
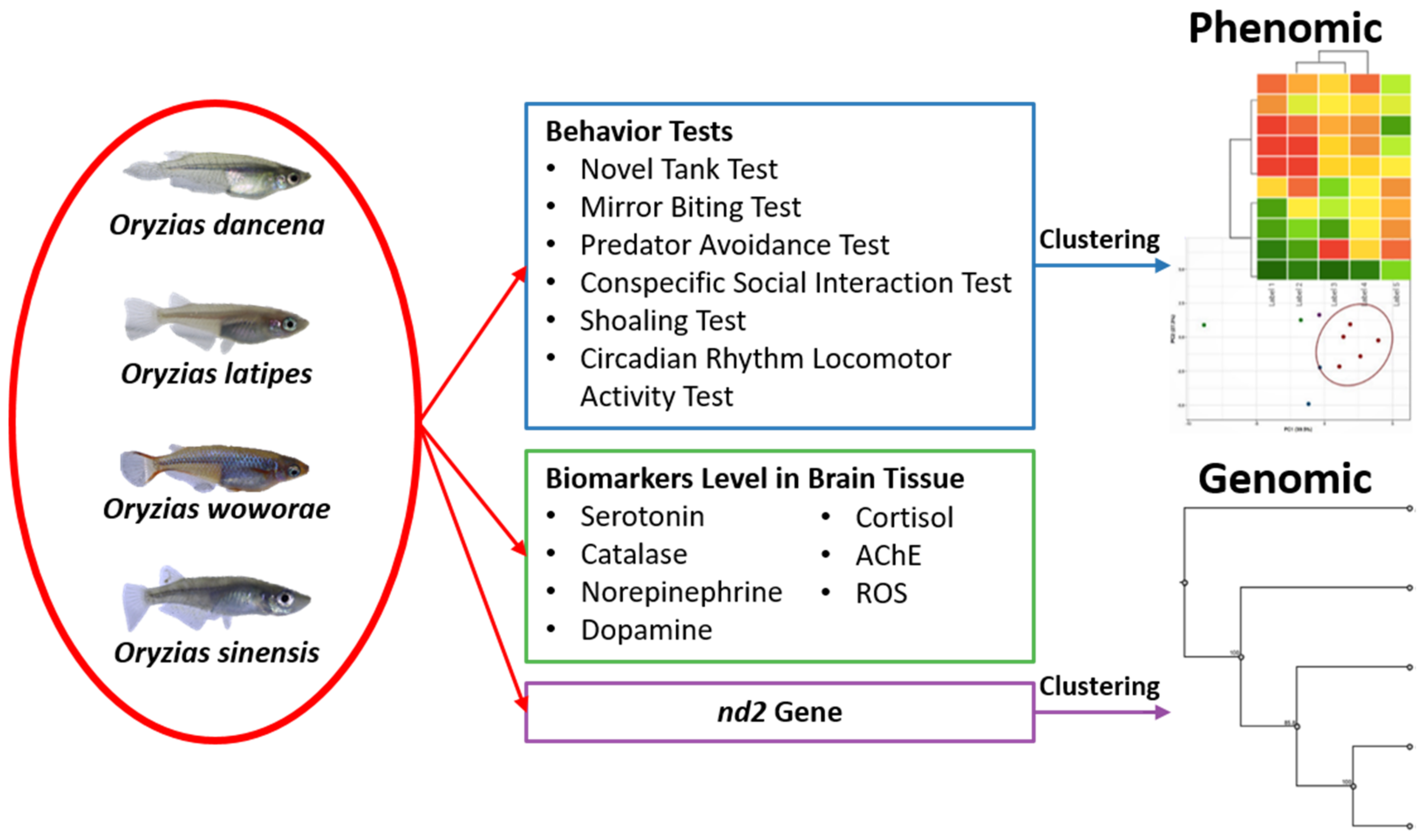
Interspecies Behavioral Variability of Medaka Fish Assessed by Comparative Phenomics


 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
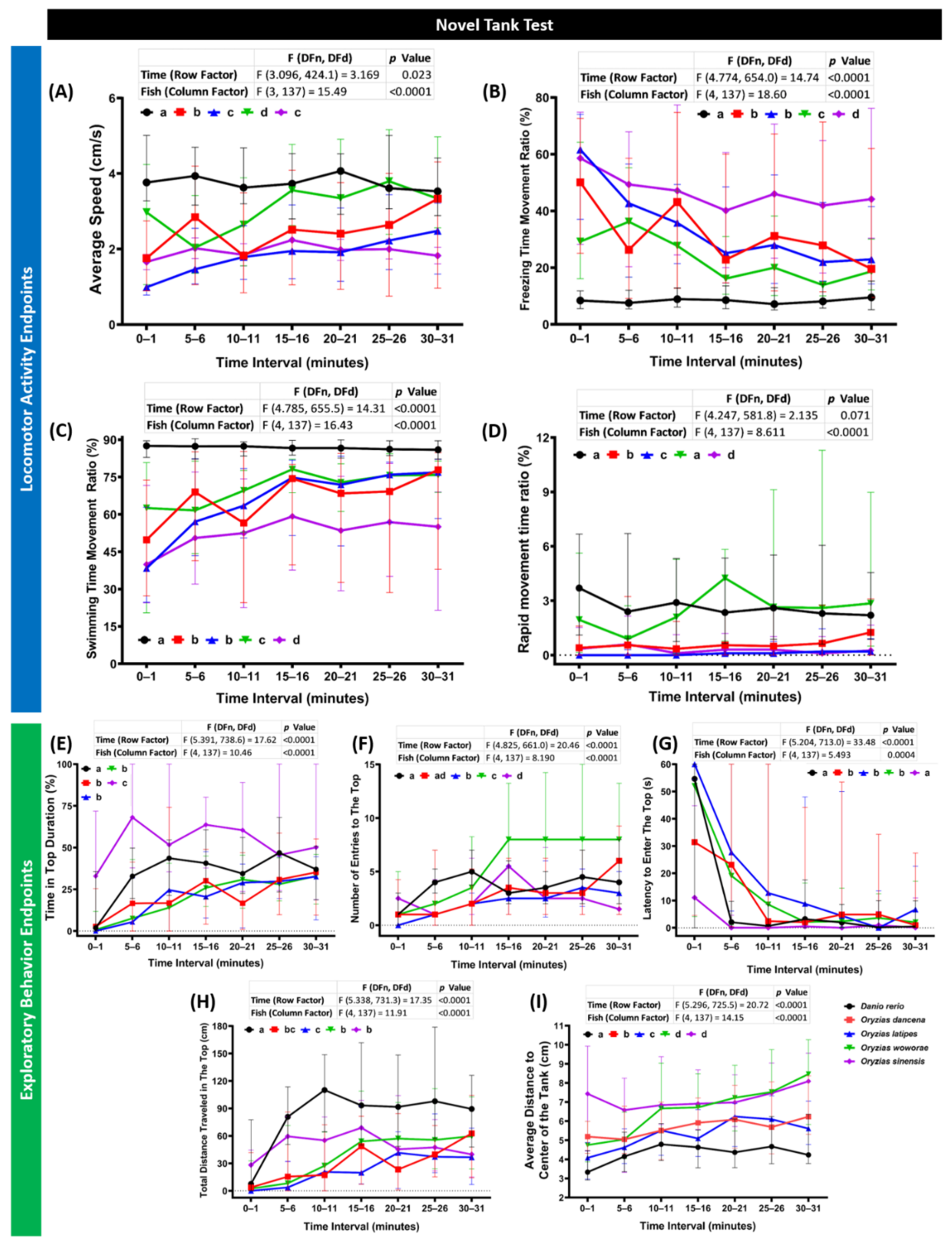
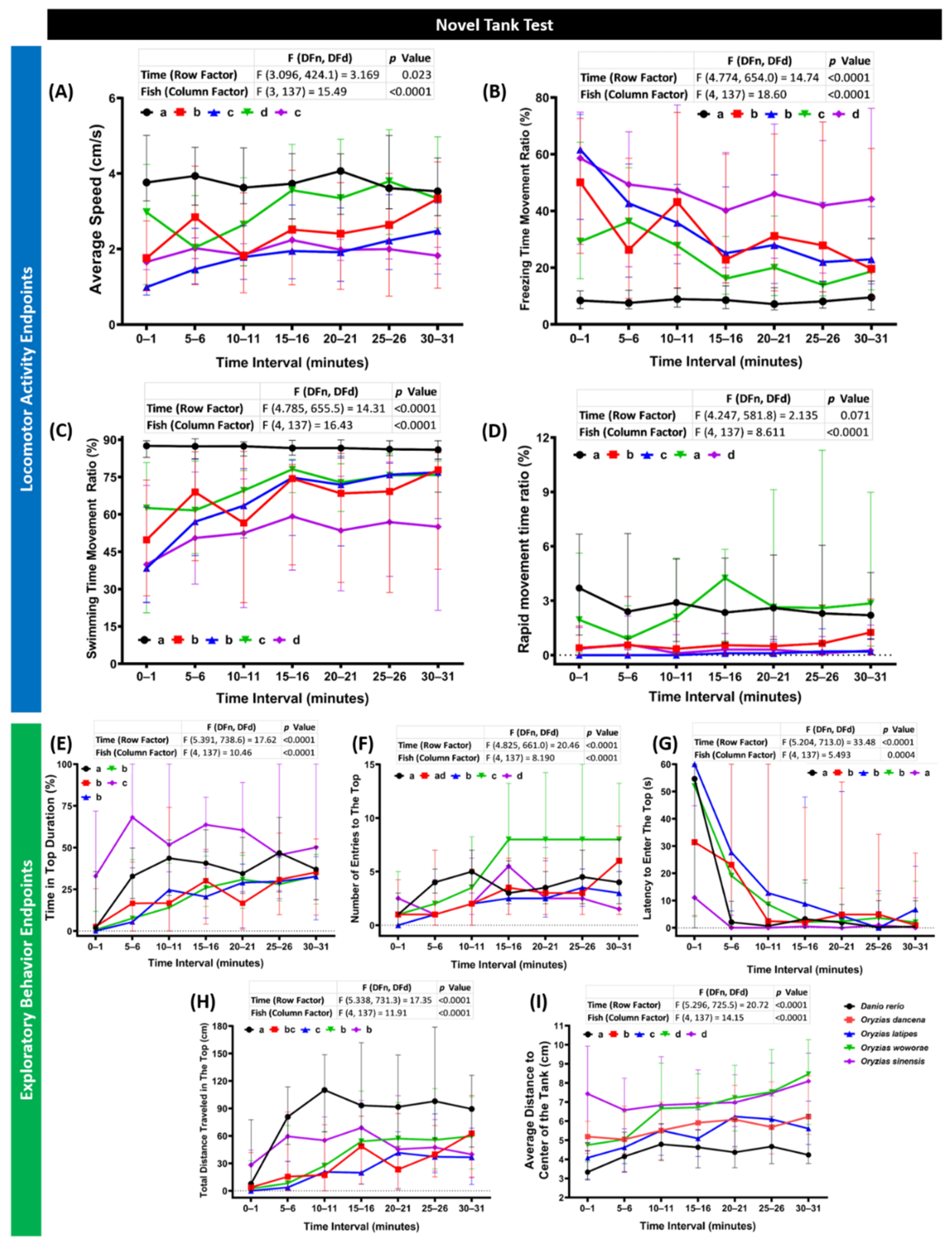
2.1. Novel Tank Assay Performance Comparison of Four Species of Medaka Fish
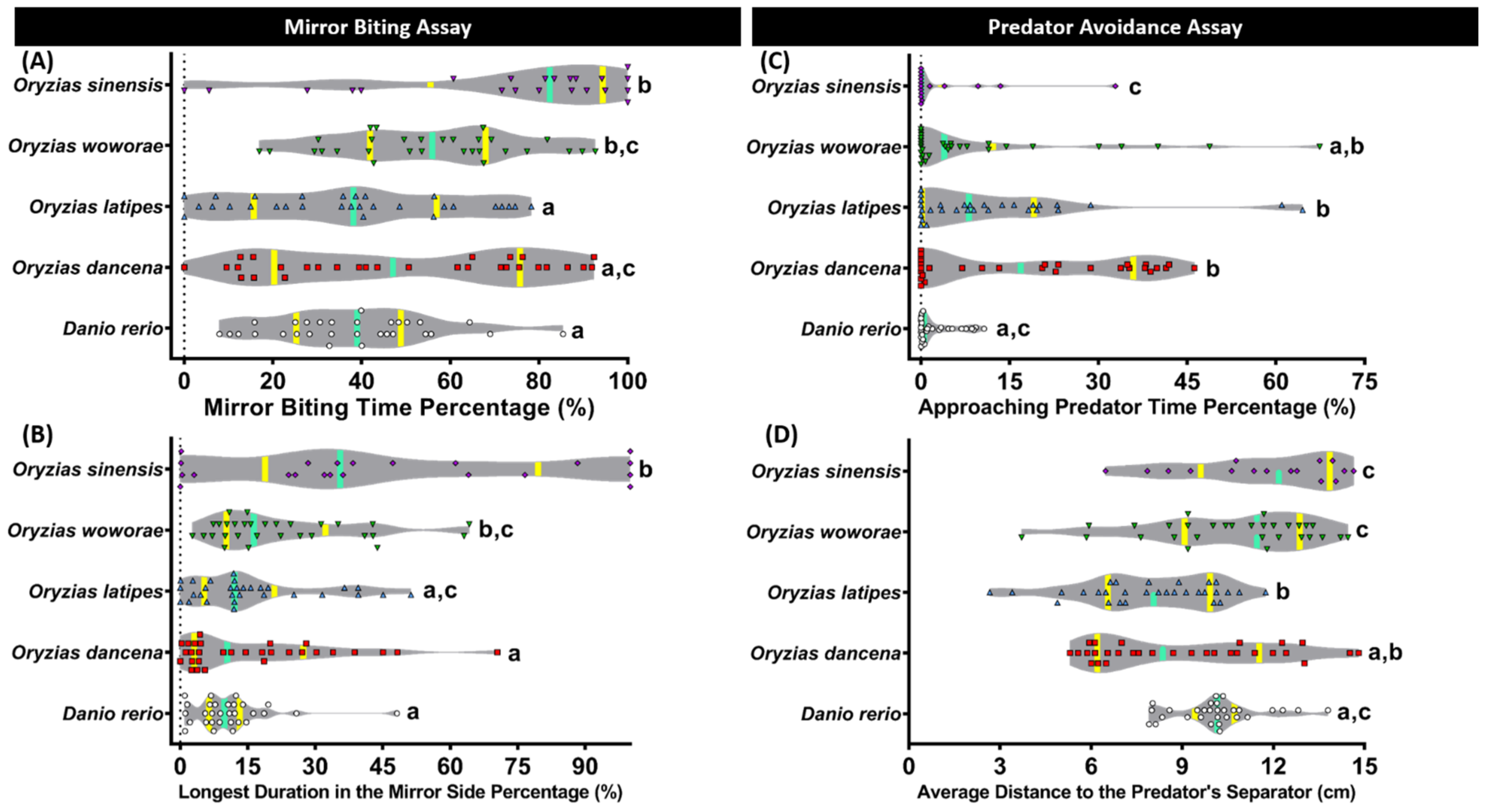
2.2. Aggressiveness Comparison of Four Species of Medaka Fish
2.3. Comparison of Predator Avoidance Test Performance for Four Species of Medaka Fish
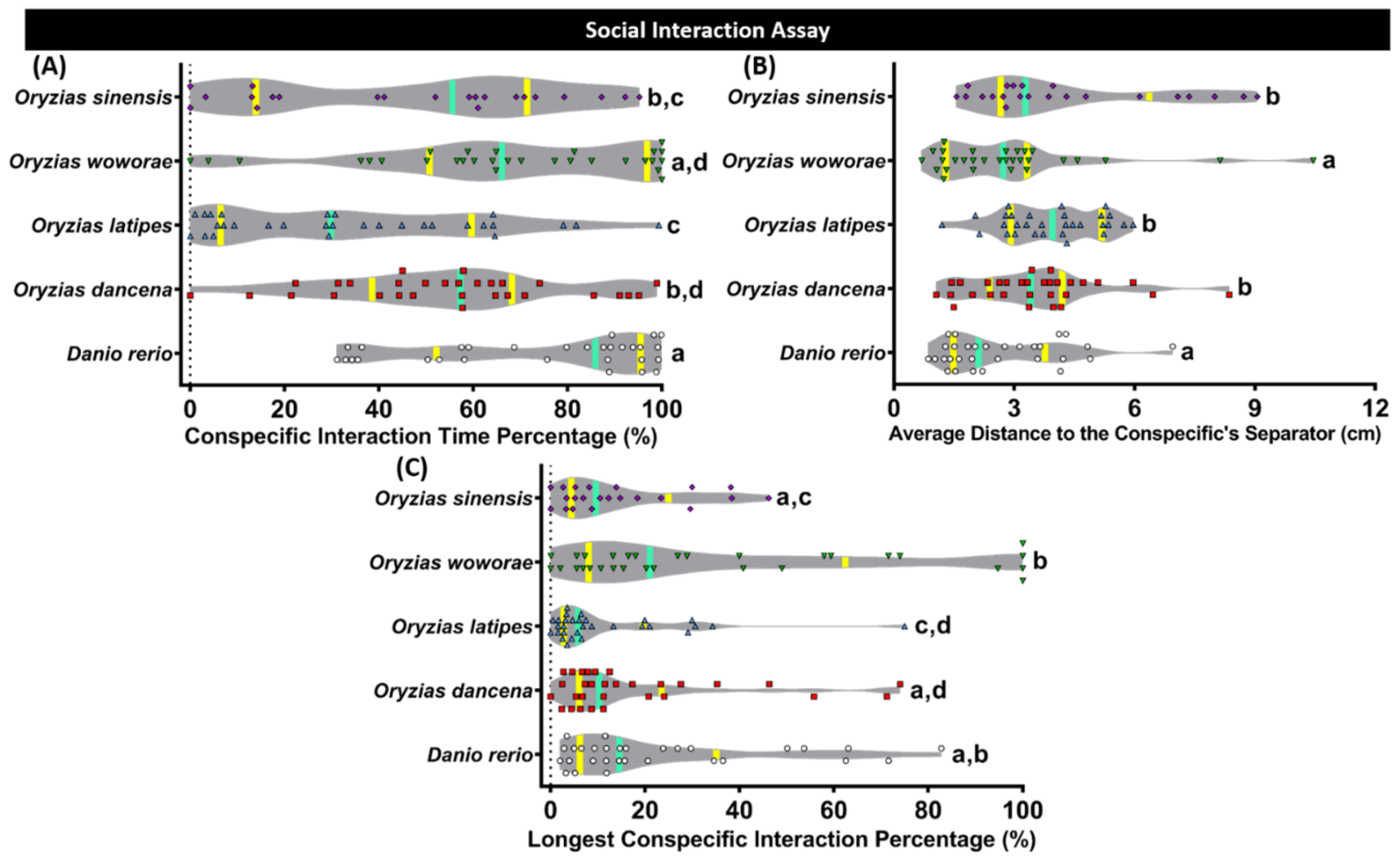
2.4. Conspecific Social Interaction Comparison of Four Species of Medaka Fish
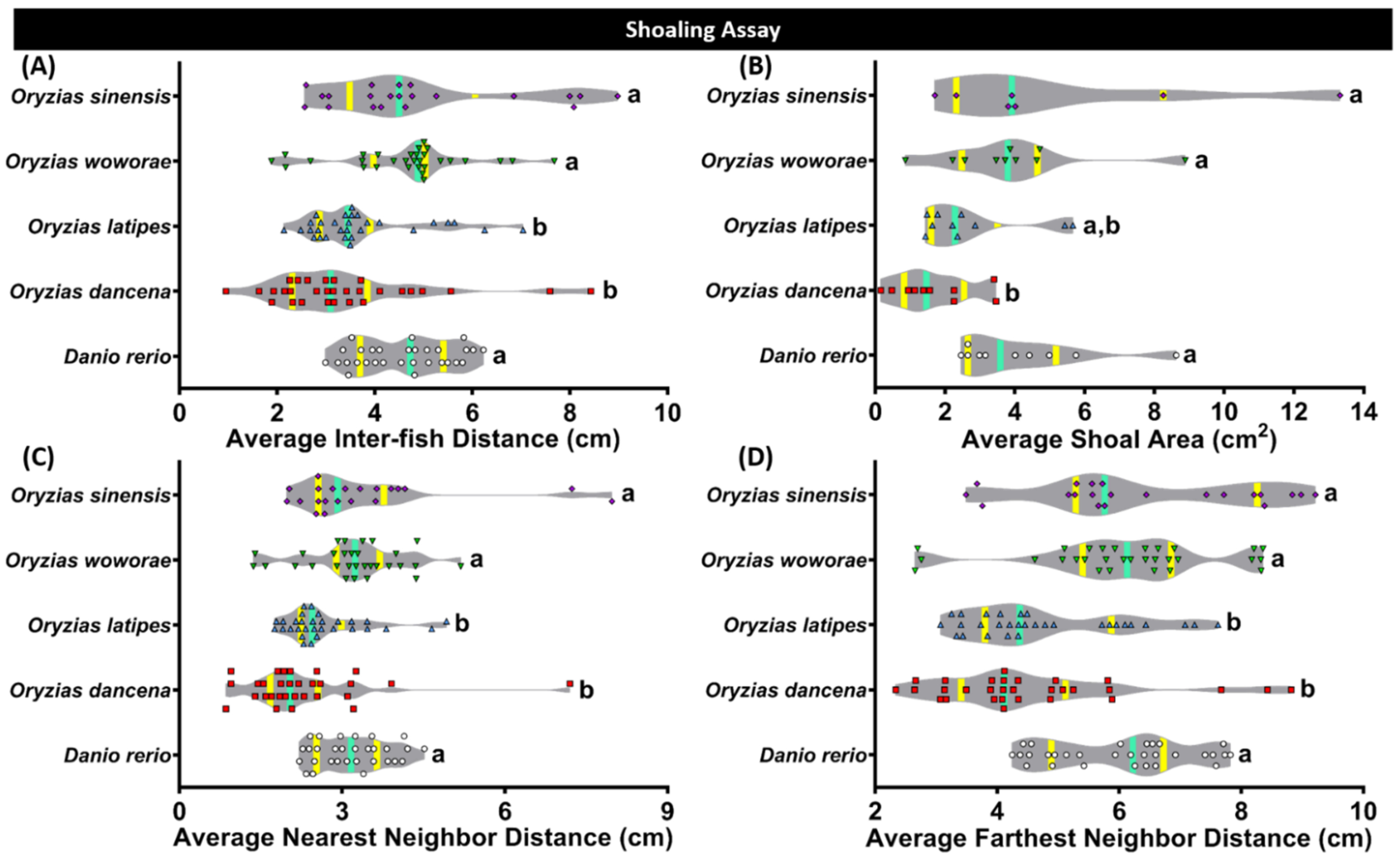
2.5. Comparison of Shoaling Behavior for Four Species of Medaka Fish
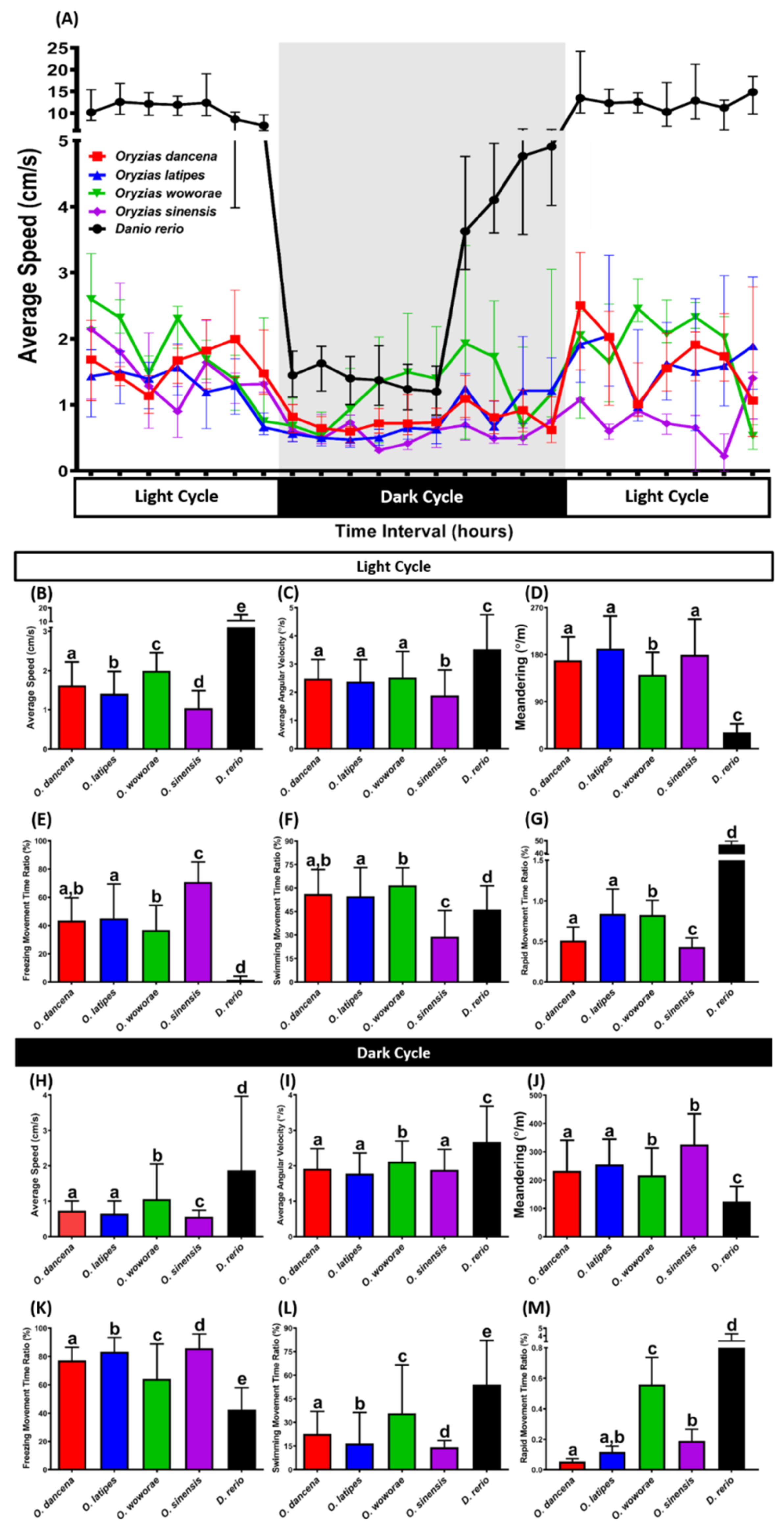
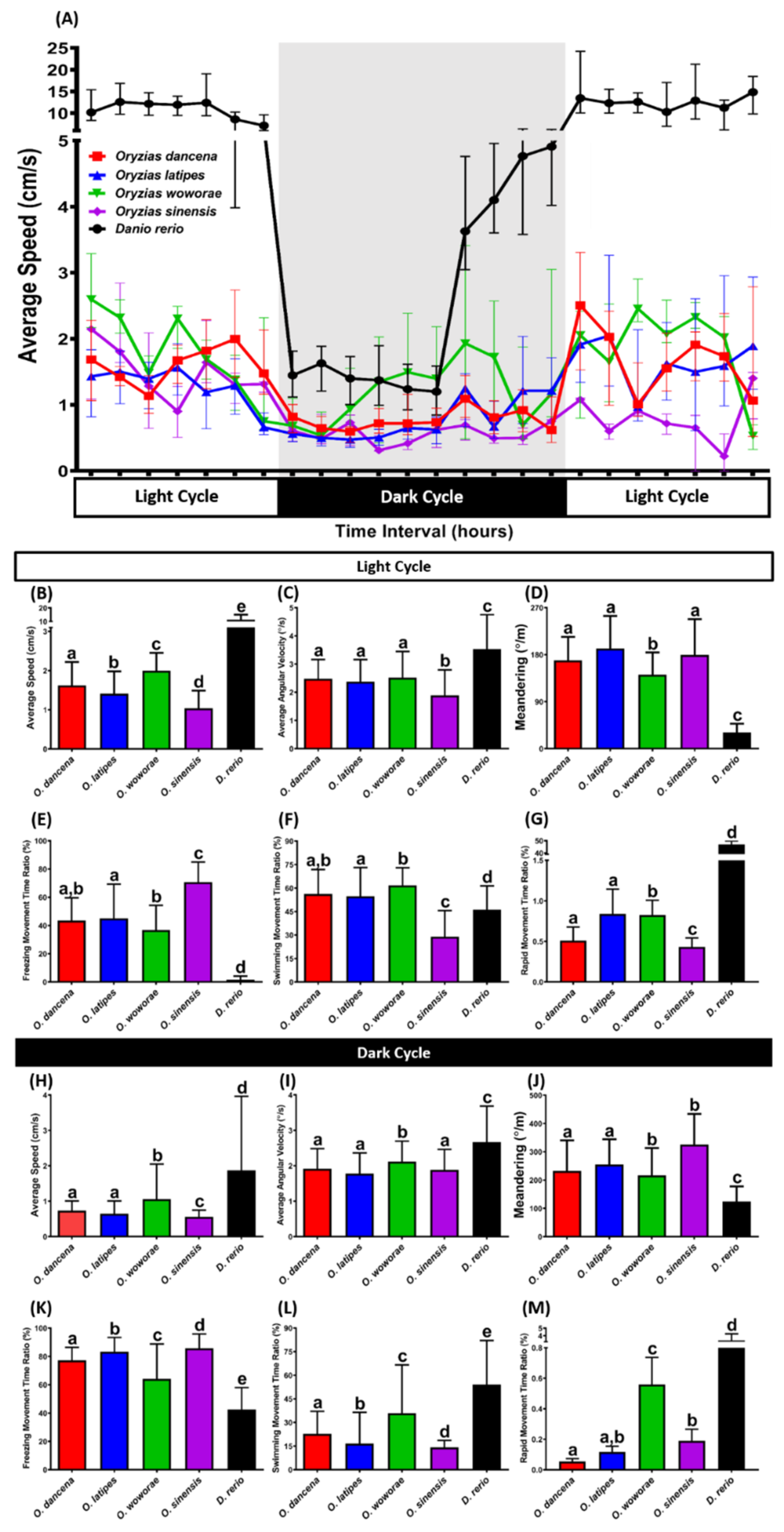
2.6. Comparison of Circadian Locomotor Activity Rhythms for Four Medaka Species
2.7. Biochemical Assay of Biomarker Expression in the Brain
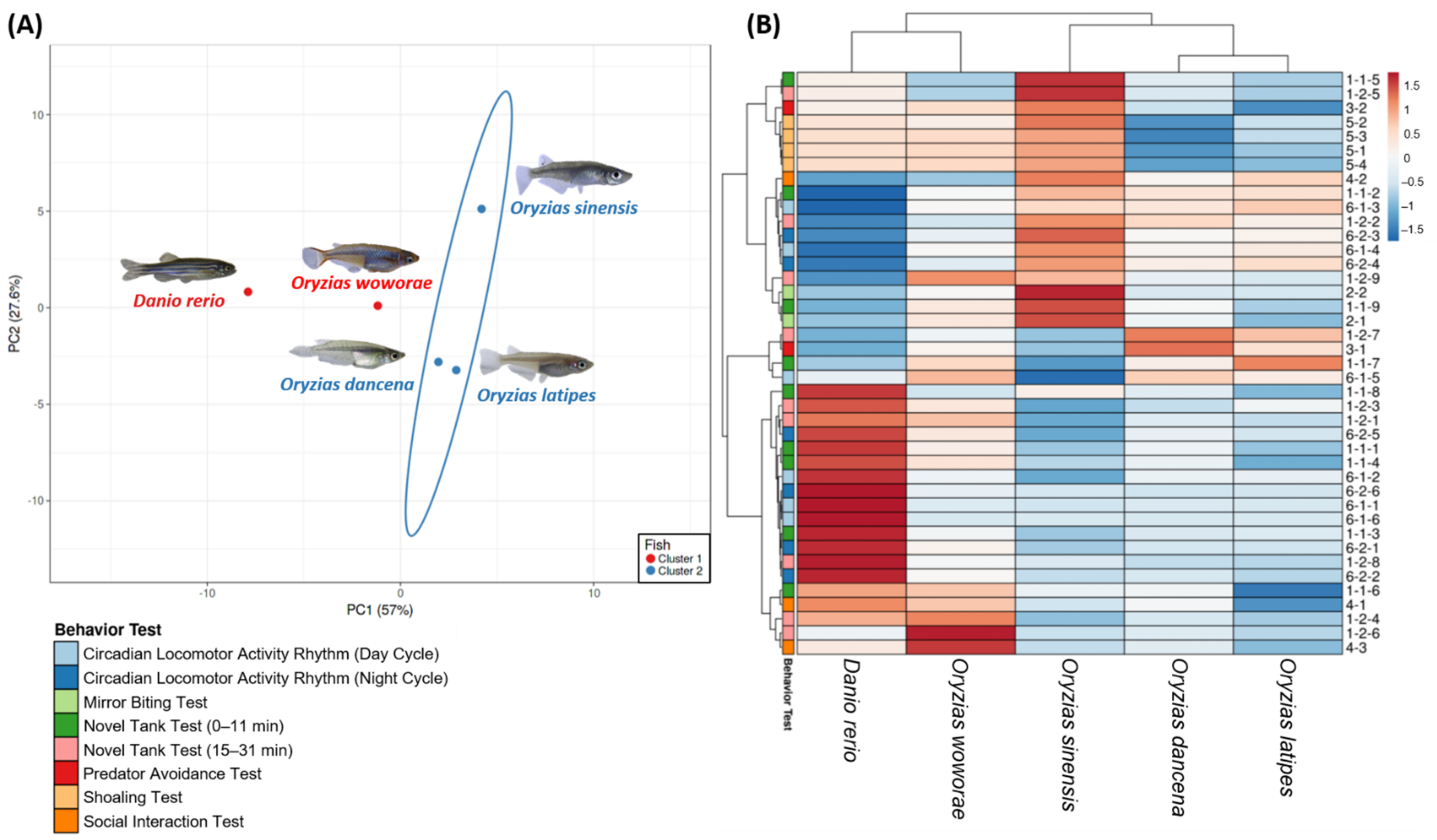
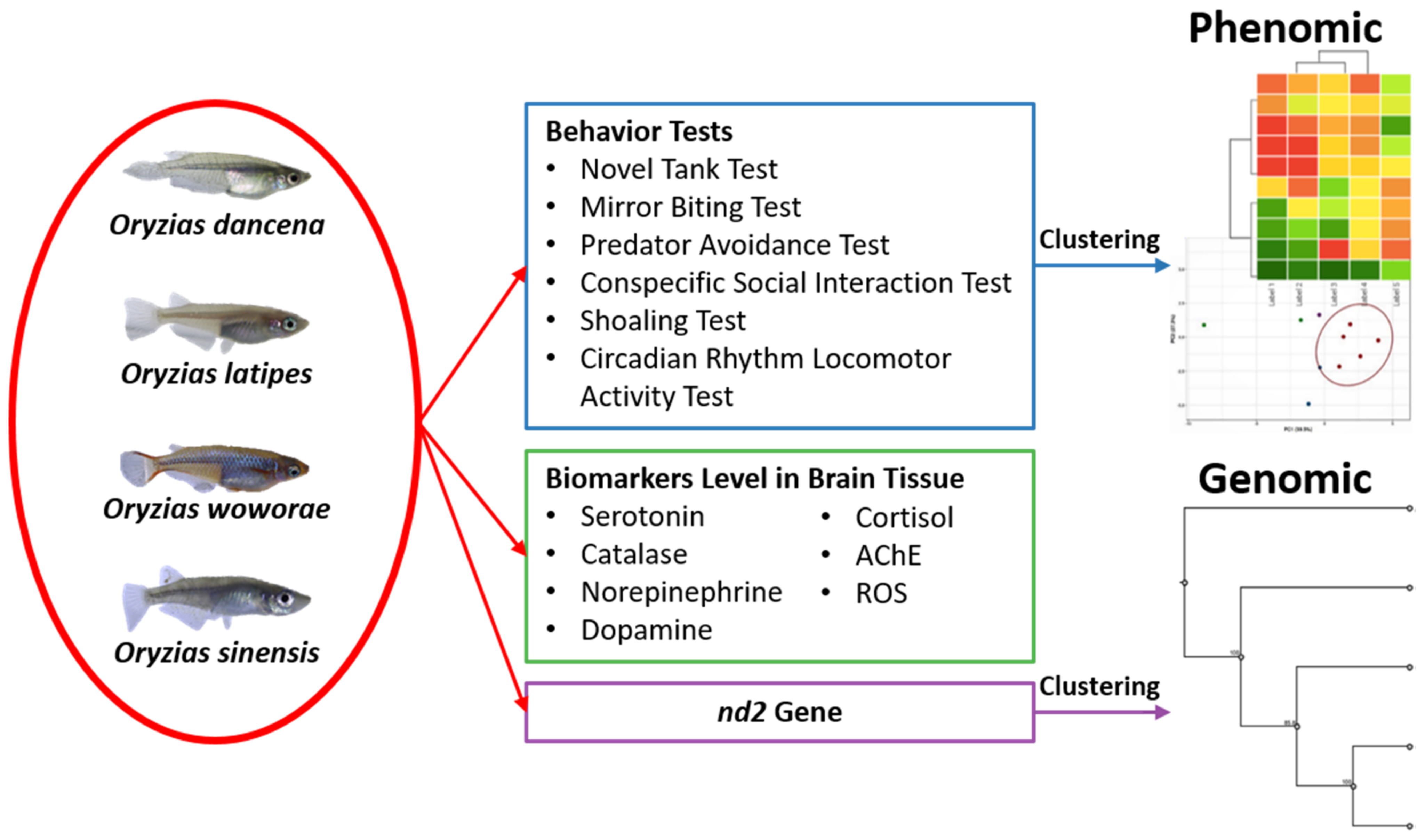
2.8. PCA Analysis and Hierarchical Clustering Analysis of Several Medaka Fish Behavioral Endpoints
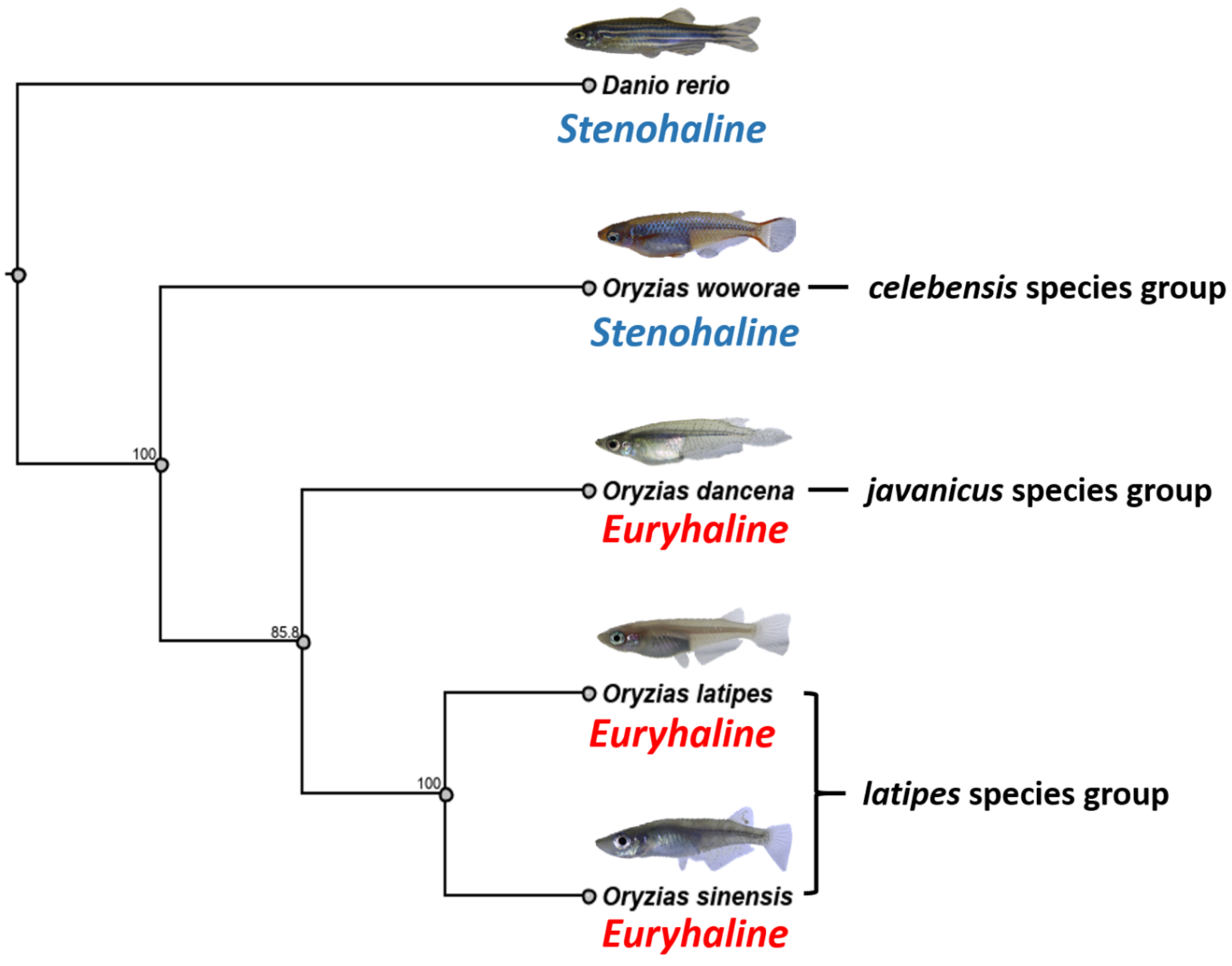
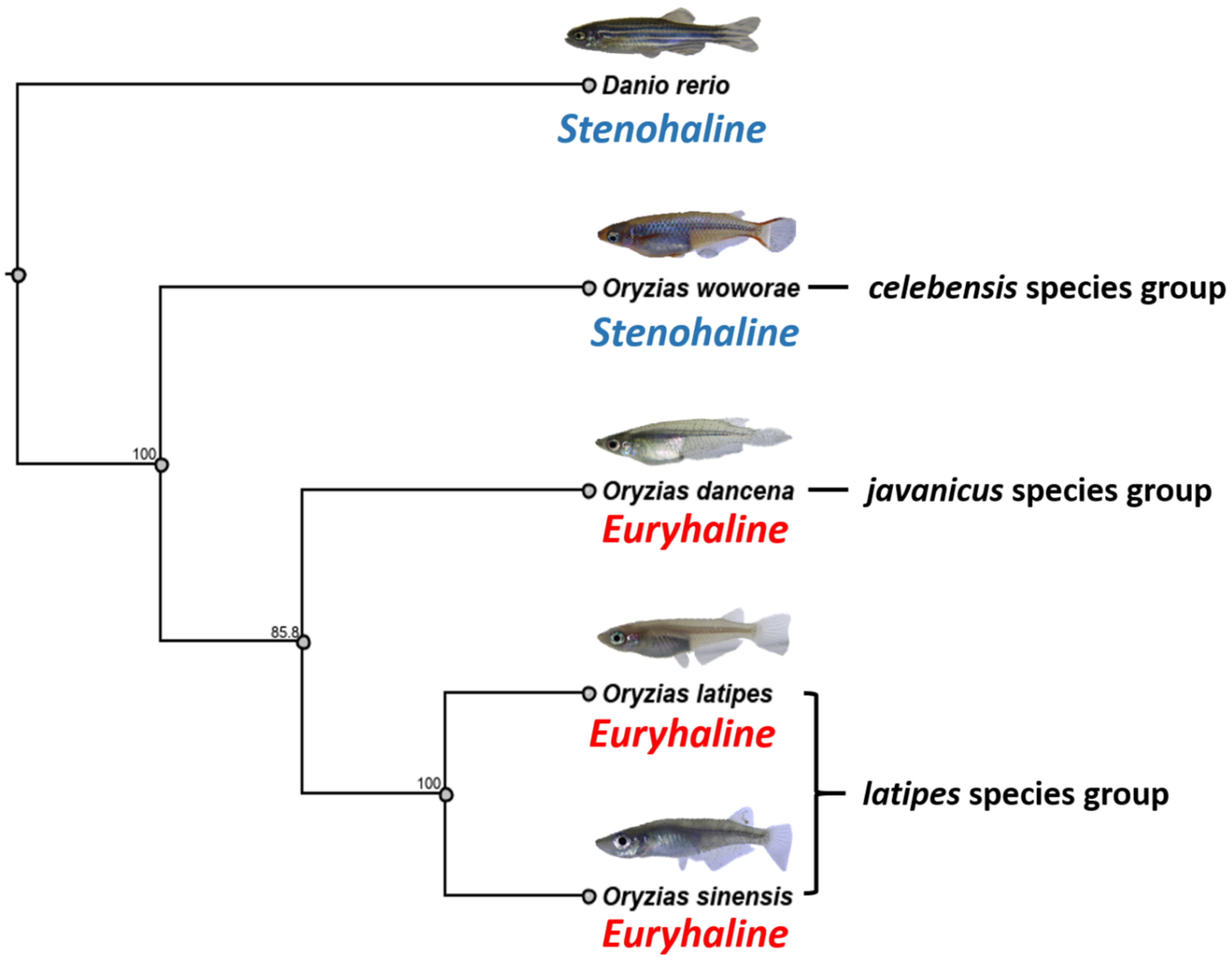
2.9. Phylogenetic Relationships of Four Medaka Species
3. Discussion
4. Materials and Methods
4.1. Animal Husbandry
4.2. Animal Ethics and Behavioral Tests
4.3. Brain Tissue Preparation, Total Protein Determination, and Quantification of Neurotransmitters, Stress Hormones, and Oxidative Stress Markers
4.4. Statistical Analyses
4.5. PCA, Heatmap, and Clustering Analysis
4.6. Phylogenetic Tree Construction
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioral Endpoints | O. dancena | O. latipes | O. woworae | O. sinensis | D. rerio | |
|---|---|---|---|---|---|---|
| Average Speed | O. dancena | 0.0097 | 0.0005 | <0.0001 | <0.0001 | |
| O. latipes | 0.0097 | <0.0001 | 0.1134 | <0.0001 | ||
| O. woworae | 0.0005 | <0.0001 | <0.0001 | <0.0001 | ||
| O. sinensis | <0.0001 | 0.1134 | <0.0001 | <0.0001 | ||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 | ||
| Freezing Time Movement Ratio | O. dancena | 0.2654 | <0.0001 | 0.0155 | <0.0001 | |
| O. latipes | 0.2654 | 0.0002 | 0.0002 | <0.0001 | ||
| O. woworae | <0.0001 | 0.0002 | <0.0001 | <0.0001 | ||
| O. sinensis | 0.0155 | 0.0002 | <0.0001 | <0.0001 | ||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 | ||
| Swimming Time Movement Ratio | O. dancena | 0.1090 | 0.0002 | 0.0286 | <0.0001 | |
| O. latipes | 0.1090 | 0.0171 | <0.0001 | <0.0001 | ||
| O. woworae | 0.0002 | 0.0171 | <0.0001 | <0.0001 | ||
| O. sinensis | 0.0286 | <0.0001 | <0.0001 | <0.0001 | ||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 | ||
| Rapid Time Movement Ratio | O. dancena | <0.0001 | <0.0001 | 0.0007 | <0.0001 | |
| O. latipes | <0.0001 | <0.0001 | 0.3643 | <0.0001 | ||
| O. woworae | <0.0001 | <0.0001 | <0.0001 | 0.4803 | ||
| O. sinensis | 0.0007 | 0.3643 | <0.0001 | <0.0001 | ||
| D. rerio | <0.0001 | <0.0001 | 0.4803 | <0.0001 | ||
| Time in Top | O. dancena | 0.0636 | 0.0595 | <0.0001 | 0.0164 | |
| O. latipes | 0.0636 | 0.9472 | <0.0001 | <0.0001 | ||
| O. woworae | 0.0595 | 0.9472 | <0.0001 | <0.0001 | ||
| O. sinensis | <0.0001 | <0.0001 | <0.0001 | <0.0001 | ||
| D. rerio | 0.0164 | <0.0001 | <0.0001 | <0.0001 | ||
| Number of Entries to The Top | O. dancena | 0.0050 | <0.0001 | 0.4143 | 0.0619 | |
| O. latipes | 0.0050 | <0.0001 | 0.0461 | <0.0001 | ||
| O. woworae | <0.0001 | <0.0001 | <0.0001 | <0.0001 | ||
| O. sinensis | 0.4143 | 0.0461 | <0.0001 | 0.0057 | ||
| D. rerio | 0.0619 | <0.0001 | <0.0001 | 0.0057 | ||
| Latency to Enter the Top | O. dancena | 0.2674 | 0.4705 | <0.0001 | <0.0001 | |
| O. latipes | 0.2674 | 0.0648 | <0.0001 | <0.0001 | ||
| O. woworae | 0.4705 | 0.0648 | <0.0001 | 0.0002 | ||
| O. sinensis | <0.0001 | <0.0001 | <0.0001 | 0.5059 | ||
| D. rerio | <0.0001 | <0.0001 | 0.0002 | 0.5059 | ||
| Total Distance Traveled in The Top | O. dancena | 0.1233 | 0.2273 | 0.0753 | <0.0001 | |
| O. latipes | 0.1233 | 0.0061 | 0.0009 | <0.0001 | ||
| O. woworae | 0.2273 | 0.0061 | 0.5744 | <0.0001 | ||
| O. sinensis | 0.0753 | 0.0009 | 0.5744 | <0.0001 | ||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 | ||
| Thigmotaxis | O. dancena | 0.0162 | <0.0001 | <0.0001 | <0.0001 | |
| O. latipes | 0.0162 | <0.0001 | <0.0001 | 0.0005 | ||
| O. woworae | <0.0001 | <0.0001 | 0.2250 | <0.0001 | ||
| O. sinensis | <0.0001 | <0.0001 | 0.2250 | <0.0001 | ||
| D. rerio | <0.0001 | 0.0005 | <0.0001 | <0.0001 |
| Behavior Tests | Behavioral Endpoints | O. dancena | O. latipes | O. woworae | O. sinensis | D. rerio | |
|---|---|---|---|---|---|---|---|
| Mirror Biting Test | Mirror Biting Time Percentage | O. dancena | 0.0988 | 0.3011 | 0.0040 | 0.1159 | |
| O. latipes | 0.0988 | 0.0073 | <0.0001 | 0.9375 | |||
| O. woworae | 0.3011 | 0.0073 | 0.0542 | 0.0091 | |||
| O. sinensis | 0.0040 | <0.0001 | 0.0542 | <0.0001 | |||
| D. rerio | 0.1159 | 0.9375 | 0.0091 | <0.0001 | |||
| Longest Duration in the Mirror Side Percentage | O. dancena | 0.7656 | 0.0466 | 0.0023 | 0.4318 | ||
| O. latipes | 0.7656 | 0.0907 | 0.0054 | 0.2782 | |||
| O. woworae | 0.0466 | 0.0907 | 0.2210 | 0.0055 | |||
| O. sinensis | 0.0023 | 0.0054 | 0.2210 | 0.0002 | |||
| D. rerio | 0.4318 | 0.2782 | 0.0055 | 0.0002 | |||
| Predator Avoidance Test | Approaching Predator Time Percentage | O. dancena | 0.7111 | 0.1669 | 0.0016 | 0.0104 | |
| O. latipes | 0.7111 | 0.3115 | 0.0044 | 0.0285 | |||
| O. woworae | 0.1669 | 0.3115 | 0.0448 | 0.2384 | |||
| O. sinensis | 0.0016 | 0.0044 | 0.0448 | 0.3064 | |||
