
The Enigmatic Metallothioneins: A Case of Upward-Looking Research




{kind=link}
{kind=link}
Abstract
1. Introduction
2. Classifying Vallee’s Experiments
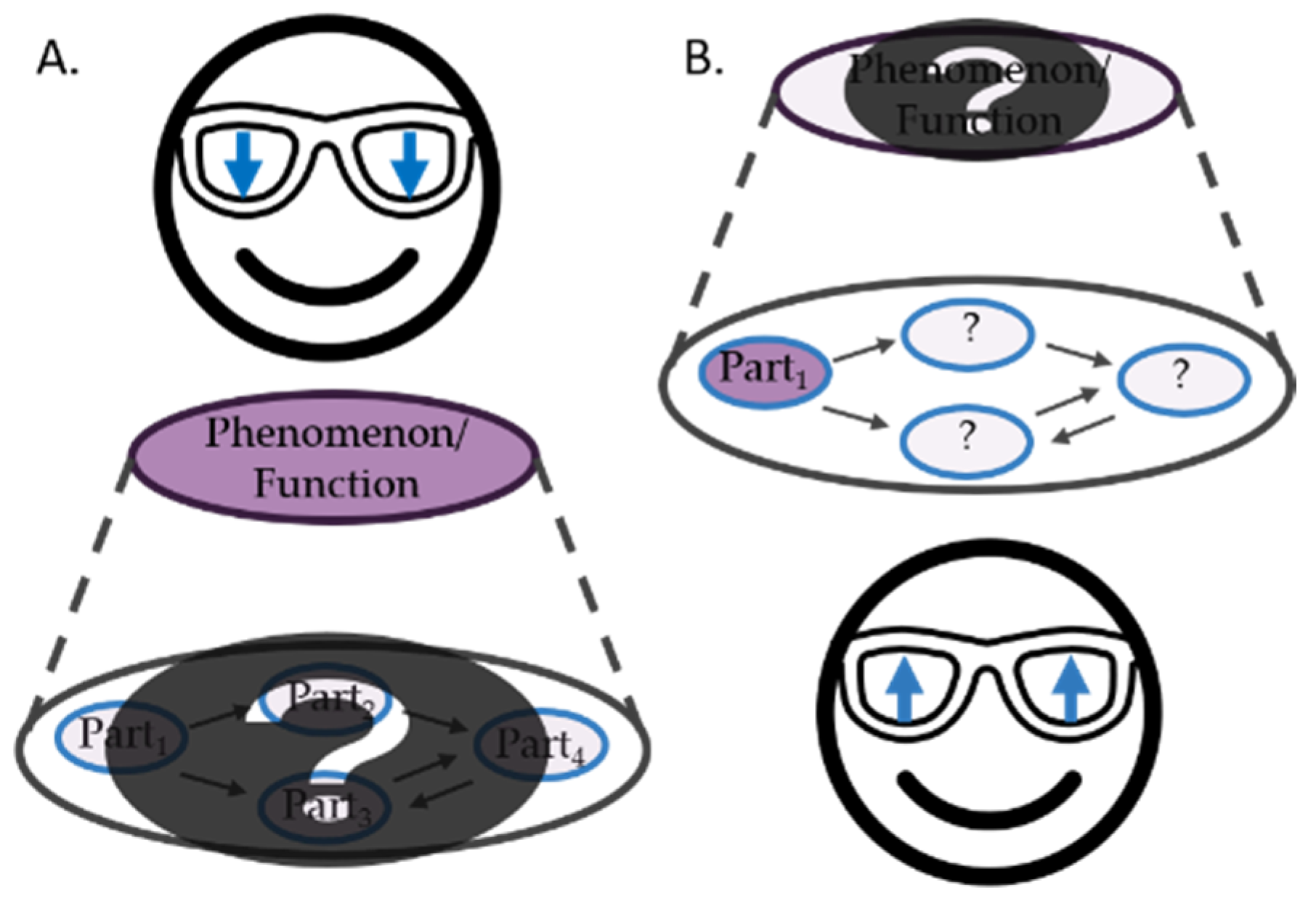
2.1. Downward-Looking versus Upward-Looking
2.2. Interventions versus Mere Interactions
2.3. Hypothesis-Testing versus Exploratory Research
3. The Enigmatic Role of Vertebrate MTs
3.1. Metal Detoxification or Toxic Metal Metabolism
3.2. Metal Homeostasis or Essential Metal Metabolism
3.3. Protection from OS or Antioxidant Scavengers
3.4. Quo Vadis?
4. The Case of Ceruloplasmin
5. Beyond One-to-One Mappings
5.1. Multiple Realization, Robustness, and Moonlighting
5.2. Evolutionary Effects: Vestiges, Exaptation, and Convergent Evolution
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vallee, B.L.; Hoch, F.L. Zinc, a component of yeast alcohol dehydrogenase. Proc. Natl. Acad. Sci. USA 1955, 41, 327–338. [Google Scholar] [CrossRef]
- Vallee, B.L. Metallothionein: Historical Review and Perspectives. Exp. Supply 1979, 34, 19–40. [Google Scholar] [CrossRef]
- Margoshes, M.; Vallee, B.L. A cadmium protein from equine kidney cortex. J. Am. Chem. Soc. 1957, 79, 4813–4814. [Google Scholar] [CrossRef]
- Kägi, J. Piscator. In Metallothionein, Proceedings of the First International Meeting on Metallothionein and Other Low Molecular Weight Metal-Binding Proteins, Zürich, Switzerland, 17–22 July 1978; Springer: Basel, Switzerland, 2013; ISBN 978-3-0348-6493-0. [Google Scholar]
- Kagi, J.H.; Valee, B.L. Metallothionein: A Cadmium- and Zinc-Containing Protein from Equine Renal Cortex. J. Biol. Chem. 1960, 235, 3460–3465. [Google Scholar] [CrossRef]
- Vallee, B.L. Implications and Inferences of Metallothionein Structure. In Metallothionein II, Proceedings of the Second International Meeting on Metallothionein and Other Low Molecular Weight Metalbinding Proteins, Zürich, Switzerland, 21–24 August 1985; Kägi, J.H.R., Kojima, Y., Eds.; Experientia Supplementum; Birkhäuser: Basel, Switzerland, 1987; pp. 5–16. ISBN 978-3-0348-6784-9. [Google Scholar]
- Palacios, O.; Atrian, S.; Capdevila, M. Zn- and Cu-Thioneins: A Functional Classification for Metallothioneins? J. Biol. Inorg. Chem. 2011, 16, 991–1009. [Google Scholar] [CrossRef] [PubMed]
- Dunn, M.A.; Blalock, T.L.; Cousins, R.J. Metallothionein. Proc. Soc. Exp. Biol. Med. 1987, 185, 107–119. [Google Scholar] [CrossRef]
