Abstract
Substance use/abuse is one of the main causes of depressive symptoms. Cannabis and synthetic cannabinoids in particular gained significant popularity in the past years. There is an increasing amount of clinical data associating such compounds with the inflammatory component of depression, indicated by the up-regulation of pro-inflammatory cytokines. Pro-inflammatory cytokines are also well-known to regulate the enzymes of the kynurenine pathway (KP), which is responsible for metabolizing tryptophan, a precursor in serotonin synthesis. Enhanced pro-inflammatory cytokine levels may over-activate the KP, leading to tryptophan depletion and reduced serotonin levels, which can subsequently precipitate depressive symptoms. Therefore, such mechanism might represent a possible link between the endocannabinoid system (ECS) and the KP in depression, via the inflammatory and dysregulated serotonergic component of the disorder. This review will summarize the data regarding those natural and synthetic cannabinoids that increase pro-inflammatory cytokines. Furthermore, the data on such cytokines associated with KP activation will be further reviewed accordingly. The interaction of the ECS and the KP has been postulated and demonstrated in some studies previously. This review will further contribute to this yet less explored connection and propose the KP to be the missing link between cannabinoid-induced inflammation and depressive symptoms.
1. Introduction
Depression affects more than 264 million people around the world [1] and social, environmental, as well as genetic factors may contribute to its development. Substance abuse is among the most frequent causes in the development of depression [2]. Recently, cannabis and synthetic cannabinoid use has gained popularity, unfortunately their misuse as well at the same time [3,4,5]. Moreover, there is a great amount of clinical evidence demonstrating that such compounds can induce depressive symptoms [3,4,5,6,7,8,9,10,11].
As with many psychiatric disorders, in depression, multiple neurotransmitter pathways, endocrine systems, and brain regions are involved. The endocannabinoid system (ECS) and the kynurenine pathway (KP) have long been strongly implicated in this disorder. Both systems contribute to the neuroinflammatory and serotonin hypothesis of depression, which will be discussed later on. In fact, there is a growing body of evidence showing potential common points or clear evidence for the interaction of the ECS and the KP. These were discussed previously by our group [12,13,14] among others [15,16,17,18]. This review will further support the link between the ECS and KP in the aspect of depression by summarizing the data of pro-inflammatory cytokines, which can be regulated by exogenous cannabinoids and at the same time which can regulate the KP. Additionally, pro-inflammatory cytokines, the ECS and KP in general, and regarding their role in depression will be also briefly discussed. Reviewing such data will allow a better understanding of the effect of cannabinoids on the neuroinflammatory component of depression.
2. Background
2.1. Pro-Inflammatory Cytokines
Pro-inflammatory cytokines are small signaling proteins which up-regulate during inflammation, as they are crucial for initiating and promoting inflammatory responses to diseases [19,20]. They are predominantly produced by macrophages, but astrocytes, microglia, and neurons can also generate them in the brain [21]. Most notable pro-inflammatory cytokines are the interleukins (IL), like IL-1, 6, 8, 12, or 18, interferons (IFN), such as IFNγ and tumor necrosis factors (TNF), like TNFα [19]. They are able to freely pass through the blood–brain barrier (BBB), via multiple mechanisms such as passive diffusion through the leaky regions of the BBB, active transport, or via nerve fibers such as the vagus or trigeminal nerves [21]. They bind to cytokine receptors which can be divided into class I and class II based on their structural differences [22]. However, both classes uniformly activate the Janus kinase-signal transducers and activators of the transcription (JAK-STAT) pathway [22]. There is increasing evidence demonstrating that pro-inflammatory cytokines have a significant role in certain neurological and psychiatric disorders. For instance, in patients with schizophrenia, Alzheimer’s, or depression, pro-inflammatory cytokine levels are elevated or dysregulated [23]. In this review, depression will be discussed in this regard.
2.2. The Endocannabinoid System
The ECS has a crucial role in depression, confirmed by behavioral, anatomical, electrophysiological, and genetic evidence reviewed thoroughly elsewhere [24,25,26,27]. The ECS includes endogenous cannabinoids or endocannabinoids [28,29,30], the enzymes which synthesize and degrade endocannabinoids [31], and finally cannabinoid receptors, namely type 1 and 2 (CB1R and CB2R) which mediate the effects of endogenous or exogenous cannabinoids [32,33,34,35]. Both CBRs belong to the G-protein coupled receptor (GPCR) family generally coupling to Gαi type G-proteins. Accordingly, they inhibit adenylyl cyclase activity, leading to the presynaptic release inhibition of neurotransmitters such as γ-aminobutyric acid (GABA), dopamine, or acetylcholine [29,36,37,38]. On the other hand, there are data demonstrating other signaling pathways activated by CBRs, involving cell and G-protein type specificity or G-protein independent mechanisms [32].
The CB1R is the most abundant GPCR in the human brain, with higher expression levels compared to other GPCRs. Apart from the central nervous system (CNS), CB1Rs are found in several peripheral tissues, such as adipocytes, gastrointestinal tract, or the reproductive system [39]. Thus, CB1Rs are responsible for multiple physiological processes such as mood, appetite, food intake, thermoregulation, cognition, and memory [29,38,39,40,41]. CB2Rs are expressed mainly in cells of the immune system in the periphery [39], but they can be found in the CNS as well, for instance, in the brainstem or cerebellum [42]. CB2Rs have a significant role in the maintenance of homeostasis, analgesia, controlling cell proliferation, differentiation, and survival of neuronal and non-neuronal cells [39,43,44].
Cannabinoid receptors—apart from endocannabinoids—are also activated by natural (or plant-derived), semi, or fully synthetic exogenous cannabinoids. Among the plant-derived or phytocannabinoids, ∆9-tetrahydrocannabinol (THC), the major psychoactive component of cannabis, and the non-psychoactive cannabidiol (CBD) are the most studied and well-known. The most relevant structural classes of synthetic cannabinoids (SC) are aminoalkylindoles (e.g., WIN 55212-2), naphtholylindoles (e.g., JWH-018), and cyclohexylphenols (e.g., CP 55940) [45,46]. Together with the CBR antagonists/inverse agonist diaryl-pyrazole derivatives, such as rimonabant [47], SCs significantly contributed to the pharmacological mapping of the ECS. Since SCs greatly mimic the effects of cannabis, aminoalkylindoles, cyclohexylphenols, and naphtholylindoles especially, are the most common SCs found in the K2/Spice products, which are the most widely abused class of drugs nowadays [11,48,49]. Indole, indazole carboxamides structured SCs (e.g., AB-PICA and AB-PINACA, respectively) were joined to this class very recently [46].
As mentioned previously, there are numerous preclinical and clinical studies pointing out that cannabis/THC and SCs use significantly contributes to the development of depression [3,6,8,50,51,52,53]. In fact, a recent study showed that SC users displayed a higher Beck Depression Inventory score [54] compared to natural cannabis users, with similar socio-demographic characteristics [51]. Another study pointed out a similar outcome, where SC use was associated with increased mental health symptomatology—including depression—compared to natural cannabinoid use [10]. These findings can be explained by the higher CB1R affinity and agonist potency of the SCs compared to THC [52,55].
2.3. The Kynurenine Pathway
Tryptophan (Trp) is an essential amino acid, which is pivotal in the brain and in mammalian cells, and is mainly metabolized via the KP (Figure 1). Kynurenic acid (KYNA) is one of the most studied and clinically relevant metabolite of the KP. KYNA is an endogenous glutamate receptor antagonist, which has neuroprotective effects and is produced by kynurenine aminotransferases (KAT)s from l-kynurenine (L-KYN) mostly in astrocytes [56,57,58]. L-KYN is formed by formamidase enzyme from N-formyl-l-kynurenine, which is created from l-Trp by two enzymes, namely the tryptophan 2,3-dioxygenase (TDO) and the indoleamine 2,3-dioxygenase 1 and 2 (IDO1 and IDO2). It is well known, that l-KYN can transform not just to KYNA, but is also able to convert into anthranilic acid by kynureninase and to 3-hydroxykynurenine by kynurenine 3-monooxygenase (KMO). Anthranilic acid can be further converted to 3-hydroxyanthranilic acid by 3-hydroxyanthranilic acid hydroxylase. In addition to this, 3-hydroxykynurenine can also convert to 3-hydroxyanthranilic acid by kynureninase enzyme. Besides that, 3-hydroxykynurenine can modify to xanthurenic acid, as well. Additionally, 3-hydroxyanthranillic acid further transforms to quinolinic acid (QUIN) by 3-hydroxyanthranillic acid 3,4-dioxygenase. In the end of the KP, QUIN is degraded to nicotinamide adenine dinucleotide by quinolinic acid phosphoribosyltransferase. Opposite to KYNA, QUIN is an endogenous glutamate receptor agonist produced by microglia [59] and it can cause lipid peroxidation [60] and has a relevant role in the neurodegenerative process [61,62].
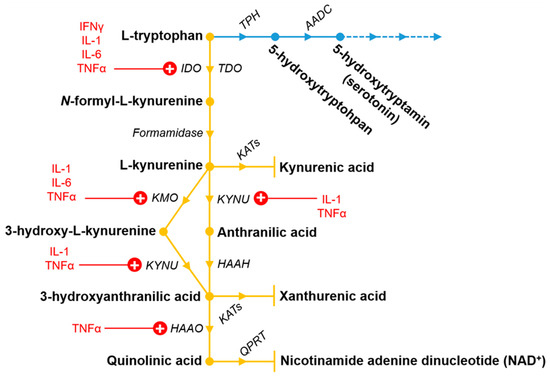
Figure 1.
The tryptophan metabolic pathway including the kynurenine (yellow) and partly the serotonin pathway (blue). Pro-inflammatory cytokines discussed in this review which upregulate (highlighted by “+”) the different enzymes are also indicated in red. For further information, see Section 4 and Table 2. Metabolites and enzymes of the pathways are shown in bold and italic, respectively. The dashed lines in the serotonin pathway indicates the further continuation of the pathway, which is not discussed here. Abbreviations: AADC: aromatic acid decarboxylase enzymes; HAAH: 3-hydroxyanthranilic acid hydroxylase; HAAO: 3-hydroxyanthranillic acid 3,4-dioxygenase; IDO: indoleamine 2,3-dioxygenase; KATs: kynurenine aminotransferases; KMO: kynurenine 3-monooxygenase; KYNU: kynureninase; QPRT: quinolinic acid phosphoribosyltransferase; TDO: tryptophan 2,3-dioxygenase; TPH: tryptophan hydroxylase.
2.4. The Serotonin and Inflammatory Hypothesis of Depression: Possible Links between ECS and KP in Depression?
The serotonin hypothesis was introduced more than 50 years ago as a possible pathological background mechanism for depression [63]. The hypothesis refers to a dysregulated serotoninergic system, implicating reduced levels of serotonin, serotonin transporters and/or receptors in patients with depression [64,65,66,67,68,69,70]. It has been long since described that reduced serotonin levels are due to Trp depletion [71]. Serotonin or 5-hydroxytryptamine is metabolized from Trp through 5-hydroxytryptophan catabolized by the tryptophan hydroxylase and aromatic acid decarboxylase enzymes [71] (Figure 1). However, only a small fraction of the Trp pool is converted to serotonin, the vast majority (~95%) is metabolized via KP, as discussed in the previous section. Thus, even a small change in the activity of the KP can have a significant impact on the Trp pool in the brain [72,73]. Indeed, there is numerous clinical evidence showing that there is an imbalance in the metabolism of KP in depression. The amount of Trp, l-KYN, and KYNA, for instance, is decreased in the serum and plasma of patient with depression, whereas QUIN is increased. These data have been reviewed in detail previously [74,75,76,77,78,79,80]. There are also several genetic mutations in the KP, which are connected to depression. Some polymorphisms of the IDO1, 2 and KMO encoded genes are identified in patients with depression [81]. On the other hand, data on increased levels of KP metabolites in blood serum and CSF in individuals with depressive disorders have been inconsistent [82,83,84,85]. It has been also proposed that changes in enzymes and metabolites of the KP are not necessarily parallel to events in the brain [86]. These data also clearly show the complexity of depression and that the serotonin hypothesis is not the only background mechanism responsible for this psychiatric disorder.