| D. rerio | 0.0104 | 0.0285 | 0.2384 | 0.3064 | |||
| Average Distance to the Predator’s Separator | O. dancena | 0.1360 | 0.0153 | 0.0017 | 0.1091 | ||
| O. latipes | 0.1360 | <0.0001 | <0.0001 | 0.0020 | |||
| O. woworae | 0.0153 | <0.0001 | 0.2628 | 0.4109 | |||
| O. sinensis | 0.0017 | <0.0001 | 0.2628 | 0.0710 | |||
| D. rerio | 0.1091 | 0.0020 | 0.4109 | 0.0710 | |||
| Social Interaction Test | Conspecific Interaction Time Percentage | O. dancena | 0.0204 | 0.0675 | 0.5032 | 0.0155 | |
| O. latipes | 0.0204 | <0.0001 | 0.1432 | <0.0001 | |||
| O. woworae | 0.0675 | <0.0001 | 0.0187 | 0.5531 | |||
| O. sinensis | 0.5032 | 0.1432 | 0.0187 | 0.0038 | |||
| D. rerio | 0.0155 | <0.0001 | 0.5531 | 0.0038 | |||
| Average Distance to the Conspecific’s Separator | O. dancena | 0.2037 | 0.0349 | 0.4902 | 0.0294 | ||
| O. latipes | 0.2037 | 0.0007 | 0.6318 | 0.0006 | |||
| O. woworae | 0.0349 | 0.0007 | 0.0085 | 0.9450 | |||
| O. sinensis | 0.4902 | 0.6318 | 0.0085 | 0.0071 | |||
| D. rerio | 0.0294 | 0.0006 | 0.9450 | 0.0071 | |||
| Longest Conspecific Interaction Percentage | O. dancena | 0.0969 | 0.0360 | 0.7053 | 0.2989 | ||
| O. latipes | 0.0969 | 0.0002 | 0.2506 | 0.0070 | |||
| O. woworae | 0.0360 | 0.0002 | 0.0211 | 0.2902 | |||
| O. sinensis | 0.7053 | 0.2506 | 0.0211 | 0.1823 | |||
| D. rerio | 0.2989 | 0.0070 | 0.2902 | 0.1823 | |||
| Shoaling Test | Average Inter-fish Distance | O. dancena | 0.3414 | <0.0001 | 0.0014 | <0.0001 | |
| O. latipes | 0.3414 | 0.0021 | 0.0193 | 0.0023 | |||
| O. woworae | <0.0001 | 0.0021 | 0.6498 | 0.9773 | |||
| O. sinensis | 0.0014 | 0.0193 | 0.6498 | 0.6685 | |||
| D. rerio | <0.0001 | 0.0023 | 0.9773 | 0.6685 | |||
| Average Shoal Area | O. dancena | 0.1811 | 0.0043 | 0.0036 | 0.0010 | ||
| O. latipes | 0.1811 | 0.1294 | 0.0888 | 0.0503 | |||
| O. woworae | 0.0043 | 0.1294 | 0.7447 | 0.6597 | |||
| O. sinensis | 0.0036 | 0.0888 | 0.7447 | 0.9410 | |||
| D. rerio | 0.0010 | 0.0503 | 0.6597 | 0.9410 | |||
| Average Nearest Neighbor Distance | O. dancena | 0.0654 | <0.0001 | 0.0001 | <0.0001 | ||
| O. latipes | 0.0654 | 0.0056 | 0.0297 | 0.0066 | |||
| O. woworae | <0.0001 | 0.0056 | 0.7350 | 0.9572 | |||
| O. sinensis | 0.0001 | 0.0297 | 0.7350 | 0.7721 | |||
| D. rerio | <0.0001 | 0.0066 | 0.9572 | 0.7721 | |||
| Average Farthest Neighbor Distance | O. dancena | 0.5503 | <0.0001 | 0.0001 | <0.0001 | ||
| O. latipes | 0.5503 | 0.0006 | 0.0009 | 0.0008 | |||
| O. woworae | <0.0001 | 0.0006 | 0.8418 | 0.9270 | |||
| O. sinensis | 0.0001 | 0.0009 | 0.8418 | 0.7774 | |||
| D. rerio | <0.0001 | 0.0008 | 0.9270 | 0.7774 | |||
| Circadian Locomotor Activity Rhythm—Day Cycle | Average Speed | O. dancena | 0.0432 | 0.0403 | <0.0001 | <0.0001 | |
| O. latipes | 0.0432 | <0.0001 | <0.0001 | <0.0001 | |||
| O. woworae | 0.0403 | <0.0001 | <0.0001 | <0.0001 | |||
| O. sinensis | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||
| Average Angular Velocity | O. dancena | 0.4015 | 0.9343 | <0.0001 | <0.0001 | ||
| O. latipes | 0.4015 | 0.3569 | <0.0001 | <0.0001 | |||
| O. woworae | 0.9343 | 0.3569 | <0.0001 | <0.0001 | |||
| O. sinensis | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||
| Meandering | O. dancena | 0.0603 | 0.0097 | 0.5545 | <0.0001 | ||
| O. latipes | 0.0603 | <0.0001 | 0.2761 | <0.0001 | |||
| O. woworae | 0.0097 | <0.0001 | 0.0037 | <0.0001 | |||
| O. sinensis | 0.5545 | 0.2761 | 0.0037 | <0.0001 | |||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||
| Freezing Movement Time Ratio | O. dancena | 0.3357 | 0.3036 | <0.0001 | <0.0001 | ||
| O. latipes | 0.3357 | 0.0464 | <0.0001 | <0.0001 | |||
| O. woworae | 0.3036 | 0.0464 | <0.0001 | <0.0001 | |||
| O. sinensis | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||
| Swimming Movement Time Ratio | O. dancena | 0.1337 | 0.2348 | <0.0001 | <0.0001 | ||
| O. latipes | 0.1337 | 0.0072 | <0.0001 | 0.0086 | |||
| O. woworae | 0.2348 | 0.0072 | <0.0001 | <0.0001 | |||
| O. sinensis | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||
| D. rerio | <0.0001 | 0.0086 | <0.0001 | <0.0001 | |||
| Rapid Movement Time Ratio | O. dancena | 0.6290 | <0.0001 | 0.0087 | <0.0001 | ||
| O. latipes | 0.6290 | <0.0001 | 0.0022 | <0.0001 | |||
| O. woworae | <0.0001 | <0.0001 | 0.0066 | <0.0001 | |||
| O. sinensis | 0.0087 | 0.0022 | 0.0066 | <0.0001 | |||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||
| Circadian Locomotor Activity Rhythm—Night Cycle | Average Speed | O. dancena | 0.0602 | 0.0013 | <0.0001 | <0.0001 | |
| O. latipes | 0.0602 | <0.0001 | 0.0043 | <0.0001 | |||
| O. woworae | 0.0013 | <0.0001 | <0.0001 | <0.0001 | |||
| O. sinensis | <0.0001 | 0.0043 | <0.0001 | <0.0001 | |||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||
| Average Angular Velocity | O. dancena | 0.1734 | 0.0331 | 0.2380 | <0.0001 | ||
| O. latipes | 0.1734 | 0.0005 | 0.9701 | <0.0001 | |||
| O. woworae | 0.0331 | 0.0005 | 0.0020 | <0.0001 | |||
| O. sinensis | 0.2380 | 0.9701 | 0.0020 | <0.0001 | |||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||
| Meandering | O. dancena | 0.4948 | 0.0310 | <0.0001 | <0.0001 | ||
| O. latipes | 0.4948 | 0.0045 | 0.0002 | <0.0001 | |||
| O. woworae | 0.0310 | 0.0045 | <0.0001 | <0.0001 | |||
| O. sinensis | <0.0001 | 0.0002 | <0.0001 | <0.0001 | |||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||
| Freezing Movement Time Ratio | O. dancena | 0.0119 | 0.0208 | <0.0001 | <0.0001 | ||
| O. latipes | 0.0119 | <0.0001 | 0.0012 | <0.0001 | |||
| O. woworae | 0.0208 | <0.0001 | <0.0001 | <0.0001 | |||
| O. sinensis | <0.0001 | 0.0012 | <0.0001 | <0.0001 | |||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||
