

RETRACTED: Thymoquinone and Curcumin Defeat Aging-Associated Oxidative Alterations Induced by D-Galactose in Rats’ Brain and Heart

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
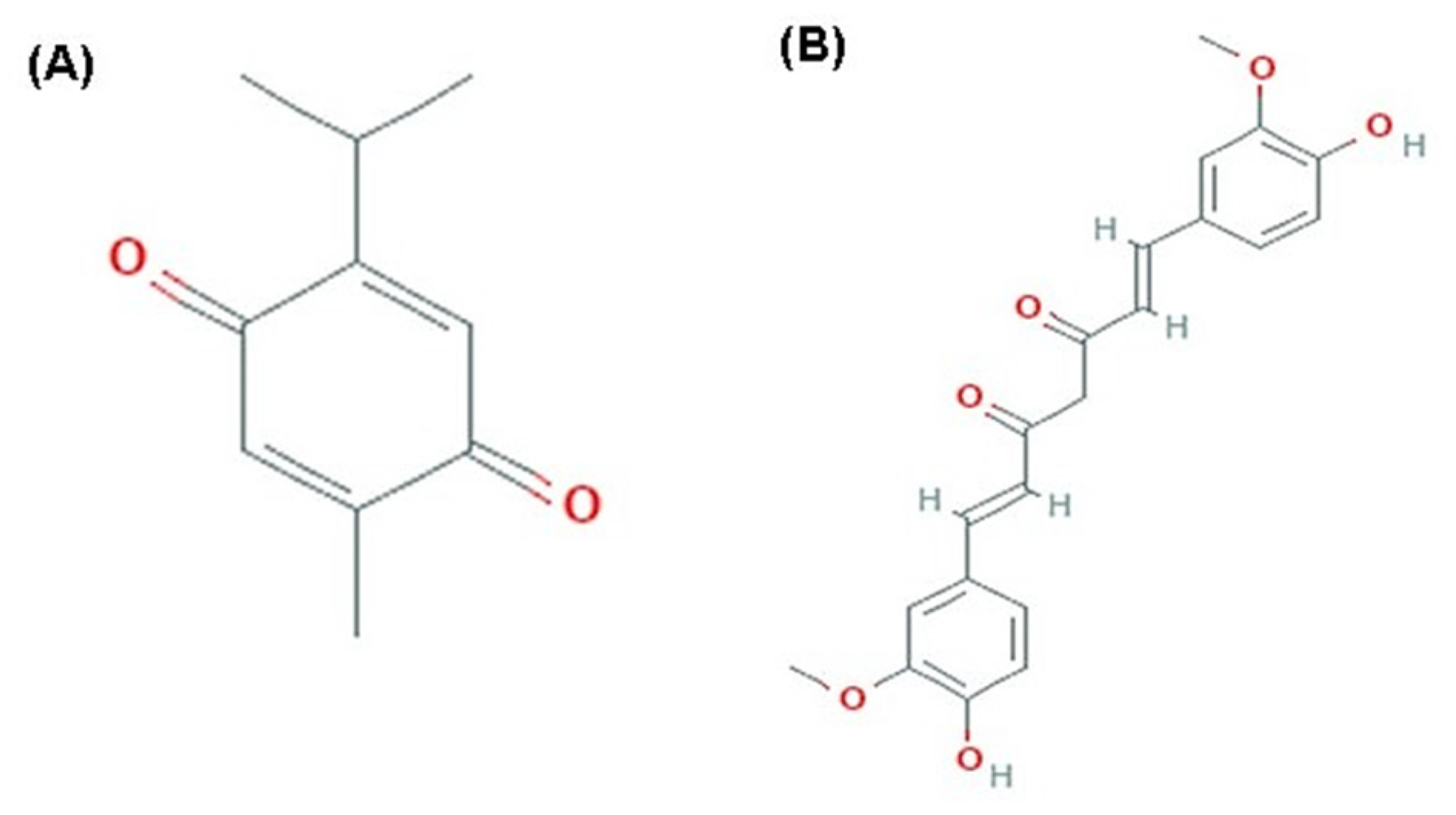
:1. Introduction
2. Results
2.1. Biochemical Parameters
2.2. Histopathological Assessment of the Rat’s Liver
2.3. Histopathological Assessment of the Rat’s Spleen
2.4. Histopathological Assessment of Rat’s Kidney
2.5. Histopathological Assessment of Rat’s Cerebellum
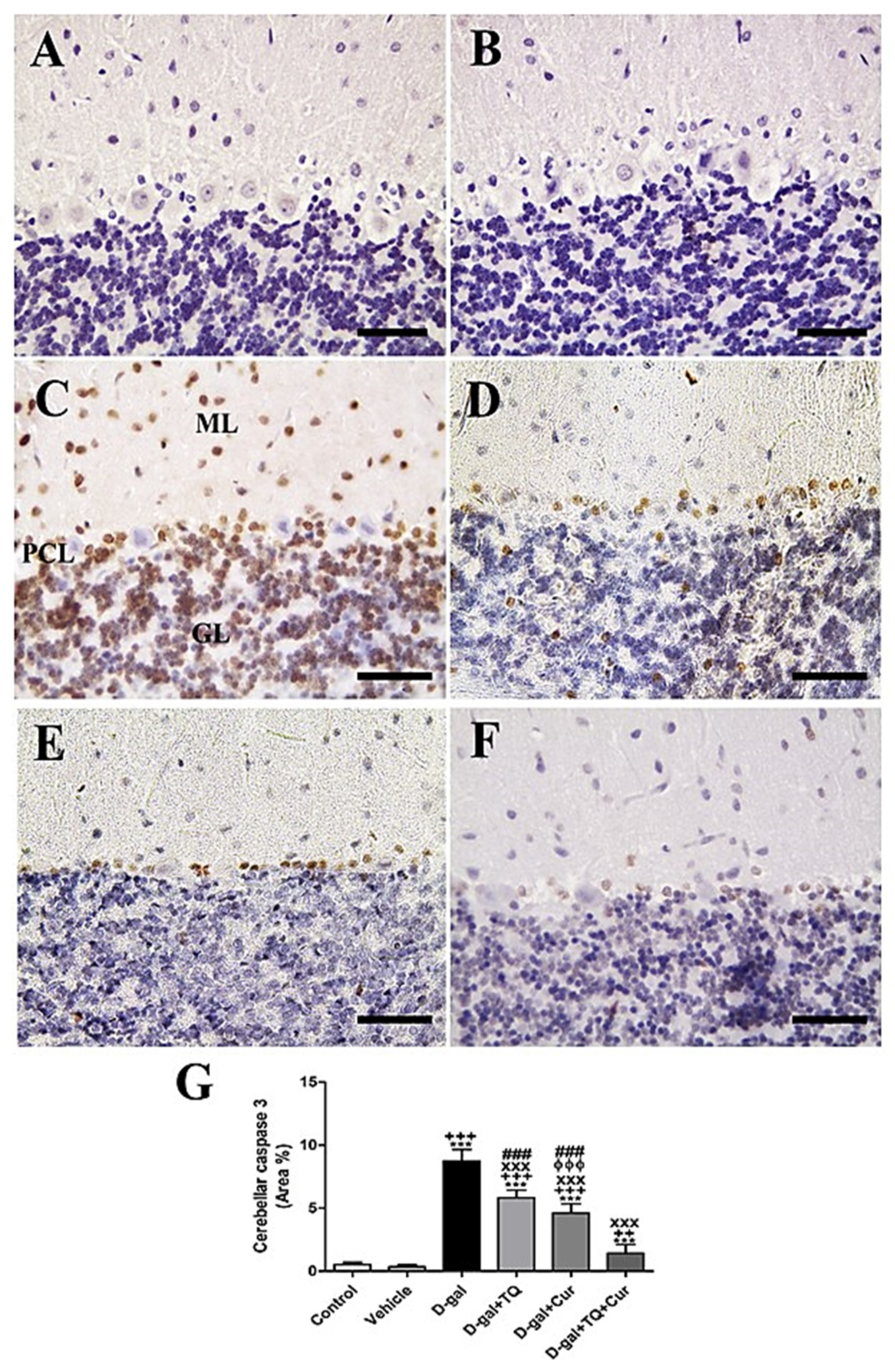
2.6. Immunohistochemistry Assessment of Cerebellum
2.7. Histopathological Assessment of Rat’s Hippocampus
2.8. Immunohistochemistry Assessment of Hippocampus
2.9. Histopathological Assessment of Rat’s Heart
2.10. Immunohistochemistry Assessment of Heart
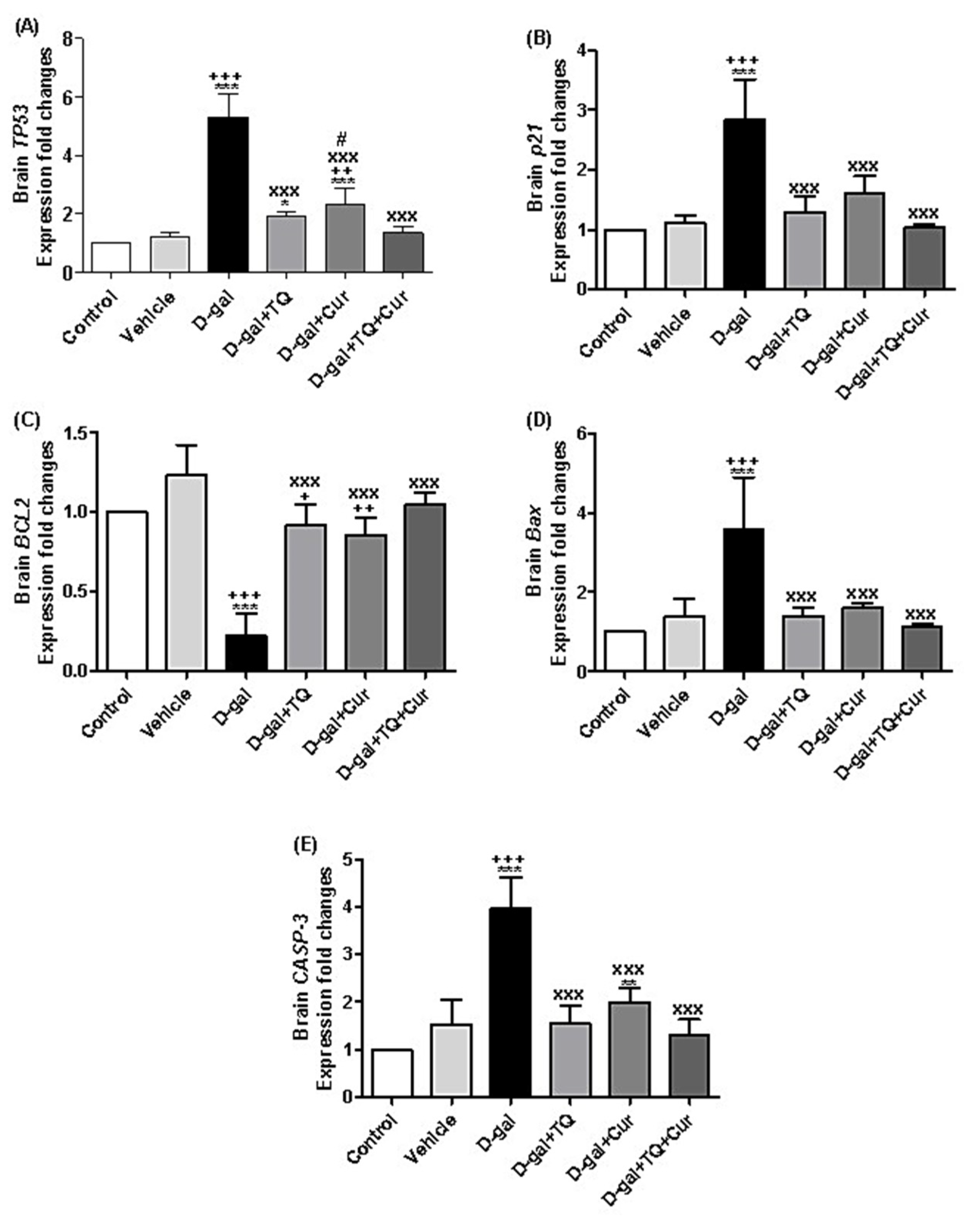
2.11. Effect of Thymoquinone and Curcumin on the Aging-Altered Genes in the Brain
2.12. Effect of Thymoquinone and Curcumin on the Aging-Altered Genes in Heart
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Experimental Design
4.3. Sampling
4.4. Biochemical, Histopathological, Immunohistochemical, and Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Assessments
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, W.K.; Tsai, Y.L.; Shibu, M.A.; Shen, C.Y.; Chang-Lee, S.N.; Chen, R.J.; Yao, C.H.; Ban, B.; Kuo, W.W.; Huang, C.Y. Exercise training augments Sirt1-signaling and attenuates cardiac inflammation in D-galactose induced-aging rats. Aging 2018, 10, 4166–4174. [Google Scholar] [CrossRef]
- Davalli, P.; Mitic, T.; Caporali, A.; Lauriola, A.; D’Arca, D. ROS, Cell Senescence, and Novel Molecular Mechanisms in Aging and Age-Related Diseases. Oxid. Med. Cell. Longev. 2016, 2016, 1–18. [Google Scholar] [CrossRef]
- Zhao, F.; Zhou, Y.; Gao, L.; Qin, X.; Du, G. [Advances in the study of the rat model of aging induced by D-galactose]. Yao Xue Xue Bao 2017, 52, 347–354. [Google Scholar]
- Hegab, Z. Role of advanced glycation end products in cardiovascular disease. World J. Cardiol. 2012, 4, 90. [Google Scholar] [CrossRef]
- Frimat, M.; Daroux, M.; Litke, R.; Nevière, R.; Tessier, F.J.; Boulanger, E. Kidney, heart and brain: Three organs targeted by ageing and glycation. Clin. Sci. 2017, 131, 1069–1092. [Google Scholar] [CrossRef] [PubMed]
- Vivarelli, S.; Falzone, L.; Basile, M.S.; Candido, S.; Libra, M. Nitric Oxide in Hematological Cancers: Partner or Rival? Antioxid. Redox Signal. 2021, 34, 383–401. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef]
