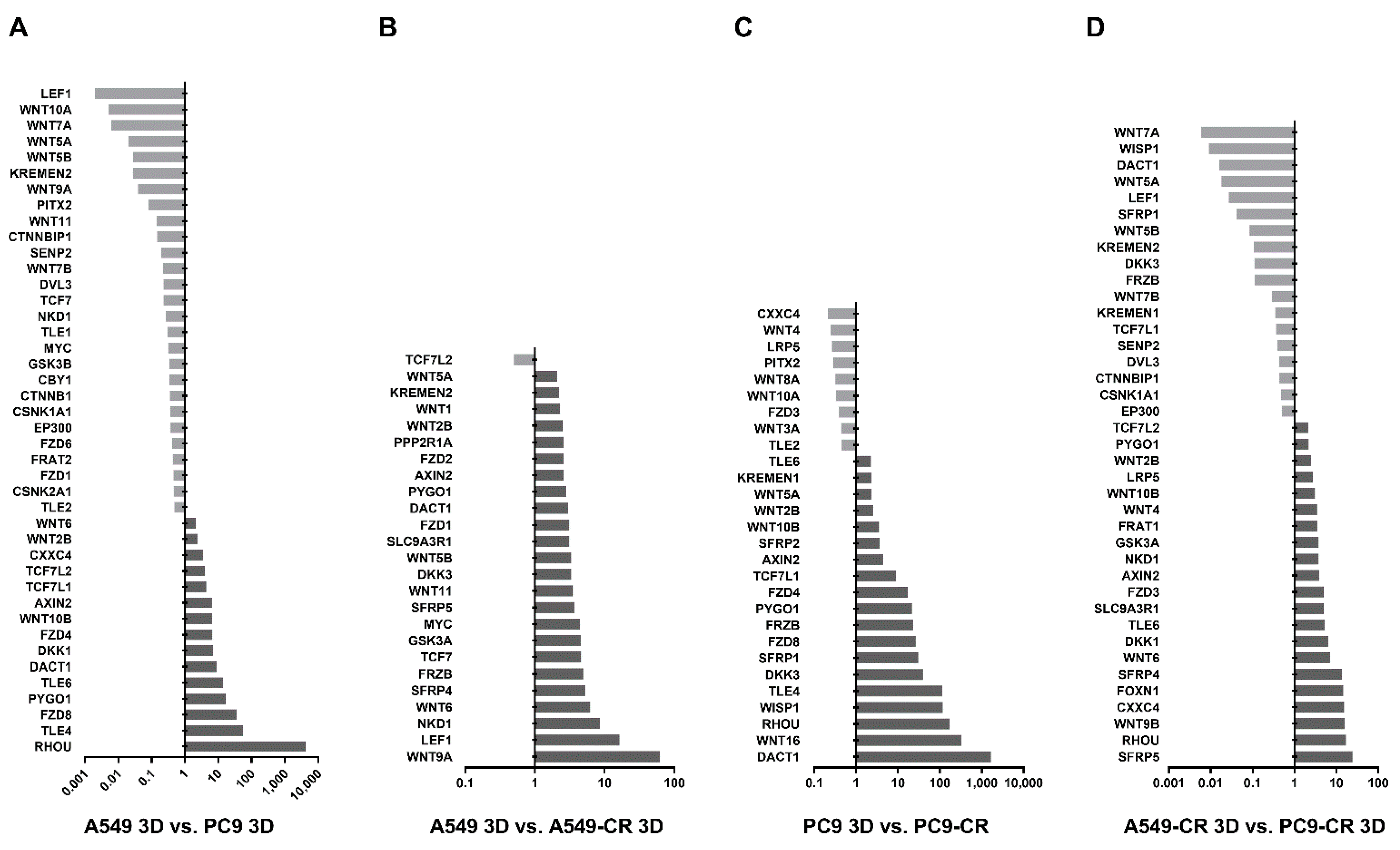
2.4. WNT Signalling in Lung AC Cell Lines Carrying KRAS or EGFR Mutations
3. Discussion
4. Materials and Methods
4.1. Human Lung Cancer Tissues
4.2. Cell Cultures
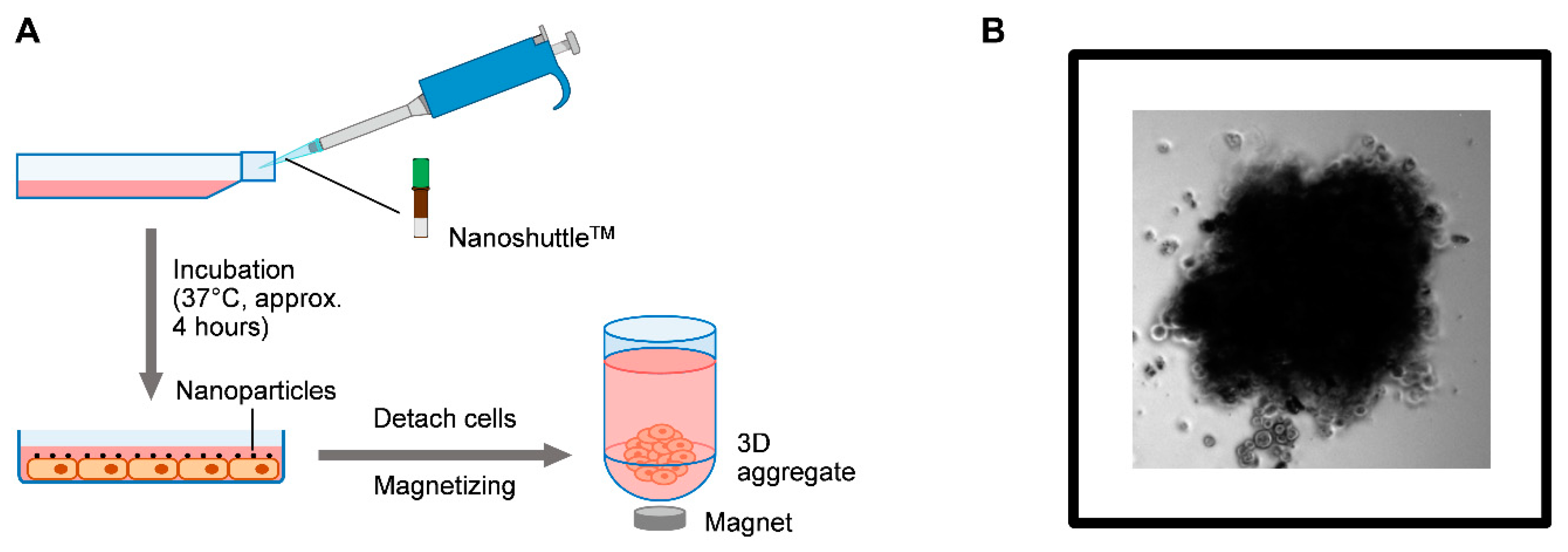
4.3. Three Dimensional (3D) Aggregate Cultures
4.4. Immune-Histochemical Staining of Drug Transporters
4.5. RNA Isolation, cDNA Synthesis and qRT-PCR
4.6. Functionality Test of ABC Drug Transporters
4.7. Drugs and Reagents
4.8. WNT Signalling Arrays
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABC transporter | ATP Binding Cassette Family of transporters |
| ABCB1 | ATP Binding Cassette Subfamily B (MDR/TAP) Member 1 |
| ABCC1 | ATP Binding Cassette Subfamily C Member 1 |
| ABCC2 | ATP Binding Cassette Subfamily C Member 2 |
| ABCC6 | ATP Binding Cassette Subfamily C Member 6 |
| ABCC10 | ATP Binding Cassette Subfamily C Member 10 |
| ABCG2 | ATP Binding Cassette Subfamily G (BCRP) Member 2 |
| AC | adenocarcinoma |
| AE | alveolar epithelium |
| ALK | anaplastic lymphoma kinase |
| BRAF | B-Raf Proto-Oncogene, Serine/Threonine Kinase |
| CR | cisplatin-resistant |
| DACT1 | Dishevelled Binding Antagonist of Beta Catenin 1 |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| DVL | Dishevelled protein |
| EGFR | epidermal growth factor receptor |
| F-12K | Kaighn’s Modification of Ham’s F-12 Medium |
| FCS | foetal calf serum |
| FGM-2 | Fibroblast Growth Medium |
| FZ | Frizzled protein |
| HEPES | 4-(2- hydroxyethyl)-1-piperazineethanesulfonic acid |
| HUVEC | Primary Umbilical Vein Endothelial Cells; Normal, Human |
| JNK | C-Jun N-terminal kinase |
| KRAS | Kirsten rat sarcoma viral oncogene homolog |
| KREMEN2 | Kringle Containing Transmembrane Protein 2 |
| LC | lung cancer |
| LCC | large cell carcinoma |
| LEF1 | lymphoid enhancer-binding factor 1 |
| MAF | multidrug resistance activity factor |
| MDR | multidrug resistance |
| NHLF | normal human lung fibroblasts |
| NKD1 | Naked Cuticle 1 |
| NSCLC | non-small cell lung cancer |
| PDL1 | programmed cell death ligand1 |
| PE | primordial epithelium |
| RHOU | Ras Homolog Family Member U |
| ROS1 | proto-oncogene 1, receptor tyrosine kinase |
| RPMI | Rosewall Park Memorial Institute 1640 Medium |
| SAEC | small airway epithelial cell |
| SCC | squamous cell carcinoma |
| SCLC | small cell lung cancer |
| TCF/LEF | T-cell specific transcription factor/lymphoid enhancer binding factor |
| TKI | tyrosine kinase inhibitor |
| TLE4 | Transducin-like enhancer protein 4 |
| WISP1 | WNT1-inducible-signaling pathway protein 1 |
| WNT | Wingless-Type MMTV Integration Site Family |