| Swimming Movement Time Ratio | O. dancena | 0.0106 | 0.0226 | <0.0001 | <0.0001 | ||
| O. latipes | 0.0106 | <0.0001 | 0.0009 | <0.0001 | |||
| O. woworae | 0.0226 | <0.0001 | <0.0001 | <0.0001 | |||
| O. sinensis | <0.0001 | 0.0009 | <0.0001 | <0.0001 | |||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||
| Rapid Movement Time Ratio | O. dancena | 0.0834 | <0.0001 | 0.0279 | <0.0001 | ||
| O. latipes | 0.0834 | <0.0001 | 0.5159 | <0.0001 | |||
| O. woworae | <0.0001 | <0.0001 | 0.0005 | <0.0001 | |||
| O. sinensis | 0.0279 | 0.5159 | 0.0005 | <0.0001 | |||
| D. rerio | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Biomarkers | Medaka Fishes | O. dancena | O. latipes | O. woworae | O. sinensis |
|---|---|---|---|---|---|
| 5-HT | O. dancena | 0.0407 | 0.1569 | 0.0488 | |
| O. latipes | 0.0407 | 0.0005 | 0.939 | ||
| O. woworae | 0.1569 | 0.0005 | 0.0007 | ||
| O. sinensis | 0.0488 | 0.939 | 0.0007 | ||
| CAT | O. dancena | 0.003 | 0.8633 | 0.0093 | |
| O. latipes | 0.0030 | 0.0052 | 0.7163 | ||
| O. woworae | 0.8633 | 0.0052 | 0.0151 | ||
| O. sinensis | 0.0093 | 0.7163 | 0.0151 | ||
| NE | O. dancena | 0.0036 | 0.5032 | 0.0583 | |
| O. latipes | 0.0036 | 0.0003 | 0.3107 | ||
| O. woworae | 0.5032 | 0.0003 | 0.0104 | ||
| O. sinensis | 0.0583 | 0.3107 | 0.0104 | ||
| DA | O. dancena | 0.0059 | 0.3199 | 0.0322 | |
| O. latipes | 0.0059 | 0.0002 | 0.5405 | ||
| O. woworae | 0.3199 | 0.0002 | 0.0017 | ||
| O. sinensis | 0.0322 | 0.5405 | 0.0017 | ||
| Cortisol | O. dancena | 0.0116 | 0.2138 | 0.0168 | |
| O. latipes | 0.0116 | 0.0002 | 0.8935 | ||
| O. woworae | 0.2138 | 0.0002 | 0.0003 | ||
| O. sinensis | 0.0168 | 0.8935 | 0.0003 | ||
| AChE | O. dancena | 0.0389 | 0.1308 | 0.3293 | |
| O. latipes | 0.0389 | 0.0003 | 0.2756 | ||
| O. woworae | 0.1308 | 0.0003 | 0.0129 | ||
| O. sinensis | 0.3293 | 0.2756 | 0.0129 | ||
| ROS | O. dancena | 0.0217 | 0.0692 | 0.0692 | |
| O. latipes | 0.0217 | <0.0001 | 0.6325 | ||
| O. woworae | 0.0692 | <0.0001 | 0.0003 | ||
| O. sinensis | 0.0692 | 0.6325 | 0.0003 |
| Index | Behavior Endpoints (Units) | Definition | Applied to |
|---|---|---|---|
| 1-1-1 & 1-2-1 | Average speed (cm s−1) | Total distance traveled by fish divided by total time duration | Novel Tank Test (1-1 for 0–11 min and 1-2 for 15–31 min) |
| 1-1-2 & 1-2-2 | Freezing time movement ratio (%) | Total percentage of time when fish’s speed less than 1 cm s−1 | |
| 1-1-3 & 1-2-3 | Swimming time movement ratio (%) | Total percentage of time when fish’s speed is between 1 and 10 cm s−1 | |
| 1-1-4 & 1-2-4 | Rapid movement ratio (%) | Total percentage of time when fish’s speed more than 10 cm s−1 | |
| 1-1-5 & 1-2-5 | Time in top duration (%) | Total time spent in the top portion of the novel tank in percentage | |
| 1-1-6 & 1-2-6 | Number of entries to the top | Total times fish enter the upper half of the tank | |
| 1-1-7 & 1-2-7 | Latency to enter the top (s) | The amount of time it takes the fish to cross into the upper half of the tank | |
| 1-1-8 & 1-2-8 | Total distance traveled in the top (cm) | Total distance traveled in the top portion of the novel tank | |
| 1-1-9 & 1-2-9 | Thigmotaxis (cm) | The average distance of the fish from the center of the tank | |
| 2-1 | Mirror biting time percentage (%) | Total percentage of time when fish stayed in the mirror biting zone | Mirror Biting Test |
| 2-2 | Longest duration in the mirror side percentage (%) | Total percentage of fish longest duration stayed in the mirror biting zone | |
| 3-1 | Approaching predator time percentage (%) | Total percentage of time when fish stayed in the approaching predator zone | Predator Avoidance Test |
| 3-2 | Average distance to the predator’s separator (cm) | Average distance of fish to the predator’s separator | |
| 4-1 | Conspecific interaction time percentage (%) | Total percentage of time when fish stayed in the conspecific interaction zone | Social Interaction Test |
| 4-2 | Average distance to the conspecific’s separator (cm) | Average distance of fish to the conspecific’s separator | |
| 4-3 | Longest conspecific interaction percentage (%) | Total percentage of fish longest duration stayed in the conspecific interaction | |
| 5-1 | Average inter-fish distance (cm) | Average distance between the body center of every member of the shoal | Shoaling Test |
| 5-2 | Average shoal area (cm2) | Average size of the shoal | |
| 5-3 | Average nearest neighbor distance (cm) | Distance for the body center of each fish to the closest neighboring fish | |
| 5-4 | Average farthest neighbor distance (cm) | Distance for the body center of each fish to the farthest neighboring fish | |
| 6-1-1 & 6-2-1 | Average speed (cm s−1) | Total distance traveled by fish divided by total time duration | Circadian Locomotor Activity Rhythm (6-1 for day cycle and 6-2 for night cycle) |
| 6-1-2 & 6-2-2 | Average Angular Velocity (°/s) | Total turning angle of fish divided by total time duration | |
| 6-1-3 & 6-2-3 | Meandering (°/m) | Total turning angle of fish divided by total distance traveled by fish | |
| 6-1-4 & 6-2-4 | Freezing time movement ratio (%) | Total percentage of time when fish’s speed less than 1 cm s−1 | |
| 6-1-5 & 6-2-5 | Swimming time movement ratio (%) | Total percentage of time when fish’s speed is between 1 and 10 cm s−1 | |
| 6-1-6 & 6-2-6 | Rapid movement ratio (%) | Total percentage of time when fish’s speed more than 10 cm s−1 |
| Index | Coefficient of Variation (%) | ||||
|---|---|---|---|---|---|
| Danio rerio | Oryzias dancena | Oryzias latipes | Oryzias woworae | Oryzias sinensis | |
