
Hyaluronan and the Fascial Frontier


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
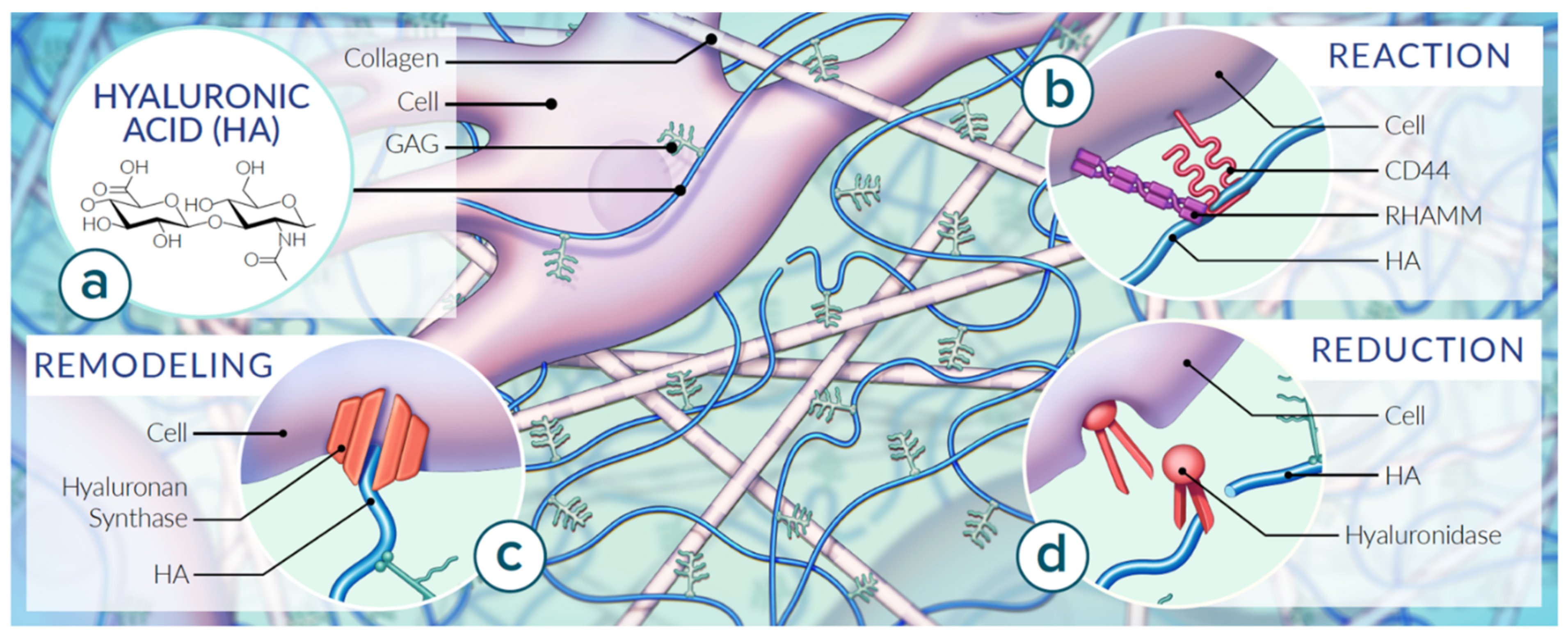
HA Structure and Function
2. Cellular Production of HA
2.1. Fibroblasts
2.2. Smooth Muscle Cells
2.3. Synoviocytes
2.4. Fasciacytes
3. HA in the Extracellular Matrix (ECM)
4. A Home for HA: Fascia
4.1. Definition of Fascia
4.2. Anatomy of Fascia
5. HA and Fascial Gliding
6. HA and Myofascial Pain
6.1. Immobility
6.2. Inflammation
7. Clinical Application of HA
7.1. Manual Therapy
7.2. HA and Treatment
8. Conclusions and Perspective
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HA | hyaluronan |
| CT | connective tissue |
| ECM | extracellular matrix |
| CD44 | cluster determinant 44 |
| RHAMM | receptor for HA-mediated motility |
| MPS | myofascial pain syndrome |
| IVD | intervertebral disc |
| Th1 | T helper 1 cell |
| TS-HA | tyramine-substituted hyaluronan |
| DRC | deep retinaculum cutis |
| SRC | superficial retinaculum cutis |
References
- Meyer, K.; Palmer, J. The Polysaccharide of the Vitreous Humor. J. Biol. Chem. 1934, 107, 629–634. [Google Scholar] [CrossRef]
- Balazs, E.A.; Laurent, T.C.; Jeanloz, R.W. Nomenclature of Hyaluronic Acid. Biochem. J. 1986, 235, 903. [Google Scholar] [CrossRef]
- Fraser, J.R.; Laurent, T.C.; Laurent, U.B. Hyaluronan: Its Nature, Distribution, Functions and Turnover. J. Intern. Med. 1997, 242, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Fakhari, A.; Berkland, C. Applications and Emerging Trends of Hyaluronic Acid in Tissue Engineering, as a Dermal Filler and in Osteoarthritis Treatment. Acta Biomater. 2013, 9, 7081–7092. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Kruglikov, I.L.; Akgul, Y.; Scherer, P.E. Hyaluronan in Adipogenesis, Adipose Tissue Physiology and Systemic Metabolism. Matrix Biol. 2019, 78–79, 284–291. [Google Scholar] [CrossRef]
- Stecco, A.; Gesi, M.; Stecco, C.; Stern, R. Fascial Components of the Myofascial Pain Syndrome. Curr. Pain Headache Rep. 2013, 17, 352–359. [Google Scholar] [CrossRef]
- Jacobetz, M.A.; Chan, D.S.; Neesse, A.; Bapiro, T.E.; Cook, N.; Frese, K.K.; Feig, C.; Nakagawa, T.; Caldwell, M.E.; Zecchini, H.I.; et al. Hyaluronan Impairs Vascular Function and Drug Delivery in a Mouse Model of Pancreatic Cancer. Gut 2013, 62, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Koyama, H.; Hibi, T.; Isogai, Z.; Yoneda, M.; Fujimori, M.; Amano, J.; Kawakubo, M.; Kannagi, R.; Kimata, K.; Taniguchi, S.; et al. Hyperproduction of Hyaluronan in Neu-Induced Mammary Tumor Accelerates Angiogenesis through Stromal Cell Recruitment: Possible Involvement of Versican/PG-M. Am. J. Pathol. 2007, 170, 1086–1099. [Google Scholar] [CrossRef]