- Davis, S.R.; Cousins, R.J. Metallothionein Expression in Animals: A Physiological Perspective on Function. J. Nutr. 2000, 130, 1085–1088. [Google Scholar] [CrossRef] [PubMed]
- Koh, J.-Y.; Lee, S.-J. Metallothionein-3 as a Multifunctional Player in the Control of Cellular Processes and Diseases. Mol. Brain 2020, 13, 116. [Google Scholar] [CrossRef]
- Rono, J.K.; Le Wang, L.; Wu, X.C.; Cao, H.W.; Zhao, Y.N.; Khan, I.U.; Yang, Z.M. Identification of a New Function of Metallothionein-like Gene OsMT1e for Cadmium Detoxification and Potential Phytoremediation. Chemosphere 2021, 265, 129136. [Google Scholar] [CrossRef]
- Jacob, C.; Maret, W.; Vallee, B.L. Control of Zinc Transfer between Thionein, Metallothionein, and Zinc Proteins. Proc. Natl. Acad. Sci. USA 1998, 95, 3489–3494. [Google Scholar] [CrossRef]
- Binz, P.-A.; Kägi, J.H.R. Metallothionein: Molecular evolution and classification. In Metallothionein I; Klaassen, C.D., Ed.; Advances in Life Sciences; Birkhäuser: Basel, Switzerland, 1999; pp. 7–13. ISBN 978-3-0348-8847-9. [Google Scholar]
- Freisinger, E. Structural Features Specific to Plant Metallothioneins. J. Biol. Inorg. Chem. 2011, 16, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Krężel, A.; Maret, W. The Functions of Metamorphic Metallothioneins in Zinc and Copper Metabolism. Int. J. Mol. Sci. 2017, 18, 1237. [Google Scholar] [CrossRef] [PubMed]
- Palmiter, R.D. The Elusive Function of Metallothioneins. Proc. Natl. Acad. Sci. USA 1998, 95, 8428–8430. [Google Scholar] [CrossRef] [PubMed]
- Kästner, L. Integration and the Mechanistic Triad: Producing, Underlying and Maintaining Mechanistic Explanations. In Neural Mechanisms: New Challenges in the Philosophy of Neuroscience; Studies in Brain and Mind, Calzavarini, F., Viola, M., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 337–361. ISBN 978-3-030-54092-0. [Google Scholar]
- Abdin, A.Y.; Jacob, C.; Kästner, L. Disambiguating “Mechanisms” in Pharmacy: Lessons from Mechanist Philosophy of Science. Int. J. Environ. Res. Public Health 2020, 17, 1833. [Google Scholar] [CrossRef]
- Craver, C.F.; Darden, L. Search of Mechanisms: Discoveries Across the Life Sciences; Chicago Press: Chicago, IL, USA, 2013; ISBN 978-0-226-03979-4. [Google Scholar]
- Bechtel, W.; Richardson, R.C. Discovering Complexity: Decomposition and Localization as Strategies in Scientific Research; The MIT Press: Cambridge, MA, USA, 2010; ISBN 978-0-262-51473-6. [Google Scholar]
