
Novel S. cerevisiae Hybrid Synthetic Promoters Based on Foreign Core Promoter Sequences



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
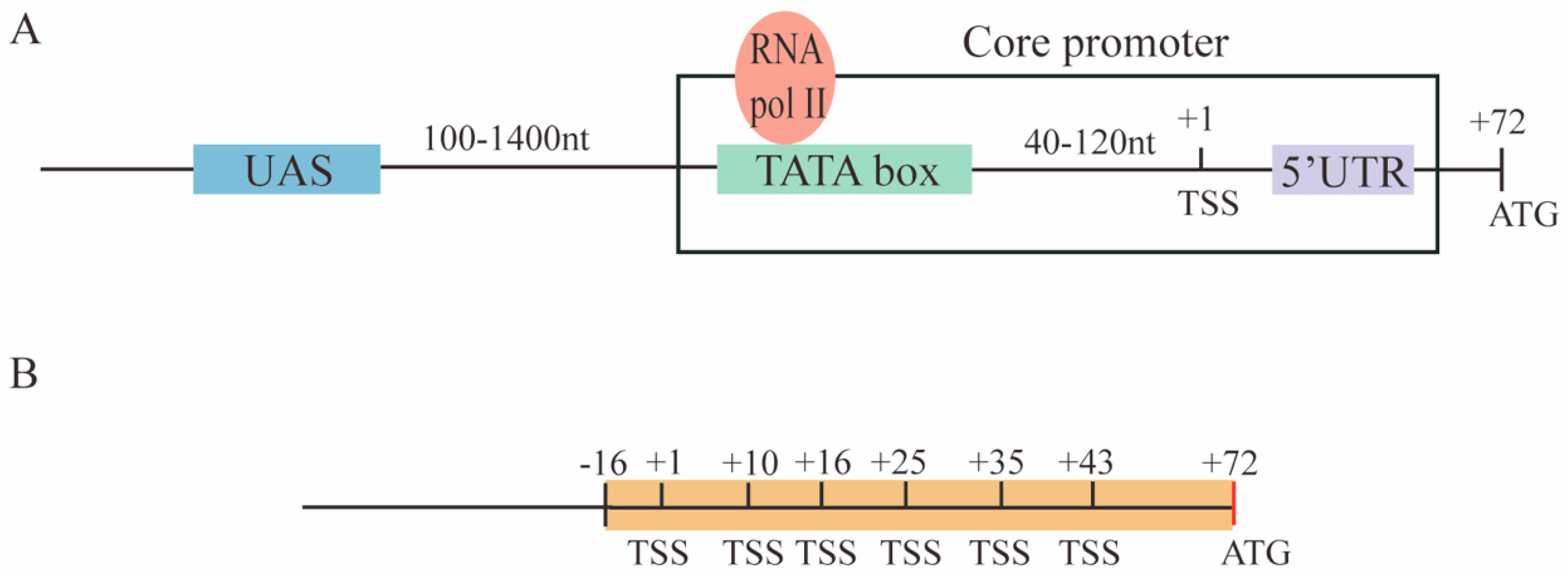
2.1. Constructing New Synthetic Core Promoters
2.2. Expanding the Promoter Library by Mutating the TATA Box
2.3. Finding the Optimal Distance between the TATA Box and the TSS
2.4. Nucleosome Removal
2.5. Building New Synthetic Promoters Via the Addition of an UAS
2.6. Final Promoter Optimization
3. Conclusions
4. Materials and Methods
4.1. Plasmids Construction
4.2. Yeast Strain Construction
4.3. Flow Cytometry
4.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Davis, R.W.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.D.; Jacq, C.; Johnston, M.; et al. Life with 6000 Genes. Science 1996, 274, 546–567. [Google Scholar] [CrossRef] [PubMed]
- Barnett, J.A. Beginnings of microbiology and biochemistry: The contribution of yeast research. Microbiology 2003, 149, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, Y.; Nielsen, J. Synthetic Biology of Yeast. Biochemistry 2019, 58, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Ro, D.-K.; Paradise, E.M.; Ouellet, M.; Fisher, K.J.; Newman, K.L.; Ndungu, J.M.; Ho, K.A.; Eachus, R.A.; Ham, T.S.; Kirby, J.; et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nat. Cell Biol. 2006, 440, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Pickens, L.B.; Tang, Y.; Chooi, Y.-H. Metabolic Engineering for the Production of Natural Products. Annu. Rev. Chem. Biomol. Eng. 2011, 2, 211–236. [Google Scholar] [CrossRef] [PubMed]
- Hubmann, G.; Thevelein, J.M.; Nevoigt, E. Natural and Modified Promoters for Tailored Metabolic Engineering of the Yeast Saccharomyces cerevisiae. Methods Mol. Biol. 2014, 1152, 17–42. [Google Scholar] [CrossRef] [PubMed]
- Van Werven, F.J.; Van Teeffelen, H.A.; Holstege, F.C.; Timmers, H.T.M. Distinct promoter dynamics of the basal transcription factor TBP across the yeast genome. Nat. Struct. Mol. Biol. 2009, 16, 1043. [Google Scholar] [CrossRef] [PubMed]
- Forsburg, S.L.; Guarente, L. Mutational analysis of upstream activation sequence 2 of the CYC1 gene of Saccharomyces cerevisiae: A HAP2-HAP3-responsive site. Mol. Cell. Biol. 1988, 8, 647–654. [Google Scholar] [CrossRef]
- Basehoar, A.D.; Zanton, S.J.; Pugh, B.F. Identification and distinct regulation of yeast TATA box-containing genes. Cell 2004, 117, 847. [Google Scholar] [CrossRef]
- Lubliner, S.; Regev, I.; Lotan-Pompan, M.; Edelheit, S.; Weinberger, A.; Segal, E. Core promoter sequence in yeast is a major determinant of expression level. Genome Res. 2015, 25, 1008–1017. [Google Scholar] [CrossRef] [PubMed]
- Wobbe, C.R.; Struhl, K. Yeast and human TATA-binding proteins have nearly identical DNA sequence requirements for transcription in vitro. Mol. Cell. Biol. 1990, 10, 3859–3867. [Google Scholar] [CrossRef]
- Mishra, A.K.; Vanathi, P.; Bhargava, P. The transcriptional activator GAL4-VP16 regulates the intra-molecular interactions of the TATA-binding protein. J. Biosci. 2003, 28, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Mogno, I.; Vallania, F.; Mitra, R.; Cohen, B.A. TATA is a modular component of synthetic promoters. Genome Res. 2010, 20, 1391–1397. [Google Scholar] [CrossRef] [PubMed]