- Toole, B.P. Hyaluronan: From Extracellular Glue to Pericellular Cue. Nat. Rev. Cancer 2004, 4, 528–539. [Google Scholar] [CrossRef]
- Avenoso, A.; D’Ascola, A.; Scuruchi, M.; Mandraffino, G.; Calatroni, A.; Saitta, A.; Campo, S.; Campo, G.M. Hyaluronan in Experimental Injured/Inflamed Cartilage: In Vivo Studies. Life Sci. 2018, 193, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Jiang, D.; Noble, P.W. Hyaluronan as a Therapeutic Target in Human Diseases. Adv. Drug Deliv. Rev. 2016, 97, 186–203. [Google Scholar] [CrossRef] [PubMed]
- Hardwick, C.; Hoare, K.; Owens, R.; Hohn, H.P.; Hook, M.; Moore, D.; Cripps, V.; Austen, L.; Nance, D.M.; Turley, E.A. Molecular Cloning of a Novel Hyaluronan Receptor that Mediates Tumor Cell Motility. J. Cell Biol. 1992, 117, 1343–1350. [Google Scholar] [CrossRef] [PubMed]
- Knudson, C.B.; Knudson, W. Hyaluronan and CD44: Modulators of Chondrocyte Metabolism. Clin. Orthop. Relat. Res. 2004, 427, S152–S162. [Google Scholar] [CrossRef]
- Knudson, C.B. Hyaluronan and CD44: Strategic Players for Cell-Matrix Interactions during Chondrogenesis and Matrix Assembly. Birth Defects Res. Part C Embryo Today Rev. 2003, 69, 174–196. [Google Scholar] [CrossRef] [PubMed]
- Nikitovic, D.; Tzardi, M.; Berdiaki, A.; Tsatsakis, A.; Tzanakakis, G.N. Cancer Microenvironment and Inflammation: Role of Hyaluronan. Front. Immunol. 2015, 6, 169. [Google Scholar] [CrossRef]
- Rilla, K.; Siiskonen, H.; Tammi, M.; Tammi, R. Hyaluronan-Coated Extracellular Vesicles—A Novel Link between Hyaluronan and Cancer. Adv. Cancer Res. 2014, 123, 121–148. [Google Scholar] [PubMed]
- Pawlina, W. Histology: A Text and Atlas: With Correlated Cell and Molecular Biology, 8th ed.; LWW: Philadelphia, PA, USA, 2018. [Google Scholar]
- Wolf, K.J.; Kumar, S. Hyaluronic Acid: Incorporating the Bio into the Material. ACS Biomater. Sci. Eng. 2019, 5, 3753–3765. [Google Scholar] [CrossRef]
- Entwistle, J.; Hall, C.L.; Turley, E.A. HA Receptors: Regulators of Signalling to the Cytoskeleton. J. Cell. Biochem. 1996, 61, 569–577. [Google Scholar] [CrossRef]
- Salwowska, N.M.; Bebenek, K.A.; Żądło, D.A.; Wcisło-Dziadecka, D.L. Physiochemical Properties and Application of Hyaluronic Acid: A Systematic Review. J. Cosmet. Dermatol. 2016, 15, 520–526. [Google Scholar] [CrossRef]
- Huang, J.; Qiu, X.; Xie, L.; Jay, G.D.; Schmidt, T.A.; Zeng, H. Probing the Molecular Interactions and Lubrication Mechanisms of Purified Full-Length Recombinant Human Proteoglycan 4 (rhPRG4) and Hyaluronic Acid (HA). Biomacromolecules 2019, 20, 1056–1067. [Google Scholar] [CrossRef]
- Yu, J.; Banquy, X.; Greene, G.W.; Lowrey, D.D.; Israelachvili, J.N. The Boundary Lubrication of Chemically Grafted and Cross-Linked Hyaluronic Acid in Phosphate Buffered Saline and Lipid Solutions Measured by the Surface Forces Apparatus. Langmuir 2012, 28, 2244–2250. [Google Scholar] [CrossRef]