| WT | wild type |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249, As part of International Association of Cancer Registries, (IACR). 2020, Global Cancer Observatory. Available online: http://gco.iarc.fr/ (accessed on 20 April 2021). [CrossRef] [PubMed]
- Scott, W.J.; Howington, J.; Feigenberg, S.; Movsas, B.; Pisters, K. American College of Chest Physicians; Treatment of Non-small Cell Lung Cancer Stage I and Stage II: ACCP Evidence-Based Clinical Practice Guidelines (2nd Edition). Chest 2007, 132, 234S–242S. [Google Scholar] [CrossRef]
- Wong, M.C.S.; Lao, X.Q.; Ho, K.F.; Goggins, W.B.; Tse, S.L.A. Incidence and mortality of lung cancer: Global trends and association with socioeconomic status. Sci. Rep. 2017, 7, 14300. [Google Scholar] [CrossRef]
- Ritter, J.H. Tumors of the Lower Respiratory Tract: Atlas of Tumor Pathology. Am. J. Clin. Pathol. 2018, 105, 129–130. [Google Scholar] [CrossRef][Green Version]
- Akinleye, A.; Rasool, Z. Immune checkpoint inhibitors of PD-L1 as cancer therapeutics. J. Hematol. Oncol. 2019, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.A.; Hughes, B.G. Targeted therapy for non-small cell lung cancer: Current standards and the promise of the future. Transl. Lung Cancer Res. 2015, 4, 36–54. [Google Scholar]
- Gettinger, S.; Hellmann, M.D.; Chow, L.Q.M.; Borghaei, H.; Antonia, S.; Brahmer, J.R.; Goldman, J.W.; Gerber, D.E.; Juergens, R.A.; Shepherd, F.A.; et al. Nivolumab Plus Erlotinib in Patients With EGFR-Mutant Advanced NSCLC. J. Thorac. Oncol. 2018, 13, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- Hiley, C.T.; Le Quesne, J.; Santis, G.; Sharpe, R.; de Castro, D.G.; Middleton, G.; Swanton, C. Challenges in molecular testing in non-small-cell lung cancer patients with advanced disease. Lancet 2016, 388, 1002–1011. [Google Scholar] [CrossRef]
- Albaba, H.; Lim, C.; Leighl, N.B. Economic Considerations in the Use of Novel Targeted Therapies for Lung Cancer: Review of Current Literature. Pharmacoeconomics 2017, 35, 1195–1209. [Google Scholar] [CrossRef]
- Youlden, D.R.; Cramb, S.M.; Baade, P.D. The International Epidemiology of Lung Cancer: Geographical distribution and secular trends. J. Thorac. Oncol. 2008, 3, 819–831. [Google Scholar] [CrossRef]
- El-Telbany, A.; Ma, P.C. Cancer Genes in Lung Cancer: Racial Disparities: Are There Any? Genes Cancer 2012, 7–8, 467–480. [Google Scholar] [CrossRef]
- Selamat, S.A.; Chung, B.S.; Girard, L.; Zhang, W.; Zhang, Y.; Campan, M.; Siegmund, K.D.; Koss, M.N.; Hagen, J.A.; Lam, W.L.; et al. Genome-scale analysis of DNA methylation in lung adenocarcinoma and integration with mRNA expression. Genome Res. 2012, 22, 1197–1211. [Google Scholar] [CrossRef] [PubMed]