- Bechtel, W.; Abrahamsen, A. Decomposing, Recomposing, and Situating Circadian Mechanisms: Three Tasks in Developing Mechanistic Explanations; De Gruyter: Berlin, Germany, 2013; pp. 177–190. ISBN 978-3-11-032885-1. [Google Scholar]
- Bush, K. Past and Present Perspectives on β-Lactamases. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef]
- Abraham, E.P.; Chain, E. An Enzyme from Bacteria Able to Destroy Penicillin. Nature 1940, 146, 837. [Google Scholar] [CrossRef]
- Abraham, E.P.; Chain, E.; Fletcher, C.M.; Gardner, A.D.; Heatley, N.G.; Jennings, A.M.; Florey, H.W. Further observations on penicillin. Lancet 1941, 2, 177–189. Available online: https://www.jameslindlibrary.org/abraham-ep-chain-e-fletcher-cm-gardner-ad-heatley-ng-jennings-am-florey-hw-1941/ (accessed on 13 November 2020). [CrossRef]
- Kirby, W.M.M. Extraction of a Highly Potent Penicillin Inactivator from Penicillin Resistant Staphylococci. Science 1944, 99, 452–453. [Google Scholar] [CrossRef]
- Woodward, J. Making Things Happen: A Theory of Causal Explanation; Oxford University Press: New York, NY, USA, 2003; ISBN 978-0-19-518953-7. [Google Scholar]
- Kästner, L.; Andersen, L.M. Intervening into Mechanisms: Prospects and Challenges. Philos. Compass 2018, 13, e12546. [Google Scholar] [CrossRef]
- Craver, C.F. Explaining the Brain: Mechanisms and the Mosaic Unity of Neuroscience; Oxford University Press: Oxford, UK; New York, NY, USA, 2007; ISBN 978-0-19-929931-7. [Google Scholar]
- Kästner, L. Philosophy of Cognitive Neuroscience: Causal Explanations, Mechanisms and Experimental Manipulations; Walter de Gruyter GmbH & Co KG: Berlin, Germany, 2017; Volume 37. [Google Scholar]
- Hacking, I. Representing and Intervening: Introductory Topics in the Philosophy of Natural Science; Cambridge University Press: Cambridge, MA, USA, 1983; ISBN 978-0-521-28246-8. [Google Scholar]
- Haueis, P.; Slaby, J. Brain in the shell. In Neuroscience and Critique: Exploring the Limits of the Neurological Turn; Routledge: London, UK, 2015; p. 117. [Google Scholar]
- Steinle, F. Entering New Fields: Exploratory Uses of Experimentation. Philos. Sci. 1997, 64, S65–S74. [Google Scholar] [CrossRef]
- Waters, C.K. Causes That Make a Difference. Available online: https://www.pdcnet.org/pdc/bvdb.nsf/purchase?openform&fp=jphil&id=jphil_2007_0104_0011_0551_0579 (accessed on 25 February 2021).