| 1-1-1 | 30.18 | 76.43 | 81.78 | 70.83 | 64.87 |
| 1-1-2 | 85.28 | 67.35 | 50.16 | 66.54 | 50.09 |
| 1-1-3 | 11.72 | 50.75 | 39.62 | 40.12 | 51.07 |
| 1-1-4 | 119.26 | 183.56 | 300.39 | 127.78 | 151.28 |
| 1-1-5 | 94.04 | 125.63 | 163.79 | 130.83 | 69.09 |
| 1-1-6 | 82.26 | 122.4 | 156.09 | 125.42 | 90.9 |
| 1-1-7 | 150.18 | 102.78 | 74.5 | 90.45 | 180.32 |
| 1-1-8 | 99.42 | 127.26 | 173.59 | 139.06 | 81.21 |
| 1-1-9 | 33.2 | 33.02 | 33.81 | 31.92 | 37.59 |
| 1-2-1 | 36.2 | 64.82 | 57.81 | 43.05 | 61.88 |
| 1-2-2 | 71.59 | 80.08 | 70.07 | 72.67 | 57.6 |
| 1-2-3 | 8.83 | 47.66 | 32.26 | 19.36 | 46.96 |
| 1-2-4 | 153.56 | 149.89 | 223.11 | 107.58 | 173.84 |
| 1-2-5 | 61.45 | 92.21 | 83.51 | 69.71 | 62.6 |
| 1-2-6 | 78.05 | 91.71 | 89.13 | 67.03 | 88.95 |
| 1-2-7 | 164.92 | 136.08 | 142.32 | 156.62 | 207.24 |
| 1-2-8 | 70.03 | 96.2 | 106.1 | 74.88 | 70.96 |
| 1-2-9 | 35.58 | 36.74 | 30.87 | 31.25 | 28.83 |
| 2-1 | 47.42 | 60.31 | 64.69 | 36.62 | 42.34 |
| 2-2 | 83.05 | 104.5 | 88.44 | 71.91 | 79.35 |
| 3-1 | 137 | 96.47 | 128.2 | 160.5 | 226 |
| 3-2 | 13.84 | 32.45 | 28.23 | 25.17 | 21.64 |
| 4-1 | 33.86 | 45.1 | 81.95 | 43.7 | 68.01 |
| 4-2 | 55.24 | 45.46 | 31.03 | 72.45 | 55.06 |
| 4-3 | 96.13 | 109.6 | 131.6 | 94.31 | 92.97 |
| 5-1 | 20.73 | 47.48 | 31.46 | 27.87 | 39.97 |
| 5-2 | 46.02 | 66.53 | 56.85 | 54.23 | 76.82 |
| 5-3 | 20.73 | 51.52 | 28.68 | 27.49 | 44.87 |
| 5-4 | 19.33 | 34.87 | 27 | 25.25 | 28.02 |
| 6-1-1 | 43.78 | 47.25 | 64.57 | 45 | 61.46 |
| 6-1-2 | 44.33 | 45.96 | 51.86 | 50.71 | 71.5 |
| 6-1-3 | 71.4 | 46.4 | 51.83 | 48.35 | 58.84 |
| 6-1-4 | 205.7 | 47.12 | 52.79 | 52.03 | 36.53 |
| 6-1-5 | 42.51 | 37.84 | 48.91 | 38.6 | 73.53 |
| 6-1-6 | 49.05 | 265 | 293.3 | 177.9 | 171.9 |
| 6-2-1 | 70.4 | 57.58 | 69.35 | 76.38 | 42.71 |
| 6-2-2 | 62.52 | 47.03 | 49.12 | 44.53 | 50.04 |
| 6-2-3 | 64.63 | 51.86 | 51.74 | 64.83 | 54.78 |
| 6-2-4 | 68.4 | 32.2 | 28.96 | 47.31 | 11.73 |
| 6-2-5 | 38.88 | 77.81 | 91.62 | 73.25 | 72.81 |
| 6-2-6 | 170.9 | 217.7 | 225.2 | 217.1 | 220.2 |
| Average | 70.52683 | 81.77098 | 89.90951 | 74.16073 | 79.91122 |
References
- Hilgers, L.; Schwarzer, J. The untapped potential of medaka and its wild relatives. eLife 2019, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Nakao, R.; Kitagawa, T. Differences in the behavior and ecology of wild type medaka (Oryzias latipes complex) and an orange commercial variety (himedaka). J. Exp. Zool. Part A Ecol. Genet. Physiol. 2015, 323, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Ansai, S.; Hosokawa, H.; Maegawa, S.; Kinoshita, M. Chronic fluoxetine treatment induces anxiolytic responses and altered social behaviors in medaka, Oryzias latipes. Behav. Brain Res. 2016, 303, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Shima, A.; Mitani, H. Medaka as a research organism: Past, present and future. Mech. Dev. 2004, 121, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Lucon-Xiccato, T.; Conti, F.; Loosli, F.; Foulkes, N.S.; Bertolucci, C. Development of Open-Field Behaviour in the Medaka, Oryzias latipes. Biology 2020, 9, 389. [Google Scholar] [CrossRef] [PubMed]
- Khalil, F.; Kang, I.J.; Undap, S.; Tasmin, R.; Qiu, X.; Shimasaki, Y.; Oshima, Y. Alterations in social behavior of Japanese medaka (Oryzias latipes) in response to sublethal chlorpyrifos exposure. Chemosphere 2013, 92, 125–130. [Google Scholar] [CrossRef]
- OECD. Test No. 203: Fish, Acute Toxicity Test; OECD Guidelines for the Testing of Chemicals, Section 2; OECD: Paris, France, 1992. [Google Scholar]
- Lee, W.; Yang, K.-L. Using medaka embryos as a model system to study biological effects of the electromagnetic fields on development and behavior. Ecotoxicol. Environ. Saf. 2014, 108, 187–194. [Google Scholar] [CrossRef]
- Kinoshita, M.; Murata, K.; Naruse, K.; Tanaka, M. Medaka: Biology, Management, and Experimental Protocols; John Wiley & Sons: Honoken, NJ, USA, 2009. [Google Scholar]
- Hong, X.; Zha, J. Fish behavior: A promising model for aquatic toxicology research. Sci. Total. Environ. 2019, 686, 311–321. [Google Scholar] [CrossRef]
- Nakayama, K.; Oshima, Y.; Hiramatsu, K.; Shimasaki, Y.; Honjo, T. Effects of Polychlorinated Biphenyls on the Schooling Behavior of Japanese Medaka (Oryzias Latipes). Environ. Toxicol. Chem. 2005, 24, 2588–2593. [Google Scholar] [CrossRef]
- Ju, Z.; Wells, M.C.; Heater, S.J.; Walter, R.B. Multiple tissue gene expression analyses in Japanese medaka (Oryzias latipes) exposed to hypoxia. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 145, 134–144. [Google Scholar] [CrossRef]
- Naruse, K.; Tanaka, M.; Takeda, H. Medaka: A Model for Organogenesis, Human Disease, and Evolution; Springer: Japan, Tokyo, 2011. [Google Scholar]
- Naruse, K.; Tanaka, M.; Mita, K.; Shima, A.; Postlethwait, J.; Mitani, H. A Medaka Gene Map: The Trace of Ancestral Vertebrate Proto-Chromosomes Revealed by Comparative Gene Mapping. Genome Res. 2004, 14, 820–828. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, Y.; Yoshimoto, M.; Ito, H. A brain atlas of a wild-type inbred strain of the medaka, Oryzias latipes. Fish Biol. J. MEDAKA 1999, 10, 1–26. [Google Scholar]
- Naruse, K.; Fukamachi, S.; Mitani, H.; Kondo, M.; Matsuoka, T.; Kondo, S.; Hanamura, N.; Morita, Y.; Hasegawa, K.; Nishigaki, R.; et al. A Detailed Linkage Map of Medaka, Oryzias latipes: Comparative Genomics and Genome Evolution. Genetics 2000, 154, 1773–1784. [Google Scholar] [CrossRef]
- Khorasani, M.Z.; Hennig, S.; Imre, G.; Asakawa, S.; Palczewski, S.; Berger, A.; Hori, H.; Naruse, K.; Mitani, H.; Shima, A.; et al. A first generation physical map of the medaka genome in BACs essential for positional cloning and clone-by-clone based genomic sequencing. Mech. Dev. 2004, 121, 903–913. [Google Scholar] [CrossRef]