- Chen, X.; Guo, C.; Kong, J. Oxidative stress in neurodegenerative diseases. Neural Regen. Res. 2012, 7, 376–385. [Google Scholar] [CrossRef]
- Sahni, S.; Hickok, J.R.; Thomas, D.D. Nitric oxide reduces oxidative stress in cancer cells by forming dinitrosyliron complexes. Nitric Oxide Biol. Chem. 2018, 76, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Ma, B.; Shang, Y.; Yin, Q.; Wang, D.; Xu, S.; Hong, Y.; Hou, X.; Liu, X. Flavonoid-Rich Ethanol Extract from the Leaves of Diospyros kaki Attenuates D-Galactose-Induced Oxidative Stress and Neuroinflammation-Mediated Brain Aging in Mice. Oxid. Med. Cell. Longev. 2018, 2018, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Atta, M.S.; Almadaly, E.A.; El-Far, A.H.; Saleh, R.M.; Assar, D.H.; Al Jaouni, S.K.; Mousa, S.A. Thymoquinone Defeats Diabetes-Induced Testicular Damage in Rats Targeting Antioxidant, Inflammatory and Aromatase Expression. Int. J. Mol. Sci. 2017, 18, 919. [Google Scholar] [CrossRef]
- Atta, M.S.; El-Far, A.H.; Farrag, F.A.; Abdel-Daim, M.M.; Al Jaouni, S.K.; Mousa, S.A. Thymoquinone attenuates cardiomyopathy in streptozotocin-treated diabetic rats. Oxid. Med. Cell. Longev. 2018, 2018, 1–10. [Google Scholar] [CrossRef]
- El-Far, A. Thymoquinone Anticancer Discovery: Possible Mechanisms. Curr. Drug Discov. Technol. 2015, 12, 80–89. [Google Scholar] [CrossRef]
- El-Far, A.H.; Al Jaouni, S.K.; Li, W.; Mousa, S.A. Protective roles of thymoquinone nanoformulations: Potential nanonutraceuticals in human diseases. Nutrients 2018, 10, 1369. [Google Scholar] [CrossRef]
- El-Far, A.H.; Korshom, M.A.; Mandour, A.A.; El-Bessoumy, A.A.; El-Sayed, Y.S. Hepatoprotective efficacy of Nigella sativa seeds dietary supplementation against lead acetate-induced oxidative damage in rabbit—Purification and characterization of glutathione peroxidase. Biomed. Pharmacother. 2017, 89, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Demiroren, K.; Basunlu, M.T.; Erten, R.; Cokluk, E. A comparison of the effects of thymoquinone, silymarin and N-acetylcysteine in an experimental hepatotoxicity. Biomed. Pharmacother. 2018, 106, 1705–1712. [Google Scholar] [CrossRef]
- Üstün, R.; Oğuz, E.K.; Şeker, A.; Korkaya, H. Thymoquinone protects DRG neurons from axotomy-induced cell death. Neurol. Res. 2018, 40, 930–937. [Google Scholar] [CrossRef]
- Menon, V.P.; Sudheer, A.R. Antioxidant and anti-inflammatory properties of curcumin. Adv. Exp. Med. Biol. 2007, 595, 105–125. [Google Scholar]