- O’Malley, M.A.; Dupré, J. Size Doesn’t Matter: Towards a More Inclusive Philosophy of Biology. Biol. Philos. 2007, 22, 155–191. [Google Scholar] [CrossRef]
- Wurtz, R.H. Recounting the Impact of Hubel and Wiesel. J. Physiol. 2009, 587, 2817–2823. [Google Scholar] [CrossRef]
- Steinle, F. Concept formation and the limits of justification: “discovering” the two electricities. In Revisiting Discovery and Justification: Historical and Philosophical Perspectives on the Context Distinction; Archimedes; Schickore, J., Steinle, F., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 183–195. ISBN 978-1-4020-4251-5. [Google Scholar]
- Kästner, L. Explaining Phenomena, Discovering Mechanisms; Talk at the Cognition Academy; Max Planck School of Cognition: Berlin, Germany, 2021. [Google Scholar]
- Suzuki, K.T.; Imura, N.; Kimura, M. Metallothionein III: Biological Roles and Medical Implications; Birkhäuser: Basel, Switzerland, 1993. [Google Scholar]
- Robbins, A.H.; McRee, D.E.; Williamson, M.; Collett, S.A.; Xuong, N.H.; Furey, W.F.; Wang, B.C.; Stout, C.D. Refined Crystal Structure of Cd, Zn Metallothionein at 2.0 Åresolution. J. Mol. Biol. 1991, 221, 1269–1293. [Google Scholar] [CrossRef]
- Calatayud, S.; Garcia-Risco, M.; Pedrini-Martha, V.; Eernisse, D.J.; Dallinger, R.; Palacios, Ò.; Capdevila, M.; Albalat, R. Modularity in Protein Evolution: Modular Organization and De Novo Domain Evolution in Mollusk Metallothioneins. Mol. Biol. Evol. 2021, 38, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Isani, G.; Carpenè, E. Metallothioneins, Unconventional Proteins from Unconventional Animals: A Long Journey from Nematodes to Mammals. Biomolecules 2014, 4, 435–457. [Google Scholar] [CrossRef] [PubMed]
- Blindauer, C.A. Chapter 21: Metallothioneins. In Binding, Transport and Storage of Metal Ions in Biological Cells; The Royal Society of Chemistry: Cambridge, UK, 2014; pp. 606–665. [Google Scholar]
- Vašák, M.; Meloni, G. Chemistry and Biology of Mammalian Metallothioneins. J. Biol. Inorg. Chem. 2011, 16, 1067. [Google Scholar] [CrossRef] [PubMed]
- Serén, N.; Glaberman, S.; Carretero, M.A.; Chiari, Y. Molecular Evolution and Functional Divergence of the Metallothionein Gene Family in Vertebrates. J. Mol. Evol. 2014, 78, 217–233. [Google Scholar] [CrossRef]
- Laukens, D.; Waeytens, A.; Bleser, P.D.; Cuvelier, C.; Vos, M.D. Human Metallothionein Expression under Normal and Pathological Conditions: Mechanisms of Gene Regulation Based on In Silico Promoter Analysis. CRE 2009, 19. [Google Scholar] [CrossRef] [PubMed]
- García-Risco, M.; Calatayud, S.; Niederwanger, M.; Albalat, R.; Palacios, Ò.; Capdevila, M.; Dallinger, R. Two Unconventional Metallothioneins in the Apple Snail Pomacea Bridgesii Have Lost Their Metal Specificity during Adaptation to Freshwater Habitats. Int. J. Mol. Sci. 2021, 22, 95. [Google Scholar] [CrossRef]