There are multiple studies showing that depression also consists of an inflammatory component centering not just in the brain but throughout the body. One of the main indications of such mechanism is the significant enhancement of circulating pro-inflammatory cytokines in animal models of depression and also in patients with depression, which can be reversed by antidepressants. These findings have been previously reviewed extensively [21,87,88,89,90,91,92]. It is well known that enzymes of the KP, especially IDO, can be activated by pro-inflammatory cytokines, which may lead to Trp depletion and possibly depression as described above. More interestingly, there are numerous data demonstrating that exogenous cannabinoids can enhance the levels of pro-inflammatory cytokines, which may over activate the KP, potentially leading to depression. These findings will be discussed later on. Therefore, there is a potential link between cannabinoids and the KP in depression, where exogenous cannabinoids potentially induce inflammation by increasing pro-inflammatory cytokines. Such effect then enhances the activity of the KP leading to Trp depletion and reduced levels of serotonin, which eventually may contribute to depression (Figure 2). This link might be a possible explanation for depressive episodes induced by natural and synthetic cannabinoids misuse. The following sections will discuss those exogenous cannabinoids which are known to increase pro-inflammatory cytokines. Additionally, such cytokines which have been associated with the activation of the KP in neuroinflammation and/or depression will be further reviewed accordingly.
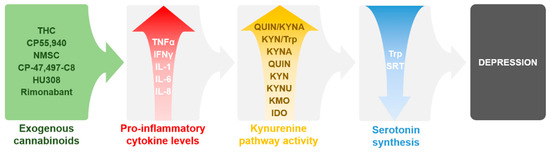
Figure 2.
Summary of the reviewed data and their proposed connections between exogenous cannabinoids, pro-inflammatory cytokines, KP, and depression as discussed in the sections below and in Table 1 and Table 2. Abbreviations: IDO: indolamine 2,3-dioxygenase; IFNγ: interferon γ; IL: interleukin; KMO: kynurenine 3-monooxygenase, KYN: l-kynurenine, KYNA: kynurenic acid; QUIN: quinolinic acid; THC: ∆9-tetrahydrocannabinol; TNFα: tumor necrosis factor α; Trp: tryptophan; SRT: serotonin.
3. Cannabinoids That Enhance Pro-Inflammatory Cytokine Levels
Cannabinoids, endogenous, synthetic, and natural types have been generally associated with anti-neuroinflammation by downregulating pro-inflammatory and/or upregulating anti-inflammatory cytokines typically through CB2Rs [93,94,95]. However, there is growing evidence demonstrating that natural and synthetic cannabinoids can indeed upregulate pro-inflammatory cytokines and thus possibly induce neuroinflammation and/or depression. This section will review these data (see Table 1). There is also substantial evidence that cytokines can induce mood alterations by regulating cannabinoid receptors [96,97,98,99], however this is out of the scope of this current review.
3.1. Natural Cannabinoids (Cannabis, THC, and CBD)
THC and CBD have been long known to regulate cytokine levels in a concentration dependent manner. In an earlier study in human peripheral blood mononuclear cells, THC and CBD in concentrations comparable to plasma levels prior to smoking marijuana (10–100 ng/mL), increased the concentration of IFNγ, while in higher concentrations (5–20 μg/mL), fully blocked the synthesis and/or release of this cytokine [100]. Another study also pointed out the biphasic effect of THC on cytokine regulation in mononuclear cells: TNF-α and IL-6 synthesis was inhibited by 3 nM THC but stimulated by 3 μM, as was with IFNγ synthesis [101]. Other studies also showed that the biphasic effect of THC on pro-inflammatory cytokines seems to be not only dependent on concentration, but also whether the experimental animals are naïve or neuroinflammation was induced [94,95,102,103,104,105]. In eosinophilic leukemia cell lines, both THC and CBD significantly increased IL-8 production, while in human T-lymphotropic virus type 1 (HTVL-1) positive B cell lines, only THC increased IL-8 levels [106]. Cutando and co-workers showed that subchronic administration of THC to mice activated cerebellar microglia and increased the expression of IL-1β and TNFα genes [107]. The neuroinflammation induced by THC was reversed by inhibiting IL-1β receptor signaling [107]. It is worth noting that CBD has a peculiar pharmacological profile which differs from THC and other natural and synthetic cannabinoids. In vitro and in vivo studies have indicated that CBD may act as a negative allosteric modulator of CB1R and an agonist of CB2R, transient receptor potential vanilloid 1 (TRPV1), 5-hydroxytryptophan1A receptors, and peroxisome proliferator-activated receptors γ (PPARy) [108]. Such multi-targeted action can help explain a prevailing anti-inflammatory action of CBD in vivo and in vitro, as reviewed elsewhere [109,110]. Briefly, CBD reduces stress and LPS-stimulated release of pro-inflammatory cytokines [109]. This anti-inflammatory effect could counteract THC-induced inflammation, thus explaining the beneficial profile of CBD in attenuating some detrimental effects of THC and in treatment conditions associated with drug abuse and dependence [111,112].
It is widely accepted that adolescence is a vulnerable period in terms of THC exposure, which can later result in psychiatric disorders in adulthood [50]. Additionally, multiple studies associated this with neuroinflammation, in particular with regulating cytokine expression. Moretti and co-workers showed that IL-1β and TNFα gene and protein expression increased in peripheral macrophages following chronic THC exposure in adolescent mice. Such was not the case when adult mice were treated chronically [113]. In fact, the opposite was observed if the same cytokines were analyzed right after the final THC treatment in both adolescent and adult animals [113]. Later on, the same findings were also confirmed in the hippocampus and hypothalamus by the same group [114]. Another study investigated the chronic effect of THC consumption in adolescent female rats. Here, the THC treatment enhanced expression levels of TNFα in microglia of the prefrontal cortex which was associated with depression-like phenotype [115].
Cannabis use disorder has its own set of definitions for diagnosis as it has been included in the latest edition of Diagnostic and Statistical Manual of Mental Disorders (DSM-5) [116]. It has been recently demonstrated that patients with cannabis use disorder have increased serum levels of IL-1β, IL-6, IL-8, and TNF-α levels [117]. In another study, they compared physically active chronic cannabis users (at least once per week for the past 6 months) and non-users in terms of the presence of depression and immune health indicated partly by IL-6 [118]. However, they found no difference between the two groups in IL-6 serum levels.
3.2. Semi- and Fully Synthetic Cannabinoids
Data regarding synthetic cannabinoids and cytokine regulation are relatively recent, but limited. CP55940 is functionally and structurally analogue to JWH-018 and to CP47497 which is a frequent component of “K2/Spice” synthetic cannabinoid blends [119]. In a study involving promyelocytic cells HL-60 transfected with CB2R, CP55940 increased TNFα mRNA after 1 h and protein levels after 24 h [120]. Both effects were CB2R mediated [120]. Very recently, Zawatsky and co-workers have shown that oropharyngeal administration of the synthetic cannabinoid CP55940 to mice significantly increased the mRNA levels of CB1Rs and induced the expression levels of IL-1β, IL-6, and TNFα in the lung [119]. In another study, they investigated a representative member of cyclohexylphenols of SCs which can bind to both CBRs, namely CP-47497-C8 (cannabicyclohexanol). Cannabicyclohexanol was also found in “Spice” in Germany and Japan [4,121] and was described to increase IL-6 and TNFα levels in peripheral blood mononuclear cells [122]. A CB2R selective synthetic cannabinoid agonist, HU308 in human primary leukocytes, was shown to induce the secretion of IL-6 via GαS coupled signaling [43]. The semi-synthetic CBD derivative 2-(methylsulfonamido)ethyl cannabidiolate (NMSC) enhanced IL-1β and IL-6 mRNA levels in RAW264.7 macrophages upon IL-17 stimulation, but only in higher concentration (10 µmol/L) [123]. In lower concentration (5 µmol/L), it showed the opposite effect.
An interesting study was conducted with the CB1R selective antagonist/inverse agonist rimonabant, which was withdrawn from the market due to its adverse psychiatric side-effects, including depression, anxiety, and suicidal ideation after long-term usage [124]. Such clinical data were strengthened by pre-clinical in vitro results. Namely, in rats which showed depressive-like phenotype, long-term rimonabant treatment increased the level of IL-6 and TNFα in the medial prefrontal cortex and in the hippocampus, respectively [125]. In a very recent study, they investigated the level of inflammation apart from oxidative stress and DNA damage in 40 synthetic cannabinoid (the exact compounds were not determined) addicts and they found that IL-1β, IL-6, and TNF-α serum cytokine levels were significantly higher compared to the healthy groups [126].

Table 1.
Summary of cannabinoids known to increase pro-inflammatory cytokines.
Table 1.
Summary of cannabinoids known to increase pro-inflammatory cytokines.
| Cannabinoid | Cytokine | Studied Sample | Ref. |
|---|---|---|---|
| THC | IFNγ | PMBC | [100,101] |
| TNFα | PMBC | [101] | |
| Adult mouse peripheral macrophage | [113] | ||
| Adult mouse hippocampus and hypothalamus | [114] | ||
| Female adol. rat microglia PFC | [115] | ||
| IL-1 | Microglia | [107] | |
| Adult mouse peripheral macrophage | [113] | ||
| Adult mouse hippocampus and hypothalamus | [114] | ||
| IL-6 | PMBC | [101] | |
| IL-8 | Eosinophilic leukemia cell line and HTLV-1 positive B cell line | [106] | |
| CBD | IFNγ | PMBC | [101] |
| IL-8 | Eosinophilic leukemia cell line | [106] | |
| Cannabis | IL-1, IL-6, IL-8 | Serum from patients with CUD | [117] |
| CP55940 | TNFα | HL-60 transfected with CB2R; mouse lung | [119,120] |
| IL-1 | Mouse lung | [119] | |
| IL-6 | Mouse lung | [119] | |
| NMSC | IL-1β, IL-6 | RAW264.7 macrophage | [123] |
| CP-47497-C8 | TNFα, IL-6 | PMBC | [122] |
| HU308 | IL-6 | Human primary leukocytes | [43] |
| Rimonabant | TNFα | rat hippocampus | [125] |
| IL-6 | rat mPFC | [125] |
CUD: cannabis used disorder; HTVL-1: human T-lymphotropic virus type 1; PFC: prefrontal cortex; PMBC: peripheral mononuclear cells; mPFC: medial prefrontal cortex; NMSC: 2-(methylsulfonamido)ethyl cannabidiolate.
4. Pro-Inflammatory Cytokines Parallelly Up-Regulated with the KP in Neuroinflammation and/or Depression
As we saw in the previous section, there are multiple studies pointing out the upregulation of IFNγ, IL-1, IL-6, IL-8, and TNFα pro-inflammatory cytokines via cannabinoid induction. This section will review the data regarding the effect of the above- mentioned cytokines on the regulation of the KP enzymes and their metabolite production. There are other reviews describing the relationship between cytokines and the KP in different disorders [127,128,129,130]. However, this section is the first to thoroughly review these data in the aspect of neuroinflammation or depression. Table 2 summarizes the data discussed below. Important to note that in contrast to multiple reports, a previous study showed reduced KP metabolism and pro-inflammatory cytokine levels in post mortem ventrolateral prefrontal cortex tissues from individuals with depressive illness [131]. The study also discussed that such unexpected result might be due to the different specific brain region investigated and/or the distinct diagnosis classification of the depressed samples used, which might have influenced the overall results. Finally, the study also pointed out that the regulation of KP in the human brain might be brain-region specific in depression.