- Samarghandian, S.; Azimi-Nezhad, M.; Farkhondeh, T.; Samini, F. Anti-oxidative effects of curcumin on immobilization-induced oxidative stress in rat brain, liver and kidney. Biomed. Pharmacother. 2017, 87, 223–229. [Google Scholar] [CrossRef]
- Chakraborty, M.; Bhattacharjee, A.; Kamath, J.V. Cardioprotective effect of curcumin and piperine combination against cyclophosphamide-induced cardiotoxicity. Indian J. Pharmacol. 2017, 49, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Banji, D.; Banji, O.J.F.F.; Dasaroju, S.; Annamalai, A.R.R. Piperine and curcumin exhibit synergism in attenuating d-galactose induced senescence in rats. Eur. J. Pharmacol. 2013, 703, 91–99. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Diagaradjane, P.; Anand, P.; Kuzhuvelil, H.B.; Deorukhkar, A.; Gelovani, J.; Guha, S.; Krishnan, S.; Aggarwal, B.B. Curcumin sensitizes human colorectal cancer to capecitabine by modulation of cyclin D1, COX-2, MMP-9, VEGF and CXCR4 expression in an orthotopic mouse model. Int. J. Cancer 2009, 125, 2187–2197. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Xu, Y.; Meng, L.; Huang, L.; Sun, H. Curcumin inhibits proliferation and promotes apoptosis of breast cancer cells. Exp. Ther. Med. 2018, 16, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; Mobasheri, A.; Lueders, C.; Busch, F.; Shayan, P.; Goel, A. Curcumin enhances the effect of chemotherapy against colorectal cancer cells by inhibition of NF-κB and Src protein kinase signaling pathways. PLoS ONE 2013, 8, e57218. [Google Scholar] [CrossRef]
- Flatt, T. A new definition of aging? Front. Genet. 2012, 3, 148. [Google Scholar] [CrossRef] [PubMed]
- Beckman, K.B.; Ames, B.N. The free radical theory of aging matures. Physiol. Rev. 1998, 78, 547–581. [Google Scholar] [CrossRef] [PubMed]
- Regulski, M.J. Cellular Senescence: What, Why, and How. Wounds 2017, 29, 168–174. [Google Scholar]
- Galanos, P.; Vougas, K.; Walter, D.; Polyzos, A.; Maya-Mendoza, A.; Haagensen, E.J.; Kokkalis, A.; Roumelioti, F.M.; Gagos, S.; Tzetis, M.; et al. Chronic p53-independent p21 expression causes genomic instability by deregulating replication licensing. Nat. Cell Biol. 2016, 18, 777–789. [Google Scholar] [CrossRef]
- Wang, Z.L.; Chen, L.B.; Qiu, Z.; Chen, X.B.; Liu, Y.; Li, J.; Wang, L.; Wang, Y.P. Ginsenoside Rg1 ameliorates testicular senescence changes in D-gal-induced aging mice via anti-inflammatory and antioxidative mechanisms. Mol. Med. Rep. 2018, 17, 6269–6276. [Google Scholar] [CrossRef]