- Cowman, M.K.; Schmidt, T.A.; Raghavan, P.; Stecco, A. Viscoelastic Properties of Hyaluronan in Physiological Conditions. F1000Research 2015, 4, 622. [Google Scholar] [CrossRef]
- Pavan, P.G.; Stecco, A.; Stern, R.; Stecco, C. Painful Connections: Densification Versus Fibrosis of Fascia. Curr. Pain Headache Rep. 2014, 18, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Juel, C. Skeletal Muscle Na+/H+ Exchange in Rats: pH Dependency and the Effect of Training. Acta Physiol. Scand. 1998, 164, 135–140. [Google Scholar] [CrossRef]
- Juel, C.; Klarskov, C.; Nielsen, J.J.; Krustrup, P.; Mohr, M.; Bangsbo, J. Effect of High-Intensity Intermittent Training on Lactate and H+ Release from Human Skeletal Muscle. Am. J. Physiol. Metab. 2004, 286, 245. [Google Scholar] [CrossRef] [PubMed]
- Juel, C.; Bangsbo, J.; Graham, T.; Saltin, B. Lactate and Potassium Fluxes from Human Skeletal Muscle during and After Intense, Dynamic, Knee Extensor Exercise. Acta Physiol. Scand. 1990, 140, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, A.; Rahmanian, M.; Rubin, K.; Heldin, P. Expression of Hyaluronan Synthase 2 Or Hyaluronidase 1 Differentially Affect the Growth Rate of Transplantable Colon Carcinoma Cell Tumors. Int. J. Cancer 2002, 102, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Lokeshwar, V.B.; Cerwinka, W.H.; Lokeshwar, B.L. HYAL1 Hyaluronidase: A Molecular Determinant of Bladder Tumor Growth and Invasion. Cancer Res. 2005, 65, 2243–2250. [Google Scholar] [CrossRef]
- Lokeshwar, V.B.; Cerwinka, W.H.; Isoyama, T.; Lokeshwar, B.L. HYAL1 Hyaluronidase in Prostate Cancer: A Tumor Promoter and Suppressor. Cancer Res. 2005, 65, 7782–7789. [Google Scholar] [CrossRef]
- Voutouri, C.; Stylianopoulos, T. Accumulation of Mechanical Forces in Tumors is Related to Hyaluronan Content and Tissue Stiffness. PLoS ONE 2018, 13, e0193801. [Google Scholar] [CrossRef]
- Fraser, J.R.; Laurent, T.C. Turnover and Metabolism of Hyaluronan. Ciba Found. Symp. 1989, 143, 41–45. [Google Scholar]
- Laurent, T.C.; Fraser, J.R. Hyaluronan. FASEB J. 1992, 6, 2397–2404. [Google Scholar] [CrossRef]
- Fede, C.; Pirri, C.; Fan, C.; Petrelli, L.; Guidolin, D.; De Caro, R.; Stecco, C. A Closer Look at the Cellular and Molecular Components of the Deep/Muscular Fasciae. Int. J. Mol. Sci. 2021, 22, 1411. [Google Scholar] [CrossRef] [PubMed]
- Bordoni, B.; Zanier, E. Understanding Fibroblasts in Order to Comprehend the Osteopathic Treatment of the Fascia. Evid.-Based Complement. Altern. Med. 2015, 2015, 860934. [Google Scholar] [CrossRef] [PubMed]
- Clarris, B.J.; Fraser, J.R. On the Pericellular Zone of some Mammalian Cells in Vitro. Exp. Cell Res. 1968, 49, 181–193. [Google Scholar] [CrossRef]