- Mounawar, M.; Mukeria, A.; Le Calvez, F.; Hung, R.J.; Renard, H.; Cortot, A.; Bollart, C.; Zaridze, D.; Brennan, P.; Boffetta, P.; et al. Patterns of EGFR, HER2, TP53, and KRAS mutations of p14arf expression in non-small cell lung cancers in relation to smoking history. Cancer Res. 2007, 67, 5667–5672. [Google Scholar] [CrossRef]
- Pao, W.; Miller, V.; Zakowski, M.; Doherty, J.; Politi, K.; Sarkaria, I.; Singh, B.; Heelan, R.; Rusch, V.; Fulton, L.; et al. EGF receptor gene mutations are common in lung cancers from “never smokers” and are associated with sensitivity of tumors to gefitinib and erlotinib. Proc. Natl. Acad. Sci. USA 2004, 101, 13306–13311. [Google Scholar] [CrossRef]
- Ahrendt, S.A.; Decker, P.A.; Alawi, E.A.; Zhu Yr, Y.R.; Sanchez-Cespedes, M.; Yang, S.C.; Haasler, G.B.; Kajdacsy-Balla, A.; Demeure, M.J.; Sidransky, D. Cigarette smoking is strongly associated with mutation of the K-ras gene in patients with primary adenocarcinoma of the lung. Cancer 2001, 92, 1525–1530. [Google Scholar] [CrossRef]
- Benesova, L.; Minarik, M.; Jancarikova, D.; Belsanova, B.; Pesek, M. Multiplicity of EGFR and KRAS mutations in non-small cell lung cancer (NSCLC) patients treated with tyrosine kinase inhibitors. Anticancer Res. 2010, 30, 1667–1671. [Google Scholar]
- Kiss, E.; Abdelwahab, E.M.M.; Steib, A.; Papp, E.; Torok, Z.; Jakab, L.; Smuk, G.; Sarosi, V.; Pongracz, J.E. Cisplatin treatment induced interleukin 6 and 8 production alters lung adenocarcinoma cell migration in an oncogenic mutation dependent manner. Respir. Res. 2020, 21, 120. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, P.; Ramlau, R.; Chorostowska-Wynimko, J.; Powrózek, T.; Lewandowska, M.A.; Limon, J.; Wasąg, B.; Pankowski, J.; Kozielski, J.; Kalinka-Warzocha, E. The efficacy of EGFR gene mutation testing in various samples from non-small cell lung cancer patients: A multicenter retrospective study. J. Cancer Res. Clin. Oncol. 2015, 141, 61–68. [Google Scholar] [CrossRef]
- Hall, M.D.; Okabe, M.; Shen, D.W.; Liang, X.J.; Gottesman, M.M. The role of cellular accumulation in determining sensitivity to platinum-based chemotherapy. Annu. Rev. Pharmacol. Toxicol. 2008, 48, 495–535. [Google Scholar] [CrossRef] [PubMed]
- Wheate, N.J.; Walker, S.; Craig, G.E.; Oun, R. The status of platinum anticancer drugs in the clinic and in clinical trials. Dalton Trans. 2010, 39, 8113–8127. [Google Scholar] [CrossRef]
- Chen, J.; Emara, N.; Solomides, C.; Parekh, H.; Simpkins, H. Resistance to platinum-based chemotherapy in lung cancer cell lines. Cancer Chemother. Pharmacol. 2010, 66, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Tsvetkova, E.; Goss, G. Drug resistance and its significance for treatment decisions in non-small-cell lung cancer. Curr. Oncol. 2012, 19 (Suppl. 1), S45–S51. [Google Scholar] [CrossRef]