- Blake, W.J.; Balázsi, G.; Kohanski, M.A.; Isaacs, F.J.; Murphy, K.F.; Kuang, Y.; Cantor, C.R.; Walt, D.R.; Collins, J.J. Phenotypic Consequences of Promoter-Mediated Transcriptional Noise. Mol. Cell 2006, 24, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Dietrich, F.S. Mapping of transcription start sites in Saccharomyces cerevisiae using 5’ SAGE. Nucleic Acids Res. 2005, 33, 2838–2851. [Google Scholar] [CrossRef] [PubMed]
- Hahn, S.; Hoar, E.T.; Guarente, L. Each of three “TATA elements” specifies a subset of the transcription initiation sites at the CYC-1 promoter of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1985, 82, 8562–8566. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, N.A.; Srikrishnan, S. Introduction and expression of genes for metabolic engineering applications in Saccharomyces cerevisiae. FEMS Yeast Res. 2012, 12, 197–214. [Google Scholar] [CrossRef] [PubMed]
- Nevoigt, E.; Kohnke, J.; Fischer, C.R.; Alper, H.; Stahl, U.; Stephanopoulos, G. Engineering of Promoter Replacement Cassettes for Fine-Tuning of Gene Expression in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2006, 72, 5266–5273. [Google Scholar] [CrossRef]
- Tang, H.; Wu, Y.; Deng, J.; Chen, N.; Zheng, Z.; Wei, Y.; Luo, X.; Keasling, J.D. Promoter Architecture and Promoter Engineering in Saccharomyces cerevisiae. Metabolites 2020, 10, 320. [Google Scholar] [CrossRef] [PubMed]
- Blazeck, J.; Garg, R.; Reed, B.; Alper, H.S. Controlling promoter strength and regulation in Saccharomyces cerevisiae using synthetic hybrid promoters. Biotechnol. Bioeng. 2012, 109, 2884–2895. [Google Scholar] [CrossRef]
- Gossen, M.; Bujard, H. Tight control of gene expression in mammalian cells by tetracycline-responsive promoters. Proc. Natl. Acad. Sci. USA 1992, 89, 5547–5551. [Google Scholar] [CrossRef] [PubMed]
- Garí, E.; Piedrafita, L.; Aldea, M.; Herrero, E. A Set of Vectors with a Tetracycline-Regulatable Promoter System for Modulated Gene Expression inSaccharomyces cerevisiae. Yeast 1997, 13, 837–848. [Google Scholar] [CrossRef]
- Belli, G. An activator/repressor dual system allows tight tetracycline-regulated gene expression in budding yeast. Nucleic Acids Res. 1998, 26, 942–947. [Google Scholar] [CrossRef] [PubMed]
- Romero-Santacreu, L.; Orozco, H.; Garre, E.; Alepuz, P. The bidirectional cytomegalovirus immediate/early promoter is regulated by Hog1 and the stress transcription factors Sko1 and Hot1 in yeast. Mol. Genet. Genom. 2010, 283, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Ede, C.; Chen, X.; Lin, M.-Y.; Chen, Y.Y. Quantitative Analyses of Core Promoters Enable Precise Engineering of Regulated Gene Expression in Mammalian Cells. ACS Synth. Biol. 2016, 5, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Li, J.; Liang, Q.; Marchisio, M.A. Can terminators be used as insulators into yeast synthetic gene circuits? J. Biol. Eng. 2016, 10, 19. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Stinski, M.F. Binding of cellular repressor protein or the IE2 protein to a cis-acting negative regulatory element upstream of a human cytomegalovirus early promoter. J. Virol. 1995, 69, 7612–7621. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Stinski, M.F. Activation of Transcription of the Human Cytomegalovirus Early UL4 Promoter by the Ets Transcription Factor Binding Element. J. Virol. 2000, 74, 9845–9857. [Google Scholar] [CrossRef][Green Version]