- Hinata, N.; Murakami, G.; Abe, S.; Shibata, S.; Morizane, S.; Honda, M.; Isoyama, T.; Sejima, T.; Takenaka, A. Coexistence of Elastic Fibers with Hyaluronic Acid in the Human Urethral Sphincter Complex: A Histological Study. J. Urol. 2013, 190, 1313–1319. [Google Scholar] [CrossRef]
- Bartok, B.; Firestein, G.S. Fibroblast-Like Synoviocytes: Key Effector Cells in Rheumatoid Arthritis. Immunol. Rev. 2010, 233, 233–255. [Google Scholar] [CrossRef] [PubMed]
- Laurent, T.C.; Laurent, U.B.; Fraser, J.R. The Structure and Function of Hyaluronan: An Overview. Immunol. Cell Biol. 1996, 74, 1. [Google Scholar] [CrossRef]
- Tamer, T.M. Hyaluronan and Synovial Joint: Function, Distribution and Healing. Interdiscip. Toxicol. 2013, 6, 111–125. [Google Scholar] [CrossRef]
- Stecco, C.; Fede, C.; Macchi, V.; Porzionato, A.; Petrelli, L.; Biz, C.; Stern, R.; De Caro, R. The Fasciacytes: A New Cell Devoted to Fascial Gliding Regulation. Clin. Anat. 2018, 31, 667–676. [Google Scholar] [CrossRef]
- Stecco, C.; Macchi, V.; Porzionato, A.; Duparc, F.; De Caro, R. The Fascia: The Forgotten Structure. Ital. J. Anat. Embryol. 2011, 116, 127–138. [Google Scholar]
- McCombe, D.; Brown, T.; Slavin, J.; Morrison, W.A. The Histochemical Structure of the Deep Fascia and its Structural Response to Surgery. J. Hand Surg. Br. 2001, 26, 89–97. [Google Scholar] [CrossRef]
- Purslow, P.P. The Structure and Role of Intramuscular Connective Tissue in Muscle Function. Front. Physiol. 2020, 11, 495. [Google Scholar] [CrossRef] [PubMed]
- Pratt, R.L. Educational Avenues for Promoting Dialog on Fascia. Clin. Anat. 2019, 32, 871–876. [Google Scholar] [CrossRef]
- Benjamin, M. The Fascia of the Limbs and Back—A Review. J. Anat. 2009, 214, 1–18. [Google Scholar] [CrossRef]
- Chaudhuri, O.; Cooper-White, J.; Janmey, P.A.; Mooney, D.J.; Shenoy, V.B. Effects of Extracellular Matrix Viscoelasticity on Cellular Behaviour. Nature 2020, 584, 535–546. [Google Scholar] [CrossRef]
- Pratt, R.L.; Kinch, M.S. Ligand Binding Up-Regulates EphA2 Messenger RNA through the Mitogen-Activated Protein/Extracellular Signal-Regulated Kinase Pathway. Mol. Cancer Res. 2003, 1, 1070–1076. [Google Scholar] [PubMed]
- Bordoni, B.; Marelli, F.; Morabito, B.; Castagna, R.; Sacconi, B.; Mazzucco, P. New Proposal to Define the Fascial System. Complement. Med. Res. 2018, 25, 257–262. [Google Scholar] [CrossRef]
- Tozzi, P. Selected Fascial Aspects of Osteopathic Practice. J. Bodyw. Mov. Ther. 2012, 16, 503–519. [Google Scholar] [CrossRef]
- Schleip, R.; Jäger, H.; Klingler, W. What is ’Fascia’? A Review of Different Nomenclatures. J. Bodyw. Mov. Ther. 2012, 16, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Stecco, C.; Schleip, R. A Fascia and the Fascial System. J. Bodyw. Mov. Ther. 2016, 20, 139–140. [Google Scholar] [CrossRef]