- Tsuboko-Ishii, S.; Kimura, T.; Shinya, M.; Suehiro, Y.; Okuyama, T.; Shimada, A.; Takeda, H.; Naruse, K.; Kubo, T.; Takeuchi, H. Genetic Control of Startle Behavior in Medaka Fish. PLoS ONE 2014, 9, e112527. [Google Scholar] [CrossRef] [Green Version]
- Matsunaga, W.; Watanabe, E. Habituation of medaka (Oryzias latipes) demonstrated by open-field testing. Behav. Process. 2010, 85, 142–150. [Google Scholar] [CrossRef]
- Réale, D.; Reader, S.M.; Sol, D.; McDougall, P.T.; Dingemanse, N.J. Integrating animal temperament within ecology and evolution. Biol. Rev. 2007, 82, 291–318. [Google Scholar] [CrossRef] [Green Version]
- Bouchard, T.J.; Loehlin, J.C. Genes, evolution, and personality. Behav. Genet. 2001, 31, 243–273. [Google Scholar] [CrossRef]
- Smith, B.R.; Blumstein, D. Fitness consequences of personality: A meta-analysis. Behav. Ecol. 2008, 19, 448–455. [Google Scholar] [CrossRef] [Green Version]
- Carter, A.J.; Feeney, W.E.; Marshall, H.H.; Cowlishaw, G.; Heinsohn, R. Animal personality: What are behavioural ecologists measuring? Biol. Rev. 2013, 88, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Sih, A.; Bell, A.; Johnson, J. Behavioral syndromes: An ecological and evolutionary overview. Trends Ecol. Evol. 2004, 19, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Koolhaas, J.; de Boer, S.; Coppens, C.; Buwalda, B. Neuroendocrinology of coping styles: Towards understanding the biology of individual variation. Front. Neuroendocrinol. 2010, 31, 307–321. [Google Scholar] [CrossRef]
- Saunders, B.T.; Robinson, E.T. Individual Variation in the Motivational Properties of Cocaine. Neuropsychopharmacology 2011, 36, 1668–1676. [Google Scholar] [CrossRef]
- Conrad, J.L.; Weinersmith, K.L.; Brodin, T.; Saltz, J.B.; Sih, A. Behavioural syndromes in fishes: A review with implications for ecology and fisheries management. J. Fish Biol. 2011, 78, 395–435. [Google Scholar] [CrossRef]
- Demin, K.A.; Lakstygal, A.M.; Alekseeva, P.A.; Sysoev, M.; de Abreu, M.S.; Alpyshov, E.T.; Serikuly, N.; Wang, D.; Wang, M.; Tang, Z.; et al. The role of intraspecies variation in fish neurobehavioral and neuropharmacological phenotypes in aquatic models. Aquat. Toxicol. 2019, 210, 44–55. [Google Scholar] [CrossRef]
- Audira, G.; Siregar, P.; Strungaru, S.-A.; Huang, J.-C.; Hsiao, C.-D. Which Zebrafish Strains Are More Suitable to Perform Behavioral Studies? A Comprehensive Comparison by Phenomic Approach. Biology 2020, 9, 200. [Google Scholar] [CrossRef]
- Hyodo-Taguchi, Y. Inbred Strains of the Medaka and their Characteristics; Egami, N., Yamagami, K., Shima, A., Eds.; Tokyo University Press: Tokyo, Japan, 1990; pp. 129–142. [Google Scholar]
- Ansai, S.; Mochida, K.; Fujimoto, S.; Mokodongan, D.F.; Sumarto, B.K.A.; Masengi, K.W.A.; Hadiaty, R.K.; Nagano, A.J.; Toyoda, A.; Naruse, K.; et al. Genome editing reveals fitness effects of a gene for sexual dichromatism in Sulawesian fishes. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Blaser, R.E.; Rosemberg, D.B. Measures of Anxiety in Zebrafish (Danio rerio): Dissociation of Black/White Preference and Novel Tank Test. PLoS ONE 2012, 7, e36931. [Google Scholar] [CrossRef] [Green Version]
- Levin, E.D.; Bencan, Z.; Cerutti, D.T. Anxiolytic effects of nicotine in zebrafish. Physiol. Behav. 2007, 90, 54–58. [Google Scholar] [CrossRef]
- Cachat, J.; Stewart, A.; Grossman, L.; Gaikwad, S.; Kadri, F.; Chung, K.M.; Wu, N.; Wong, K.; Roy, S.; Suciu, C.; et al. Measuring behavioral and endocrine responses to novelty stress in adult zebrafish. Nat. Protoc. 2010, 5, 1786–1799. [Google Scholar] [CrossRef]
- Pham, M.; Raymond, J.; Hester, J.; Kyzar, E.; Gaikwad, S.; Bruce, I.; Fryar, C.; Chanin, S.; Enriquez, J.; Bagawandoss, S. Assessing social behavior phenotypes in adult zebrafish: Shoaling, social preference, and mirror biting tests. Zebrafish Protoc. Neurobehav. Res. 2012, 66, 231–246. [Google Scholar] [CrossRef]
- Ahmed, O.; Séguin, D.; Gerlai, R. An automated predator avoidance task in zebrafish. Behav. Brain Res. 2011, 216, 166–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audira, G.; Sampurna, B.P.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Hsiao, C.-D. A Versatile Setup for Measuring Multiple Behavior Endpoints in Zebrafish. Inventions 2018, 3, 75. [Google Scholar] [CrossRef] [Green Version]
- Engeszer, R.E.; Da Barbiano, L.A.; Ryan, M.J.; Parichy, D.M. Timing and plasticity of shoaling behaviour in the zebrafish, Danio rerio. Anim. Behav. 2007, 74, 1269–1275. [Google Scholar] [CrossRef] [Green Version]
- Miller, N.; Gerlai, R. Quantification of shoaling behaviour in zebrafish (Danio rerio). Behav. Brain Res. 2007, 184, 157–166. [Google Scholar] [CrossRef]
- Gwinner, F. Testosterone Induces “Splitting” of Circadian Locomotor Activity Rhythms in Birds. Science 1974, 185, 72–74. [Google Scholar] [CrossRef]
- Baker, M.R.; Goodman, A.C.; Santo, J.B.; Wong, R.Y. Repeatability and reliability of exploratory behavior in proactive and reactive zebrafish, Danio rerio. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Wong, K.; Elegante, M.; Bartels, B.; Elkhayat, S.; Tien, D.; Roy, S.; Goodspeed, J.; Suciu, C.; Tan, J.; Grimes, C.; et al. Analyzing habituation responses to novelty in zebrafish (Danio rerio). Behav. Brain Res. 2010, 208, 450–457. [Google Scholar] [CrossRef]
- Sataa, N.S.A.M.; Bakar, N.A.; Hodin, N.A.S.; Ramlan, N.F.; Ibrahim, W.N.W. Behavioral Responses of Javanese Medaka (Oryzias Javanicus) Versus Zebrafish (Danio Rerio) in Open Field Test. Res. Sq. 2020, 1–24. [Google Scholar] [CrossRef]