- Palacios, Ò.; Pagani, A.; Pérez-Rafael, S.; Egg, M.; Höckner, M.; Brandstätter, A.; Capdevila, M.; Atrian, S.; Dallinger, R. Shaping Mechanisms of Metal Specificity in a Family of Metazoan Metallothioneins: Evolutionary Differentiation of Mollusc Metallothioneins. BMC Biol. 2011, 9, 4. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rafael, S.; Monteiro, F.; Dallinger, R.; Atrian, S.; Palacios, Ò.; Capdevila, M. Cantareus Aspersus Metallothionein Metal Binding Abilities: The Unspecific CaCd/CuMT Isoform Provides Hints about the Metal Preference Determinants in Metallothioneins. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2014, 1844, 1694–1707. [Google Scholar] [CrossRef]
- Dallinger, R.; Zerbe, O.; Baumann, C.; Egger, B.; Capdevila, M.; Palacios, Ò.; Albalat, R.; Calatayud, S.; Ladurner, P.; Schlick-Steiner, B.C.; et al. Metallomics Reveals a Persisting Impact of Cadmium on the Evolution of Metal-Selective Snail Metallothioneins. Metallomics 2020, 12, 702–720. [Google Scholar] [CrossRef] [PubMed]
- Jacob, C.; Maret, W.; Vallee, B.L. Ebselen, a Selenium-Containing Redox Drug, Releases Zinc from Metallothionein. Biochem. Biophys. Res. Commun. 1998, 248, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Jacob, C.; Vallee, B.L.; Fischer, E.H. Inhibitory Sites in Enzymes: Zinc Removal and Reactivation by Thionein. Proc. Natl. Acad. Sci. USA 1999, 96, 1936–1940. [Google Scholar] [CrossRef]
- Jacob, C.; Maret, W.; Vallee, B.L. Selenium Redox Biochemistry of Zinc–Sulfur Coordination Sites in Proteins and Enzymes. Proc. Natl. Acad. Sci. USA 1999, 96, 1910–1914. [Google Scholar] [CrossRef] [PubMed]
- Quaife, C.J.; Kelly, E.J.; Masters, B.A.; Brinster, R.L.; Palmiter, R.D. Ectopic Expression of Metallothionein-III Causes Pancreatic Acinar Cell Necrosis in Transgenic Mice. Toxicol. Appl. Pharmacol. 1998, 148, 148–157. [Google Scholar] [CrossRef]
- Klein, J.B.; Wang, G.-W.; Zhou, Z.; Buridi, A.; Kang, Y.J. Inhibition of Tumor Necrosis Factor-α-Dependent Cardiomyocyte Apoptosis by Metallothionein. Cardiovasc. Toxicol. 2002, 2, 209–217. [Google Scholar] [CrossRef]
- Kang, Y.J.; Li, Y.; Sun, X.; Sun, X. Antiapoptotic Effect and Inhibition of Ischemia/Reperfusion-Induced Myocardial Injury in Metallothionein-Overexpressing Transgenic Mice. Am. J. Pathol. 2003, 163, 1579–1586. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Y.; Habeebu, S.S.; Klaassen, C.D. Susceptibility of MT-Null Mice to Chronic CdCl2-Induced Nephrotoxicity Indicates That Renal Injury Is Not Mediated by the CdMT Complex. Toxicol. Sci. 1998, 46, 197–203. [Google Scholar] [CrossRef]
- Zhang, B.; Georgiev, O.; Hagmann, M.; Günes, Ç.; Cramer, M.; Faller, P.; Vasák, M.; Schaffner, W. Activity of Metal-Responsive Transcription Factor 1 by Toxic Heavy Metals and H2O2 In Vitro Is Modulated by Metallothionein. Mol. Cell Biol. 2003, 23, 8471–8485. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, B.; Harmon, P.M.; Schaffner, W.; Peterson, D.O.; Giedroc, D.P. A Novel Cysteine Cluster in Human Metal-Responsive Transcription Factor 1 Is Required for Heavy Metal-Induced Transcriptional Activation in Vivo. J. Biol. Chem. 2004, 279, 4515–4522. [Google Scholar] [CrossRef]
