
Dual Actions of A2A and A3 Adenosine Receptor Ligand Prevents Obstruction-Induced Kidney Fibrosis in Mice



Abstract
:1. Introduction
2. Results
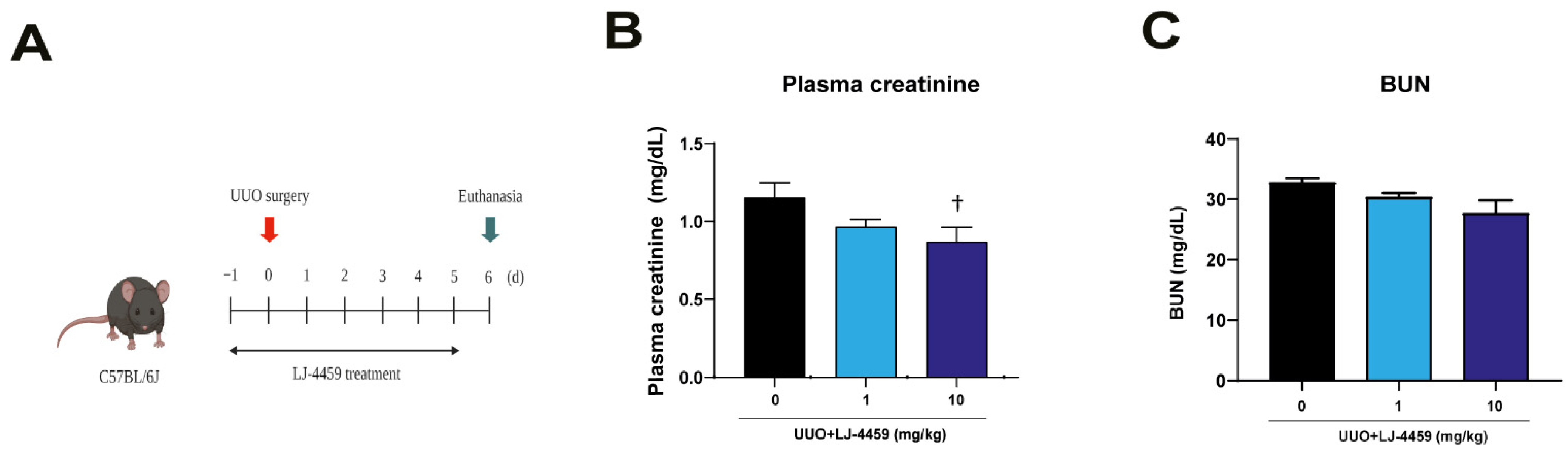
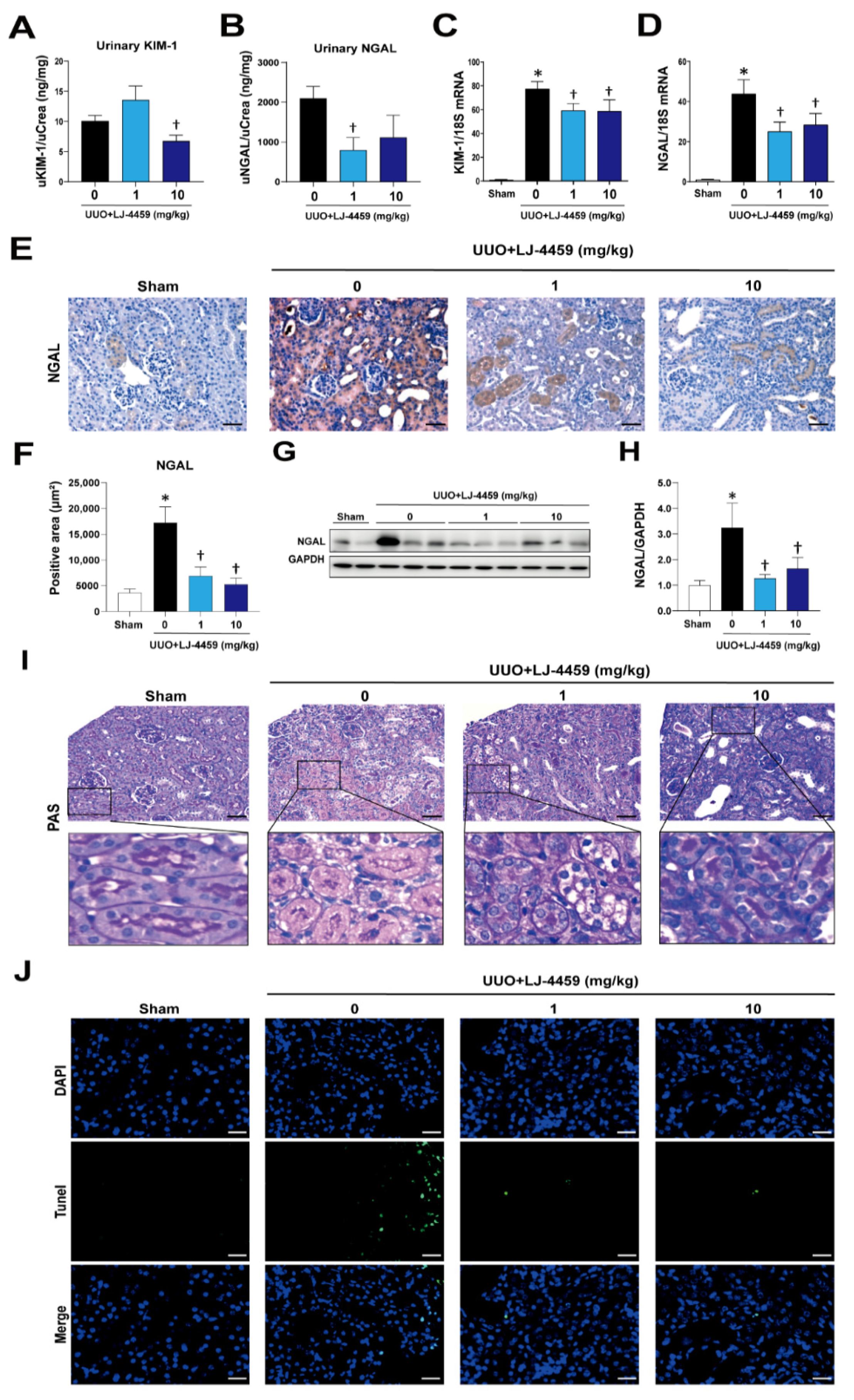
2.1. Pretreatment of LJ-4459 Improves Kidney Function and Attenuates Kidney Tubular Injury in the Obstructed Kidney
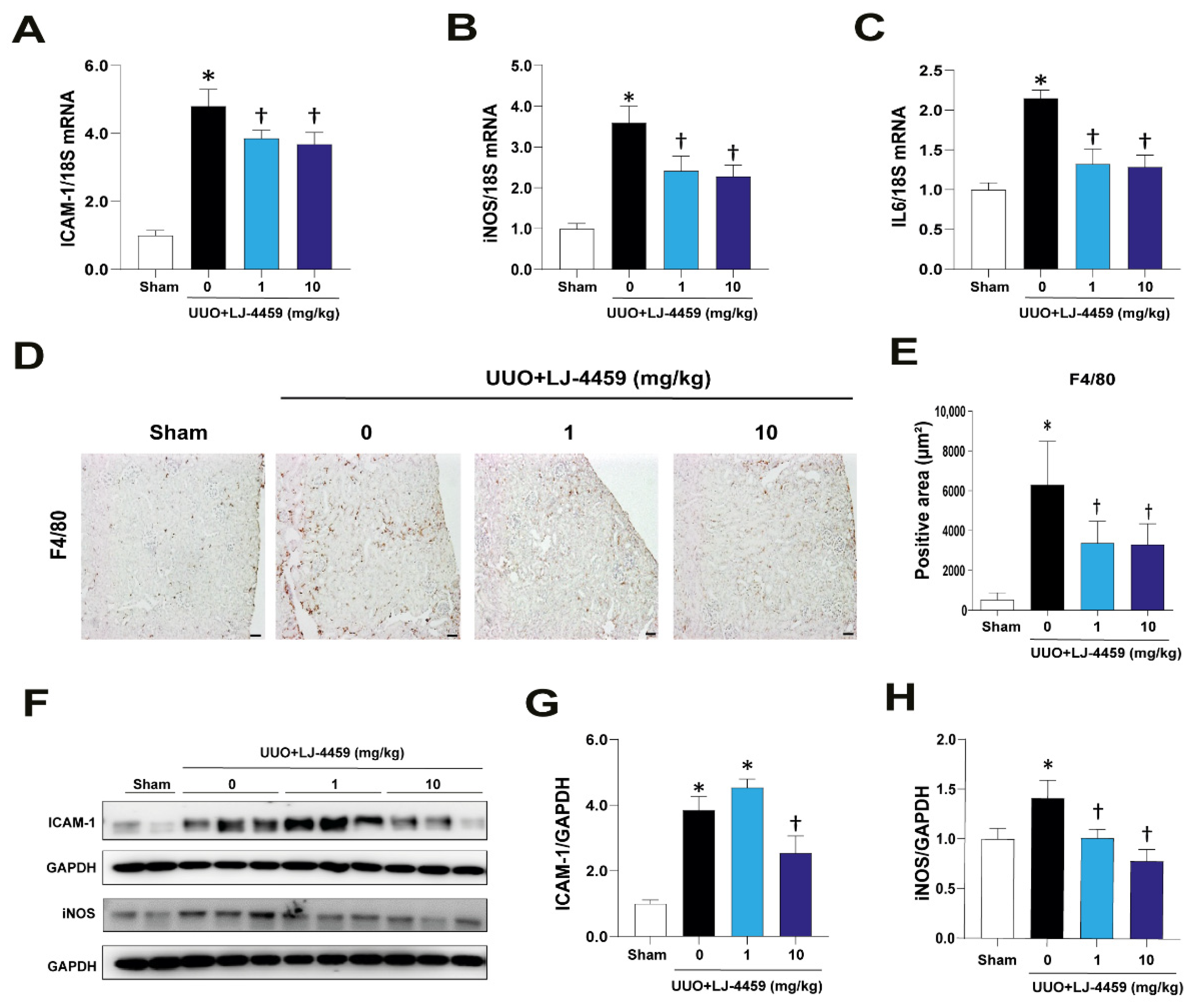
2.2. Pretreatment of LJ-4459 Decreases Kidney Inflammation in the Obstructed Kidney
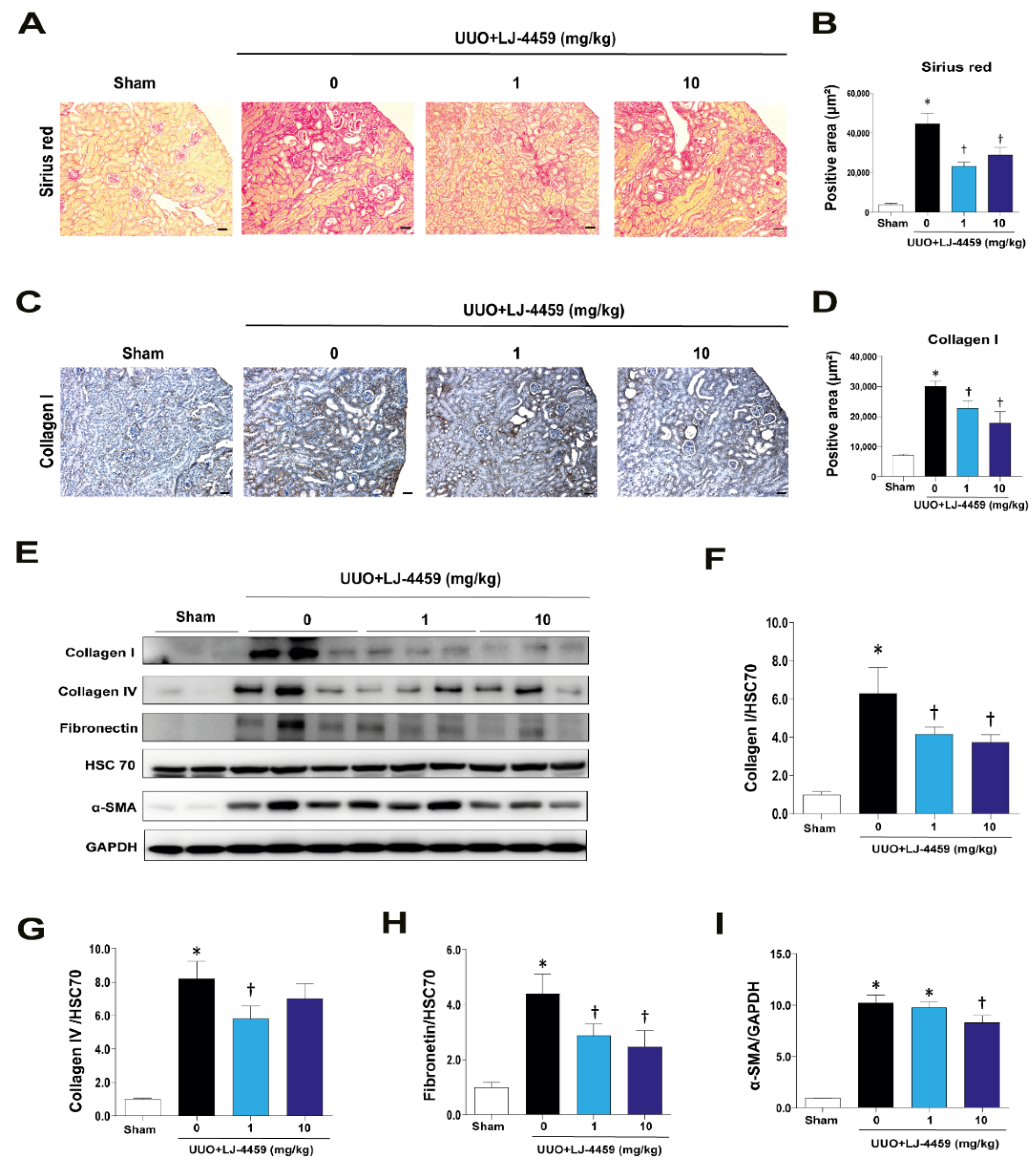
2.3. Pretreatment with LJ-4459 Inhibits Tubulointerstitial Fibrosis in the Obstructed Kidney
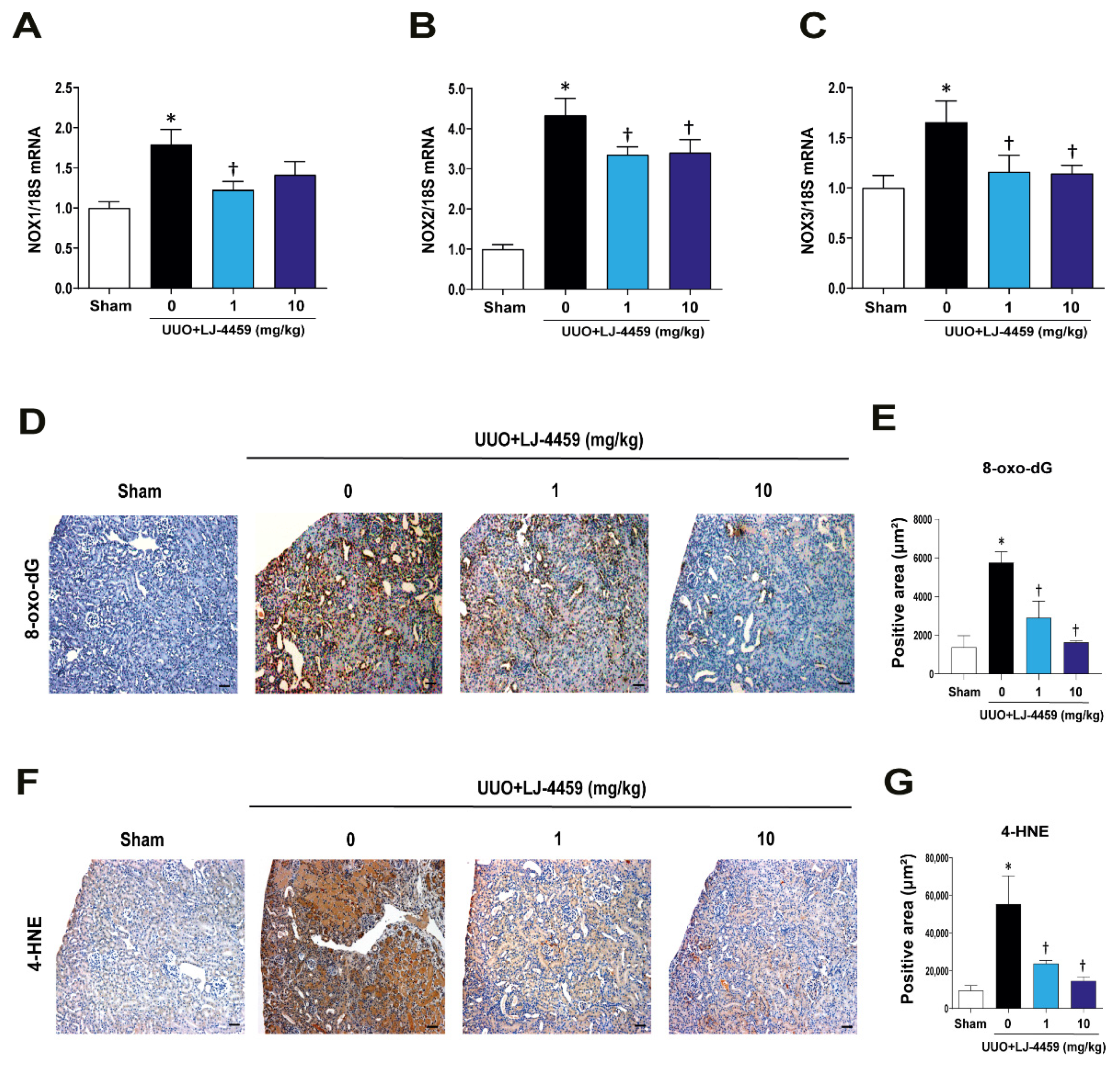
2.4. Pretreatment with LJ-4459 Reduces Oxidative Stress in the Obstructed Kidney
2.5. Pretreatment with LJ-4459 Suppresses ERK and NF-κB Phosphorylation in the Obstructed Kidney
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animal Experiments
4.3. Measurements of Blood Parameters
4.4. Measurements of Urine Parameters
4.5. Histology and Immunohistochemistry
4.6. Terminal Transferase-dUTP-Nick-End Labeling (TUNEL) Assay
4.7. Western Blot Analysis
4.8. Quantitative Real Time Reverse Transcriptase Polymerase Chain Reaction
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Y. Renal fibrosis: New insights into the pathogenesis and therapeutics. Kidney Int. 2006, 69, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Hewitson, T.D. Fibrosis in the kidney: Is a problem shared a problem halved? Fibrogenesis Tissue Repair 2012, 5, S14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlondorff, D.O. Overview of factors contributing to the pathophysiology of progressive renal disease. Kidney Int. 2008, 74, 860–866. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y. Cellular and molecular mechanisms of renal fibrosis. Nat. Rev. Nephrol. 2011, 7, 684–696. [Google Scholar] [CrossRef] [PubMed]