- Schnörr, S.; Steenbergen, P.; Richardson, M.; Champagne, D. Measuring thigmotaxis in larval zebrafish. Behav. Brain Res. 2012, 228, 367–374. [Google Scholar] [CrossRef]
- Peitsaro, N.; Kaslin, J.; Anichtchik, O.V.; Panula, P. Modulation of the histaminergic system and behaviour by α-fluoromethylhistidine in zebrafish. J. Neurochem. 2004, 86, 432–441. [Google Scholar] [CrossRef]
- Nakayasu, T.; Yasugi, M.; Shiraishi, S.; Uchida, S.; Watanabe, E. Three-dimensional computer graphic animations for studying social approach behaviour in medaka fish: Effects of systematic manipulation of morphological and motion cues. PLoS ONE 2017, 12, e0175059. [Google Scholar] [CrossRef]
- Tsubokawa, T.; Saito, K.; Kawano, H.; Kawamura, K.; Shinozuka, K.; Watanabe, S. Pharmacological effects on mirror approaching behavior and neurochemical aspects of the telencephalon in the fish, medaka (Oryzias latipes). Soc. Neurosci. 2009, 4, 276–286. [Google Scholar] [CrossRef]
- Otsuka, A.; Inahata, M.; Shimomura, Y.; Kagawa, N. Physiological changes in response to social isolation in male medaka fish. Fish. Sci. 2020, 86, 1–7. [Google Scholar] [CrossRef]
- De Abreu, M.S.; Koakoski, G.; Ferreira, D.; Oliveira, T.A.; Da Rosa, J.G.S.; Gusso, D.; Giacomini, A.C.V.; Piato, A.L.; Gil Barcellos, L.J. Diazepam and Fluoxetine Decrease the Stress Response in Zebrafish. PLoS ONE 2014, 9, e103232. [Google Scholar] [CrossRef]
- Dickinson, B.C.; Chang, C.J. Chemistry and biology of reactive oxygen species in signaling or stress responses. Nat. Chem. Biol. 2011, 7, 504–511. [Google Scholar] [CrossRef] [Green Version]
- Mocelin, R.N.; Herrmann, A.P.; Marcon, M.; Rambo, C.L.; Rohden, A.; Bevilaqua, F.; de Abreu, M.S.; Zanatta, L.; Elisabetsky, E.; Barcellos, L.J.; et al. N-acetylcysteine prevents stress-induced anxiety behavior in zebrafish. Pharmacol. Biochem. Behav. 2015, 139, 121–126. [Google Scholar] [CrossRef]
- Polavarapu, R.; Spitz, D.R.; Sim, J.E.; Follansbee, M.H.; Oberley, L.W.; Rahemtulla, A.; Nanji, A.A. Increased lipid peroxidation and impaired antioxidant enzyme function is associated with pathological liver injury in experimental alcoholic liver disease in rats fed diets high in corn oil and fish oil. Hepatology 1998, 27, 1317–1323. [Google Scholar] [CrossRef]
- Abdel-Salam, O.M.E.; Youness, E.R.; Morsy, F.A.; Mahfouz, M.M.; Kenawy, S.A. Study of the effect of antidepressant drugs and donepezil on aluminum-induced memory impairment and biochemical alterations in rats. Comp. Haematol. Int. 2015, 24, 847–860. [Google Scholar] [CrossRef]
- Lillesaar, C. The serotonergic system in fish. J. Chem. Neuroanat. 2011, 41, 294–308. [Google Scholar] [CrossRef]
- Mathuru, A.S. Conspecific injury raises an alarm in medaka. Sci. Rep. 2016, 6, 36615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reznick, D.N.; Ghalambor, C.K.; Crooks, K. Experimental studies of evolution in guppies: A model for understanding the evolutionary consequences of predator removal in natural communities. Mol. Ecol. 2008, 17, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, M.; Dudai, Y. Reconsolidation of fresh, remote, and extinguished fear memory in Medaka: Old fears don’t die. Eur. J. Neurosci. 2004, 20, 3397–3403. [Google Scholar] [CrossRef] [PubMed]
- Ladu, F.; Bartolini, T.; Panitz, S.G.; Chiarotti, F.; Butail, S.; Macrí, S.; Porfiri, M. Live Predators, Robots, and Computer-Animated Images Elicit Differential Avoidance Responses in Zebrafish. Zebrafish 2015, 12, 205–214. [Google Scholar] [CrossRef]
- Bass, S.L.; Gerlai, R. Zebrafish (Danio rerio) responds differentially to stimulus fish: The effects of sympatric and allopatric predators and harmless fish. Behav. Brain Res. 2008, 186, 107–117. [Google Scholar] [CrossRef]
- Mosienko, V.; Bert, B.; Beis, D.; Matthes, S.; Fink, H.; Bader, M.; Alenina, N. Exaggerated aggression and decreased anxiety in mice deficient in brain serotonin. Transl. Psychiatry 2012, 2, e122. [Google Scholar] [CrossRef] [Green Version]
- Ball, G.F.; Balthazart, J.; McCarthy, M.M. Is it useful to view the brain as a secondary sexual characteristic? Neurosci. Biobehav. Rev. 2014, 46, 628–638. [Google Scholar] [CrossRef]
- Mason, W. Ontogeny of social behavior. In Social Behavior and Communication; Springer: Berlin/Heidelberg, Germany, 1979; pp. 1–28. [Google Scholar]
- Isoe, Y.; Konagaya, Y.; Yokoi, S.; Kubo, T.; Takeuchi, H. Ontogeny and Sexual Differences in Swimming Proximity to Conspecifics in Response to Visual Cues in Medaka Fish. Zool. Sci. 2016, 33, 246–254. [Google Scholar] [CrossRef]
- Shibai, A.; Arimoto, T.; Yoshinaga, T.; Tsuchizawa, Y.; Khureltulga, D.; Brown, Z.P.; Kakizuka, T.; Hosoda, K. Attraction of posture and motion-trajectory elements of conspecific biological motion in medaka fish. Sci. Rep. 2018, 8, 8589. [Google Scholar] [CrossRef] [Green Version]
- Saverino, C.; Gerlai, R. The social zebrafish: Behavioral responses to conspecific, heterospecific, and computer animated fish. Behav. Brain Res. 2008, 191, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Imada, H.; Hoki, M.; Suehiro, Y.; Okuyama, T.; Kurabayashi, D.; Shimada, A.; Naruse, K.; Takeda, H.; Kubo, T.; Takeuchi, H. Coordinated and Cohesive Movement of Two Small Conspecific Fish Induced by Eliciting a Simultaneous Optomotor Response. PLoS ONE 2010, 5, e11248. [Google Scholar] [CrossRef]
- Nakayasu, T.; Watanabe, E. Biological motion stimuli are attractive to medaka fish. Anim. Cogn. 2013, 17, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Winberg, S.; Nilsson, G.E. Roles of brain monoamine neurotransmitters in agonistic behaviour and stress reactions, with particular reference to fish. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1993, 106, 597–614. [Google Scholar] [CrossRef]