- Sun, J.; Jing, S.; Jiang, R.; Wang, C.; Zhang, C.; Chen, J.; Li, H. Metabolomics study of the therapeutic mechanism of schisandra chinensis lignans on aging rats induced by D-galactose. Clin. Interv. Aging 2018, 13, 829–841. [Google Scholar] [CrossRef]
- Li, Q.; Zeng, J.; Su, M.; He, Y.; Zhu, B. Acetylshikonin from Zicao attenuates cognitive impairment and hippocampus senescence in D-galactose-induced aging mouse model via upregulating the expression of SIRT1. Brain Res. Bull. 2018, 137, 311–318. [Google Scholar] [CrossRef]
- Gali-Muhtasib, H.; Kuester, D.; Mawrin, C.; Bajbouj, K.; Diestel, A.; Ocker, M.; Habold, C.; Foltzer-Jourdainne, C.; Schoenfeld, P.; Peters, B.; et al. Thymoquinone triggers inactivation of the stress response pathway sensor CHEK1 and contributes to apoptosis in colorectal cancer cells. Cancer Res. 2008, 68, 5609–5618. [Google Scholar] [CrossRef] [PubMed]
- Isaev, N.K.; Genrikhs, E.E.; Oborina, M.V.; Stelmashook, E.V. Accelerated aging and aging process in the brain. Rev. Neurosci. 2018, 29, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Shwe, T.; Pratchayasakul, W.; Chattipakorn, N.; Chattipakorn, S.C. Role of D-galactose-induced brain aging and its potential used for therapeutic interventions. Exp. Gerontol. 2018, 101, 13–36. [Google Scholar] [CrossRef]
- Olivetti, G.; Melissari, M.; Capasso, J.M.; Anversa, P. Cardiomyopathy of the aging human heart. Myocyte loss and reactive cellular hypertrophy. Circ. Res. 1991, 68, 1560–1568. [Google Scholar] [CrossRef]
- Kajstura, J.; Cheng, W.; Sarangarajan, R.; Li, P.; Li, B.; Nitahara, J.A.; Chapnick, S.; Reiss, K.; Olivetti, G.; Anversa, P. Necrotic and apoptotic myocyte cell death in the aging heart of Fischer 344 rats. Am. J. Physiol. 1996, 271, H1215–H1228. [Google Scholar] [CrossRef]
- Kung, G.; Konstantinidis, K.; Kitsis, R.N. Programmed necrosis, not apoptosis, in the heart. Circ. Res. 2011, 108, 1017–1036. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, J.Z.; Lin, Z.X.; Yuan, Q.J.; Li, Y.C.; Liang, J.L.; Zhan, J.Y.X.; Xie, Y.L.; Su, Z.R.; Liu, Y.H. Ameliorative effect of supercritical fluid extract of Chrysanthemum indicum Linnén against D-galactose induced brain and liver injury in senescent mice via suppression of oxidative stress, inflammation and apoptosis. J. Ethnopharmacol. 2019, 234, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Ullah, F.; Ali, T.; Ullah, N.; Kim, M.O. Caffeine prevents d-galactose-induced cognitive deficits, oxidative stress, neuroinflammation and neurodegeneration in the adult rat brain. Neurochem. Int. 2015, 90, 114–124. [Google Scholar] [CrossRef]