- Cai, L.; Li, X.-K.; Song, Y.; Cherian, M.G. Essentiality, Toxicology and Chelation Therapy of Zinc and Copper. Curr. Med. Chem. 2005, 12, 2753–2763. [Google Scholar] [CrossRef]
- Maret, W. Zinc in Cellular Regulation: The Nature and Significance of “Zinc Signals”. Int. J. Mol. Sci. 2017, 18, 2285. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Flight, R.M.; Rouchka, E.C.; Moseley, H.N.B. A Less-biased Analysis of Metalloproteins Reveals Novel Zinc Coordination Geometries. Proteins 2015, 83, 1470–1487. [Google Scholar] [CrossRef]
- Sutherland, D.E.K.; Stillman, M.J. The “Magic Numbers” of Metallothionein. Metallomics 2011, 3, 444–463. [Google Scholar] [CrossRef]
- Balamurugan, K.; Schaffner, W. 2:Regulation of Metallothionein Gene Expression. In Metallothioneins and Related Chelators; The Royal Society of Chemistry: Cambridge, UK, 2009; pp. 31–49. [Google Scholar]
- Chiaverini, N.; Ley, M.D. Protective Effect of Metallothionein on Oxidative Stress-Induced DNA Damage. Free Radic. Res. 2010, 44, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.J.; Li, G.; Saari, J.T. Metallothionein Inhibits Ischemia-Reperfusion Injury in Mouse Heart. Am. J. Physiol. Heart Circ. Physiol. 1999, 276, H993–H997. [Google Scholar] [CrossRef] [PubMed]
- Zangger, K.; Oz, G.; Otvos, J.D.; Armitage, I.M. Three-Dimensional Solution Structure of Mouse [Cd7]-Metallothionein-1 by Homonuclear and Heteronuclear NMR Spectroscopy. Protein Sci. 1999, 8, 2630–2638. [Google Scholar] [CrossRef]
- Schultze, P.; Wörgötter, E.; Braun, W.; Wagner, G.; Vašák, M.; Kägi, J.H.R.; Wüthrich, K. Conformation of [Cd7]-Metallothionein-2 from Rat Liver in Aqueous Solution Determined by Nuclear Magnetic Resonance Spectroscopy. J. Mol. Biol. 1988, 203, 251–268. [Google Scholar] [CrossRef]
- Braun, W.; Vasák, M.; Robbins, A.H.; Stout, C.D.; Wagner, G.; Kägi, J.H.; Wüthrich, K. Comparison of the NMR Solution Structure and the X-Ray Crystal Structure of Rat Metallothionein-2. Proc. Natl. Acad. Sci. USA 1992, 89, 10124–10128. [Google Scholar] [CrossRef]
- Bertini, I.; Luchinat, C.; Messori, L.; Vasak, M. Proton NMR Studies of the Cobalt(II)-Metallothionein System. J. Am. Chem. Soc. 1989, 111, 7296–7300. [Google Scholar] [CrossRef]
- Hünefeld Friedr, L. Der Chem.ismus in der Thierischen Organisation: Physiologisch-Chem.ische Untersuchungen der Materiellen Veränderungen oder des Bildungslebens im Thierischen Organismus, Insbesondere des Blutbildungsprocesses, der Natur der Blutkörperchen und Ihrer Kernchen: Ein Beitrag zur Physiologie und Heilmittellehre; Brockhaus: Leipzig, Germany, 1840. [Google Scholar]
- Edsall, J.T. Blood and Hemoglobin: The Evolution of Knowledge of Functional Adaptation in a Biochemical System. J. Hist. Biol. 1972, 5, 205–257. [Google Scholar] [CrossRef] [PubMed]
- Leçons Sur Les Effets Des Substances Toxiques et Médicamenteuses: Bernard, Claude, 1813–1878: Free Download, Borrow, and Streaming: Internet Archive. Available online: https://archive.org/details/leonssurlesef00bern/page/n11/mode/2up (accessed on 11 February 2021).