- Eltzschig, H.K. Adenosine: An old drug newly discovered. Anesthesiology 2009, 111, 904–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredholm, B.B.; Ijzerman, A.P.; Jacobson, K.A.; Klotz, K.N.; Linden, J. International Union of Pharmacology. XXV. Nomenclature and classification of adenosine receptors. Pharmacol. Rev. 2001, 53, 527–552. [Google Scholar]
- Jacobson, K.A.; Gao, Z.-G. Adenosine receptors as therapeutic targets. Nat. Rev. Drug Discov. 2006, 5, 247–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallon, V.; Mühlbauer, B.; Osswald, H. Adenosine and Kidney Function. Physiol. Rev. 2006, 86, 901–940. [Google Scholar] [CrossRef] [Green Version]
- Oyarzún, C.; Garrido, W.; Alarcón, S.; Yáñez, A.; Sobrevia, L.; Quezada, C.; Martín, R.S. Adenosine contribution to normal renal physiology and chronic kidney disease. Mol. Asp. Med. 2017, 55, 75–89. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Zhang, W.; Wen, J.; Zhang, Y.; Kellems, R.E.; Xia, Y. A2B Adenosine Receptor-Mediated Induction of IL-6 Promotes CKD. J. Am. Soc. Nephrol. 2011, 22, 890–901. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Jiang, X.; Zhou, Y.; Xia, B.; Dai, Y. Increased adenosine levels contribute to ischemic kidney fibrosis in the unilateral ureteral obstruction model. Exp. Ther. Med. 2015, 9, 737–743. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Hwang, I.; Lee, J.H.; Lee, H.W.; Jeong, L.-S.; Ha, H. The Selective A3AR Antagonist LJ-1888 Ameliorates UUO-Induced Tubulointerstitial Fibrosis. Am. J. Pathol. 2013, 183, 1488–1497. [Google Scholar] [CrossRef]
