
The Mildew Resistance Locus O 4 Interacts with CaM/CML and Is Involved in Root Gravity Response

Abstract
:1. Introduction
2. Results
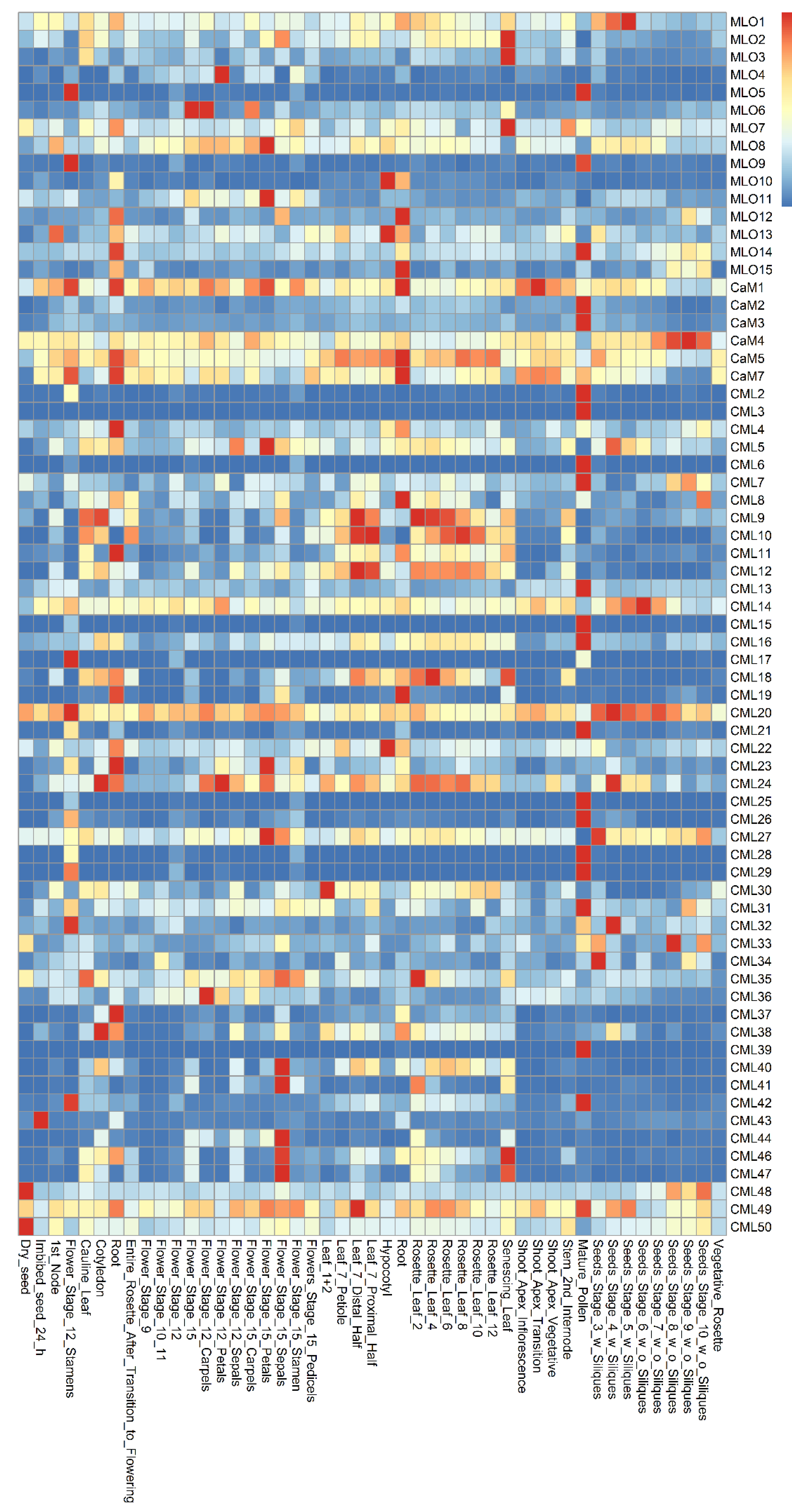
2.1. The Expression Patterns of MLO and CaM/CML Genes
2.2. Screening for Interactive Pairs of MLO and CaM/CML Proteins
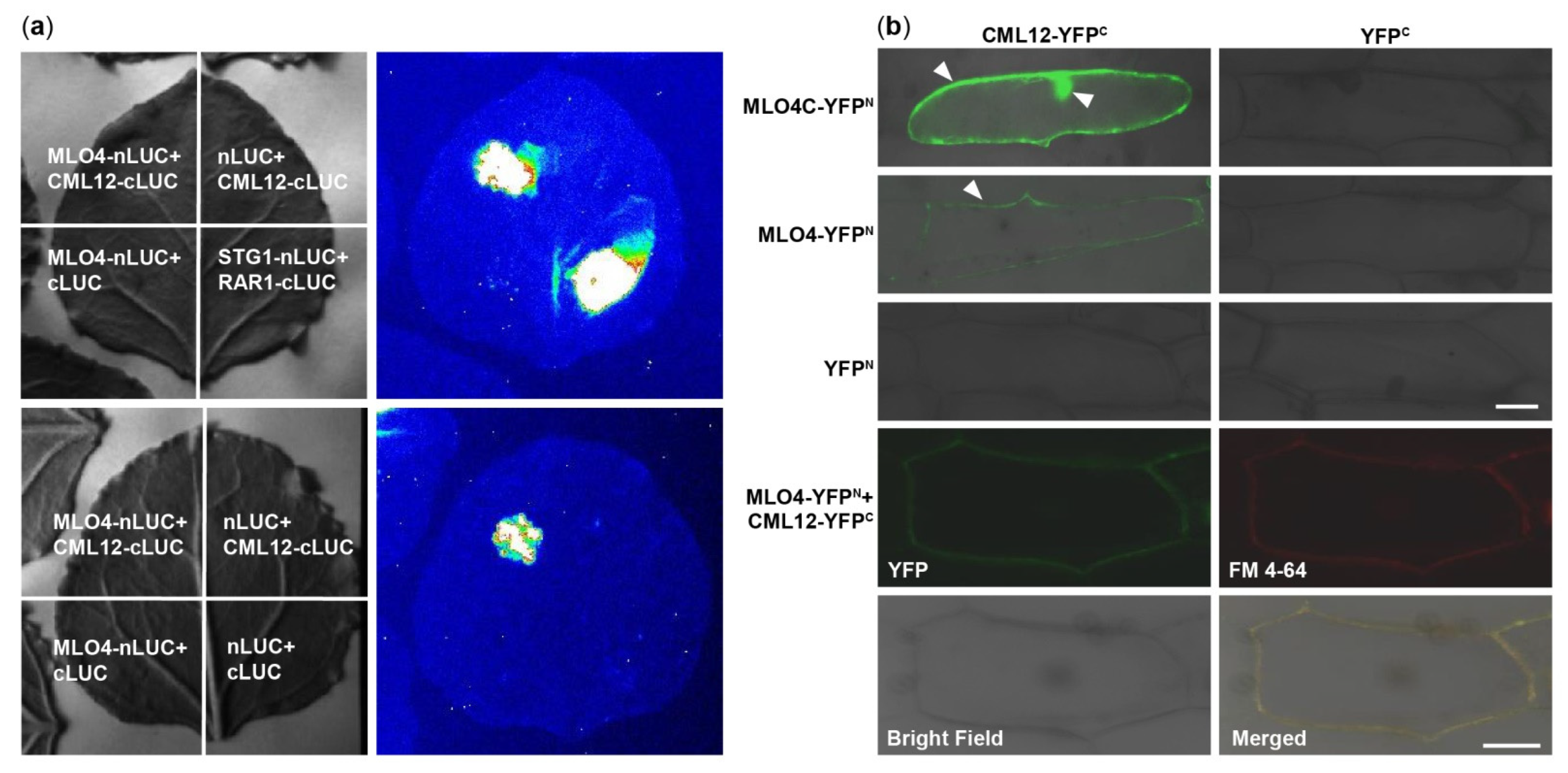
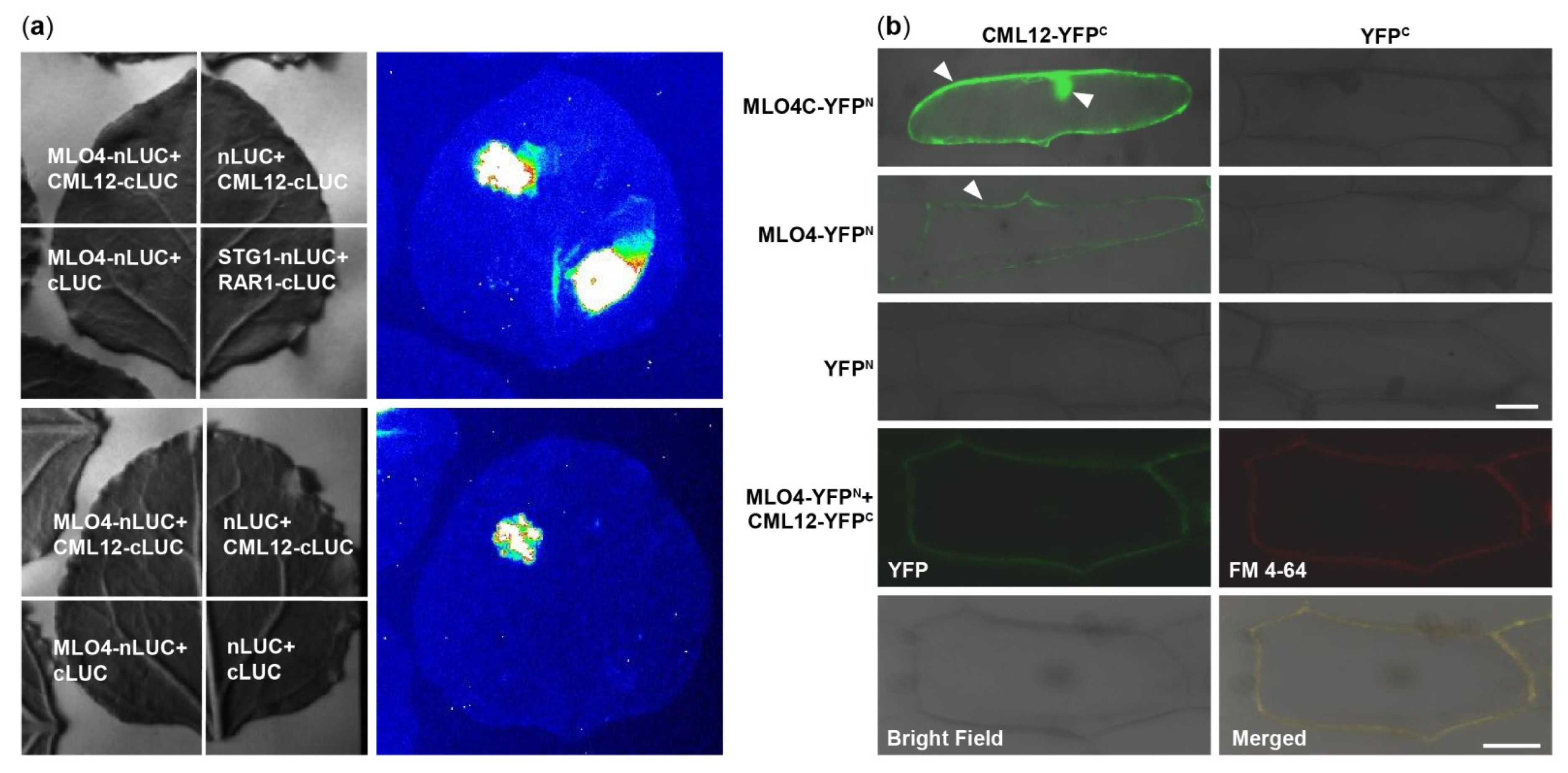
2.3. LCI and BiFC Assays Further Demonstrated That MLO4 Interacted with CML12
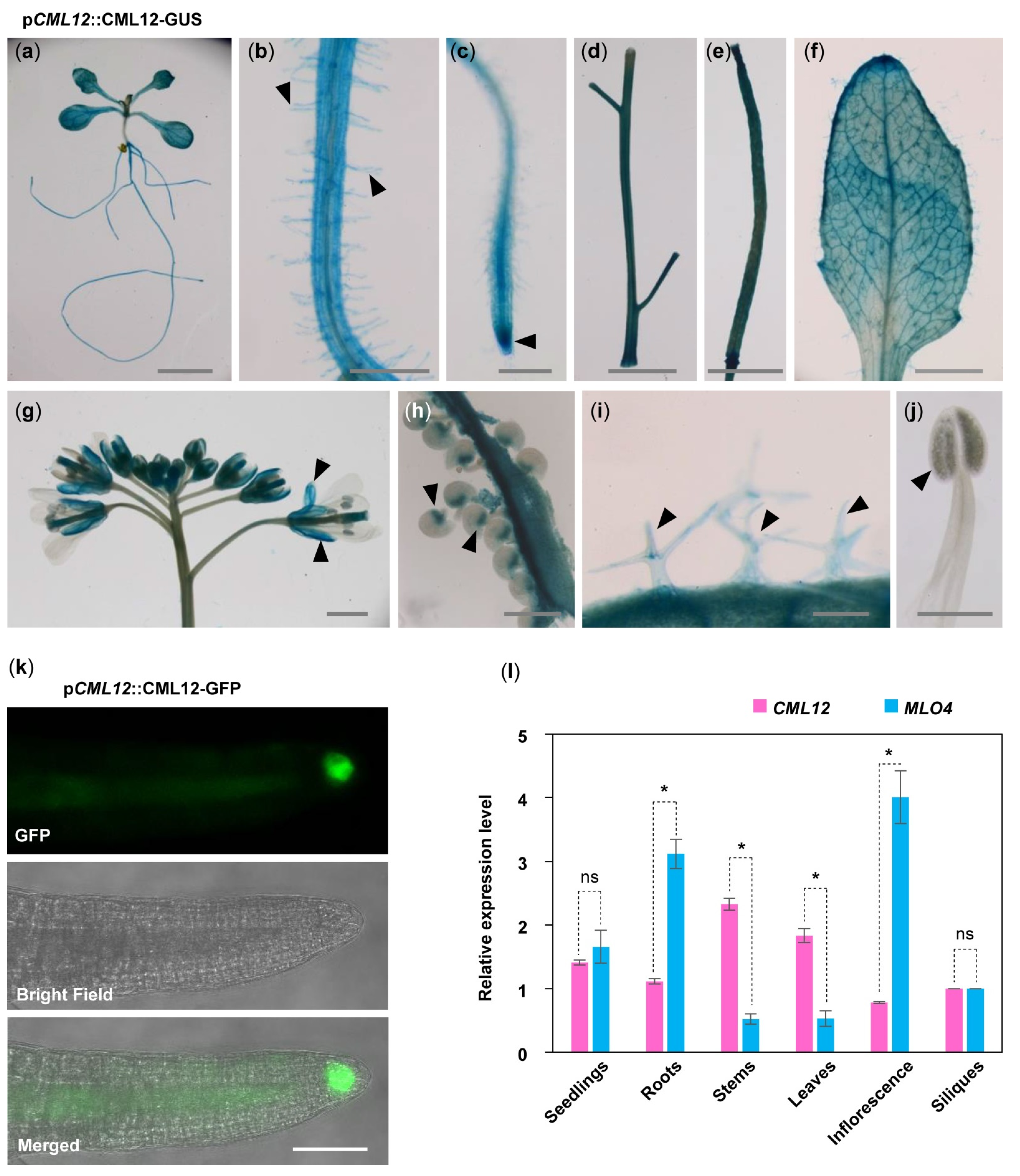
2.4. MLO4 and CML12 Displayed a Similar Expression Pattern
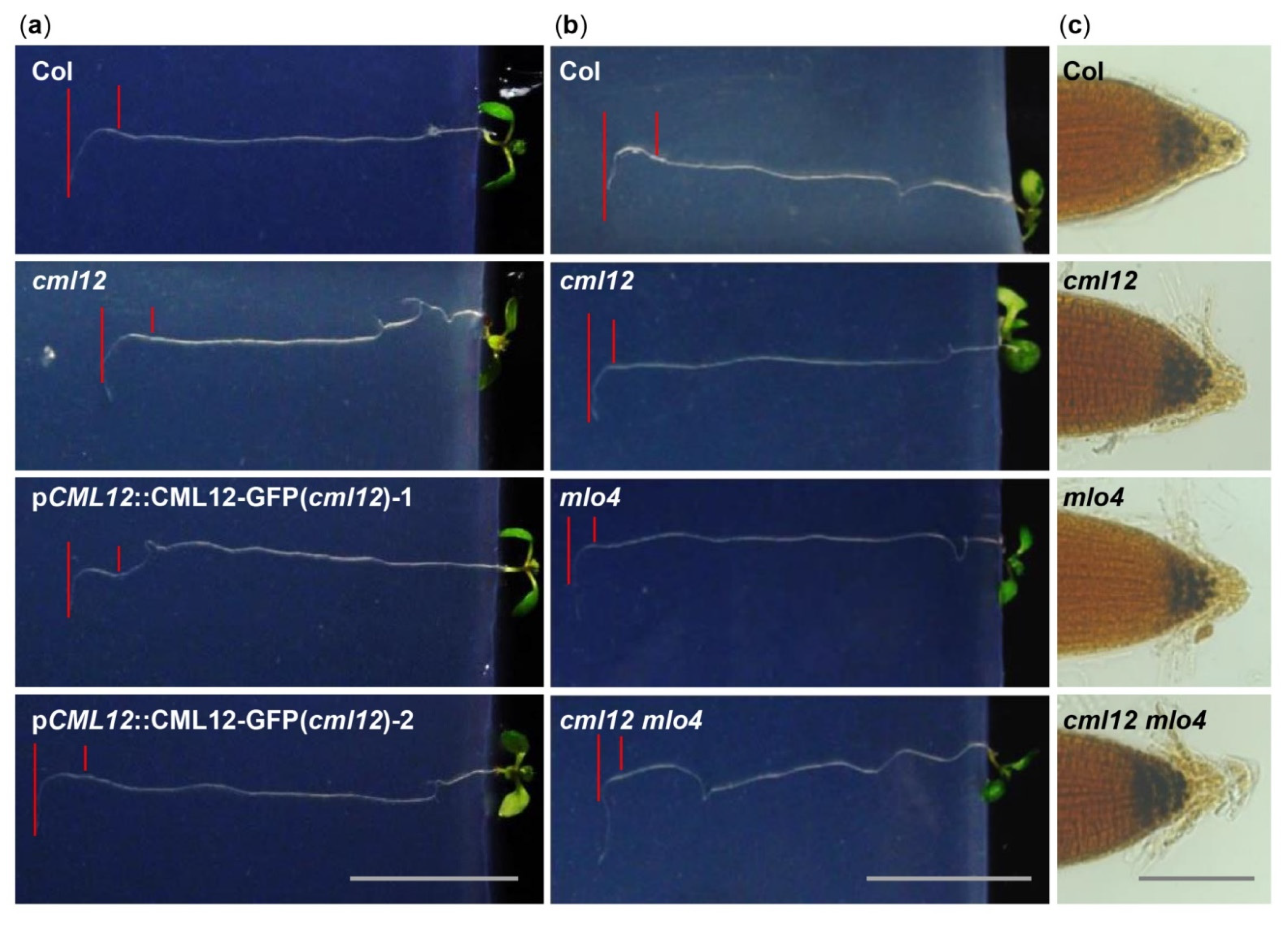
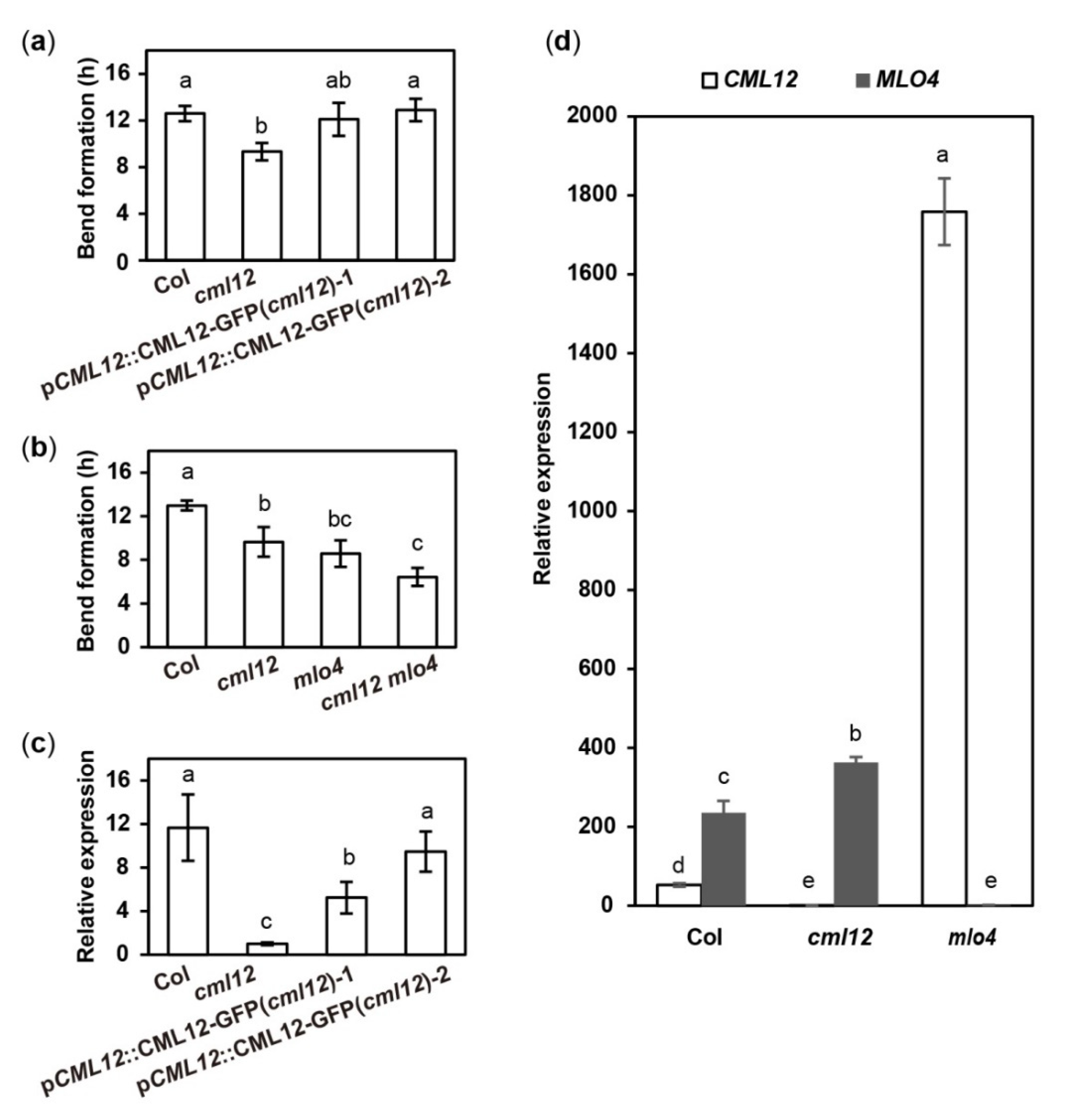
2.5. The mlo4 and cml12 Mutants Shared Similar Phenotypes
2.6. Mutation in MLO4 Affected the Expression of CML12
3. Discussion
3.1. MLO4 and CML12 Function Coordinately in Root Thigmomorphogenesis
3.2. The Conservative Amino Acids of CaMBDs and C-terminal Sequence of MLO Protein May Determine Their Specific Interacting CaM/CMLs
3.3. MLO-CaM/CML Interaction May Relate to Ca2+ Signaling and Mediate Vesicle Transport
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Root Phenotype Analysis
4.3. Real-Time PCR Assay
4.4. GUS Assay for Expression Pattern
4.5. Subcellular Localization of CML12 Protein
4.6. Yeasts Two Hybrid (Y2H) Assay
4.7. Luciferase Complementation Imaging (LCI) Assay
4.8. Bimolecular Fluorescence Complementation (BiFC) Assays Luciferase Complementation Imaging
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miklis, M.; Consonni, C.; Bhat, R.A.; Lipka, V.; Schulze-Lefert, P.; Panstruga, R. Barley MLO modulates actin-dependent and actin-independent antifungal defense pathways at the cell periphery. Plant Physiol. 2007, 144, 1132–1143. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Garcia, J.; Kusch, S.; Panstruga, R. Magical mystery tour: MLO proteins in plant immunity and beyond. New Phytol. 2014, 204, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Bidzinski, P.; Noir, S.; Shahi, S.; Reinstadler, A.; Gratkowska, D.M.; Panstruga, R. Physiological characterization and genetic modifiers of aberrant root thigmomorphogenesis in mutants of Arabidopsis thaliana MILDEW LOCUS O genes. Plant Cell Environ. 