- Holmberg, C.G. On the Presence of n Laccase-like Enzyme in Nerum and Its Relation to the Copper in Serum. Acta Physiol. Scand. 1944, 8, 227–229. [Google Scholar] [CrossRef]
- Lee, G.R.; Nacht, S.; Lukens, J.N.; Cartwright, G.E. Iron Metabolism in Copper-Deficient Swine. J. Clin. Investig. 1968, 47, 2058–2069. [Google Scholar] [CrossRef]
- Hellman, N.E.; Gitlin, J.D. Ceruloplasmin Metabolism and Function. Annu. Rev. Nutr. 2002, 22, 439–458. [Google Scholar] [CrossRef] [PubMed]
- The Laboratory Diagnosis of Haemophilia. Abstract—Europe PMC. Available online: https://europepmc.org/article/med/16810890 (accessed on 13 November 2020).
- Biggs, R.; Douglas, A.S.; Macfarlane, R.G.; Dacie, J.V.; Pitney, W.R.; Merskey, C.; O’Brien, J.R. Christmas Disease. Br. Med. J. 1952, 2, 1378–1382. [Google Scholar] [CrossRef]
- Félix, M.-A.; Barkoulas, M. Pervasive Robustness in Biological Systems. Nat. Rev. Genet. 2015, 16, 483–496. [Google Scholar] [CrossRef]
- Bedner, P.; Steinhäuser, C.; Theis, M. Functional Redundancy and Compensation among Members of Gap Junction Protein Families? Biochim. Biophys. Acta (BBA) Biomembr. 2012, 1818, 1971–1984. [Google Scholar] [CrossRef]
- Manthey, D.; Banach, K.; Desplantez, T.; Lee, C.G.; Kozak, C.A.; Traub, O.; Weingart, R.; Willecke, K. Intracellular Domains of Mouse Connexin26 and -30 Affect Diffusional and Electrical Properties of Gap Junction Channels. J. Membr. Biol. 2001, 181, 137–148. [Google Scholar] [CrossRef]
- Plum, A.; Hallas, G.; Magin, T.; Dombrowski, F.; Hagendorff, A.; Schumacher, B.; Wolpert, C.; Kim, J.-S.; Lamers, W.H.; Evert, M.; et al. Unique and Shared Functions of Different Connexins in Mice. Curr. Biol. 2000, 10, 1083–1091. [Google Scholar] [CrossRef]
- Sweeny, E.A.; Singh, A.B.; Chakravarti, R.; Martinez-Guzman, O.; Saini, A.; Haque, M.M.; Garee, G.; Dans, P.D.; Hannibal, L.; Reddi, A.R.; et al. Glyceraldehyde-3-Phosphate Dehydrogenase Is a Chaperone That Allocates Labile Heme in Cells. J. Biol. Chem. 2018, 293, 14557–14568. [Google Scholar] [CrossRef] [PubMed]
- Kosova, A.A.; Khodyreva, S.N.; Lavrik, O.I. Role of Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) in DNA Repair. Biochemistry (Moscow) 2017, 82, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Tarze, A.; Deniaud, A.; Le Bras, M.; Maillier, E.; Molle, D.; Larochette, N.; Zamzami, N.; Jan, G.; Kroemer, G.; Brenner, C. GAPDH, a Novel Regulator of the pro-Apoptotic Mitochondrial Membrane Permeabilization. Oncogene 2007, 26, 2606–2620. [Google Scholar] [CrossRef] [PubMed]
- Boradia, V.M.; Raje, M.; Raje, C.I. Protein Moonlighting in Iron Metabolism: Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH). Biochem. Soc. Trans. 2014, 42, 1796–1801. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, C.J. Protein Moonlighting: What Is It, and Why Is It Important? Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20160523. [Google Scholar] [CrossRef]
- Huberts, D.H.E.W.; van der Klei, I.J. Moonlighting Proteins: An Intriguing Mode of Multitasking. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2010, 1803, 520–525. [Google Scholar] [CrossRef]
- Jeffery, C.J. Moonlighting Proteins. Trends Biochem. Sci. 1999, 24, 8–11. [Google Scholar] [CrossRef]
- Grzmil, P.; Kim, Y.; Shamsadin, R.; Neesen, J.; Adham, I.M.; Heinlein, U.A.; Schwarzer, U.J.; Engel, W. Human Cyritestin Genes (CYRN1 and CYRN2) Are Non-Functional. Biochem. J. 2001, 357, 551–556. [Google Scholar] [CrossRef]