- Sun, S.L.; Guo, L.; Ren, Y.C.; Wang, B.; Li, R.H.; Qi, Y.S.; Yu, H.; Chang, N.D.; Li, M.H.; Peng, H.S. Anti-apoptosis effect of polysaccharide isolated from the seeds of Cuscuta chinensis Lam on cardiomyocytes in aging rats. Mol. Biol. Rep. 2014, 41, 6117–6124. [Google Scholar] [CrossRef]
- Abulfadl, Y.S.; El-Maraghy, N.N.; Ahmed, A.A.E.; Nofal, S.; Badary, O.A. Protective effects of thymoquinone on D-galactose and aluminum chloride induced neurotoxicity in rats: Biochemical, histological and behavioral changes. Neurol. Res. 2018, 40, 324–333. [Google Scholar] [CrossRef]
- Banji, D.; Banji, O.J.F.F.; Dasaroju, S.; Kumar Ch, K. Curcumin and piperine abrogate lipid and protein oxidation induced by d-galactose in rat brain. Brain Res. 2013, 1515, 1–11. [Google Scholar] [CrossRef]
- Banji, O.J.F.F.; Banji, D.; Ch, K. Curcumin and hesperidin improve cognition by suppressing mitochondrial dysfunction and apoptosis induced by D-galactose in rat brain. Food Chem. Toxicol. 2014, 74, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Fairless, R.; Williams, S.K.; Diem, R. Dysfunction of neuronal calcium signalling in neuroinflammation and neurodegeneration. Cell Tissue Res. 2014, 357, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Fairless, R.; Williams, S.K.; Diem, R. Calcium-Binding Proteins as Determinants of Central Nervous System Neuronal Vulnerability to Disease. Int. J. Mol. Sci. 2019, 20, 2146. [Google Scholar] [CrossRef] [PubMed]
- Ouh, I.-O.; Kim, Y.-M.; Gim, S.-A.; Koh, P.-O. Focal cerebral ischemic injury decreases calbindin expression in brain tissue and HT22 cells. Lab. Anim. Res. 2013, 29, 156. [Google Scholar] [CrossRef]
- Tai, Y.H.; Lin, Y.Y.; Wang, K.C.; Chang, C.L.; Chen, R.Y.; Wu, C.C.; Cheng, I.H. Curcuminoid submicron particle ameliorates cognitive deficits and decreases amyloid pathology in Alzheimer’s disease mouse model. Oncotarget 2018, 9, 10681–10697. [Google Scholar] [CrossRef]
- Sasaki, Y.; Ohsawa, K.; Kanazawa, H.; Kohsaka, S.; Imai, Y. Iba1 is an actin-cross-linking protein in macrophages/microglia. Biochem. Biophys. Res. Commun. 2001, 286, 292–297. [Google Scholar] [CrossRef]
- Ito, D.; Tanaka, K.; Suzuki, S.; Dembo, T.; Fukuuchi, Y. Enhanced Expression of Iba1, Ionized Calcium-Binding Adapter Molecule 1, After Transient Focal Cerebral Ischemia In Rat Brain. Stroke 2001, 32, 1208–1215. [Google Scholar] [CrossRef]