- Byrne, B.J.; Davis, M.S.; Yamaguchi, J.; Bergsma, D.J.; Subramanian, K.N. Definition of the simian virus 40 early promoter region and demonstration of a host range bias in the enhancement effect of the simian virus 40 72-base-pair repeat. Proc. Natl. Acad. Sci. USA 1983, 80, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Maundrell, K. nmt1 of fission yeast. A highly transcribed gene completely repressed by thiamine. J. Biol. Chem. 1990, 265, 10857–10864. [Google Scholar] [CrossRef]
- Redden, H.; Alper, H.S. The development and characterization of synthetic minimal yeast promoters. Nat. Commun. 2015, 6, 7810. [Google Scholar] [CrossRef] [PubMed]
- Ribas, J.C.; Moreno, M.B.; Durán, A. A family of multifunctional thiamine-repressible expression vectors for fission yeast. Yeast 2000, 16, 861–872. [Google Scholar] [CrossRef]
- Lewis, E.D.; Manley, J.L. Control of adenovirus late promoter expression in two human cell lines. Mol. Cell. Biol. 1985, 5, 2433–2442. [Google Scholar] [CrossRef] [PubMed]
- Odell, J.T.; Nagy, F.; Chua, N.-H. Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter. Nat. Cell Biol. 1985, 313, 810–812. [Google Scholar] [CrossRef] [PubMed]
- McKnight, S.L.; Gavis, E.R.; Kingsbury, R.; Axel, R. Analysis of transcriptional regulatory signals of the HSV thymidine kinase gene: Identification of an upstream control region. Cell 1981, 25, 385–398. [Google Scholar] [CrossRef]
- Nakajima, K.; Kusafuka, T.; Takeda, T.; Fujitani, Y.; Nakae, K.; Hirano, T. Identification of a novel interleukin-6 response element containing an Ets-binding site and a CRE-like site in the junB promoter. Mol. Cell. Biol. 1993, 13, 3027–3041. [Google Scholar] [CrossRef]
- Yu, L.; Marchisio, M.A. Saccharomyces cerevisiae Synthetic Transcriptional Networks Harnessing dCas12a and Type V-A anti-CRISPR Proteins. ACS Synth. Biol. 2021, 10, 870–883. [Google Scholar] [CrossRef]
- Raveh-Sadka, T.; Levo, M.; Shabi, U.; Shany, B.; Keren, L.; Lotan-Pompan, M.; Zeevi, D.; Sharon, E.; Weinberger, A.; Segal, E. Manipulating nucleosome disfavoring sequences allows fine-tune regulation of gene expression in yeast. Nat. Genet. 2012, 44, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Lubliner, S.; Keren, L.; Segal, E. Sequence features of yeast and human core promoters that are predictive of maximal promoter activity. Nucleic Acids Res. 2013, 41, 5569–5581. [Google Scholar] [CrossRef] [PubMed]
- Curran, K.A.; Crook, N.C.; Karim, A.S.; Gupta, A.R.; Wagman, A.M.; Alper, H.S. Design of synthetic yeast promoters via tuning of nucleosome architecture. Nat. Commun. 2014, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.; Fondufe-Mittendorf, Y.; Xia, L.; Flatow, J.; Widom, J.; Wang, J.-P. Predicting nucleosome positioning using a duration Hidden Markov Model. BMC Bioinform. 2010, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bitter, G.A.; Chang, K.K.H.; Egan, K.M. A multi-component upstream activation sequence of the Saccharomyces cerevisiae glyceraldehyde-3-phosphate dehydrogenase gene promoter. Mol. Genet. Genom. 1991, 231, 22–32. [Google Scholar] [CrossRef]