- Lemjabbar-Alaoui, H.; Hassan, O.; Yang, Y.W.; Buchanan, P. Lung cancer: Biology and treatment options. Biochim. Biophys. Acta 2015, 1856, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Linton, K.J. Structure and function of ABC transporters. Physiology 2007, 22, 122–130. [Google Scholar] [CrossRef]
- Locher, K.P. Structure and mechanism of ATP-binding cassette transporters. Philos. Trans. R. Soc. B Biol. Sci. 2009, 1514, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Sharom, F.J. The P-glycoprotein multidrug transporter. Essays Biochem. 2011, 50, 161–178. [Google Scholar] [CrossRef]
- Fletcher, J.I.; Haber, M.; Henderson, M.J.; Norris, M.D. ABC transporters in cancer: More than just drug efflux pumps. Nat. Rev. Cancer 2010, 10, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.L.; Patel, A.; Kumar, P.; Chen, Z.S. Role of ABC transporters in cancer chemotherapy. Chin. J. Cancer 2012, 2, 51–57. [Google Scholar] [CrossRef]
- Chen, B.; Zheng, J.; Zeng, Y.; Li, B.; Xie, B.; Zheng, J.; Zhou, J.; Zhang, W. Sequence-dependent antiproliferative effects of gefitinib and docetaxel on non-small cell lung cancer (NSCLC) cells and the possible mechanism. PLoS ONE 2014, 9, e114074. [Google Scholar] [CrossRef]
- Vesel, M.; Rapp, J.; Feller, D.; Kiss, E.; Jaromi, L.; Meggyes, M.; Miskei, G.; Duga, B.; Smuk, G.; Laszlo, T.; et al. ABCB1 and ABCG2 drug transporters are differentially expressed in non-small cell lung cancers (NSCLC) and expression is modified by cisplatin treatment via altered Wnt signaling. Respir. Res. 2017, 18, 52. [Google Scholar] [CrossRef]
- Zhou, S.F.; Wang, L.L.; Di, Y.M.; Xue, C.C.; Duan, W.; Li, C.G.; Li, Y. Substrates and inhibitors of human multidrug resistance associated proteins and the implications in drug development. Curr. Med. Chem. 2008, 15, 1981–2039. [Google Scholar] [CrossRef] [PubMed]
- Sprowl, J.; Gregorc, V.; Lazzari, C.; Mathijssen, R.H.; Loos, W.J.; Sparreboom, A. Association of ABCC2 polymorphisms with cisplatin disposition and efficacy. Clin. Pharm. 2012, 91, 1022–1026. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.K.; Wang, Y.J.; Gupta, P.; Chen, Z.S. Multidrug Resistance Proteins (MRPs) and Cancer Therapy. AAPS J. 2015, 17, 802–812. [Google Scholar] [CrossRef]
- Yin, J.Y.; Huang, Q.; Yang, Y.; Zhang, J.T.; Zhong, M.Z.; Zhou, H.H.; Liu, Z.Q. Characterization and analyses of multidrug resistance-associated protein 1 (MRP1/ABCC1) polymorphisms in Chinese population. Pharm. Genom. 2009, 19, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.K.; To, K.K.; Huang, L.Y.; Xu, J.H.; Yang, K.; Wang, F.; Huang, Z.C.; Ye, S.; Fu, L.W. Afatinib circumvents multidrug resistance via dually inhibiting ATP binding cassette subfamily G member 2 in vitro and in vivo. Oncotarget 2014, 5, 11971–11985. [Google Scholar] [CrossRef]