4.1. IFN-γ
IFN-γ has long been known for regulating IDO activity [132,133], which has been discussed extensively in a previous review in 2014 [127]. Since then, additional research has been done in this area. The synergistic effect between IFN-γ and IL-1 is well-known in regulating IDO enzyme activity and transcription [134,135,136]. Moreover, in THP-1 human monocytic cell lines, Fujigaki and co-workers also demonstrated that LPS-induced IDO enzyme activity was upregulated when IFN-γ together with IL-1β, IL-6, and TNFα were present [137]. However, galectins, which also play an important role in neuroinflammation, and corticosteroids have also been shown to enhance the effect of IFN-γ in controlling IDO expression. In the mouse hippocampus, it has been shown that galectin-9, dexamethasone, corticosterone, and aldosterone interacted with IFN-γ to further enhance the mRNA expression of different IDO variants [138,139]. In a chronic social defeat mouse model, which models the anhedonic and social-avoidance aspect of depression, IFN-γ plasma levels increased together with KYN, 3-HK [140]. On the other hand, KYNA plasma levels were also enhanced, which seem to be in contrast with the elevated QUIN/KYNA ratio attributed to depression. The study did not further elaborate on this result. Another animal model of depression, the chronic mild stress procedure significantly increased IFNγ and IDO mRNA and decreased KAT II mRNA in the rat cortex [141]. The latter case may project ahead the increased QUIN/KYNA ratio observed in depression, since due to reduced KAT II availability, KYN conversion is more likely to be directed towards QUIN rather than KYNA production.
4.2. IL-1
Apart from IFN-γ, IL-1 is the most significant pro-inflammatory cytokine to regulate IDO. IL-1 alone transcriptionally activates the IDO gene in primary macrophages and is able to enhance the activity of the enzyme but only in the IFN-γ pretreated THP1 monocytic leukemia cell line [142].
Fractalkine receptor (CX3CR1) deficient mice have been demonstrated to display depressive-like behavior following LPS treatment [143]. In such mice, increased microglial mRNA expression of IL-1β, IDO, and KMO after LPS treatment was observed [143]. In a Bacille Calmette Guérin (BCG) depressive-like behavior mouse model, both IL-1β and KMO but not IDO-1 and -2 mRNA were upregulated in microglia [144]. Upon LPS stimulation, mRNA expression of IL-1β was dose-dependently increased parallelly with IDO-1 and KMO in murine microglia [145]. Additionally, KMO deletion prevented the LPS-induction of IL-1β.
Laumet and co-workers demonstrated the involvement of IL-1β in nerve injury-induced depression associated with enhanced KMO mRNA brain expression and activity in mice brain [146]. Additionally, functional IDO-positive dendritic cells produced significantly more IL-1β than IDO-negative cells upon CD40L stimulation [147]. IL-1β treatment in human hippocampal progenitor cells induced the transcription of IDO, KMO, and KYNU, which resulted in an increase in KYN production [148]. In the same study, inhibiting the KMO enzyme reversed the reduction of neurogenesis in human hippocampal progenitor cells induced by IL-1β. In another study involving the hippocampus, IDO1 mRNA expression was also enhanced by the upregulation of IL-1β production in the hippocampus of rats with coexisting chronic temporal lobe epilepsy and depressive behavior [149]. The two forms of the alarmin protein, high mobility group box-1 (HMGB1)—the fully reduced (fr-HMGB1) and the disulfide (ds-HMGB1) form—are known to induce depressive-like behavior [150]. Recently, it has been shown in mouse hippocampal tissues ds-HMGB1 directly activated IDO, KMO, and KYNU in parallel with IL-1β upregulation [151]. With fr-HMGB1, the same observations were made following H2O2 treatment. In the study, both forms of HMGB1 induced depressive-like behavior.
4.3. IL-6
The correlation between IL-6 and KP metabolites and enzymes has been long known. For instance, lower Trp levels in patients with depression are known to be inversely correlated to serum concentrations of IL-6 [152]. There is evidence that depressive and anxiety symptoms in the early puerperium in fare causally related to an increased catabolism of Trp into KYN, which may be associated with increased plasma levels of IL-6 [153]. Schwieler showed that in patients with unipolar treatment-resistant depression, IL-6 plasma levels and the QUIN/KYNA ratio in the plasma significantly increased compared to healthy volunteers [154]. Kruse and co-workers demonstrated that in a human experimental model of endotoxin-induced depressed mood, there was a positive correlation in plasma concentrations of KYN and QUIN and IL-6. However, changes in the KP metabolites did not mediate the correlation between cytokines and depressed mood [80]. In a recent study, IL-6 and QUIN plasma levels were positively correlated in women with peripartum onset depression (PPD) [155]. In another recent study with frail patients, it was found that the KYN/Trp ratio and KYN levels were strongly correlated with IL-6 plasma levels [156]. The authors concluded that these results are in accordance with the serotonin-KYN hypothesis of depression and also may explain the high prevalence of depression among individuals with frailty status [156].
IL-6 may also contribute to cortisol’s induction of TDO, as increased IL-6 in depression are implicated in elevated hypothalamic–pituitary–adrenal activity and cortisol levels, which in turn activates TDO [157,158]. Bay-Richter and colleagues found that cerebrospinal fluid levels of QUIN and KYNA increased and decreased, respectively, in suicide attempters, which remained over time and also high IL-6 cerebrospinal fluid levels correlated with more severe suicidal symptoms [159].
In another study with microglia, LPS stimulation dose-dependently increased the mRNA expression of IL-6 and parallelly of IDO-1 and KMO [145]. There is also multiple evidence for IL-6 regulating KP enzymes in the brain. Kim and co-workers have shown that intra-hippocampal administration of IL-6 in rats induces IDO1 expression through the JAK/STAT pathway [160]. Xie and co-workers demonstrated in rats with coexisting chronic temporal lobe epilepsy and depressive behavior that the upregulation of IL-6 production in the hippocampus enhanced IDO1 mRNA expression too in the same brain area [149]. In rats with ovariectomy-induced depressive-like behavior, showed parallelly elevated IL-6 and IDO protein levels in the hippocampus [161]. In another model, the enhancement of LPS induced IDO and KMO mRNA expression was accompanied by a significant increase in IL-6 expression in the rat hippocampus and cortex and in cultured glial cells [162].
In the Netherlands Study of Depression and Anxiety (NESDA), a cohort consisting almost 3000 participants, no indications were found in KYN/Trp ratio for mediating the relationship between changes in IL-6 levels and depressive symptoms [83].
4.4. IL-8
Maes and colleagues demonstrated that hepatitis C patients who received IFNα treatment showed an increase in depressive symptoms and KYN/Trp quotient along with elevations in IL-8 plasma levels [163]. In the previously mentioned study, where they investigated the relation between KYNUs, immune activity and depressive and anxiety symptoms in the early puerperium, they observed enhanced IL-8 plasma levels parallelly with increased KYN/Trp quotient [153].
4.5. TNFα
TNFα and IDO serum levels parallelly increased in major depressive disorder (MDD) patients, which was reduced by post-treatment [164]. Chronic social defeat depression mouse model led to increased plasma levels of TNF-α in parallel with KYN, 3-HK, and KYNA as seen with IFNγ [140]. Similar to IL-6, there was also a positive correlation in plasma concentrations of KYN and QUIN and TNFα in the human experimental model of endotoxin-induced depressed mood. However, changes in the KP metabolites did not mediate the correlation between cytokines and the depressed mood [80]. TNFα levels correlated positively with QUIN plasma levels in women with PPD [155]. Haroon and co-workers found a correlation among peripheral and central KP metabolites and inflammation in depression in a study involving 72 unmedicated depressed patients. Accordingly, plasma TNFα was robustly associated with plasma KYN and KYN/Trp ratio levels, which was in turn significantly correlated with CSF KYN, KYNA, and QUIN [165]. Additionally high TNFα-kynurenine/tryptophan subjects showed enhanced depression severity, anhedonia, and treatment nonresponse [165]. In the study involving frailty patients, they also found that the KYN/Trp ratio and KYN levels were strongly correlated with TNFα and TNFαR1 levels too, while Trp and KYNA alone were also strongly correlated with TNFαR1 levels [156].
O’Connor and co-workers demonstrated that in the BCG mice model, TNFα, IDO, and HAO mRNA significantly increased in the brain [166]. The same group also showed that IFNγ and TNFα synergistically induce IDO in primary microglia cells and they are both necessary for the induction of IDO and depressive-like behavior in mice after BCG infection [167]. There is also further evidence that TNFα together with IFNγ can transcriptionally activate IDO [136,168,169].
The enhancement of LPS induced IDO and KMO expression was also associated with increased TNFα expression in the rat hippocampus and cortex and in cultured glial cells [162]. In murine microglia upon LPS stimulation, mRNA expression of TNF-α was dose-dependently increased together with IDO1 and KMO mRNA, and KMO deletion eliminated the LPS-induced TNFα elevation [145]. The two forms of the high mobility group box-1 (HMGB1) protein (fr-HMGB1 and ds-HMGB1) mentioned regarding IL-1β cytokine, are also known to upregulate TNFα besides inducing depressive-like behavior [150]. Additionally, alongside IL-β, TNFα was also upregulated in mice hippocampal tissues in parallel with IDO, KMO, and KYNU, which were activated by both forms of HMGB1 [151].
Table 2.
Summarizing the data regarding upregulated pro-inflammatory cytokines associated with altered KP enzymes or metabolites.
Table 2.
Summarizing the data regarding upregulated pro-inflammatory cytokines associated with altered KP enzymes or metabolites.