2014, 37, 2738–2753. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.C.; Jones, D.S.; Dino, A.J.; Cejda, N.I.; Yuan, J.; Willoughby, A.C.; Kessler, S.A. Arabidopsis thaliana MLO genes are expressed in discrete domains during reproductive development. Plant Reprod. 2017, 30, 185–195. [Google Scholar] [CrossRef]
- Kusch, S.; Pesch, L.; Panstruga, R. Comprehensive phylogenetic analysis sheds light on the diversity and origin of the MLO family of integral membrane proteins. Genome Biol. Evol. 2016, 8, 878–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, S.A.; Shimosato-Asano, H.; Keinath, N.F.; Wuest, S.E.; Ingram, G.; Panstruga, R.; Grossniklaus, U. Conserved molecular components for pollen tube reception and fungal invasion. Science 2010, 330, 968–971. [Google Scholar] [CrossRef]
- Jones, D.S.; Yuan, J.; Smith, B.E.; Willoughby, A.C.; Kumimoto, E.L.; Kessler, S.A. Mildew Resistance Locus O function in pollen tube reception is linked to its oligomerization and subcellular distribution. Plant Physiol. 2017, 175, 172–185. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.S.; Liang, L.; Jia, P.F.; Wang, Y.C.; Li, H.C.; Yang, W.C. Integration of ovular signals and exocytosis of a Ca2+ channel by MLOs in pollen tube guidance. Nat. Plant 2020, 6, 143–153. [Google Scholar] [CrossRef]
- Consonni, C.; Humphry, M.E.; Hartmann, H.A.; Livaja, M.; Durner, J.; Westphal, L.; Vogel, J.; Lipka, V.; Kemmerling, B.; Schulze-Lefert, P.; et al. Conserved requirement for a plant host cell protein in powdery mildew pathogenesis. Nat. Genet. 2006, 38, 716–720. [Google Scholar] [CrossRef]
- Cui, F.; Wu, H.; Safronov, O.; Zhang, P.; Kumar, R.; Kollist, H.; Salojarvi, J.; Panstruga, R.; Overmyer, K. Arabidopsis MLO2 is a negative regulator of sensitivity to extracellular reactive oxygen species. Plant Cell Environ. 2018, 41, 782–796. [Google Scholar] [CrossRef]