- Oguri, T.; Ozasa, H.; Uemura, T.; Bessho, Y.; Miyazaki, M.; Maeno, K.; Maeda, H.; Sato, S.; Ueda, R. MRP7/ABCC10 expression is a predictive biomarker for the resistance to paclitaxel in non-small cell lung cancer. Mol. Cancer 2008, 7, 1150–1155. [Google Scholar] [CrossRef] [PubMed]
- Hopper-Borge, E.A.; Churchill, T.; Paulose, C.; Nicolas, E.; Jacobs, J.D.; Ngo, O.; Kuang, Y.; Grinberg, A.; Westphal, H.; Chen, Z.S.; et al. Contribution of Abcc10 (Mrp7) to in vivo paclitaxel resistance as assessed in Abcc10(-/-) mice. Cancer Res. 2011, 71, 3649–3657. [Google Scholar] [CrossRef]
- Sun, Y.L.; Chen, J.J.; Kumar, P.; Chen, K.; Sodani, K.; Patel, A.; Chen, Y.L.; Chen, S.D.; Jiang, W.Q.; Chen, Z.S. Reversal of MRP7 (ABCC10)-mediated multidrug resistance by tariquidar. PLoS ONE 2013, 8, e55576. [Google Scholar] [CrossRef]
- Scannell, J.W.; Blanckley, A.; Boldon, H.; Warrington, B. Diagnosing the decline in pharmaceutical R&D efficiency. Nat. Rev. Drug Discov. 2012, 11, 191–200. [Google Scholar]
- Yildirim, O.; Gottwald, M.; Schüler, P.; Michel, M.C. Opportunities and Challenges for Drug Development: Public–Private Partnerships, Adaptive Designs and Big Data. Front. Pharmacol. 2016, 7, 461. [Google Scholar] [CrossRef]
- Fang, Y.; Eglen, R.M. Three-Dimensional Cell Cultures in Drug Discovery and Development. SLAS Discov. 2017, 5, 456–472. [Google Scholar]
- Centeno, E.G.Z.; Cimarosti, H.; Bithell, A. 2D versus 3D human induced pluripotent stem cell-derived cultures for neurodegenerative disease modelling. Mol. Neurodegener. 2018, 13, 1–15. [Google Scholar] [CrossRef]
- Central South Coast Cancer Network (CSCCN). Lung Chemotherapy Protocols. 2014. Updated: 2020. Available online: https://www.uhs.nhs.uk/HealthProfessionals/Chemotherapy-protocols/Lung-chemotherapy-protocols.aspx (accessed on 20 April 2021).
- Moldvay, J.; Rokszin, G.; Abonyi-Tóth, Z.; Katona, L.; Fábián, K.; Kovács, G. Lung cancer drug therapy in Hungary—3-year experience. Onco Targets Ther. 2015, 8, 1031–1038. [Google Scholar]
- Haymarket Media, I. The National Comprehensive Cancer Network Guidelines® (NCCN) Non-Small Cell Lung Cancer Treatment Regimens. Non-Small Cell Lung Cancer Guidelines v 7.2019. 2019. Available online: https://www.cancertherapyadvisor.com/home/cancer-topics/lung-cancer/lung-cancer-treatment-regimens-landing-page/non-small-cell-lung-cancer-treatment-regimens/ (accessed on 20 April 2021).