- Dorotea, D.; Cho, A.; Lee, G.; Kwon, G.; Lee, J.; Sahu, P.K.; Jeong, L.S.; Cha, D.R.; Ha, H. Orally active, spe-cies-independent novel A3 adenosine receptor antagonist protects against kidney injury in db/db mice. Exp. Mol. Med. 2018, 50, 38–52. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.T.; Emala, C.W. Protective effects of renal ischemic preconditioning and adenosine pretreatment: Role of A1 and A3receptors. Am. J. Physiol. Physiol. 2000, 278, F380–F387. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.T.; Ota-Setlik, A.; Xu, H.; D’Agati, V.D.; Jacobson, M.A.; Emala, C.W. A3 adenosine receptor knockout mice are protected against ischemia- and myoglobinuria-induced renal failure. Am. J. Physiol. Physiol. 2003, 284, F267–F273. [Google Scholar] [CrossRef] [Green Version]
- Kretschmar, C.; Oyarzún, C.; Villablanca, C.; Jaramillo, C.; Alarcón, S.; Perez, G.; Díaz-Encarnación, M.M.; Pastor-Anglada, M.; Garrido, W.; Quezada, C.; et al. Reduced Adenosine Uptake and Its Contribution to Signaling that Mediates Profibrotic Activation in Renal Tubular Epithelial Cells: Implication in Diabetic Nephropathy. PLoS ONE 2016, 11, e0147430. [Google Scholar] [CrossRef] [Green Version]
- Min, H.S.; Cha, J.J.; Kim, K.; Kim, J.E.; Ghee, J.Y.; Kim, H.; Lee, J.E.; Han, J.-Y.; Jeong, L.S.; Cha, D.R.; et al. Renoprotective Effects of a Highly Selective A3 Adenosine Receptor Antagonist in a Mouse Model of Adriamycin-induced Nephropathy. J. Korean Med. Sci. 2016, 31, 1403–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haskó, G.; Pacher, P. A2A receptors in inflammation and injury: Lessons learned from transgenic animals. J. Leukoc. Biol. 2008, 83, 447–455. [Google Scholar] [CrossRef] [Green Version]
- Haskó, G.; Pacher, P.; Deitch, E.A.; Vizi, E.S. Shaping of monocyte and macrophage function by adenosine receptors. Pharmacol. Ther. 2007, 113, 264–275. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, G.W.; Linden, J.; Buster, B.L.; Scheld, W.M. Neutrophil A2A Adenosine Receptor Inhibits Inflammation in a Rat Model of Meningitis: Synergy with the Type IV Phosphodiesterase Inhibitor, Rolipram. J. Infect. Dis. 1999, 180, 1550–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshiba, M.; Rosin, D.L.; Hayashi, N.; Linden, J.; Sitkovsky, M.V. Patterns of A2A extracellular adenosine receptor expression in different functional subsets of human peripheral T cells. Flow cytometry studies with anti-A2A receptor monoclonal antibodies. Mol. Pharmacol. 1999, 55, 614–624. [Google Scholar]
- Raskovalova, T.; Huang, X.; Sitkovsky, M.; Zacharia, L.C.; Jackson, E.K.; Gorelik, E. GsProtein-Coupled Adenosine Receptor Signaling and Lytic Function of Activated NK Cells. J. Immunol. 2005, 175, 4383–4391. [Google Scholar] [CrossRef] [Green Version]
- Okusa, M.D.; Linden, J.; Macdonald, T.; Huang, L. Selective A2A adenosine receptor activation reduces ischemia-reperfusion injury in rat kidney. Am. J. Physiol. Renal. Physiol. 1999, 277, F404–F412. [Google Scholar] [CrossRef] [PubMed]
- Okusa, M.D.; Linden, J.; Huang, L.; Rieger, J.M.; Macdonald, T.L.; Huynh, L.P. A2A adenosine receptor-mediated inhibition of renal injury and neutrophil adhesion. Am. J. Physiol. Renal. Physiol. 2000, 279, F809–F818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okusa, M.D.; Linden, J.; Huang, L.; Rosin, D.L.; Smith, D.F.; Sullivan, G. Enhanced protection from renal ischemia-reperfusion [correction of ischemia:reperfusion] injury with A2A-adenosine receptor activation and PDE 4 inhibition. Kidney Int. 2001, 59, 2114–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awad, A.S.; Huang, L.; Ye, H.; Duong, E.T.A.; Bolton, W.K.; Linden, J.; Okusa, M.D. Adenosine A2A receptor activation attenuates inflammation and injury in diabetic nephropathy. Am. J. Physiol. Physiol. 2006, 290, F828–F837. [Google Scholar] [CrossRef] [Green Version]