- Chen, Z.; Noir, S.; Kwaaitaal, M.; Hartmann, H.A.; Wu, M.J.; Mudgil, Y.; Sukumar, P.; Muday, G.; Panstruga, R.; Jones, A.M. Two seven-transmembrane domain MILDEW RESISTANCE LOCUS O proteins cofunction in Arabidopsis root thigmomorphogenesis. Plant Cell 2009, 21, 1972–1991. [Google Scholar] [CrossRef] [Green Version]
- Panstruga, R.; Schulze-Lefert, P. Corruption of host seven-transmembrane proteins by pathogenic microbes: A common theme in animals and plants? Microbes Infect. 2003, 5, 429–437. [Google Scholar] [CrossRef]
- La Verde, V.; Dominici, P.; Astegno, A. Towards understanding plant calcium signaling through calmodulin-like proteins: A biochemical and structural perspective. Int. J. Mol. Sci. 2018, 19, 1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.C.; Panstruga, R.; Elliott, C.; Muller, J.; Devoto, A.; Yoon, H.W.; Park, H.C.; Cho, M.J.; Schulze-Lefert, P. Calmodulin interacts with MLO protein to regulate defence against mildew in barley. Nature 2002, 416, 447–451. [Google Scholar] [CrossRef] [PubMed]
- McCormack, E.; Tsa, Y.C.; Braam, J. Handling calcium signaling: Arabidopsis CaMs and CMLs. Trends Plant Sci. 2005, 10, 383–389. [Google Scholar] [CrossRef]
- Perochon, A.; Aldon, D.; Galaud, J.P.; Ranty, B. Calmodulin and calmodulin-like proteins in plant calcium signaling. Biochimie 2011, 93, 2048–2053. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, R.; Singh, A.; Chattopadhyay, S. Calmodulin7 plays an important role as transcriptional regulator in Arabidopsis seedling development. Plant Cell 2008, 20, 1747–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, N.; Maurya, J.P.; Senapati, D.; Gangappa, S.N.; Chattopadhyay, S. Arabidopsis CAM7 and HY5 physically interact and directly bind to the HY5 promoter to regulate its expression and thereby promote photomorphogenesis. Plant Cell 2014, 26, 1036–1052. [Google Scholar] [CrossRef] [Green Version]
- Landoni, M.; De Francesco, A.; Galbiati, M.; Tonelli, C. A loss-of-function mutation in Calmodulin2 gene affects pollen germination in Arabidopsis thaliana. Plant Mol. Biol. 2010, 74, 235–247. [Google Scholar] [CrossRef]