- Hung, T.H.; Hsu, S.C.; Cheng, C.Y.; Choo, K.B.; Tseng, C.P.; Chen, T.C.; Lan, Y.W.; Huang, T.T.; Lai, H.C.; Chen, C.M. Wnt5A regulates ABCB1 expression in multidrug-resistant cancer cells through activation of the non-canonical PKA/beta-catenin pathway. Oncotarget 2014, 5, 12273–12290. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, D.; Kuai, J.; Cheng, M.; Fang, X.; Li, G. Upregulation of miR-199a/b contributes to cisplatin resistance via Wnt/beta-catenin-ABCG2 signaling pathway in ALDHA1(+) colorectal cancer stem cells. Tumour Biol. 2017, 39, 1010428317715155. [Google Scholar] [CrossRef]
- Wang, Q.; Geng, F.; Zhou, H.; Chen, Y.; Du, J.; Zhang, X.; Song, D.; Zhao, H. MDIG promotes cisplatin resistance of lung adenocarcinoma by regulating ABC transporter expression via activation of the WNT/beta-catenin signaling pathway. Oncol. Lett. 2019, 18, 4294–4307. [Google Scholar] [PubMed]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug Resistance in Cancer: An Overview. Cancers 2014, 3, 1769–1792. [Google Scholar] [CrossRef] [PubMed]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Zawadzka, I.; Jeleń, A.; Pietrzak, J.; Żebrowska-Nawrocka, M.; Michalska, K.; Szmajda-Krygier, D.; Mirowski, M.; Łochowski, M.; Kozak, J.; Balcerczak, E. The impact of ABCB1 gene polymorphism and its expression on non-small-cell lung cancer development, progression and therapy—preliminary report. Sci. Rep. 2020, 10, 6188. [Google Scholar] [CrossRef] [PubMed]
- Dohse, M.; Scharenberg, C.; Shukla, S.; Robey, R.W.; Volkmann, T.; Deeken, J.F.; Brendel, C.; Ambudkar, S.V.; Neubauer, A.; Bates, S.E. Comparison of ATP-binding cassette transporter interactions with the tyrosine kinase inhibitors imatinib, nilotinib, and dasatinib. Drug Metab. Dispos. 2010, 38, 1371–1380. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, A.; Sawers, L.; Gannon, A.L.; Chakravarty, P.; Scott, A.L.; Bray, S.E.; Ferguson, M.J.; Smith, G. ABCB1 (MDR1) induction defines a common resistance mechanism in paclitaxel- and olaparib-resistant ovarian cancer cells. Br. J. Cancer 2016, 115, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Johnatty, S.E.; Beesley, J.; Paul, J.; Fereday, S.; Spurdle, A.B.; Webb, P.M.; Byth, K.; Marsh, S.; McLeod, H. AOCS Study Group ABCB1 (MDR 1) polymorphisms and progression-free survival among women with ovarian cancer following paclitaxel/carboplatin chemotherapy. Clin. Cancer Res. 2008, 14, 5594–5601. [Google Scholar] [CrossRef]
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug resistance in cancer: Role of ATP-dependent transporters. Nat. Rev. Cancer 2002, 2, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Ohnuma, S.; Ambudkar, S.V. Improving cancer chemotherapy with modulators of ABC drug transporters. Curr. Drug Targets 2011, 12, 621–630. [Google Scholar] [CrossRef]
- Fox, E.; Bates, S.E. Tariquidar (XR9576): A P-glycoprotein drug efflux pump inhibitor. Expert Rev. Anticancer 2007, 7, 447–459. [Google Scholar] [CrossRef]
- Kelly, R.J.; Draper, D.; Chen, C.C.; Robey, R.W.; Figg, W.D.; Piekarz, R.L.; Chen, X.; Gardner, E.R.; Balis, F.M.; Venkatesan, A.M.; et al. A Pharmacodynamic Study of Docetaxel in Combination with the P-glycoprotein Antagonist, Tariquidar (XR9576) in Patients with Lung, Ovarian, and Cervical Cancer. Clin. Cancer Res. 2011, 17, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Russell, A.; Beesley, J.; Chen, X.Q.; Healey, S.; Henderson, M.; Wong, M.; Emmanuel, C.; Galletta, L.; Johnatty, S.E.; et al. Paclitaxel sensitivity in relation to ABCB1 expression, efflux and single nucleotide polymorphisms in ovarian cancer. Sci. Rep. 2014, 4, 4669. [Google Scholar] [CrossRef]
- Rapp, J.; Jaromi, L.; Kvell, K.; Miskei, G.; Pongracz, J.E. WNT signalling—lung cancer is no exception. Respir. Res. 2017, 18, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Pongracz, J.E.; Stockley, R.A. Wnt signalling in lung development and diseases. Respir. Res. 2006, 7, 15. [Google Scholar] [CrossRef]