- Øverli, Ø.; Harris, C.A.; Winberg, S. Short-Term Effects of Fights for Social Dominance and the Establishment of Dominant-Subordinate Relationships on Brain Monoamines and Cortisol in Rainbow Trout. Brain Behav. Evol. 1999, 54, 263–275. [Google Scholar] [CrossRef]
- Hare, A.J.; Waheed, A.; Hare, J.F.; Anderson, W.G. Cortisol and catecholamine responses to social context and a chemical alarm signal in juvenile lake sturgeon, Acipenser fulvescens. Can. J. Zool. 2015, 93, 605–613. [Google Scholar] [CrossRef]
- Shams, S.; Amlani, S.; Buske, C.; Chatterjee, D.; Gerlai, R. Developmental social isolation affects adult behavior, social interaction, and dopamine metabolite levels in zebrafish. Dev. Psychobiol. 2018, 60, 43–56. [Google Scholar] [CrossRef]
- Saif, M.; Chatterjee, D.; Buske, C.; Gerlai, R. Sight of conspecific images induces changes in neurochemistry in zebrafish. Behav. Brain Res. 2013, 243, 294–299. [Google Scholar] [CrossRef]
- Egami, N. Effect of artificial photoperiodicity on time of oviposition in the fish, Oryzias latipes. Anim. Behav. 1954, 27, 57–62. [Google Scholar]
- Ueda, M.; Oishi, T. Circadian oviposition rhythm and locomotor activity in the medaka, Oryzias latipes. J. Interdiscip. Cycle Res. 1982, 13, 97–104. [Google Scholar] [CrossRef]
- Karigo, T.; Kanda, S.; Takahashi, A.; Abe, H.; Okubo, K.; Oka, Y. Time-of-Day-Dependent Changes in GnRH1 Neuronal Activities and Gonadotropin mRNA Expression in a Daily Spawning Fish, Medaka. Endocrinology 2012, 153, 3394–3404. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.A. Rhythms in Fishes; Springer: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Weber, D.N.; Spieler, R.E. Effects of the light-dark cycle and scheduled feeding on behavioral and reproductive rhythms of the cyprinodont fish, Medaka, Oryzias latipes. Cell. Mol. Life Sci. 1987, 43, 621–624. [Google Scholar] [CrossRef]
- Thi, N.H.B.; Audira, G.; Siregar, P.; Liang, S.-T.; Huang, J.-C.; Hsiao, C.-D.; Thi, N.A.N.; Thi, N.B.; Thi, N.N. Chronic Exposure to Low Concentration Lead Chloride-Induced Anxiety and Loss of Aggression and Memory in Zebrafish. Int. J. Mol. Sci. 2020, 21, 1844. [Google Scholar] [CrossRef] [Green Version]
- Roshchina, V.V. Evolutionary Considerations of Neurotransmitters in Microbial, Plant, and Animal Cells. Microb. Endocrinol. 2010, 17–52. [Google Scholar] [CrossRef]
- Van Ginneken, V.; Nieveen, M.; Van Eersel, R.; Thillart, G.V.D.; Addink, A. Neurotransmitter levels and energy status in brain of fish species with and without the survival strategy of metabolic depression. Comp. Biochem. Physiol. Part A Physiol. 1996, 114, 189–196. [Google Scholar] [CrossRef]
- Porteus, C.S.; Brink, D.L.; Milsom, W.K. Neurotransmitter profiles in fish gills: Putative gill oxygen chemoreceptors. Respir. Physiol. Neurobiol. 2012, 184, 316–325. [Google Scholar] [CrossRef]
- Reid, S.G.; Bernier, N.J.; Perry, S.F. The adrenergic stress response in fish: Control of catecholamine storage and release. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1998, 120, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Aprison, M.; Shank, R.; Davidoff, R.; Werman, R. The distribution of glycine, a neurotransmitter suspect in the central nervous system of several vertebrate species. Life Sci. 1968, 7, 583–590. [Google Scholar] [CrossRef]
- Wilkinson, G.S.; Breden, F.; Mank, J.; Ritchie, M.G.; Higginson, A.D.; Radwan, J.; Jaquiery, J.; Salzburger, W.; Arriero, E.; Barribeau, S.M.; et al. The locus of sexual selection: Moving sexual selection studies into the post-genomics era. J. Evol. Biol. 2015, 28, 739–755. [Google Scholar] [CrossRef] [Green Version]
- Yoon, M.; Kim, K.-Y.; Bang, I.-C.; Nam, Y.K.; Kim, D.S. Complete mitogenome sequence of the Chinese medaka Oryzias sinensis (Teleostei: Beloniformes) and its phylogenetic analysis. Genes Genom. 2011, 33, 307–312. [Google Scholar] [CrossRef]
- Sakamoto, T.; Kozaka, T.; Takahashi, A.; Kawauchi, H.; Ando, M. Medaka (Oryzias latipes) as a model for hypoosmoregulation of euryhaline fishes. Aquaculture 2001, 193, 347–354. [Google Scholar] [CrossRef]
- Kim, Y.K.; Lee, S.Y.; Kim, B.S.; Kim, D.S.; Nam, Y.K. Isolation and mRNA expression analysis of aquaporin isoforms in marine medaka Oryzias dancena, a euryhaline teleost. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2014, 171, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Parenti, L.R. A phylogenetic analysis and taxonomic revision of ricefishes, Oryzias and relatives (Beloniformes, Adrianichthyidae). Zool. J. Linn. Soc. 2008, 154, 494–610. [Google Scholar] [CrossRef] [Green Version]
- Juo, J.-J.; Kang, C.-K.; Yang, W.-K.; Yang, S.-Y.; Lee, T.-H. A Stenohaline Medaka, Oryzias woworae, Increases Expression of Gill Na, K-ATPase and Na, K, 2Cl-Cotransporter 1 to Tolerate Osmotic Stress. Zool. Sci. 2016, 33, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.-H.; Lin, L.-Y.; Tseng, Y.-C.; Horng, J.-L.; Hwang, P.-P. A new model for fish ion regulation: Identification of ionocytes in freshwater- and seawater-acclimated medaka (Oryzias latipes). Cell Tissue Res. 2014, 357, 225–243. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y. Taxonomy and distribution of the genus Oryzias in Yunnan, China. Acta Taxonom. Sin. 1989, 14, 239–246. [Google Scholar]
- Takehana, Y.; Naruse, K.; Sakaizumi, M. Molecular phylogeny of the medaka fishes genus Oryzias (Beloniformes: Adrianichthyidae) based on nuclear and mitochondrial DNA sequences. Mol. Phylogenet. Evol. 2005, 36, 417–428. [Google Scholar] [CrossRef]