- Dobney, S.; Chiasson, D.; Lam, P.; Smith, S.P.; Snedden, W.A. The calmodulin-related calcium sensor CML42 plays a role in trichome branching. J. Biol. Chem. 2009, 284, 31647–31657. [Google Scholar] [CrossRef] [Green Version]
- Delk, N.A.; Johnson, K.A.; Chowdhury, N.I.; Braam, J. CML24, regulated in expression by diverse stimuli, encodes a potential Ca2+ sensor that functions in responses to abscisic acid, daylength, and ion stress. Plant Physiol. 2005, 139, 240–253. [Google Scholar] [CrossRef] [Green Version]
- Tsai, Y.C.; Delk, N.A.; Chowdhury, N.I.; Braam, J. Arabidopsis potential calcium sensors regulate nitric oxide levels and the transition to flowering. Plant Signal. Behav. 2007, 2, 446–454. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Zhou, R.G.; Gao, Y.J.; Zheng, S.Z.; Xu, P.; Zhang, S.Q.; Sun, D.Y. Molecular and genetic evidence for the key role of AtCaM3 in heat-shock signal transduction in Arabidopsis. Plant Physiol. 2009, 149, 1773–1784. [Google Scholar] [CrossRef] [Green Version]
- Chu, M.; Li, J.; Zhang, J.; Shen, S.; Li, C.; Gao, Y.; Zhang, S. AtCaM4 interacts with a Sec14-like protein, PATL1, to regulate freezing tolerance in Arabidopsis in a CBF-independent manner. J. Exp. Bot. 2018, 69, 5241–5253. [Google Scholar] [CrossRef] [Green Version]
- Magnan, F.; Ranty, B.; Charpenteau, M.; Sotta, B.; Galaud, J.P.; Aldon, D. Mutations in AtCML9, a calmodulin-like protein from Arabidopsis thaliana, alter plant responses to abiotic stress and abscisic acid. Plant J. Cell Mol. Biol. 2008, 56, 575–589. [Google Scholar] [CrossRef]
- Wu, X.; Qiao, Z.; Liu, H.; Acharya, B.R.; Li, C.; Zhang, W. CML20, an Arabidopsis calmodulin-like protein, negatively regulates guard cell ABA signaling and drought stress tolerance. Front. Plant Sci. 2017, 8, 824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Perez, M.; Aldon, D.; Galaud, J.P. Respective contribution of CML8 and CML9, two Arabidopsis calmodulin-like proteins, to plant stress responses. Plant Signal. Behav. 2018, 12, e1322246. [Google Scholar] [CrossRef]