- Ton, B.H.T.; Chen, Q.; Gaina, G.; Tucureanu, C.; Georgescu, A.; Strungaru, C.; Flonta, M.L.; Sah, D.; Ristoiu, V. Activation profile of dorsal root ganglia Iba-1 (+) macrophages varies with the type of lesion in rats. Acta Histochem. 2013, 115, 840–850. [Google Scholar] [CrossRef]
- Romero-Sandoval, A.; Chai, N.; Nutile-McMenemy, N.; DeLeo, J.A. A comparison of spinal Iba1 and GFAP expression in rodent models of acute and chronic pain. Brain Res. 2008, 1219, 116–126. [Google Scholar] [CrossRef]
- Eitzen, U.V.; Egensperger, R.; Kösel, S.; Grasbon-Frodl, E.M.; Imai, Y.; Bise, K.; Kohsaka, S.; Mehraein, P.; Graeber, M.B. Microglia and the Development of Spongiform Change in Creutzfeldt-Jakob Disease. J. Neuropathol. Exp. Neurol. 1998, 57, 246–256. [Google Scholar] [CrossRef]
- Ullah, R.; Jo, M.H.; Riaz, M.; Alam, S.I.; Saeed, K.; Ali, W.; Rehman, I.U.; Ikram, M.; Kim, M.O. Glycine, the smallest amino acid, confers neuroprotection against D-galactose-induced neurodegeneration and memory impairment by regulating c-Jun N-terminal kinase in the mouse brain. J. Neuroinflamm. 2020, 17, 303. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Wang, X.; Chen, G.; Quan, C.; Qu, S.; Tong, J. Inhibiting RIPK1 Limits Neuroinflammation and Alleviates Postoperative Cognitive Impairments in D-Galactose-Induced Aged Mice. Front. Behav. Neurosci. 2018, 12, 138. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Yang, X.; Li, J.; Shu, Z.; Dai, J.; Liu, X.; Li, B.; Jia, S.; Kou, X.; Yang, Y.; et al. Spermidine coupled with exercise rescues skeletal muscle atrophy from D-gal-induced aging rats through enhanced autophagy and reduced apoptosis via AMPK-FOXO3a signal pathway. Oncotarget 2017, 8, 17475–17490. [Google Scholar] [CrossRef] [PubMed]
- Abulfadl, Y.; El-Maraghy, N.; Ahmed, A.E.; Nofal, S.; Abdel-Mottaleb, Y.; Badary, O. Thymoquinone alleviates the experimentally induced Alzheimer’s disease inflammation by modulation of TLRs signaling. Hum. Exp. Toxicol. 2018, 37, 1092–1104. [Google Scholar] [CrossRef]
- Xu, Y.; Lin, D.; Li, S.; Li, G.; Shyamala, S.G.; Barish, P.A.; Vernon, M.M.; Pan, J.; Ogle, W.O. Curcumin reverses impaired cognition and neuronal plasticity induced by chronic stress. Neuropharmacology 2009, 57, 463–471. [Google Scholar] [CrossRef]
- Bancroft, J.; Layton, C. The Hematoxylin and eosin. In Theory Practice of Histological Techniques, 7th ed.; Suvarna, S.K., Layton, C., Bancroft, J.D., Eds.; Churchill Livingstone of El Sevier: Philadelphia, PA, USA, 2013. [Google Scholar]