- Persson, P.; Friederich-Persson, M.; Fasching, A.; Hansell, P.; Inagi, R.; Palm, F. Adenosine A2a receptor stimulation prevents proteinuria in diabetic rats by promoting an anti-inflammatory phenotype without affecting oxidative stress. Acta Physiol. 2015, 214, 311–318. [Google Scholar] [CrossRef]
- Awad, A.S.; Rouse, M.; Liu, L.; Vergis, A.L.; Rosin, D.L.; Linden, J.; Sedor, J.R.; Okusa, M.D. Activation of Adenosine 2A Receptors Preserves Structure and Function of Podocytes. J. Am. Soc. Nephrol. 2007, 19, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Garcia, G.E.; Truong, L.D.; Chen, J.-F.; Johnson, R.J.; Feng, L. Adenosine A2A receptor activation prevents progressive kidney fibrosis in a model of immune-associated chronic inflammation. Kidney Int. 2011, 80, 378–388. [Google Scholar] [CrossRef] [Green Version]
- Xiao, H.; Shen, H.Y.; Liu, W.; Xiong, R.P.; Li, P.; Meng, G.; Yang, N.; Chen, X.; Si, L.Y.; Zhou, Y.G. Adenosine A2A receptor: A target for regulating renal interstitial fibrosis in obstructive nephropathy. PLoS ONE 2013, 8, e60173. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Majik, M.S.; Kim, K.; Pyee, Y.; Lee, Y.; Alexander, V.; Chung, H.-J.; Lee, H.W.; Chandra, G.; Lee, J.H.; et al. Structure–Activity Relationships of Truncated C2- or C8-Substituted Adenosine Derivatives as Dual Acting A2A and A3 Adenosine Receptor Ligands. J. Med. Chem. 2011, 55, 342–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Oh, D.-S.; Dorotea, D.; Son, E.; Kim, D.-S.; Ha, H. Dojuksan ameliorates tubulointerstitial fibrosis through irisin-mediated muscle-kidney crosstalk. Phytomedicine 2021, 80, 153393. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.-M.; Nikolic-Paterson, D.J.; Lan, H.Y. Inflammatory processes in renal fibrosis. Nat. Rev. Nephrol. 2014, 10, 493–503. [Google Scholar] [CrossRef]