- Adham, I.M.; Kim, Y.; Shamsadin, R.; Heinlein, U.A.; Von Beust, G.; Mattei, M.G.; Engel, W. Molecular Cloning, Chromosomal Localization, and Expression Analysis of CYRN1 and CYRN2, Two Human Genes Coding for Cyritestin, a Sperm Protein Involved in Gamete Interaction. DNA Cell Biol. 1998, 17, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Shamsadin, R.; Adham, I.M.; Nayernia, K.; Heinlein, U.A.; Oberwinkler, H.; Engel, W. Male Mice Deficient for Germ-Cell Cyritestin Are Infertile. Biol. Reprod. 1999, 61, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- Adamo, G.M.; Lotti, M.; Tamás, M.J.; Brocca, S. Amplification of the CUP1 Gene Is Associated with Evolution of Copper Tolerance in Saccharomyces Cerevisiae. Microbiology (Read.) 2012, 158, 2325–2335. [Google Scholar] [CrossRef]
- Pedrini-Martha, V.; Köll, S.; Dvorak, M.; Dallinger, R. Cadmium Uptake, MT Gene Activation and Structure of Large-Sized Multi-Domain Metallothioneins in the Terrestrial Door Snail Alinda Biplicata (Gastropoda, Clausiliidae). Int. J. Mol. Sci. 2020, 21, 1631. [Google Scholar] [CrossRef] [PubMed]
- de Francisco, P.; Martín-González, A.; Turkewitz, A.P.; Gutiérrez, J.C. Extreme Metal Adapted, Knockout and Knockdown Strains Reveal a Coordinated Gene Expression among Different Tetrahymena Thermophila Metallothionein Isoforms. PLoS ONE 2017, 12, e0189076. [Google Scholar] [CrossRef] [PubMed]
- de Francisco, P.; Martín-González, A.; Turkewitz, A.P.; Gutiérrez, J.C. Genome Plasticity in Response to Stress in Tetrahymena Thermophila: Selective and Reversible Chromosome Amplification and Paralogous Expansion of Metallothionein Genes. Environ. Microbiol. 2018, 20, 2410–2421. [Google Scholar] [CrossRef]
- Ziller, A.; Fraissinet-Tachet, L. Metallothionein Diversity and Distribution in the Tree of Life: A Multifunctional Protein. Metallomics 2018, 10, 1549–1559. [Google Scholar] [CrossRef]
- Laudan, L. From Theories to Research Traditions. Read. Philos. Sci. 1989, 368–379. [Google Scholar]
- Šešelja, D.; Weber, E. Rationality and Irrationality in the History of Continental Drift: Was the Hypothesis of Continental Drift Worthy of Pursuit? Stud. Hist. Philos. Sci. Part A 2012, 43, 147–159. [Google Scholar] [CrossRef]
- Laudan, L. Progress and Its Problems: Toward a Theory of Scientific Growth; University of California Press: Berkeley, CA, USA, 1977. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdin, A.Y.; Jacob, C.; Kästner, L. The Enigmatic Metallothioneins: A Case of Upward-Looking Research. Int. J. Mol. Sci. 2021, 22, 5984. https://doi.org/10.3390/ijms22115984
Abdin AY, Jacob C, Kästner L. The Enigmatic Metallothioneins: A Case of Upward-Looking Research. International Journal of Molecular Sciences. 2021; 22(11):5984. https://doi.org/10.3390/ijms22115984
Chicago/Turabian StyleAbdin, Ahmad Yaman, Claus Jacob, and Lena Kästner. 2021. "The Enigmatic Metallothioneins: A Case of Upward-Looking Research" International Journal of Molecular Sciences 22, no. 11: 5984. https://doi.org/10.3390/ijms22115984
APA StyleAbdin, A. Y., Jacob, C., & Kästner, L. (2021). The Enigmatic Metallothioneins: A Case of Upward-Looking Research. International Journal of Molecular Sciences, 22(11), 5984. https://doi.org/10.3390/ijms22115984