- Noreldin, A.E.; Elewa, Y.H.A.; Kon, Y.; Warita, K.; Hosaka, Y.Z. Immunohistochemical localization of osteoblast activating peptide in the mouse kidney. Acta Histochem. 2018, 120, 323–328. [Google Scholar] [CrossRef]
- Noreldin, A.E.; Sogabe, M.; Yamano, Y.; Uehara, M.; Mahdy, M.A.A.; Elnasharty, M.A.; Sayed-Ahmed, A.; Warita, K.; Hosaka, Y.Z. Spatial distribution of osteoblast activating peptide in the rat stomach. Acta Histochem. 2016, 118, 109–117. [Google Scholar] [CrossRef]
- El-Far, A.H.; Lebda, M.A.; Noreldin, A.E.; Atta, M.S.; Elewa, Y.H.A.; Elfeky, M.; Mousa, S.A. Quercetin Attenuates Pancreatic and Renal D-Galactose-Induced Aging-Related Oxidative Alterations in Rats. Int. J. Mol. Sci. 2020, 21, 4348. [Google Scholar] [CrossRef]
- Zaragozá, R.; García, C.; Rus, A.D.; Pallardó, F.V.; Barber, T.; Torres, L.; Miralles, V.J.; Viña, J.R. Inhibition of liver trans-sulphuration pathway by propargylglycine mimics gene expression changes found in the mammary gland of weaned lactating rats: Role of glutathione. Biochem. J. 2003, 373, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Dkhil, M.A.; Al-Khalifa, M.S.; Al-Quraishy, S.; Zrieq, R.; Moneim, A.E.A. Indigofera oblongifolia mitigates lead-acetateinduced kidney damage and apoptosis in a rat model. Drug Des. Dev. Ther. 2016, 10, 1847–1856. [Google Scholar] [CrossRef]
- Mohseni, M.; Mihandoost, E.; Shirazi, A.; Sepehrizadeh, Z.; Bazzaz, J.T.; Ghazi-khansari, M. Melatonin may play a role in modulation of bax and bcl-2 expression levels to protect rat peripheral blood lymphocytes from gamma irradiation-induced apoptosis. Mutat. Res. Mol. Mech. Mutagen. 2012, 738–739, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Sarna, L.K.; Siow, Y.L.; O, K. Regulation of hepatic cholesterol biosynthesis by berberine during hyperhomocysteinemia. Am. J. Physiol. Integr. Comp. Physiol. 2011, 300, R635–R643. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | g/kg Diet |
|---|---|
| Corn flour | 529.5 |
| Casein | 200 |
| Sucrose | 100 |
| Soybean oil | 70 |
| Cellulose | 50 |
| Mineral mix | 35 |
| Vitamin mix | 10 |
| L-cystine | 3 |
| Choline | 2.5 |
| Antibody | Source | Dilution | Antigen Retrieval | Heating Condition |
|---|---|---|---|---|
| Rabbit polyclonal anti-caspase 3 | (9662, Cell Signaling Technology, Danvers, MA, USA | 1:300 | 10 mM citrate buffer (pH 6.0) | 105 °C, 20 min |