- Takabatake, R.; Karita, E.; Seo, S.; Mitsuhara, I.; Kuchitsu, K.; Ohashi, Y. Pathogen-induced calmodulin isoforms in basal resistance against bacterial and fungal pathogens in tobacco. Plant Cell Physiol. 2007, 48, 414–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiasson, D.; Ekengren, S.K.; Martin, G.B.; Dobney, S.L.; Snedden, W.A. Calmodulin-like proteins from Arabidopsis and tomato are involved in host defense against Pseudomonas syringae pv. tomato. Plant Mol. Biol. 2005, 58, 887–897. [Google Scholar] [CrossRef]
- Vitha, S.; Yang, M.; Sack, F.D.; Kiss, J.Z. Gravitropism in the starch excess mutant of Arabidopsis thaliana. Am. J. Bot. 2007, 94, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, L.; Squires, S.; Bisgrove, S.R. The microtubule associated protein END BINDING 1 represses root responses to mechanical cues. Plant Sci. 2012, 187, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Barlow, P.W. Gravity perception in plants: A multiplicity of systems derived by evolution? Plant Cell Environ. 1995, 18, 951–962. [Google Scholar] [CrossRef]
- Blancaflor, E.B.; Fasano, J.M.; Gilroy, S. Mapping the functional roles of cap cells in the response of Arabidopsis primary roots to gravity. Plant Physiol. 1998, 116, 213–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsugeki, R.; Fedoroff, N.V. Genetic ablation of root cap cells in Arabidopsis. Proc. Natl. Acad. Sci. USA 1999, 96, 12941–12946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, A.; Kobayashi, Y.; Hase, Y.; Watanabe, H. Positional effect of cell inactivation on root gravitropism using heavy-ion microbeams. J. Exp. Bot. 2002, 53, 683–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, K.L.; Strohm, A.K.; Masson, P. Gravity sensing and signal transduction in vascular plant primary poots. Am. J. Bot. 2013, 100, 126–142. [Google Scholar] [CrossRef]
- Fitzelle, K.; Kiss, J. Restoration of gravitropic sensitivity in starch-deficient mutants of Arabidopsis by hypergravity. J. Exp. Bot. 2001, 52, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Sistrunk, M.L.; Antosiewicz, D.M.; Purugganan, M.M.; Braam, J. Arabidopsis TCH3 encodes a novel Ca2+ binding protein and shows environmentally induced and tissue-specific regulation. Plant Cell 1994, 6, 1553–1565. [Google Scholar] [CrossRef] [Green Version]
