
Optogenetic Manipulation of Olfactory Responses in Transgenic Zebrafish: A Neurobiological and Behavioral Study

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
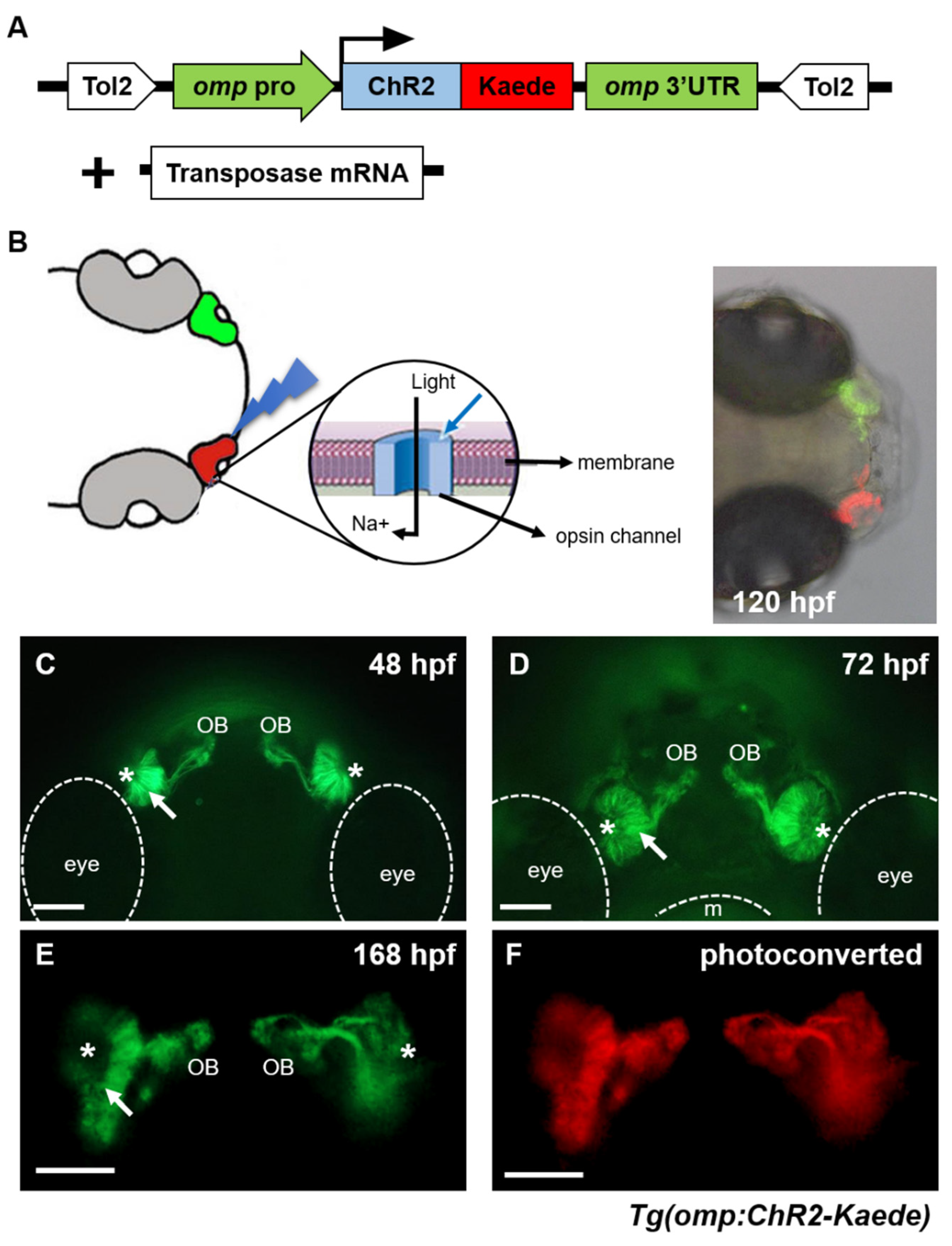
2.1. Establishment and Characterization of Transgenic Zebrafish