| Rabbit polyclonal anti-Iba1 | (019-19741, Wako Osaka, Japan) | 1:1200 | 10 mM citrate buffer (pH 6.0) | 105 °C, 20 min |
| Rabbit polyclonal anti-calbindin antibody | (E10340, Spring Bioscience, Pleasanton, CA, USA | 1:500 | 10 mM citrate buffer (pH 6.0) | 105 °C, 20 min |
| Rabbit polyclonal anti-bcl2 | (SC-492, Santa Cruz, CA, USA) | 1:300 | 10 mM citrate buffer (pH 6.0) | 90 °C, 20 min |
| Genes | Genes Forward (5′-3′) | Reverse (5′-3′) | Accession No./References |
|---|---|---|---|
| p21 | GTGAGACACCAGAGTGCAAGA | ACAGCGATATCGAGACACTCA | [62] |
| TP53 | CACAGTCGGATATGAGCATC | GTCGTCCAGATACTCAGCAT | |
| BCL2 | GATTGTGGCCTTCTTTGAGT | ATAGTTCCACAAAGGCATCC | NM_016993/[63] |
| Bax | GGCGAATTGGCGATGAACTG | ATGGTTCTGATCAGCTCGGG | NM_017059/[64] |
| CASP-3 | TTTGCGCCATGCTGAAACT | ACGAGTGAGGATGTGCATGAATT | NM_012922.2 |
| GAPDH (Housekeeping) | TCAAGAAGGTGGTGAAGCAG | AGGTGGAAGAATGGGAGTTG | NM_017008.4/[65] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Far, A.H.; Elewa, Y.H.A.; Abdelfattah, E.-Z.A.; Alsenosy, A.-W.A.; Atta, M.S.; Abou-Zeid, K.M.; Al Jaouni, S.K.; Mousa, S.A.; Noreldin, A.E. RETRACTED: Thymoquinone and Curcumin Defeat Aging-Associated Oxidative Alterations Induced by D-Galactose in Rats’ Brain and Heart. Int. J. Mol. Sci. 2021, 22, 6839. https://doi.org/10.3390/ijms22136839
El-Far AH, Elewa YHA, Abdelfattah E-ZA, Alsenosy A-WA, Atta MS, Abou-Zeid KM, Al Jaouni SK, Mousa SA, Noreldin AE. RETRACTED: Thymoquinone and Curcumin Defeat Aging-Associated Oxidative Alterations Induced by D-Galactose in Rats’ Brain and Heart. International Journal of Molecular Sciences. 2021; 22(13):6839. https://doi.org/10.3390/ijms22136839
Chicago/Turabian StyleEl-Far, Ali H., Yaser H. A. Elewa, Elsayeda-Zeinab A. Abdelfattah, Abdel-Wahab A. Alsenosy, Mustafa S. Atta, Khalid M. Abou-Zeid, Soad K. Al Jaouni, Shaker A. Mousa, and Ahmed E. Noreldin. 2021. "RETRACTED: Thymoquinone and Curcumin Defeat Aging-Associated Oxidative Alterations Induced by D-Galactose in Rats’ Brain and Heart" International Journal of Molecular Sciences 22, no. 13: 6839. https://doi.org/10.3390/ijms22136839
APA StyleEl-Far, A. H., Elewa, Y. H. A., Abdelfattah, E.-Z. A., Alsenosy, A.-W. A., Atta, M. S., Abou-Zeid, K. M., Al Jaouni, S. K., Mousa, S. A., & Noreldin, A. E. (2021). RETRACTED: Thymoquinone and Curcumin Defeat Aging-Associated Oxidative Alterations Induced by D-Galactose in Rats’ Brain and Heart. International Journal of Molecular Sciences, 22(13), 6839. https://doi.org/10.3390/ijms22136839