- Noh, H.; Ha, H. Reactive oxygen species and oxidative stress. Contrib. Nephrol. 2011, 170, 102–112. [Google Scholar] [CrossRef]
- Forbes, M.S.; Thornhill, B.A.; Minor, J.J.; Gordon, K.A.; Galarreta, C.I.; Chevalier, R.L. Fight-or-flight: Murine unilateral ureteral obstruction causes extensive proximal tubular degeneration, collecting duct dilatation, and minimal fibrosis. Am. J. Physiol. Physiol. 2012, 303, F120–F129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevalier, R.L.; Forbes, M.S.; Thornhill, B.A. Ureteral obstruction as a model of renal interstitial fibrosis and obstructive nephropathy. Kidney Int. 2009, 75, 1145–1152. [Google Scholar] [CrossRef] [Green Version]
- Borea, P.A.; Gessi, S.; Merighi, S.; Vincenzi, F.; Varani, K. Pathological overproduction: The bad side of adenosine. Br. J. Pharmacol. 2017, 174, 1945–1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, B.; Xu, S.; Hou, X.; Pimentel, D.R.; Brecher, P.; Cohen, R.A. Temporal control of NF-kappaB activation by ERK differentially regulates interleukin-1beta-induced gene expression. J. Biol. Chem. 2004, 279, 1323–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tashiro, K.; Tamada, S.; Kuwabara, N.; Komiya, T.; Takekida, K.; Asai, T.; Iwao, H.; Sugimura, K.; Matsumura, Y.; Takaoka, M.; et al. Attenuation of renal fibrosis by proteasome inhibition in rat obstructive nephropathy: Possible role of nuclear factor kappaB. Int. J. Mol. Med. 2003, 12, 587–592. [Google Scholar] [CrossRef]
- Giambelluca, M.S.; Pouliot, M. Early tyrosine phosphorylation events following adenosine A2A receptor in human neutrophils: Identification of regulated pathways. J. Leukoc. Biol. 2017, 102, 829–836. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Gao, Z.; Xu, F.; Liu, L.; Luo, Q.; Shen, Y.; Wu, X.; Wu, X.; Sun, Y.; Wu, X.; et al. A novel combination of astilbin and low-dose methotrexate respectively targeting A2AAR and its ligand adenosine for the treatment of collagen-induced arthritis. Biochem. Pharmacol. 