| Cytokine | KP Enzyme or Metabolite | Studied Sample | Comment | Ref. |
|---|---|---|---|---|
| IFNγ | IDO mRNA ↑ | mouse hippocampus | galectin-9 synergism | [138] |
| mouse hippocampus | dexamethasone, corticosterone and aldosterone synergism | [139] | ||
| rat cortex | CMS model | [141] | ||
| KAT II mRNA ↓ | rat cortex | CMS model | [141] | |
| KYN, 3-HK, KYNA ↑ | mouse plasma | CSD model | [140] | |
| IL-1 | IDO mRNA ↑ | primary macrophage | [142] | |
| IDO activity ↑ | THP1 monocytic leukemia cell line | IFNγ pretreatment | [142] | |
| IDO, KMO mRNA ↑ | CX3CR1 K.O. mouse microglia | [143] | ||
| KMO mRNA ↑ | mouse microglia | BCG model | [144] | |
| IDO, KMO mRNA ↑ | murine microglia | LPS-induction | [145] | |
| KMO mRNA ↑ | mouse brain | nerve injury-induced depression | [146] | |
| IDO, KMO, KYNU mRNA, KYN ↑ | human hippocampal progenitor cells | [148] | ||
| IDO1 mRNA ↑ | rat hippocampus | coexisting chronic temporal lobe epilepsy and depressive behavior | [149] | |
| IDO, KMO and KYNU activity ↑ | mouse hippocampus | HMGB1 induced depressive like behavior model | [151] | |
| IL-6 | Trp ↓ | human serum | in patients with depression | [152] |
| KYN ↑ | female human serum | in early puerperium associated with anxiety and depression | [153] | |
| QUIN/KYNA ratio ↑ | human plasma | in patients with unipolar treatment-resistant depression | [154] | |
| KYN and QUIN ↑ | human plasma | did not mediate the correlation between cytokines and depressed mood | [80] | |
| QUIN ↑ | female human plasma | women with PPD | [155] | |
| KYN/Trp ratio, KYN ↑ | plasma from frailty patients | may explain high prevalence depression in frailty patients | [156] | |
| QUIN ↑ KYNA ↓ | human CSF | in suicide attempters | [159] | |
| IDO1, KMO mRNA ↑ | murine microglia | following LPS-stimulation | [145] | |
| IDO1 protein ↑ | rat hippocampus | through JAK/STAT pathway | [160] | |
| IDO1 mRNA ↑ | rat hippocampus | coexisting chronic temporal lobe epilepsy and depressive behavior | [149] | |
| IDO1 protein ↑ | rat hippocampus | ovariectomy-induced depression model | [161] | |
| IDO, KMO mRNA ↑ | rat hpc., ctx., and cultured glia cells | [162] | ||
| IL-8 | KYN/Trp quotient ↑ | human plasma | IFNα-induced depressive symptoms | [163] |
| female human serum | in early puerperium associated with anxiety and depression | [153] | ||
| TNFα | IDO ↑ | human serum | in MDD patients | [164] |
| KYN, 3-HK, KYNA ↑ | mouse plasma | CSD mouse model | [140] | |
| KYN and QUIN ↑ | human plasma | did not mediate the correlation between cytokines and depressed mood | [80] | |
| QUIN ↑ | female human plasma | women with PPD | [155] | |
| KYN, KYN/Trp ratio ↑ | plasma | associated with enhanced depression, anhedonia, and treatment nonresponse | [165] | |
| KYN, KYNA, QUIN ↑ | CSF | in unmedicated depressed patients | [165] | |
| KYN/Trp ratio, KYN ↑ | plasma from frailty patients | may explain high prevalence depression in frailty patients | [156] | |
| IDO, HAAO mRNA ↑ | mouse brain | BCG model | [166] | |
| IDO activity ↑ | mouse microglia cells | BCG model | [167] | |
| IDO, KMO mRNA ↑ | rat hpc., ctx., and cultured glia cells | [162] | ||
| IDO1, KMO mRNA ↑ | murine microglia | following LPS-stimulation | [145] | |
| IDO, KMO and KYNU activity ↑ | mouse hippocampus | HMGB1 induced depressive like behavior model | [151] |
↑: increase; ↓: decrease; BCG: Bacille Calmette Guérin mice model of depression; CMS: chronic mild stress model; ctx.: cortex; CSD: chronic social defeat model; HMGB1: high mobility group box-1 protein; hpc.: hippocampus; MDD: major depressive disorder; PPD: peripartum onset depression.
5. Summary and Conclusions
This paper summarized pre-clinical and clinical evidence on pro-inflammatory cytokines which are upregulated by natural and synthetic cannabinoids, thus might be contributing to the inflammatory component of depression induced by such compounds. Additionally, the manuscript further reviewed those cytokines which are parallelly upregulated with certain enzymes and metabolites of the KP, possibly leading to the over-activation of the KP. This over-activation may significantly contribute to the downregulated serotoninergic system attributed to depression.
Cannabis use has been increasing rapidly over the past few years, due to its legalization in a growing number of US states and other countries. SC consumption has also gained significant popularity over the years, however, in contrast to natural cannabinoids, for many SCs, the receptor preference, affinity, metabolic mechanisms, and pharmacodynamics are unknown [45,46]. Additionally, the discrepancies in the pro- and anti-inflammatory effects of THC and CBD are known and it is explained by the difference in the applied concentrations and model systems (naïve vs. inflammatory-induced) and by the complex pharmacological profile in case of CBD. The investigation on the molecular mechanisms by which cannabinoids could lead to increased inflammatory effects could potentially unravel important targets for controlling neuroinflammation associated with drug abuse and dependence and its emotional consequences. Both natural and synthetic cannabinoids significantly contribute to the development of depression based on multiple pre-clinical and clinical studies. The data reviewed here may reveal a possible link between the ECS and the KP and help to overview the connection between cannabinoids, inflammation, and KP in relation to the pathophysiology of depression. Although, there is no direct evidence so far that exogenous cannabinoids induce depression via inflammation-stimulated KP in one experimental system, the data gathered in this review clearly demonstrate its strong possibility. Nevertheless, reviewing such data may raise interest to study the inflammatory component of depression by pharmacological and/or genetic manipulation of either the ECS or the KP. Selective exogenous cannabinoids [170,171] and enzyme inhibitors of the KP as well as CBR and KP enzyme knock-out animals are available and widely used to study the function of the ECS and KP [172,173,174,175,176,177,178,179]. Applying these tools may reveal the response of each system to one another when manipulated in inflammatory-induced depression.
Author Contributions
Conceptualization, F.Z., S.J.; Writing—original draft preparation; F.Z., S.J., G.N.-G., S.D., E.S.; Writing—review and editing, F.Z., S.J., G.N.-G., S.D., E.S., L.V., C.T., S.B.; Visualization, F.Z.; Project administration, F.Z., G.N.-G., L.V.; Funding acquisition, G.N.-G., L.V. All authors have read and agreed to the published version of the manuscript.
Funding
G.N.-G. was supported by the ÚNKP-20-4 New National Excellence Program of the Ministry for Innovation and Technology from the source of the National Research, Development and Innovation Fund. S.J. is supported by the Aarhus University Research Foundation (AUFF starting grant) and FAPESP (17/24304-0). This research was supported by the EU-funded Hungarian grant EFOP-3.6.1-16-2016-00008 and by the TUDFO/47138-1/2019-ITM, GINOP 2.3.2-15-2016-00034.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- GBD 2017 Disease and Injury Incidence and Prevalence Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef]
- Volkow, N.D. The reality of comorbidity: Depression and drug abuse. Biol. Psychiatry 2004, 56, 714–717. [Google Scholar] [CrossRef]
- Seely, K.A.; Lapoint, J.; Moran, J.H.; Fattore, L. Spice drugs are more than harmless herbal blends: A review of the pharmacology and toxicology of synthetic cannabinoids. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2012, 39, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Auwärter, V.; Dresen, S.; Weinmann, W.; Müller, M.; Pütz, M.; Ferreirós, N. “Spice” and other herbal blends: Harmless incense or cannabinoid designer drugs? J. Mass Spectrom. 2009, 44, 832–837. [Google Scholar] [CrossRef] [PubMed]
- Stinson, F.S.; Ruan, W.J.; Pickering, R.; Grant, B.F. Cannabis use disorders in the USA: Prevalence, correlates and co-morbidity. Psychol. Med. 2006, 36, 1447–1460. [Google Scholar] [CrossRef] [PubMed]
- Dierker, L.; Selya, A.; Lanza, S.; Li, R.; Rose, J. Depression and marijuana use disorder symptoms among current marijuana users. Addict. Behav. 2018, 76, 161–168. [Google Scholar] [CrossRef]
- Armenian, P.; Darracq, M.; Gevorkyan, J.; Clark, S.; Kaye, B.; Brandehoff, N.P. Intoxication from the novel synthetic cannabinoids AB-PINACA and ADB-PINACA: A case series and review of the literature. Neuropharmacology 2018, 134, 82–91. [Google Scholar] [CrossRef]
- Lev-Ran, S.; Roerecke, M.; Le Foll, B.; George, T.P.; McKenzie, K.; Rehm, J. The association between cannabis use and depression: A systematic review and meta-analysis of longitudinal studies. Psychol. Med. 2014, 44, 797–810. [Google Scholar] [CrossRef]
- Richter, L.; Pugh, B.S.; Ball, S.A. Assessing the risk of marijuana use disorder among adolescents and adults who use marijuana. Am. J. Drug Alcohol Abus. 2017, 43, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Mensen, V.T.; Vreeker, A.; Nordgren, J.; Atkinson, A.; de la Torre, R.; Farré, M.; Ramaekers, J.G.; Brunt, T.M. Psychopathological symptoms associated with synthetic cannabinoid use: A comparison with natural cannabis. Psychopharmacology 2019, 236, 2677–2685. [Google Scholar] [CrossRef]
- Evren, C.; Bozkurt, M. Synthetic Cannabinoids: Crisis of The Decade. Dusunen Adam J. Psychiatry Neurol. Sci. 2013, 26, 1. [Google Scholar]
- Nagy-Grócz, G.; Zádor, F.; Dvorácskó, S.; Bohár, Z.; Benyhe, S.; Tömböly, C.; Párdutz, Á.; Vécsei, L. Interactions between the Kynurenine and the Endocannabinoid System with Special Emphasis on Migraine. Int. J. Mol. Sci. 2017, 18, 1617. [Google Scholar] [CrossRef] [PubMed]
- Zádor, F.; Nagy-Grócz, G.; Kekesi, G.; Dvorácskó, S.; Szűcs, E.; Tömböly, C.; Horvath, G.; Benyhe, S.; Vécsei, L. Kynurenines and the Endocannabinoid System in Schizophrenia: Common Points and Potential Interactions. Molecules 2019, 24, 3709. [Google Scholar] [CrossRef] [PubMed]