- Kang, C.-K.; Tsai, S.-C.; Lee, T.-H.; Hwang, P.-P. Differential expression of branchial Na+/K+-ATPase of two medaka species, Oryzias latipes and Oryzias dancena, with different salinity tolerances acclimated to fresh water, brackish water and seawater. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 151, 566–575. [Google Scholar] [CrossRef]
- Cui, L.; Fan, M.; Belanger, S.; Li, J.; Wang, X.; Fan, B.; Li, W.; Gao, X.; Chen, J.; Liu, Z. Oryzias sinensis, a new model organism in the application of eco-toxicity and water quality criteria (WQC). Chemosphere 2020, 261, 127813. [Google Scholar] [CrossRef]
- Myosho, T.; Takahashi, H.; Yoshida, K.; Sato, T.; Hamaguchi, S.; Sakamoto, T.; Sakaizumi, M. Hyperosmotic tolerance of adult fish and early embryos are determined by discrete, single loci in the genus Oryzias. Sci. Rep. 2018, 8, 6897. [Google Scholar] [CrossRef]
- Chen, W.-J.; Orti, G.; Meyer, A. Novel evolutionary relationship among four fish model systems. Trends Genet. 2004, 20, 424–431. [Google Scholar] [CrossRef] [Green Version]
- Colón-Cruz, L.; Kristofco, L.; Crooke-Rosado, J.; Acevedo, A.; Torrado, A.; Brooks, B.W.; Sosa, M.A.; Behra, M. Alterations of larval photo-dependent swimming responses (PDR): New endpoints for rapid and diagnostic screening of aquatic contamination. Ecotoxicol. Environ. Saf. 2018, 147, 670–680. [Google Scholar] [CrossRef]
- Spence, R.; Gerlach, G.; Lawrence, C.; Smith, C. The behaviour and ecology of the zebrafish, Danio rerio. Biol. Rev. 2007, 83, 13–34. [Google Scholar] [CrossRef]
- Avdesh, A.; Chen, M.; Martin-Iverson, M.T.; Mondal, A.; Ong, D.; Rainey-Smith, S.; Taddei, K.; Lardelli, M.; Groth, D.M.; Verdile, G.; et al. Regular Care and Maintenance of a Zebrafish (Danio rerio) Laboratory: An Introduction. J. Vis. Exp. 2012, e4196. [Google Scholar] [CrossRef] [Green Version]
- Audira, G.; Sampurna, B.P.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Han, L.; Hsiao, C.-D. Establishing simple image-based methods and cost-effective instrument for toxicity assessment on circadian rhythm dysregulation in fish. Biol. Open 2019, 8, 041871. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Escudero, A.; Vicente-Page, J.; Hinz, R.C.; Arganda, S.; De Polavieja, G.G. idTracker: Tracking individuals in a group by automatic identification of unmarked animals. Nat. Methods 2014, 11, 743–748. [Google Scholar] [CrossRef]
- Adams, D.C.; Anthony, C.D. Using randomization techniques to analyse behavioural data. Anim. Behav. 1996, 51, 733–738. [Google Scholar] [CrossRef] [Green Version]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Furutani-Seiki, M.; Wittbrodt, J. Medaka and zebrafish, an evolutionary twin study. Mech. Dev. 2004, 121, 629–637. [Google Scholar] [CrossRef]
- Wittbrodt, J.; Shima, A.; Schartl, M. Medaka—A model organism from the far East. Nat. Rev. Genet. 2002, 3, 53–64. [Google Scholar] [CrossRef]
- Sasado, T.; Tanaka, M.; Kobayashi, K.; Sato, T.; Sakaizumi, M.; Naruse, K. The National BioResource Project Medaka (NBRP Medaka): An Integrated Bioresource for Biological and Biomedical Sciences. Exp. Anim. 2010, 59, 13–23. [Google Scholar] [CrossRef] [Green Version]




| Biomarkers | O. dancena | O. latipes | O. woworae | O. sinensis | Unit | ||||
|---|---|---|---|---|---|---|---|---|---|
| Concentration | SSD * Letter | Concentration | SSD Letter | Concentration | SSD Letter | Concentration | SSD Letter | ||
| 5-HT (Serotonin) | 36.21 ± 11.25 | a | 25.95 ± 5.634 | b | 40.59 ± 7.444 | a | 27.16 ± 7.525 | b | ng/total protein (mg) |
| CAT (Catalase) | 15.83 ± 5.214 | a, c | 10.16 ± 2.194 | b | 14.35 ± 2.980 | a | 10.77 ± 2.829 | b, c | ng/total protein (mg) |
| NE (Norepinephrine) | 1.512 ± 0.6271 | a | 0.8307 ± 0.2756 | b | 1.653 ± 0.5085 | a | 1.011 ± 0.4499 | b | ng/total protein (mg) |
| DA (Dopamine) | 22.86 ± 9.282 | a | 12.84 ± 3.738 | b | 24.93 ± 5.888 | a | 14.45 ± 4.942 | b | pg/total protein (mg) |
| Cortisol | 228.1 ± 69.08 | a | 135.5 ± 44.60 | b | 283.3 ± 68.17 | a | 146.9 ± 68.29 | b | pg/total protein (mg) |
| AChE (Acetylcholinesterase) | 29.82 ± 6.940 | a, c | 17.09 ± 9.654 | b | 35.92 ± 7.873 | a | 23.44 ± 15.40 | b, c | U/total protein (mg) |
| ROS (Reactive Oxygen Species) | 133.5 ± 44.48 | a, c | 80.81 ± 31.65 | b | 199 ± 69.21 | a | 89.78 ± 44.00 | b, c | IU/total protein (mg) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Audira, G.; Siregar, P.; Chen, K.H.-C.; Roldan, M.J.M.; Huang, J.-C.; Lai, H.-T.; Hsiao, C.-D. Interspecies Behavioral Variability of Medaka Fish Assessed by Comparative Phenomics. Int. J. Mol. Sci. 2021, 22, 5686. https://doi.org/10.3390/ijms22115686
Audira G, Siregar P, Chen KH-C, Roldan MJM, Huang J-C, Lai H-T, Hsiao C-D. Interspecies Behavioral Variability of Medaka Fish Assessed by Comparative Phenomics. International Journal of Molecular Sciences. 2021; 22(11):5686. https://doi.org/10.3390/ijms22115686
Chicago/Turabian StyleAudira, Gilbert, Petrus Siregar, Kelvin H.-C. Chen, Marri Jmelou M. Roldan, Jong-Chin Huang, Hong-Thih Lai, and Chung-Der Hsiao. 2021. "Interspecies Behavioral Variability of Medaka Fish Assessed by Comparative Phenomics" International Journal of Molecular Sciences 22, no. 11: 5686. https://doi.org/10.3390/ijms22115686
APA StyleAudira, G., Siregar, P., Chen, K. H.-C., Roldan, M. J. M., Huang, J.-C., Lai, H.-T., & Hsiao, C.-D. (2021). Interspecies Behavioral Variability of Medaka Fish Assessed by Comparative Phenomics. International Journal of Molecular Sciences, 22(11), 5686. https://doi.org/10.3390/ijms22115686