2018, 153, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Che, J.; Chan, E.S.L.; Cronstein, B.N. Adenosine A2A Receptor Occupancy Stimulates Collagen Expression by Hepatic Stellate Cells via Pathways Involving Protein Kinase A, Src, and Extracellular Signal-Regulated Kinases 1/2 Signaling Cascade or p38 Mitogen-Activated Protein Kinase Signaling Pathway. Mol. Pharmacol. 2007, 72, 1626–1636. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, R.A.; Agha, A.M.; Abdel-Rahman, A.A.; Nassar, N.N. Role of adenosine A2A receptor in cerebral ischemia reperfusion injury: Signaling to phosphorylated extracellular signal-regulated protein kinase (pERK1/2). Neuroscience 2016, 314, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Jhun, B.S.; Oh, Y.T.; Lee, J.H.; Choe, W.; Baik, H.H.; Ha, J.; Yoon, K.S.; Kim, S.S.; Kang, I. Activation of adenosine A3 receptor suppresses lipopolysaccharide-induced TNF-alpha production through inhibition of PI 3-kinase/Akt and NF-kappaB activation in murine BV2 microglial cells. Neurosci. Lett. 2006, 396, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Varani, K.; Maniero, S.; Vincenzi, F.; Targa, M.; Stefanelli, A.; Maniscalco, P.; Martini, F.; Tognon, M.; Borea, P.A. A3 receptors are overexpressed in pleura from patients with mesothelioma and reduce cell growth via Akt/nuclear factor-κB pathway. Am. J. Respir. Crit. Care Med. 2011, 183, 522–530. [Google Scholar] [CrossRef]
- Lee, H.T.; Kim, M.; Joo, J.D.; Gallos, G.; Chen, J.-F.; Emala, C.W. A3 adenosine receptor activation decreases mortality and renal and hepatic injury in murine septic peritonitis. Am. J. Physiol. Integr. Comp. Physiol. 2006, 291, R959–R969. [Google Scholar] [CrossRef] [Green Version]