- Zádor, F.; Nagy-Grócz, G.; Dvorácskó, S.; Bohár, Z.; Cseh, E.K.; Zádori, D.; Párdutz, Á.; Szűcs, E.; Tömböly, C.; Borsodi, A.; et al. Long-term systemic administration of kynurenic acid brain region specifically elevates the abundance of functional CB1 receptors in rats. Neurochem. Int. 2020, 138, 104752. [Google Scholar] [CrossRef]
- Beggiato, S.; Borelli, A.C.; Tomasini, M.C.; Morgano, L.; Antonelli, T.; Tanganelli, S.; Cuomo, V.; Ferraro, L. Long-lasting alterations of hippocampal GABAergic neurotransmission in adult rats following perinatal Δ9-THC exposure. Neurobiol. Learn. Mem. 2017, 139, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Secci, M.E.; Mascia, P.; Sagheddu, C.; Beggiato, S.; Melis, M.; Borelli, A.C.; Tomasini, M.C.; Panlilio, L.V.; Schindler, C.W.; Tanda, G.; et al. Astrocytic Mechanisms Involving Kynurenic Acid Control Δ9-Tetrahydrocannabinol-Induced Increases in Glutamate Release in Brain Reward-Processing Areas. Mol. Neurobiol. 2018, 56, 3563–3575. [Google Scholar] [CrossRef] [PubMed]
- Beggiato, S.; Ieraci, A.; Tomasini, M.C.; Schwarcz, R.; Ferraro, L. Prenatal THC exposure raises kynurenic acid levels in the prefrontal cortex of adult rats. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2020, 100, 109883. [Google Scholar] [CrossRef] [PubMed]
- Colín-González, A.L.; Aguilera, G.; Santamaría, A. Cannabinoids: Glutamatergic Transmission and Kynurenines. Adv. Neurobiol. 2016, 12, 173–198. [Google Scholar]
- Dinarello, C.A. Proinflammatory Cytokines. Chest 2000, 118, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.-Q.; Yu, J. Inflammation: A mechanism of depression? Neurosci. Bull. 2014, 30, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Brooks, A.J.; Dehkhoda, F.; Kragelund, B.B. Cytokine Receptors; Springer: Cham, Switzerland, 2017; pp. 1–29. [Google Scholar]
- Kronfol, Z.; Remick, D.G. Cytokines and the Brain: Implications for Clinical Psychiatry. Am. J. Psychiatry 2000, 157, 683–694. [Google Scholar] [CrossRef]
- Huang, W.-J.; Chen, W.-W.; Zhang, X. Endocannabinoid system: Role in depression, reward and pain control (Review). Mol. Med. Rep. 2016, 14, 2899–2903. [Google Scholar] [CrossRef]
- Vinod, K.Y.; Hungund, B.L. Role of the endocannabinoid system in depression and suicide. Trends Pharmacol. Sci. 2006, 27, 539–545. [Google Scholar] [CrossRef]
- Patel, S.; Hillard, C.J. Role of endocannabinoid signaling in anxiety and depression. Curr. Top. Behav. Neurosci. 2009, 1, 347–371. [Google Scholar] [PubMed]
- Valverde, O.; Torrens, M. CB1 receptor-deficient mice as a model for depression. Neuroscience 2012, 204, 193–206. [Google Scholar] [CrossRef]
- Battista, N.; Di Tommaso, M.; Bari, M.; Maccarrone, M. The endocannabinoid system: An overview. Front. Behav. Neurosci. 2012, 6, 9. [Google Scholar] [CrossRef]
- Rodríguez de Fonseca, F.; Del Arco, I.; Bermudez-Silva, F.J.; Bilbao, A.; Cippitelli, A.; Navarro, M. The endocannabinoid system: Physiology and pharmacology. Alcohol Alcohol. 2005, 40, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Bisogno, T. Endogenous cannabinoids: Structure and metabolism. J. Neuroendocr. 2008, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Basavarajappa, B.S. Critical enzymes involved in endocannabinoid metabolism. Protein Pept. Lett. 2007, 14, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Busquets-Garcia, A.; Bains, J.; Marsicano, G. CB1 Receptor Signaling in the Brain: Extracting Specificity from Ubiquity. Neuropsychopharmacology 2018, 43, 4–20. [Google Scholar] [CrossRef]
- Howlett, A.C.; Abood, M.E. CB1 and CB2 Receptor Pharmacology. Adv. Pharmacol. 2017, 80, 169–206. [Google Scholar]
- Pertwee, R.G. The pharmacology of cannabinoid receptors and their ligands: An overview. Int. J. Obes. 2006, 30 (Suppl. 1), S13–S18. [Google Scholar] [CrossRef]
- Pertwee, R.G.; Howlett, A.C.; Abood, M.E.; Alexander, S.P.H.; Di Marzo, V.; Elphick, M.R.; Greasley, P.J.; Hansen, H.S.; Kunos, G.; Mackie, K.; et al. International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid Receptors and Their Ligands: Beyond CB1 and CB2. Pharmacol. Rev. 2010, 62, 588–631. [Google Scholar] [CrossRef]
- Demuth, D.G.; Molleman, A. Cannabinoid signalling. Life Sci. 2006, 78, 549–563. [Google Scholar] [CrossRef]
- Bidaut-Russell, M.; Devane, W.A.; Howlett, A.C. Cannabinoid receptors and modulation of cyclic AMP accumulation in the rat brain. J. Neurochem. 1990, 55, 21–26. [Google Scholar] [CrossRef]
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, W.A.; Felder, C.C.; Herkenham, M.; Mackie, K.; Martin, B.R.; et al. International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol. Rev. 2002, 54, 161–202. [Google Scholar] [CrossRef]
- Maccarrone, M.; Bab, I.; Bíró, T.; Cabral, G.A.; Dey, S.K.; Di Marzo, V.; Konje, J.C.; Kunos, G.; Mechoulam, R.; Pacher, P.; et al. Endocannabinoid signaling at the periphery: 50 years after THC. Trends Pharmacol. Sci. 2015, 36, 277–296. [Google Scholar] [CrossRef]
- Pertwee, R.G. Cannabinoid receptors and pain. Prog. Neurobiol. 2001, 63, 569–611. [Google Scholar] [CrossRef]
- Pagotto, U.; Marsicano, G.; Cota, D.; Lutz, B.; Pasquali, R. The emerging role of the endocannabinoid system in endocrine regulation and energy balance. Endocr. Rev. 2006, 27, 73–100. [Google Scholar] [CrossRef]
- Chen, D.; Gao, M.; Gao, F.; Su, Q.; Wu, J. Brain cannabinoid receptor 2: Expression, function and modulation. Acta Pharmacol. Sin. 2017, 38, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Saroz, Y.; Kho, D.T.; Glass, M.; Graham, E.S.; Grimsey, N.L. Cannabinoid Receptor 2 (CB2) Signals via G-alpha-s and Induces IL-6 and IL-10 Cytokine Secretion in Human Primary Leukocytes. bioRxiv 2019, 2, 414–428. [Google Scholar] [CrossRef]
- Atwood, B.K.; Mackie, K. CB2: A cannabinoid receptor with an identity crisis. Br. J. Pharmacol. 2010, 160, 467–479. [Google Scholar] [CrossRef]
- Walsh, K.B.; Andersen, H.K. Molecular Pharmacology of Synthetic Cannabinoids: Delineating CB1 Receptor-Mediated Cell Signaling. Int. J. Mol. Sci. 2020, 21, 6115. [Google Scholar] [CrossRef] [PubMed]
- Potts, A.J.; Cano, C.; Thomas, S.H.L.; Hill, S.L. Synthetic cannabinoid receptor agonists: Classification and nomenclature. Clin. Toxicol. 2020, 58, 82–98. [Google Scholar] [CrossRef]
- Muccioli, G.G.; Lambert, D.M. Current knowledge on the antagonists and inverse agonists of cannabinoid receptors. Curr. Med. Chem. 2005, 12, 1361–1394. [Google Scholar] [CrossRef]
- Fattore, L. Synthetic Cannabinoids—Further Evidence Supporting the Relationship Between Cannabinoids and Psychosis. Biol. Psychiatry 2016, 79, 539–548. [Google Scholar] [CrossRef]
- Brents, L.K.; Prather, P.L. The K2/Spice Phenomenon: Emergence, identification, legislation and metabolic characterization of synthetic cannabinoids in herbal incense products. Drug Metab. Rev. 2014, 46, 72. [Google Scholar] [CrossRef]
- Volkow, N.D.; Baler, R.D.; Compton, W.M.; Weiss, S.R.B. Adverse Health Effects of Marijuana Use. N. Engl. J. Med. 2014, 370, 2219–2227. [Google Scholar] [CrossRef]
- Serap Akdeniz, G.; Can Sait, S. Cannabinoid Use and Depression: Comparison of Natural and Synthetic Cannabinoids. Clin. Med. Rev. Case Rep. 2020, 7, 7. [Google Scholar] [CrossRef]
- Castaneto, M.S.; Gorelick, D.A.; Desrosiers, N.A.; Hartman, R.L.; Pirard, S.; Huestis, M.A. Synthetic cannabinoids: Epidemiology, pharmacodynamics, and clinical implications. Drug Alcohol Depend. 2014, 144, 12–41. [Google Scholar] [CrossRef]
- Escelsior, A.; Belvederi Murri, M.; Pietro, C.G.; Serafini, G.; Aguglia, A.; Zampogna, D.; Cattedra, S.; Nebbia, J.; Trabucco, A.; Prestia, D.; et al. Cannabinoid use and self-injurious behaviours: A systematic review and meta-analysis. J. Affect. Disord. 2021, 278, 85–98. [Google Scholar] [CrossRef]
- Sajatovic, M.; Chen, P.; Young, R.C. Rating Scales in Bipolar Disorder. Clin. Trial Des. Chall. Mood Disord. 2015, 105–136. [Google Scholar] [CrossRef]
- Martinotti, G.; Santacroce, R.; Papanti, D.; Elgharably, Y.; Prilutskaya, M.; Corazza, O. Synthetic Cannabinoids: Psychopharmacology, Clinical Aspects, Psychotic Onset. CNS Neurol. Disord. Drug Targets 2017, 16, 567–575. [Google Scholar] [CrossRef]
- Guillemin, G.J.; Kerr, S.J.; Smythe, G.A.; Smith, D.G.; Kapoor, V.; Armati, P.J.; Croitoru, J.; Brew, B.J. Kynurenine pathway metabolism in human astrocytes: A paradox for neuronal protection. J. Neurochem. 2001, 78, 842–853. [Google Scholar] [CrossRef] [PubMed]
- Guillemin, G.J.; Smith, D.G.; Kerr, S.J.; Smythe, G.A.; Kapoor, V.; Armati, P.J.; Brew, B.J. Characterisation of kynurenine pathway metabolism in human astrocytes and implications in neuropathogenesis. Redox Rep. 2000, 5, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Guillemin, G.J.; Cullen, K.M.; Lim, C.K.; Smythe, G.A.; Garner, B.; Kapoor, V.; Takikawa, O.; Brew, B.J. Characterization of the Kynurenine Pathway in Human Neurons. J. Neurosci. 2007, 27, 12884–12892. [Google Scholar] [CrossRef] [PubMed]
- Guillemin, G.J.; Smith, D.G.; Smythe, G.A.; Armati, P.J.; Brew, G.J. Expression of The Kynurenine Pathway Enzymes in Human Microglia and Macrophages. Adv. Exp. Med. Biol. 2003, 527, 105–112. [Google Scholar] [PubMed]
- Behan, W.M.H.; McDonald, M.; Darlington, L.G.; Stone, T.W. Oxidative stress as a mechanism for quinolinic acid-induced hippocampal damage: Protection by melatonin and deprenyl. Br. J. Pharmacol. 1999, 128, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Lugo-Huitrón, R.; Ugalde Muñiz, P.; Pineda, B.; Pedraza-Chaverrí, J.; Ríos, C.; Pérez-de la Cruz, V. Quinolinic acid: An endogenous neurotoxin with multiple targets. Oxid. Med. Cell. Longev. 2013, 2013, 104024. [Google Scholar] [CrossRef]
- Vécsei, L.; Szalárdy, L.; Fülöp, F.; Toldi, J. Kynurenines in the CNS: Recent advances and new questions. Nat. Rev. Drug. Discov. 2013, 12, 64–82. [Google Scholar] [CrossRef] [PubMed]
- Coppen, A. The biochemistry of affective disorders. Br. J. Psychiatry 1967, 113, 1237–1264. [Google Scholar] [CrossRef] [PubMed]