- Cardama, G.A.; Gonzalez, N.; Maggio, J.; Menna, P.L.; Gomez, D.E. Rho GTPases as therapeutic targets in cancer (Review). Int. J. Oncol. 2020, 51, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, T.Y.; Peng, H.T.; Wu, Y.Q.; Zhang, L.L.; Lin, X.H.; Lai, Y.H. Up-regulation of Wnt7b rather than Wnt1, Wnt7a, and Wnt9a indicates poor prognosis in breast cancer. Int. J. Clin. Exp. Pathol. 2018, 11, 4552–4561. [Google Scholar] [PubMed]
- Yuan, G.; Wang, C.; Ma, C.; Chen, N.; Tian, Q.; Zhang, T.; Fu, W. Oncogenic function of DACT1 in colon cancer through the regulation of beta-catenin. PLoS ONE 2012, 7, e34004. [Google Scholar]
- Cheyette, B.N.; Waxman, J.S.; Miller, J.R.; Takemaru, K.; Sheldahl, L.C.; Khlebtsova, N.; Fox, E.P.; Earnest, T.; Moon, R.T. Dapper, a Dishevelled-associated antagonist of beta-catenin and JNK signaling, is required for notochord formation. Dev. Cell 2002, 2, 449–461. [Google Scholar] [CrossRef]
- Yin, X.; Xiang, T.; Li, L.; Su, X.; Shu, X.; Luo, X.; Huang, J.; Yuan, Y.; Peng, W.; Oberst, M.; et al. DACT1, an antagonist to Wnt/β-catenin signaling, suppresses tumor cell growth and is frequently silenced in breast cancer. Breast Cancer Res. 2013, 2, R23. [Google Scholar] [CrossRef]
- Tuo, H.; Wang, Y.; Wang, L.; Yao, B.; Li, Q.; Wang, C.; Liu, Z.; Han, S.; Yin, G.; Tu, K.; et al. MiR-324-3p promotes tumor growth through targeting DACT1 and activation of Wnt/β-catenin pathway in hepatocellular carcinoma. Oncotarget 2017, 8, 65687–65698. [Google Scholar] [CrossRef]
- Yau, T.O.; Chan, C.Y.; Chan, K.L.; Lee, M.F.; Wong, C.M.; Fan, S.T.; Ng, I.O. HDPR1, a novel inhibitor of the WNT/beta-catenin signaling, is frequently downregulated in hepatocellular carcinoma: Involvement of methylation-mediated gene silencing. Oncogene 2005, 24, 1607–1614. [Google Scholar] [CrossRef]
- Yang, Z.Q.; Zhao, Y.; Liu, Y.; Zhang, J.Y.; Zhang, S.; Jiang, G.Y.; Zhang, P.X.; Yang, L.H.; Liu, D.; Li, Q.C.; et al. Downregulation of HDPR1 is associated with poor prognosis and affects expression levels of p120-catenin and beta-catenin in nonsmall cell lung cancer. Mol. Carcinog. 2010, 49, 508–519. [Google Scholar] [CrossRef]
- Sun, Y.; Zhu, D.; Chen, F.; Qian, M.; Wei, H.; Chen, W.; Xu, J. SFRP2 augments WNT16B signaling to promote therapeutic resistance in the damaged tumor microenvironment. Oncogene 2016, 35, 4321–4334. [Google Scholar] [CrossRef]
- Yao, X.; Ireland, S.K.; Pham, T.; Temple, B.; Chen, R.; Raj, M.H.; Biliran, H. TLE1 promotes EMT in A549 lung cancer cells through suppression of E-cadherin. Biochem. Biophys. Res. Commun. 2014, 455, 277–284. [Google Scholar] [CrossRef]
- Wang, S.Y.; Gao, K.; Deng, D.L.; Cai, J.J.; Xiao, Z.Y.; He, L.Q.; Jiao, H.L.; Ye, Y.P.; Yang, R.W.; Li, T.T.; et al. TLE4 promotes colorectal cancer progression through activation of JNK/c-Jun signaling pathway. Oncotarget 2016, 7, 2878–2888. [Google Scholar] [CrossRef]
- Yang, R.-W.; Zeng, Y.-Y.; Wei, W.-T.; Cui, Y.M.; Sun, H.Y.; Cai, Y.L.; Nian, X.X.; Hu, Y.T.; Quan, Y.P.; Jiang, S.L.; et al. TLE3 represses colorectal cancer proliferation by inhibiting MAPK and AKT signaling pathways. J. Exp. Clin. Cancer Res. 2016, 35, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yin, J.; Li, X.; Wang, Y.; Zheng, Y.; Qian, C.; Xiao, L.; Zou, T.; Wang, Z.; Liu, J.; et al. WISP1 Polymorphisms Contribute to Platinum-Based Chemotherapy Toxicity in Lung Cancer Patients. Int. J. Mol. Sci. 2014, 15, 21011–21027. [Google Scholar] [CrossRef]
- Stewart, D.J. Wnt Signaling Pathway in Non-Small Cell Lung Cancer. J. Nat. Cancer Inst. 2014, 106, djt356. [Google Scholar] [CrossRef] [PubMed]