- Karmouty-Quintana, H.; Xia, Y.; Blackburn, M.R. Adenosine signaling during acute and chronic disease states. J. Mol. Med. 2013, 91, 173–181. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′ → 3′) | Reverse (5′ → 3′) |
|---|---|---|
| 18S | CGAAAGCATTTGCCAAGAAT | AGTCGGCATCGTTTATGGTC |
| NGAL | GGCCAGTTCACTCTGGGAAA | TGGCGAACTGGTTGTAGTCC3 |
| ICAM-1 | CTTCCAGCTACCATGCCAAA | CTTCAGAGGCAGGAAACAGG |
| iNOS | GGCAGCCTGTGAGACCTTTG | CATTGGAAGTGAAGCGTTTCG |
| IL-6 | AGTTGCCTTCTTGGGACTGA | TCCACGATTTCCCAGAGAAC |
| NOX1 | AGCCATTGGATCACAACCTC | AGAAGCGAGAGATCCATCCA |
| NOX2 | TGCACCATGATGAGGAGAAA | CCACACAGGAAAACGCCTAT |
| NOX3 | ATTTCACTACCCCGTGAGCG | TCAGGCAGGCTCTGTGATTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pak, E.S.; Jeong, L.S.; Hou, X.; Tripathi, S.K.; Lee, J.; Ha, H. Dual Actions of A2A and A3 Adenosine Receptor Ligand Prevents Obstruction-Induced Kidney Fibrosis in Mice. Int. J. Mol. Sci. 2021, 22, 5667. https://doi.org/10.3390/ijms22115667
Pak ES, Jeong LS, Hou X, Tripathi SK, Lee J, Ha H. Dual Actions of A2A and A3 Adenosine Receptor Ligand Prevents Obstruction-Induced Kidney Fibrosis in Mice. International Journal of Molecular Sciences. 2021; 22(11):5667. https://doi.org/10.3390/ijms22115667
Chicago/Turabian StylePak, Eun Seon, Lak Shin Jeong, Xiyan Hou, Sushil K. Tripathi, Jiyoun Lee, and Hunjoo Ha. 2021. "Dual Actions of A2A and A3 Adenosine Receptor Ligand Prevents Obstruction-Induced Kidney Fibrosis in Mice" International Journal of Molecular Sciences 22, no. 11: 5667. https://doi.org/10.3390/ijms22115667
APA StylePak, E. S., Jeong, L. S., Hou, X., Tripathi, S. K., Lee, J., & Ha, H. (2021). Dual Actions of A2A and A3 Adenosine Receptor Ligand Prevents Obstruction-Induced Kidney Fibrosis in Mice. International Journal of Molecular Sciences, 22(11), 5667. https://doi.org/10.3390/ijms22115667