- Joensuu, M.; Tolmunen, T.; Saarinen, P.I.; Tiihonen, J.; Kuikka, J.; Ahola, P.; Vanninen, R.; Lehtonen, J. Reduced midbrain serotonin transporter availability in drug-naïve patients with depression measured by SERT-specific [(123)I] nor-beta-CIT SPECT imaging. Psychiatry Res. 2007, 154, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Sargent, P.A.; Kjaer, K.H.; Bench, C.J.; Rabiner, E.A.; Messa, C.; Meyer, J.; Gunn, R.N.; Grasby, P.M.; Cowen, P.J. Brain serotonin1A receptor binding measured by positron emission tomography with [11C]WAY-100635: Effects of depression and antidepressant treatment. Arch. Gen. Psychiatry 2000, 57, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Drevets, W.C.; Thase, M.E.; Moses-Kolko, E.L.; Price, J.; Frank, E.; Kupfer, D.J.; Mathis, C. Serotonin-1A receptor imaging in recurrent depression: Replication and literature review. Nucl. Med. Biol. 2007, 34, 865–877. [Google Scholar] [CrossRef] [PubMed]
- Yatham, L.N.; Liddle, P.F.; Shiah, I.S.; Scarrow, G.; Lam, R.W.; Adam, M.J.; Zis, A.P.; Ruth, T.J. Brain serotonin2 receptors in major depression: A positron emission tomography study. Arch. Gen. Psychiatry 2000, 57, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Albert, P.R.; Benkelfat, C.; Descarries, L. The neurobiology of depression--revisiting the serotonin hypothesis. I. Cellular and molecular mechanisms. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 2378–2381. [Google Scholar] [CrossRef] [PubMed]
- Reimold, M.; Batra, A.; Knobel, A.; Smolka, M.N.; Zimmer, A.; Mann, K.; Solbach, C.; Reischl, G.; Schwärzler, F.; Gründer, G.; et al. Anxiety is associated with reduced central serotonin transporter availability in unmedicated patients with unipolar major depression: A [11C]DASB PET study. Mol. Psychiatry 2008, 13, 606–613. [Google Scholar] [CrossRef]
- Cowen, P.J.; Browning, M. What has serotonin to do with depression? World Psychiatry 2015, 14, 158–160. [Google Scholar] [CrossRef]
- Lapin, I.P.; Oxenkrug, G.F. Intensification of the central serotoninergic processes as a possible determinant of the thymoleptic effect. Lancet 1969, 293, 132–136. [Google Scholar] [CrossRef]
- Salter, M.; Knowles, R.G.; Pogson, C.I. How does displacement of albumin-bound tryptophan cause sustained increases in the free tryptophan concentration in plasma and 5-hydroxytryptamine synthesis in brain? Biochem. J. 1989, 262, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Höglund, E.; Øverli, Ø.; Winberg, S. Tryptophan Metabolic Pathways and Brain Serotonergic Activity: A Comparative Review. Front. Endocrinol. 2019, 10, 158. [Google Scholar] [CrossRef] [PubMed]
- Ogyu, K.; Kubo, K.; Noda, Y.; Iwata, Y.; Tsugawa, S.; Omura, Y.; Wada, M.; Tarumi, R.; Plitman, E.; Moriguchi, S.; et al. Kynurenine pathway in depression: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2018, 90, 16–25. [Google Scholar] [CrossRef]
- Wu, H.; Gong, J.; Liu, Y. Indoleamine 2,3-dioxygenase regulation of immune response (Review). Mol. Med. Rep. 2018, 17, 4867–4873. [Google Scholar] [CrossRef] [PubMed]
- Myint, A.M. Kynurenines: From the perspective of major psychiatric disorders. FEBS J. 2012, 279, 1375–1385. [Google Scholar] [CrossRef]
- Myint, A.-M.; Kim, Y.K.; Verkerk, R.; Scharpé, S.; Steinbusch, H.; Leonard, B. Kynurenine pathway in major depression: Evidence of impaired neuroprotection. J. Affect. Disord. 2007, 98, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Myint, A.-M.; Schwarz, M.J.; Müller, N. The role of the kynurenine metabolism in major depression. J. Neural Transm. 2012, 119, 245–251. [Google Scholar] [CrossRef]
- Oxenkrug, G. Serotonin-kynurenine hypothesis of depression: Historical overview and recent developments. Curr. Drug Targets 2013, 14, 514–521. [Google Scholar] [CrossRef]
- Kruse, J.L.; Cho, J.H.-J.; Olmstead, R.; Hwang, L.; Faull, K.; Eisenberger, N.I.; Irwin, M.R. Kynurenine metabolism and inflammation-induced depressed mood: A human experimental study. Psychoneuroendocrinology 2019, 109, 104371. [Google Scholar] [CrossRef]
- Boros, F.A.; Bohár, Z.; Vécsei, L. Genetic alterations affecting the genes encoding the enzymes of the kynurenine pathway and their association with human diseases. Mutat. Res. Mutat. Res. 2018, 776, 32–45. [Google Scholar] [CrossRef]
- Hughes, M.M.; Carballedo, A.; McLoughlin, D.M.; Amico, F.; Harkin, A.; Frodl, T.; Connor, T.J. Tryptophan depletion in depressed patients occurs independent of kynurenine pathway activation. Brain. Behav. Immun. 2012, 26, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Quak, J.; Doornbos, B.; Roest, A.M.; Duivis, H.E.; Vogelzangs, N.; Nolen, W.A.; Penninx, B.W.J.H.; Kema, I.P.; de Jonge, P. Does tryptophan degradation along the kynurenine pathway mediate the association between pro-inflammatory immune activity and depressive symptoms? Psychoneuroendocrinology 2014, 45, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Dahl, J.; Andreassen, O.A.; Verkerk, R.; Malt, U.F.; Sandvik, L.; Brundin, L.; Ormstad, H. Ongoing episode of major depressive disorder is not associated with elevated plasma levels of kynurenine pathway markers. Psychoneuroendocrinology 2015, 56, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Savitz, J.; Drevets, W.C.; Wurfel, B.E.; Ford, B.N.; Bellgowan, P.S.F.; Victor, T.A.; Bodurka, J.; Teague, T.K.; Dantzer, R. Reduction of kynurenic acid to quinolinic acid ratio in both the depressed and remitted phases of major depressive disorder. Brain Behav. Immun. 2015, 46, 55–59. [Google Scholar] [CrossRef]
- Schwarcz, R.; Bruno, J.P.; Muchowski, P.J.; Wu, H.-Q. Kynurenines in the mammalian brain: When physiology meets pathology. Nat. Rev. Neurosci. 2012, 13, 465–477. [Google Scholar] [CrossRef]
- Gururajan, A.; Clarke, G.; Dinan, T.G.; Cryan, J.F. Molecular biomarkers of depression. Neurosci. Biobehav. Rev. 2016, 64, 101–133. [Google Scholar] [CrossRef]
- Leonard, B.E. Inflammation and depression: A causal or coincidental link to the pathophysiology? Acta Neuropsychiatr. 2018, 30, 1–16. [Google Scholar] [CrossRef]
- Leonard, B.E. Inflammation, depression and dementia: Are they connected? Neurochem. Res. 2007, 32, 1749–1756. [Google Scholar] [CrossRef]
- Martínez-Cengotitabengoa, M.; Carrascón, L.; O’Brien, J.; Díaz-Gutiérrez, M.-J.; Bermúdez-Ampudia, C.; Sanada, K.; Arrasate, M.; González-Pinto, A. Peripheral Inflammatory Parameters in Late-Life Depression: A Systematic Review. Int. J. Mol. Sci. 2016, 17, 2022. [Google Scholar] [CrossRef] [PubMed]
- Strawbridge, R.; Young, A.H.; Cleare, A.J. Biomarkers for depression: Recent insights, current challenges and future prospects. Neuropsychiatr. Dis. Treat. 2017, 13, 1245–1262. [Google Scholar] [CrossRef]
- Hacimusalar, Y.; Eşel, E. Suggested Biomarkers for Major Depressive Disorder. Noro Psikiyatr. Ars. 2018, 55, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Ranieri, R.; Laezza, C.; Bifulco, M.; Marasco, D.; Malfitano, A.M. Cannabinoids and Neuro-Inflammation: Regulation of Brain Immune Response. Recent Pat. CNS Drug Discov. 2016, 10, 178–203. [Google Scholar] [CrossRef]
- Klein, T.; Lane, B.; Newton, C.; Friedman, H. The cannabinoid system and cytokine network. Proc. Soc. Exp. Biol. Med. 2000, 225, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Massi, P.; Vaccani, A.; Parolaro, D. Cannabinoids, immune system and cytokine network. Curr. Pharm. Des. 2006, 12, 3135–3146. [Google Scholar] [CrossRef] [PubMed]
- Jean-Gilles, L.; Braitch, M.; Latif, M.L.; Aram, J.; Fahey, A.J.; Edwards, L.J.; Robins, R.A.; Tanasescu, R.; Tighe, P.J.; Gran, B.; et al. Effects of pro-inflammatory cytokines on cannabinoid CB1 and CB2 receptors in immune cells. Acta Physiol. 2015, 214, 63. [Google Scholar] [CrossRef]
- Mandolesi, G.; Bullitta, S.; Fresegna, D.; Gentile, A.; De Vito, F.; Dolcetti, E.; Rizzo, F.R.; Strimpakos, G.; Centonze, D.; Musella, A. Interferon-γ causes mood abnormalities by altering cannabinoid CB1 receptor function in the mouse striatum. Neurobiol. Dis. 2017, 108, 45–53. [Google Scholar] [CrossRef]
- Rossi, S.; Motta, C.; Musella, A.; Centonze, D. The interplay between inflammatory cytokines and the endocannabinoid system in the regulation of synaptic transmission. Neuropharmacology 2015, 96, 105–112. [Google Scholar] [CrossRef]
- Rossi, S.; Sacchetti, L.; Napolitano, F.; De Chiara, V.; Motta, C.; Studer, V.; Musella, A.; Barbieri, F.; Bari, M.; Bernardi, G.; et al. Interleukin-1β causes anxiety by interacting with the endocannabinoid system. J. Neurosci. 2012, 32, 13896–13905. [Google Scholar] [CrossRef]
- Watzl, B.; Scuderi, P.; Watson, R.R. Marijuana components stimulate human peripheral blood mononuclear cell secretion of interferon-gamma and suppress interleukin-1 alpha in vitro. Int. J. Immunopharmacol. 1991, 13, 1091–1097. [Google Scholar] [CrossRef]
- Berdyshev, E.V.; Boichot, E.; Germain, N.; Allain, N.; Anger, J.-P.; Lagente, V. Influence of fatty acid ethanolamides and Δ9-tetrahydrocannabinol on cytokine and arachidonate release by mononuclear cells. Eur. J. Pharmacol. 1997, 330, 231–240. [Google Scholar] [CrossRef]
- Klein, T.W.; Friedman, H.; Specter, S. Marijuana, immunity and infection. J. Neuroimmunol. 1998, 83, 102–115. [Google Scholar] [CrossRef]
- Klein, T.W.; Newton, C.; Zhu, W.; Daaka, Y.; Friedman, H. Delta 9-Tetrahydrocannabinol, Cytokines, and Immunity to Legionella pneumophila. Exp. Biol. Med. 1995, 209, 205–212. [Google Scholar] [CrossRef]
- Suárez-Pinilla, P.; López-Gil, J.; Crespo-Facorro, B. Immune system: A possible nexus between cannabinoids and psychosis. Brain Behav. Immun. 2014, 40, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Cabral, G.A.; Griffin-Thomas, L. Emerging Role of the CB2 Cannabinoid Receptor in Immune Regulation and Therapeutic Prospects. Expert Rev. Mol. Med. 2010, 11, e3. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, M.D.; Srivastava, B.I.; Brouhard, B. Δ9 Tetrahydrocannabinol and cannabidiol alter cytokine production by human immune cells. Immunopharmacology 1998, 40, 179–185. [Google Scholar] [CrossRef]