- Stein, M.; Somerville, S.C. MLO, a novel modulator of plant defenses and cell death, binds calmodulin. Trends Plant Sci. 2002, 7, 379–380. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Hartmann, H.A.; Wu, M.J.; Friedman, E.J.; Chen, J.G.; Pulley, M.; Schulze-Lefert, P.; Panstruga, R.; Jones, A.M. Expression analysis of the AtMLO gene family encoding plant-specific seven-transmembrane domain proteins. Plant Mol. Biol. 2006, 60, 583–597. [Google Scholar] [CrossRef] [Green Version]
- Devoto, A.; Piffanelli, P.; Nilsson, I.; Wallin, E.; Panstruga, R.; Von Heijne, G.; Schulze-Lefert, P. Topology, subcellular localization, and sequence diversity of the Mlo family in plants. J. Biol. Chem. 1999, 274, 34993–35004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devoto, A.; Hartmann, H.A.; Piffanelli, P.; Elliott, C.; Simmons, C.; Taramino, G.; Goh, C.S.; Cohen, F.E.; Emerson, B.C.; Schulze-Lefert, P.; et al. Molecular phylogeny and evolution of the plant specific seven-transmembrane MLO family. J. Mol. Evol. 2003, 56, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Diao, M.; Qu, X.L.; Huang, S.J. Calcium imaging in Arabidopsis pollen cells using G-CaMP5. J. Integr. Plant Biol. 2018, 60, 897–906. [Google Scholar] [CrossRef] [Green Version]
- Ngo, Q.A.; Vogler, H.; Lituiev, D.S.; Nestorova, A.; Grossniklaus, U. A calcium dialog mediated by the FERONIA signal transduction pathway controls plant sperm delivery. Dev. Cell 2014, 29, 491–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denninger, P.; Bleckmann, A.; Lausser, A.; Vogler, F.; Ott, T.; Ehrhardt, D.W.; Frommer, W.B.; Sprunck, S.; Dresselhaus, T.; Grossmann, G. Male–female communication triggers calcium signatures during fertilization in Arabidopsis. Nat. Commun. 2014, 5, 4645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitão, N.; Dangeville, P.; Carter, R.; Charpentier, M. Nuclear calcium signatures are associated with root development. Nat. Commun. 2019, 10, 4865. [Google Scholar] [CrossRef]