- Decoene, T.; De Maeseneire, S.L.; De Mey, M. Modulating transcription through development of semi-synthetic yeast core promoters. PLoS ONE 2019, 14, e0224476. [Google Scholar] [CrossRef]
- Vignais, M.L.; Huet, J.; Buhler, J.M.; Sentenac, A. Contacts between the factor TUF and RPG sequences. J. Biol. Chem. 1990, 265, 14669–14674. [Google Scholar] [CrossRef]
- Kurtz, S.; Shore, D. RAP1 protein activates and silences transcription of mating-type genes in yeast. Genes Dev. 1991, 5, 616–628. [Google Scholar] [CrossRef]
- Qiu, C.; Jin, H.; Vvedenskaya, I.; Llenas, J.A.; Zhao, T.; Malik, I.; Visbisky, A.M.; Schwartz, S.L.; Cui, P.; Čabart, P.; et al. Universal promoter scanning by Pol II during transcription initiation in Saccharomyces cerevisiae. Genome Biol. 2020, 21, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Chee, M.K.; Haase, S.B. New and Redesigned pRS Plasmid Shuttle Vectors for Genetic Manipulation of Saccharomycescerevisiae. G3 (Bethesda) 2012, 2, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Sheff, M.A.; Thorn, K.S. Optimized cassettes for fluorescent protein tagging in Saccharomyces cerevisiae. Yeast 2004, 21, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Mumberg, D.; Müller, R.; Funk, M. Yeast vectors for the controlled expression of heterologous proteins in different genetic backgrounds. Gene 1995, 156, 119–122. [Google Scholar] [CrossRef]
- Gibson, D. One-step enzymatic assembly of DNA molecules up to several hundred kilobases in size. Protoc. Exch. 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Touchdown Polymerase Chain Reaction (PCR). Cold Spring Harb. Protoc. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R. (Ed.) Molecular Cloning, 4th ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012. [Google Scholar]
- Gietz, R.D.; Woods, R.A. Transformation of yeast by lithium acetate/single-stranded carrier DNA/polyethylene glycol method. Methods Enzymol. 2002, 350, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Hahne, F.; LeMeur, N.; Brinkman, R.R.; Ellis, B.; Haaland, P.; Sarkar, D.; Spidlen, J.; Strain, E.; Gentleman, R. flowCore: A Bioconductor package for high throughput flow cytometry. BMC Bioinform. 2009, 10, 106. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, X.; Marchisio, M.A. Novel S. cerevisiae Hybrid Synthetic Promoters Based on Foreign Core Promoter Sequences. Int. J. Mol. Sci. 2021, 22, 5704. https://doi.org/10.3390/ijms22115704
Feng X, Marchisio MA. Novel S. cerevisiae Hybrid Synthetic Promoters Based on Foreign Core Promoter Sequences. International Journal of Molecular Sciences. 2021; 22(11):5704. https://doi.org/10.3390/ijms22115704
Chicago/Turabian StyleFeng, Xiaofan, and Mario Andrea Marchisio. 2021. "Novel S. cerevisiae Hybrid Synthetic Promoters Based on Foreign Core Promoter Sequences" International Journal of Molecular Sciences 22, no. 11: 5704. https://doi.org/10.3390/ijms22115704
APA StyleFeng, X., & Marchisio, M. A. (2021). Novel S. cerevisiae Hybrid Synthetic Promoters Based on Foreign Core Promoter Sequences. International Journal of Molecular Sciences, 22(11), 5704. https://doi.org/10.3390/ijms22115704