- Cutando, L.; Busquets-Garcia, A.; Puighermanal, E.; Gomis-González, M.; Delgado-García, J.M.; Gruart, A.; Maldonado, R.; Ozaita, A. Microglial activation underlies cerebellar deficits produced by repeated cannabis exposure. J. Clin. Investig. 2013, 123, 2816–2831. [Google Scholar] [CrossRef] [PubMed]
- García-Gutiérrez, M.S.; Navarrete, F.; Gasparyan, A.; Austrich-Olivares, A.; Sala, F.; Manzanares, J. Cannabidiol: A Potential New Alternative for the Treatment of Anxiety, Depression, and Psychotic Disorders. Biomolecules 2020, 10, 1575. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.M.; Kaplan, B.L.F. Immune Responses Regulated by Cannabidiol. Cannabis Cannabinoid Res. 2020, 5, 12–31. [Google Scholar] [CrossRef] [PubMed]
- Burstein, S. Cannabidiol (CBD) and its analogs: A review of their effects on inflammation. Bioorg. Med. Chem. 2015, 23, 1377–1385. [Google Scholar] [CrossRef]
- Hahn, B. The Potential of Cannabidiol Treatment for Cannabis Users With Recent-Onset Psychosis. Schizophr. Bull. 2018, 44, 46–53. [Google Scholar] [CrossRef]
- Calpe-López, C.; García-Pardo, M.P.; Aguilar, M.A. Cannabidiol Treatment Might Promote Resilience to Cocaine and Methamphetamine Use Disorders: A Review of Possible Mechanisms. Molecules 2019, 24, 2583. [Google Scholar] [CrossRef] [PubMed]
- Moretti, S.; Castelli, M.; Franchi, S.; Raggi, M.A.; Mercolini, L.; Protti, M.; Somaini, L.; Panerai, A.E.; Sacerdote, P. Δ9-Tetrahydrocannabinol-induced anti-inflammatory responses in adolescent mice switch to proinflammatory in adulthood. J. Leukoc. Biol. 2014, 96, 523–534. [Google Scholar] [CrossRef]
- Moretti, S.; Franchi, S.; Castelli, M.; Amodeo, G.; Somaini, L.; Panerai, A.; Sacerdote, P. Exposure of Adolescent Mice to Delta-9-Tetrahydrocannabinol Induces Long-Lasting Modulation of Pro- and Anti-Inflammatory Cytokines in Hypothalamus and Hippocampus Similar to that Observed for Peripheral Macrophages. J. Neuroimmune Pharmacol. 2015, 10, 371–379. [Google Scholar] [CrossRef]
- Zamberletti, E.; Gabaglio, M.; Prini, P.; Rubino, T.; Parolaro, D. Cortical neuroinflammation contributes to long-term cognitive dysfunctions following adolescent delta-9-tetrahydrocannabinol treatment in female rats. Eur. Neuropsychopharmacol. 2015, 25, 2404–2415. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of DSM-5, 5th ed.; American Psychiatric Publishing: London, UK; Washington, DC, USA, 2013; ISBN 978-0-89042-554-1. [Google Scholar]
- Bayazit, H.; Selek, S.; Karababa, I.F.; Cicek, E.; Aksoy, N. Evaluation of Oxidant/Antioxidant Status and Cytokine Levels in Patients with Cannabis Use Disorder. Clin. Psychopharmacol. Neurosci. 2017, 15, 237–242. [Google Scholar] [CrossRef]
- Lisano, J.K.; Kisiolek, J.N.; Smoak, P.; Phillips, K.T.; Stewart, L.K. Chronic cannabis use and circulating biomarkers of neural health, stress, and inflammation in physically active individuals. Appl. Physiol. Nutr. Metab. 2020, 45, 258–263. [Google Scholar] [CrossRef]
- Zawatsky, C.N.; Abdalla, J.; Cinar, R. Synthetic cannabinoids induce acute lung inflammation via cannabinoid receptor 1 activation. ERJ Open Res. 2020, 6, 00121–02020. [Google Scholar] [CrossRef] [PubMed]
- Derocq, J.-M.; Jbilo, O.; Bouaboula, M.; Ségui, M.; Clère, C.; Casellas, P. Genomic and Functional Changes Induced by the Activation of the Peripheral Cannabinoid Receptor CB2 in the Promyelocytic Cells HL-60. J. Biol. Chem. 2000, 275, 15621–15628. [Google Scholar] [CrossRef]
- Nahoko, U.; Ruri, K.-H.; Nobuo, K.; Yuji, H.; Yukihiro, G. Identification of a cannabinoid analog as a new type of designer drug in a herbal product. Chem. Pharm. Bull. 2009, 57, 439–441. [Google Scholar]
- Bileck, A.; Ferk, F.; Al-Serori, H.; Koller, V.J.; Muqaku, B.; Haslberger, A.; Auwärter, V.; Gerner, C.; Knasmüller, S. Impact of a synthetic cannabinoid (CP-47,497-C8) on protein expression in human cells: Evidence for induction of inflammation and DNA damage. Arch. Toxicol. 2016, 90, 1369–1382. [Google Scholar] [CrossRef]
- Zádor, F.; Mohammadzadeh, A.; Balogh, M.; Zádori, Z.S.; Király, K.; Barsi, S.; Galambos, A.R.; László, S.B.; Hutka, B.; Váradi, A.; et al. Comparisons of In Vivo and In Vitro Opioid Effects of Newly Synthesized 14-Methoxycodeine-6-O-sulfate and Codeine-6-O-sulfate. Molecules 2020, 25, 1370. [Google Scholar] [CrossRef]
- Christensen, R.; Kristensen, P.K.; Bartels, E.M.; Bliddal, H.; Astrup, A. Efficacy and safety of the weight-loss drug rimonabant: A meta-analysis of randomised trials. Lancet 2007, 370, 1706–1713. [Google Scholar] [CrossRef]
- Beyer, C.E.; Dwyer, J.M.; Piesla, M.J.; Platt, B.J.; Shen, R.; Rahman, Z.; Chan, K.; Manners, M.T.; Samad, T.A.; Kennedy, J.D.; et al. Depression-like phenotype following chronic CB1 receptor antagonism. Neurobiol. Dis. 2010, 39, 148–155. [Google Scholar] [CrossRef]
- Guler, E.; Bektay, M.; Akyildiz, A.; Sisman, B.; Izzettin, F.; Kocyigit, A. Investigation of DNA damage, oxidative stress, and inflammation in synthetic cannabinoid users. Hum. Exp. Toxicol. 2020, 39, 1454–1462. [Google Scholar] [CrossRef]
- Campbell, B.M.; Charych, E.; Lee, A.W.; Möller, T. Kynurenines in CNS disease: Regulation by inflammatory cytokines. Front. Neurosci. 2014, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Strasser, B.; Becker, K.; Fuchs, D.; Gostner, J.M. Kynurenine pathway metabolism and immune activation: Peripheral measurements in psychiatric and co-morbid conditions. Neuropharmacology 2017, 112, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, R.; Forteza, M.J.; Ketelhuth, D.F.J. The interplay between cytokines and the Kynurenine pathway in inflammation and atherosclerosis. Cytokine 2019, 122, 154148. [Google Scholar] [CrossRef]
- Mándi, Y.; Vécsei, L. The kynurenine system and immunoregulation. J. Neural Transm. 2012, 119, 197–209. [Google Scholar] [CrossRef]
- Clark, S.M.; Pocivavsek, A.; Nicholson, J.D.; Notarangelo, F.M.; Langenberg, P.; McMahon, R.P.; Kleinman, J.E.; Hyde, T.M.; Stiller, J.; Postolache, T.T.; et al. Reduced kynurenine pathway metabolism and cytokine expression in the prefrontal cortex of depressed individuals. J. Psychiatry Neurosci. 2016, 41, 386–394. [Google Scholar] [CrossRef]
- Byrne, G.I.; Lehmann, L.K.; Landry, G.J. Induction of tryptophan catabolism is the mechanism for gamma-interferon-mediated inhibition of intracellular Chlamydia psittaci replication in T24 cells. Infect. Immun. 1986, 53, 347–351. [Google Scholar] [CrossRef]
- Pfefferkorn, E.R.; Rebhun, S.; Eckel, M. Characterization of an indoleamine 2,3-dioxygenase induced by gamma-interferon in cultured human fibroblasts. J. Interferon Res. 1986, 6, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Hissong, B.D.; Byrne, G.I.; Padilla, M.L.; Carlin, J.M. Upregulation of interferon-induced indoleamine 2,3-dioxygenase in human macrophage cultures by lipopolysaccharide, muramyl tripeptide, and interleukin-1. Cell. Immunol. 1995, 160, 264–269. [Google Scholar] [CrossRef]
- Hissong, B.D.; Carlin, J.M. Potentiation of Interferon-Induced Indoleamine 2,3-Dioxygenase mRNA in Human Mononuclear Phagocytes by Lipopolysaccharide and Interleukin-1. J. Interf. Cytokine Res. 1997, 17, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Babcock, T.A.; Carlin, J.M. Transcriptional activation of indoleamine dioxygenase by interleukin 1 and tumor necrosis factor α in interferon-treated epithelial cells. Cytokine 2000, 12, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Fujigaki, H.; Saito, K.; Fujigaki, S.; Takemura, M.; Sudo, K.; Ishiguro, H.; Seishima, M. The Signal Transducer and Activator of Transcription 1α and Interferon Regulatory Factor 1 Are Not Essential for the Induction of Indoleamine 2,3-Dioxygenase by Lipopolysaccharide: Involvement of p38 Mitogen-Activated Protein Kinase and Nuclear Factor-κB. J. Biochem. 2006, 139, 655–662. [Google Scholar] [CrossRef]
- Brooks, A.K.; Lawson, M.A.; Rytych, J.L.; Yu, K.C.; Janda, T.M.; Steelman, A.J.; McCusker, R.H. Immunomodulatory Factors Galectin-9 and Interferon-Gamma Synergize to Induce Expression of Rate-Limiting Enzymes of the Kynurenine Pathway in the Mouse Hippocampus. Front. Immunol. 2016, 7, 422. [Google Scholar] [CrossRef]
- Brooks, A.K.; Lawson, M.A.; Smith, R.A.; Janda, T.M.; Kelley, K.W.; McCusker, R.H. Interactions between inflammatory mediators and corticosteroids regulate transcription of genes within the Kynurenine Pathway in the mouse hippocampus. J. Neuroinflamm. 2016, 13, 98. [Google Scholar] [CrossRef]
- Fuertig, R.; Azzinnari, D.; Bergamini, G.; Cathomas, F.; Sigrist, H.; Seifritz, E.; Vavassori, S.; Luippold, A.; Hengerer, B.; Ceci, A.; et al. Mouse chronic social stress increases blood and brain kynurenine pathway activity and fear behaviour: Both effects are reversed by inhibition of indoleamine 2,3-dioxygenase. Brain Behav. Immun. 2016, 54, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Duda, W.; Curzytek, K.; Kubera, M.; Connor, T.J.; Fagan, E.M.; Basta-Kaim, A.; Trojan, E.; Papp, M.; Gruca, P.; Budziszewska, B.; et al. Interaction of the immune-inflammatory and the kynurenine pathways in rats resistant to antidepressant treatment in model of depression. Int. Immunopharmacol. 2019, 73, 527–538. [Google Scholar] [CrossRef]
- Hu, B.; Hissong, B.D.; Carlin, J.M. Interleukin-1 Enhances Indoleamine 2,3-Dioxygenase Activity by Increasing Specific mRNA Expression in Human Mononuclear Phagocytes. J. Interf. Cytokine Res. 1995, 15, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Corona, A.W.; Huang, Y.; O’Connor, J.C.; Dantzer, R.; Kelley, K.W.; Popovich, P.G.; Godbout, J.P. Fractalkine receptor (CX3CR1) deficiency sensitizes mice to the behavioral changes induced by lipopolysaccharide. J. Neuroinflamm. 2010, 7, 93. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Pena, D.; Nixon, S.E.; O’Connor, J.C.; Southey, B.R.; Lawson, M.A.; McCusker, R.H.; Borras, T.; Machuca, D.; Hernandez, A.G.; Dantzer, R.; et al. Microglia Transcriptome Changes in a Model of Depressive Behavior after Immune Challenge. PLoS ONE 2016, 11, e150858. [Google Scholar] [CrossRef]