- Bordoni, B.; Mahabadi, N.; Varacallo, M. Anatomy, Fascia. In StatPearls; StatPearls Publishing LLC: Treasure Island, FL, USA, 2021. [Google Scholar]
- Adstrum, S.; Hedley, G.; Schleip, R.; Stecco, C.; Yucesoy, C.A. Defining the Fascial System. J. Bodyw. Mov. Ther. 2017, 21, 173–177. [Google Scholar] [CrossRef]
- Stecco, C.; Stern, R.; Porzionato, A.; Macchi, V.; Masiero, S.; Stecco, A.; De Caro, R. Hyaluronan within Fascia in the Etiology of Myofascial Pain. Surg. Radiol. Anat. 2011, 33, 891–896. [Google Scholar] [CrossRef]
- Piehl-Aulin, K.; Laurent, C.; Engström-Laurent, A.; Hellström, S.; Henriksson, J. Hyaluronan in Human Skeletal Muscle of Lower Extremity: Concentration, Distribution, and Effect of Exercise. J. Appl. Physiol. 1991, 71, 2493–2498. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Dubey, D.K. Effect of Aggrecan Degradation on the Nanomechanics of Hyaluronan in Extra-Fibrillar Matrix of Annulus Fibrosus: A Molecular Dynamics Investigation. J. Mech. Behav. Biomed. Mater. 2020, 107, 103752. [Google Scholar] [CrossRef]
- Fede, C.; Angelini, A.; Stern, R.; Macchi, V.; Porzionato, A.; Ruggieri, P.; De Caro, R.; Stecco, C. Quantification of Hyaluronan in Human Fasciae: Variations with Function and Anatomical Site. J. Anat. 2018, 233, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Roman, M.; Chaudhry, H.; Bukiet, B.; Stecco, A.; Findley, T.W. Mathematical Analysis of the Flow of Hyaluronic Acid Around Fascia during Manual Therapy Motions. J. Am. Osteopath. Assoc. 2013, 113, 600–610. [Google Scholar] [CrossRef]
- Matteini, P.; Dei, L.; Carretti, E.; Volpi, N.; Goti, A.; Pini, R. Structural Behavior of Highly Concentrated Hyaluronan. Biomacromolecules 2009, 10, 1516–1522. [Google Scholar] [CrossRef] [PubMed]
- Dintenfass, L. Lubrication in Synovial Joints. Nature 1963, 197, 496–497. [Google Scholar] [CrossRef] [PubMed]
- Tadmor, R.; Chen, N.; Israelachvili, J.N. Thin Film Rheology and Lubricity of Hyaluronic Acid Solutions at a Normal Physiological Concentration. J. Biomed. Mater. Res. 2002, 61, 514–523. [Google Scholar] [CrossRef]
- Petrey, A.C.; de la Motte, C.A. Hyaluronan, a Crucial Regulator of Inflammation. Front. Immunol. 2014, 5, 101. [Google Scholar] [CrossRef]
- Litwiniuk, M.; Krejner, A.; Speyrer, M.S.; Gauto, A.R.; Grzela, T. Hyaluronic Acid in Inflammation and Tissue Regeneration. Wounds Compend. Clin. Res. Pract. 2016, 28, 78–88. [Google Scholar]
- Litwiniuk, M.; Cioffi, A. Hyaluronic Acid as Regulator of Inflammatory and Immune Responses. Ital. J. Allergy Clin. Immunol. 2011, 21, 83–88. [Google Scholar]