- Santiago, L.; Daniels, G.; Wang, D.; Deng, F.M.; Lee, P. Wnt signaling pathway protein LEF1 in cancer, as a biomarker for prognosis and a target for treatment. Am. J. Cancer Res. 2017, 7, 1389–1406. [Google Scholar]
- Rousset, R.; Mack, J.A.; Wharton, K.A., Jr.; Axelrod, J.D.; Cadigan, K.M.; Fish, M.P.; Nusse, R.; Scott, M.P. Naked cuticle targets dishevelled to antagonize Wnt signal transduction. Genes Dev. 2001, 15, 658–671. [Google Scholar] [CrossRef] [PubMed]
- Van Raay, T.J.; Fortino, N.J.; Miller, B.W.; Ma, H.; Lau, G.; Li, C.; Franklin, J.L.; Attisano, L.; Solnica-Krezel, L.; Coffey, R.J. Naked1 antagonizes Wnt signaling by preventing nuclear accumulation of beta-catenin. PLoS ONE 2011, 6, e18650. [Google Scholar] [CrossRef]
- Larraguibel, J.; Weiss, A.R.E.; Pasula, D.J.; Dhaliwal, R.S.; Kondra, R.; Van Raay, T.J. Wnt ligand–dependent activation of the negative feedback regulator Nkd1. Mol. Biol. Cell 2015, 26, 2375–2384. [Google Scholar] [CrossRef] [PubMed]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 2001. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Mutation | Histology | T | N | M | Age | Gender |
|---|---|---|---|---|---|---|---|
| 1 | EGFR/KRAS WT | AC | T2 | N1 | Mx | 65 | F |
| 2 | EGFR/KRAS WT | AC | T1 | N1 | Mx | 69 | M |
| 3 | EGFR MUTANT | AC | T2b | N1 | Mx | 73 | F |
| 4 | EGFR MUTANT | AC | T1 | N1 | Mx | 60 | M |
| 5 | KRAS MUTANT | AC | T1 | N1b | Mx | 65 | M |
| 6 | KRAS MUTANT | AC | T2b | N2 | M0 | 62 | F |
| 7 | KRAS MUTANT | AC | T1 | N2 | Mx | 51 | F |
| 8 | KRAS MUTANT | AC | T3 | N2 | Mx | 57 | F |
| 9 | KRAS MUTANT | AC | T2 | N0 | Mx | 72 | M |
| 10 | KRAS MUTANT | AC | T2 | N2 | Mx | 62 | M |
| 11 | KRAS MUTANT | AC | T2 | N2 | Mx | 68 | M |
| 12 | KRAS MUTANT | AC | T2 | N1 | Mx | 59 | M |
| Target | Forward Primer | Reverse Primer |
|---|---|---|
| human beta-actin | 5′-GCGCGGCTACAGCTTCA-3′ | 5′-CTTAATGTCACGCACGATTTCC-3′ |
| human ABCB1 | 5′-GCAGCTGGAAGACAAATACACAA-3′ | 5′-CCCAACATCGTGCACATCA-3′ |
| human ABCG2 | 5′-AACCTGGTCTCAACGCCATC-3′ | 5′-GTCGCGGTGCTCCATTTATC-3′ |
| human ABCC1 | 5′-GCTGGAGTGTGTGGGCAACT-3′ | 5′-CTGAGGCGTTGCCTGGAGAT-3′ |
| human ABCC2 | 5′-GCAAACTGTTCTGGTGTGGGA-3′ | 5′-CCAGCTCTATGGCTGCTAGA-3′ |
| human ABCC6 | 5′-GAATGGCCTGGTGTTTGCAG-3′ | 5′-CAGTTGCGAACAACCCACTG-3′ |
| human ABCC10 | 5′-AACGCTTTGCCAACAAGACA-3′ | 5′-CCAGCACCCGGTCTGAGTT-3′ |
| human CK7 | 5′-AGGATGTGGATGCTGCCTAC-3′ | 5′-GGGACTGCAGCTCTGTCAAC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaromi, L.; Csongei, V.; Vesel, M.; Abdelwahab, E.M.M.; Soltani, A.; Torok, Z.; Smuk, G.; Sarosi, V.; Pongracz, J.E. KRAS and EGFR Mutations Differentially Alter ABC Drug Transporter Expression in Cisplatin-Resistant Non-Small Cell Lung Cancer. Int. J. Mol. Sci. 2021, 22, 5384. https://doi.org/10.3390/ijms22105384
Jaromi L, Csongei V, Vesel M, Abdelwahab EMM, Soltani A, Torok Z, Smuk G, Sarosi V, Pongracz JE. KRAS and EGFR Mutations Differentially Alter ABC Drug Transporter Expression in Cisplatin-Resistant Non-Small Cell Lung Cancer. International Journal of Molecular Sciences. 2021; 22(10):5384. https://doi.org/10.3390/ijms22105384
Chicago/Turabian StyleJaromi, Luca, Veronika Csongei, Monika Vesel, ElHusseiny Mohamed Mahmud Abdelwahab, Amina Soltani, Zsofia Torok, Gabor Smuk, Veronika Sarosi, and Judit Erzsebet Pongracz. 2021. "KRAS and EGFR Mutations Differentially Alter ABC Drug Transporter Expression in Cisplatin-Resistant Non-Small Cell Lung Cancer" International Journal of Molecular Sciences 22, no. 10: 5384. https://doi.org/10.3390/ijms22105384
APA StyleJaromi, L., Csongei, V., Vesel, M., Abdelwahab, E. M. M., Soltani, A., Torok, Z., Smuk, G., Sarosi, V., & Pongracz, J. E. (2021). KRAS and EGFR Mutations Differentially Alter ABC Drug Transporter Expression in Cisplatin-Resistant Non-Small Cell Lung Cancer. International Journal of Molecular Sciences, 22(10), 5384. https://doi.org/10.3390/ijms22105384