- Brost, C.; Studtrucker, T.; Reimann, R.; Denninger, P.; Czekalla, J.; Krebs, M.; Fabry, B.; Schumacher, K.; Grossmann, G.; Dietrich, P. Multiple cyclic nucleotide-gated channels coordinate calcium oscillations and polar growth of root hairs. Plant J. 2019, 99, 910–923. [Google Scholar] [CrossRef]
- Hamamura, Y.; Nishimaki, M.; Takeuchi, H.; Geitmann, A.; Kurihara1, D.; Higashiyama, T. Live imaging of calcium spikes during double fertilization in Arabidopsis. Nat. Commun. 2014, 5, 4722. [Google Scholar] [CrossRef] [Green Version]
- Day, I.S.; Reddy, V.S.; Ali, G.S.; Reddy, A.S.N. Analysis of EF-hand-containing proteins in Arabidopsis. Genome Biol. 2002, 3, 1–24. [Google Scholar] [CrossRef]
- Wang, L.; Wang, W.; Wang, Y.Q.; Liu, Y.Y.; Wang, J.X.; Zhang, X.Q.; Ye, D.; Chen, L.Q. Arabidopsis galacturonosyltransferase (GAUT) 13 and GAUT14 have redundant functions in pollen tube growth. Mol. Plant 2013, 6, 1131–1148. [Google Scholar] [CrossRef] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Ye, D.; Xu, J.; Sundaresan, V. The SPOROCYTELESS gene of Arabidopsis is required for initiation of sporogenesis and encodes a novel nuclear protein. Gene Dev. 1999, 13, 2108–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Chu, L.C.; Liang, Y.; Zhang, X.Q.; Chen, L.Q.; Ye, D. The Arabidopsis CrRLK1L protein kinases BUPS1 and BUPS2 are required for normal growth of pollen tubes in the pistil. Plant J. 2018, 95, 474–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Zou, Y.; Shang, Y.; Lin, H.; Wang, Y.; Cai, R.; Tang, X.; Zhou, J. Firefly luciferase complementation imaging assay for protein-protein interactions in plants. Plant Physiol. 2007, 146, 368–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, D.; Wang, C.; He, J.; Liao, H.; Duan, Y.; Zhu, Z.; Guo, Y.; Chen, Z.; Gong, Z. A plasma membrane receptor kinase, GHR1, mediates abscisic acid- and hydrogen peroxide-regulated stomatal movement in Arabidopsis. Plant Cell 2012, 24, 2546–2561. [Google Scholar] [CrossRef] [Green Version]