- Garrison, A.M.; Parrott, J.M.; Tuñon, A.; Delgado, J.; Redus, L.; O’connor, J.C. Kynurenine pathway metabolic balance influences microglia activity: Targeting kynurenine monooxygenase to dampen neuroinflammation HHS Public Access. Psychoneuroendocrinology 2018, 94, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Laumet, G.; Zhou, W.; Dantzer, R.; Edralin, J.D.; Huo, X.; Budac, D.P.; O’connor, J.C.; Lee, A.W.; Heijnen, C.J.; Kavelaars, A. Upregulation of neuronal kynurenine 3-monooxygenase mediates depression-like behavior in a mouse model of neuropathic pain HHS Public Access. Brain Behav. Immun. 2017, 66, 94–102. [Google Scholar] [CrossRef]
- Von Bubnoff, D.; Scheler, M.; Wilms, H.; Fimmers, R.; Bieber, T. Identification of IDO-positive and IDO-negative human dendritic cells after activation by various proinflammatory stimuli. J. Immunol. 2011, 186, 6701–6709. [Google Scholar] [CrossRef] [PubMed]
- Zunszain, P.A.; Anacker, C.; Cattaneo, A.; Choudhury, S.; Musaelyan, K.; Myint, A.M.; Thuret, S.; Price, J.; Pariante, C.M. Interleukin-1β: A new regulator of the kynurenine pathway affecting human hippocampal neurogenesis. Neuropsychopharmacology 2012, 37, 939–949. [Google Scholar] [CrossRef]
- Xie, W.; Cai, L.; Yu, Y.; Gao, L.; Xiao, L.; He, Q.; Ren, Z.; Liu, Y. Activation of brain indoleamine 2,3-dioxygenase contributes to epilepsy-associated depressive-like behavior in rats with chronic temporal lobe epilepsy. J. Neuroinflamm. 2014, 11, 41. [Google Scholar] [CrossRef]
- Lian, Y.-J.; Gong, H.; Wu, T.-Y.; Su, W.-J.; Zhang, Y.; Yang, Y.-Y.; Peng, W.; Zhang, T.; Zhou, J.-R.; Jiang, C.-L.; et al. Ds-HMGB1 and fr-HMGB induce depressive behavior through neuroinflammation in contrast to nonoxid-HMGB1. Brain. Behav. Immun. 2017, 59, 322–332. [Google Scholar] [CrossRef]
- Wang, B.; Lian, Y.; Su, W.; Liu, L.; Li, J.; Jiang, C.; Wang, Y. Fr-HMGB1 and ds-HMGB1 activate the kynurenine pathway via different mechanisms in association with depressive-like behavior. Mol. Med. Rep. 2019, 20, 359–367. [Google Scholar] [CrossRef]
- Maes, M.; Meltzer, H.Y.; Scharpè, S.; Bosmans, E.; Suy, E.; De Meester, I.; Calabrese, J.; Cosyns, P. Relationships between lower plasma L-tryptophan levels and immune-inflammatory variables in depression. Psychiatry Res. 1993, 49, 151–165. [Google Scholar] [CrossRef]
- Maes, M.; Verkerk, R.; Bonaccorso, S.; Ombelet, W.; Bosmans, E.; Scharpé, S. Depressive and anxiety symptoms in the early puerperium are related to increased degradation of tryptophan into kynurenine, a phenomenon which is related to immune activation. Life Sci. 2002, 71, 1837–1848. [Google Scholar] [CrossRef]
- Schwieler, L.; Samuelsson, M.; Frye, M.A.; Bhat, M.; Schuppe-Koistinen, I.; Jungholm, O.; Johansson, A.G.; Landén, M.; Sellgren, C.M.; Erhardt, S. Electroconvulsive therapy suppresses the neurotoxic branch of the kynurenine pathway in treatment-resistant depressed patients. J. Neuroinflamm. 2016, 13, 51. [Google Scholar] [CrossRef]
- Achtyes, E.; Keaton, S.A.; Smart, L.; Burmeister, A.R.; Heilman, P.L.; Krzyzanowski, S.; Nagalla, M.; Guillemin, G.J.; Escobar Galvis, M.L.; Lim, C.K.; et al. Inflammation and kynurenine pathway dysregulation in post-partum women with severe and suicidal depression. Brain Behav. Immun. 2020, 83, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Westbrook, R.; Chung, T.; Lovett, J.; Ward, C.; Joca, H.; Yang, H.; Khadeer, M.; Tian, J.; Xue, Q.-L.; Le, A.; et al. Kynurenines link chronic inflammation to functional decline and physical frailty. JCI Insight 2020, 5. [Google Scholar] [CrossRef]
- Anderson, G.; Kubera, M.; Duda, W.; Lasoń, W.; Berk, M.; Maes, M. Increased IL-6 trans-signaling in depression: Focus on the tryptophan catabolite pathway, melatonin and neuroprogression. Pharmacol. Rep. 2013, 65, 1647–1654. [Google Scholar] [CrossRef]
- Maes, M. Evidence for an immune response in major depression: A review and hypothesis. Prog. Neuro Psychopharmacol. Biol. Psychiatry 1995, 19, 11–38. [Google Scholar] [CrossRef]
- Bay-Richter, C.; Linderholm, K.R.; Lim, C.K.; Samuelsson, M.; Träskman-Bendz, L.; Guillemin, G.J.; Erhardt, S.; Brundin, L. A role for inflammatory metabolites as modulators of the glutamate N-methyl-d-aspartate receptor in depression and suicidality. Brain Behav. Immun. 2015, 43, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Chen, L.; Lim, G.; Sung, B.; Wang, S.; McCabe, M.F.; Rusanescu, G.; Yang, L.; Tian, Y.; Mao, J. Brain indoleamine 2,3-dioxygenase contributes to the comorbidity of pain and depression. J. Clin. Investig. 2012, 122, 2940–2954. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Sheng, H.; Tang, Z.; Lu, J.; Ni, X. Inflammation and increased IDO in hippocampus contribute to depression-like behavior induced by estrogen deficiency. Behav. Brain Res. 2015, 288, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Connor, T.J.; Starr, N.; O’Sullivan, J.B.; Harkin, A. Induction of indolamine 2,3-dioxygenase and kynurenine 3-monooxygenase in rat brain following a systemic inflammatory challenge: A role for IFN-γ? Neurosci. Lett. 2008, 441, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Bonaccorso, S.; Marino, V.; Puzella, A.; Pasquini, M.; Biondi, M.; Artini, M.; Almerighi, C.; Meltzer, H. Treatment with interferon-alpha (IFNα) of hepatitis C patients induces lower serum dipeptidyl peptidase IV activity, which is related to IFNα-induced depressive and anxiety symptoms and immune activation. Mol. Psychiatry 2001, 6, 475–480. [Google Scholar] [CrossRef]
- Zoga, M.; Oulis, P.; Chatzipanagiotou, S.; Masdrakis, V.G.; Pliatsika, P.; Boufidou, F.; Foteli, S.; Soldatos, C.R.; Nikolaou, C.; Papageorgiou, C. Indoleamine 2,3-dioxygenase and immune changes under antidepressive treatment in major depression in females. In Vivo 2014, 28, 633–638. [Google Scholar]
- Haroon, E.; Welle, J.R.; Woolwine, B.J.; Goldsmith, D.R.; Baer, W.; Patel, T.; Felger, J.C.; Miller, A.H. Associations among peripheral and central kynurenine pathway metabolites and inflammation in depression. Neuropsychopharmacology 2020, 45, 998–1007. [Google Scholar] [CrossRef]
- O’Connor, J.C.; Lawson, M.A.; André, C.; Briley, E.M.; Szegedi, S.S.; Lestage, J.; Castanon, N.; Herkenham, M.; Dantzer, R.; Kelley, K.W. Induction of IDO by bacille Calmette-Guérin is responsible for development of murine depressive-like behavior. J. Immunol. 2009, 182, 3202–3212. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.C.; André, C.; Wang, Y.; Lawson, M.A.; Szegedi, S.S.; Lestage, J.; Castanon, N.; Kelley, K.W.; Dantzer, R. Interferon-gamma and tumor necrosis factor-alpha mediate the upregulation of indoleamine 2,3-dioxygenase and the induction of depressive-like behavior in mice in response to bacillus Calmette-Guerin. J. Neurosci. 2009, 29, 4200–4209. [Google Scholar] [CrossRef]
- Currier, A.R.; Ziegler, M.H.; Riley, M.M.; Babcock, T.A.; Telbis, V.P.; Carlin, J.M. Tumor Necrosis Factor-alpha and Lipopolysaccharide Enhance Interferon-Induced Antichlamydial Indoleamine Dioxygenase Activity Independently. J. Interferon Cytokine Res. 2000, 20, 369–376. [Google Scholar] [CrossRef]
- Robinson, C.M.; Hale, P.T.; Carlin, J.M. The role of IFN-gamma and TNF-alpha-responsive regulatory elements in the synergistic induction of indoleamine dioxygenase. J. Interferon Cytokine Res. 2005, 25, 20–30. [Google Scholar] [CrossRef]
- Pacher, P.; Kunos, G. Modulating the endocannabinoid system in human health and disease—Successes and failures. FEBS J. 2013, 280, 1918–1943. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Bátkai, S.; Kunos, G. The endocannabinoid system as an emerging target of pharmacotherapy. Pharmacol. Rev. 2006, 58, 389–462. [Google Scholar] [CrossRef]
- Zimmer, A.; Zimmer, A.M.; Hohmann, A.G.; Herkenham, M.; Bonner, T.I. Increased mortality, hypoactivity, and hypoalgesia in cannabinoid CB1 receptor knockout mice. Proc. Natl. Acad. Sci. USA 1999, 96, 5780–5785. [Google Scholar] [CrossRef]
- Buckley, N.E.; McCoy, K.L.; Mezey, É.; Bonner, T.; Zimmer, A.; Felder, C.C.; Glass, M.; Zimmer, A. Immunomodulation by cannabinoids is absent in mice deficient for the cannabinoid CB2 receptor. Eur. J. Pharmacol. 2000, 396, 141–149. [Google Scholar] [CrossRef]
- Giorgini, F.; Huang, S.-Y.; Sathyasaikumar, K.V.; Notarangelo, F.M.; Thomas, M.A.R.; Tararina, M.; Wu, H.-Q.; Schwarcz, R.; Muchowski, P.J. Targeted deletion of kynurenine 3-monooxygenase in mice: A new tool for studying kynurenine pathway metabolism in periphery and brain. J. Biol. Chem. 2013, 288, 36554–36566. [Google Scholar] [CrossRef]
- Erhardt, S.; Pocivavsek, A.; Repici, M.; Liu, X.-C.; Imbeault, S.; Maddison, D.C.; Thomas, M.A.R.; Smalley, J.L.; Larsson, M.K.; Muchowski, P.J.; et al. Adaptive and Behavioral Changes in Kynurenine 3-Monooxygenase Knockout Mice: Relevance to Psychotic Disorders. Biol. Psychiatry 2017, 82, 756–765. [Google Scholar] [CrossRef]
- Járai, Z.; Wagner, J.A.; Varga, K.; Lake, K.D.; Compton, D.R.; Martin, B.R.; Zimmer, A.M.; Bonner, T.I.; Buckley, N.E.; Mezey, E.; et al. Cannabinoid-induced mesenteric vasodilation through an endothelial site distinct from CB1 or CB2 receptors. Proc. Natl. Acad. Sci. USA 1999, 96, 14136–14141. [Google Scholar] [CrossRef]
- Hirata, N.; Hattori, S.; Shoji, H.; Funakoshi, H.; Miyakawa, T. Comprehensive behavioral analysis of indoleamine 2,3-dioxygenase knockout mice. Neuropsychopharmacol. Rep. 2018, 38, 133–144. [Google Scholar] [CrossRef]
- Yu, P.; Di Prospero, N.A.; Sapko, M.T.; Cai, T.; Chen, A.; Melendez-Ferro, M.; Du, F.; Whetsell, W.O.; Guidetti, P.; Schwarcz, R.; et al. Biochemical and phenotypic abnormalities in kynurenine aminotransferase II-deficient mice. Mol. Cell. Biol. 2004, 24, 6919–6930. [Google Scholar] [CrossRef]
- Bondulich, M.K.; Fan, Y.; Song, Y.; Giorgini, F.; Bates, G.P. Ablation of kynurenine 3-monooxygenase rescues plasma inflammatory cytokine levels in the R6/2 mouse model of Huntington’s disease. Sci. Rep. 2021, 11, 5484. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).