- Cylwik, B.; Gruszewska, E.; Gindzienska-Sieskiewicz, E.; Kowal-Bielecka, O.; Chrostek, L. Serum Profile of Transferrin Isoforms in Rheumatoid Arthritis Treated with Biological Drugs. Clin. Biochem. 2019, 74, 31–35. [Google Scholar] [CrossRef]
- Hughes, E.J.; McDermott, K.; Funk, M.F. Evaluation of Hyaluronan Content in Areas of Densification Compared to Adjacent Areas of Fascia. J. Bodyw. Mov. Ther. 2019, 23, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Langevin, H.M.; Huijing, P.A. Communicating about Fascia: History, Pitfalls, and Recommendations. Int. J. Ther. Massage Bodyw. 2009, 2, 3–8. [Google Scholar]
- Klingler, W.; Velders, M.; Hoppe, K.; Pedro, M.; Schleip, R. Clinical Relevance of Fascial Tissue and Dysfunctions. Curr. Pain Headache Rep. 2014, 18, 439. [Google Scholar] [CrossRef]
- Ingber, D.E. Tensegrity and Mechanotransduction. J. Bodyw. Mov. Ther. 2008, 12, 198–200. [Google Scholar] [CrossRef] [PubMed]
- Al-Khateeb, R.; Prpic, J. Hyaluronic Acid: The Reason for its Variety of Physiological and Biochemical Functional Properties. Appl. Clin. Res. Clin. Trials Regul. Aff. 2019, 6, 112–159. [Google Scholar] [CrossRef]
- Kumai, T.; Samoto, N.; Hasegawa, A.; Noguchi, H.; Shiranita, A.; Shiraishi, M.; Ikeda, S.; Sugimoto, K.; Tanaka, Y.; Takakura, Y. Short-Term Efficacy and Safety of Hyaluronic Acid Injection for Plantar Fasciopathy. Knee Surg. Sports Traumatol. Arthrosc. 2018, 26, 903–911. [Google Scholar] [CrossRef]
- Chin, L.; Calabro, A.; Rodriguez, E.R.; Tan, C.D.; Walker, E.; Derwin, K.A. Characterization of and Host Response to Tyramine Substituted-Hyaluronan Enriched Fascia Extracellular Matrix. J. Mater. Sci. Mater. Med. 2011, 22, 1465–1477. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chin, L.; Calabro, A.; Walker, E.; Derwin, K.A. Mechanical Properties of Tyramine Substituted-Hyaluronan Enriched Fascia Extracellular Matrix. J. Biomed. Mater. Res. Part A 2012, 100, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Chhetri, D.K.; Mendelsohn, A.H. Hyaluronic Acid for the Treatment of Vocal Fold Scars. Curr. Opin. Otolaryngol. Head Neck Surg. 2010, 18, 498–502. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pratt, R.L. Hyaluronan and the Fascial Frontier. Int. J. Mol. Sci. 2021, 22, 6845. https://doi.org/10.3390/ijms22136845
Pratt RL. Hyaluronan and the Fascial Frontier. International Journal of Molecular Sciences. 2021; 22(13):6845. https://doi.org/10.3390/ijms22136845
Chicago/Turabian StylePratt, Rebecca L. 2021. "Hyaluronan and the Fascial Frontier" International Journal of Molecular Sciences 22, no. 13: 6845. https://doi.org/10.3390/ijms22136845
APA StylePratt, R. L. (2021). Hyaluronan and the Fascial Frontier. International Journal of Molecular Sciences, 22(13), 6845. https://doi.org/10.3390/ijms22136845