- Gookin, E.; Assmann, S.M. Significant reduction of BiFC non-specific assembly facilitates in planta assessment of heterotrimeric G-protein interactors. Plant J. 2014, 80, 553–567. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MLOs | CaM/CML Proteins that Interacted with Corresponding MLO Proteins |
|---|---|
| MLO1C | CaM2, CML29 |
| MLO2C | CML9, CML18 |
| MLO3C | CML18, CML20, CML23, CML26, CML32, CML40, CML44 |
| MLO4C | CML12, CML40, CML44 |
| MLO5C | CML18, CML26, CML40, CML44 |
| MLO6C | CML8, CML10, CML20, CML26, CML73, CML40, CML44, CML49 |
| MLO7C | CML8, CML9, CML10, CML11, CML12, CML26, CML30, CML37, CML38, CML40, CML44 |
| MLO8C | CML8, CML9, CML23, CML26, CML37, CML40, CML44 |
| MLO9C | CML10, CML42, CML44, CML49 |
| MLO10C | CML8, CML9, CML10, CML26, CML37, CML44 |
| MLO11C | CML47 |
| MLO12C | CML10, CML12, CML30, CML35, CML36, CML37, CML44 |
| MLO13C | CML32, CML40, CML41 |
| MLO14C | CML17, CML18, CML49 |
| MLO15C | CML10, CML44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, L.; Zhang, X.-Q.; Ye, D.; Chen, L.-Q. The Mildew Resistance Locus O 4 Interacts with CaM/CML and Is Involved in Root Gravity Response. Int. J. Mol. Sci. 2021, 22, 5962. https://doi.org/10.3390/ijms22115962
Zhu L, Zhang X-Q, Ye D, Chen L-Q. The Mildew Resistance Locus O 4 Interacts with CaM/CML and Is Involved in Root Gravity Response. International Journal of Molecular Sciences. 2021; 22(11):5962. https://doi.org/10.3390/ijms22115962
Chicago/Turabian StyleZhu, Lei, Xue-Qin Zhang, De Ye, and Li-Qun Chen. 2021. "The Mildew Resistance Locus O 4 Interacts with CaM/CML and Is Involved in Root Gravity Response" International Journal of Molecular Sciences 22, no. 11: 5962. https://doi.org/10.3390/ijms22115962
APA StyleZhu, L., Zhang, X. -Q., Ye, D., & Chen, L. -Q. (2021). The Mildew Resistance Locus O 4 Interacts with CaM/CML and Is Involved in Root Gravity Response. International Journal of Molecular Sciences, 22(11), 5962. https://doi.org/10.3390/ijms22115962