Expressing Channelrhodopsin-2 in Olfactory Sensory Neurons
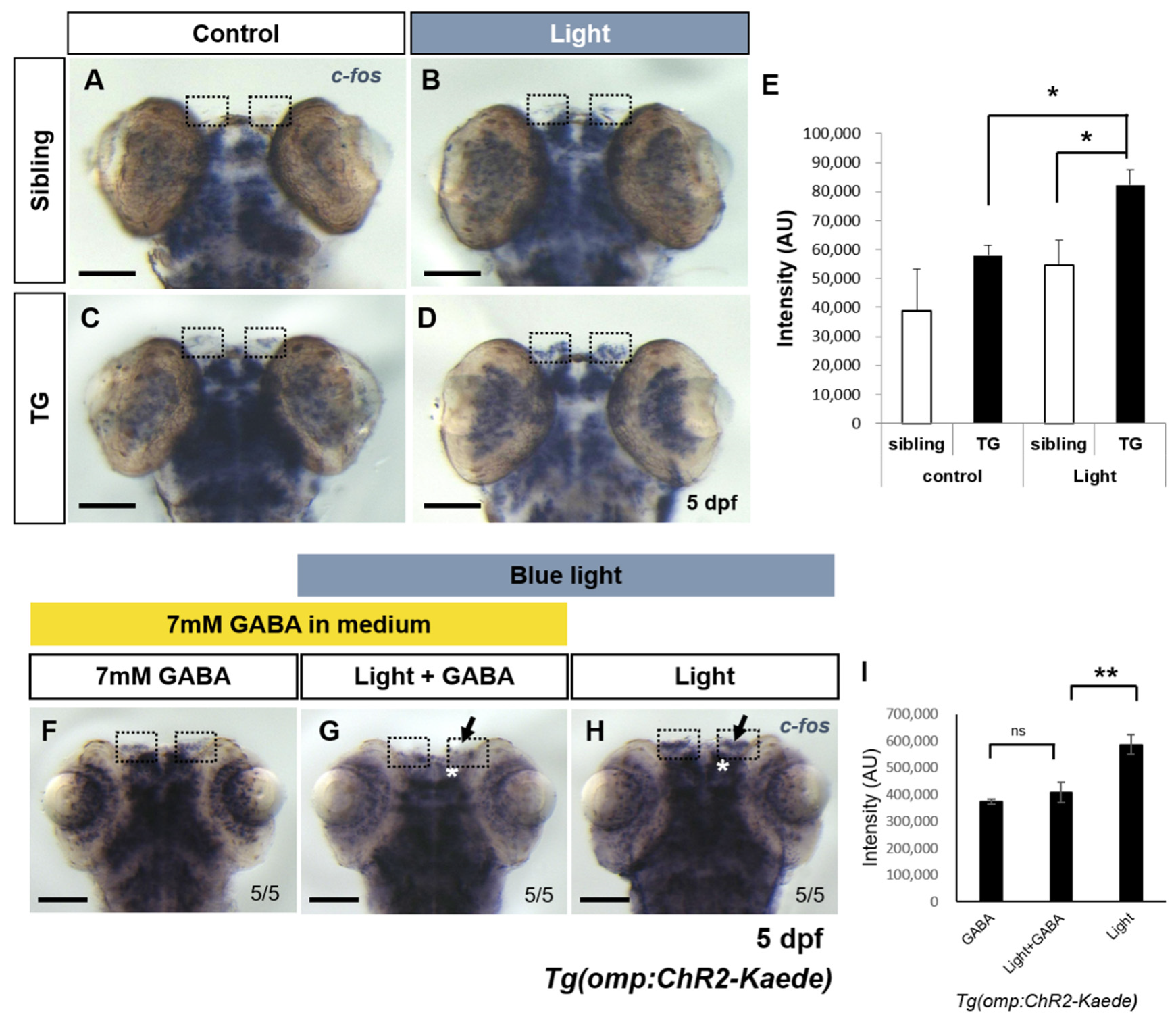
2.2. Evaluation of Light Induced Neuronal Activity Using Immediate Early Gene in the Transgenic Fish
2.3. Inhibition of Light-Evoked Activity by GABA Agonist in Transgenic Fish
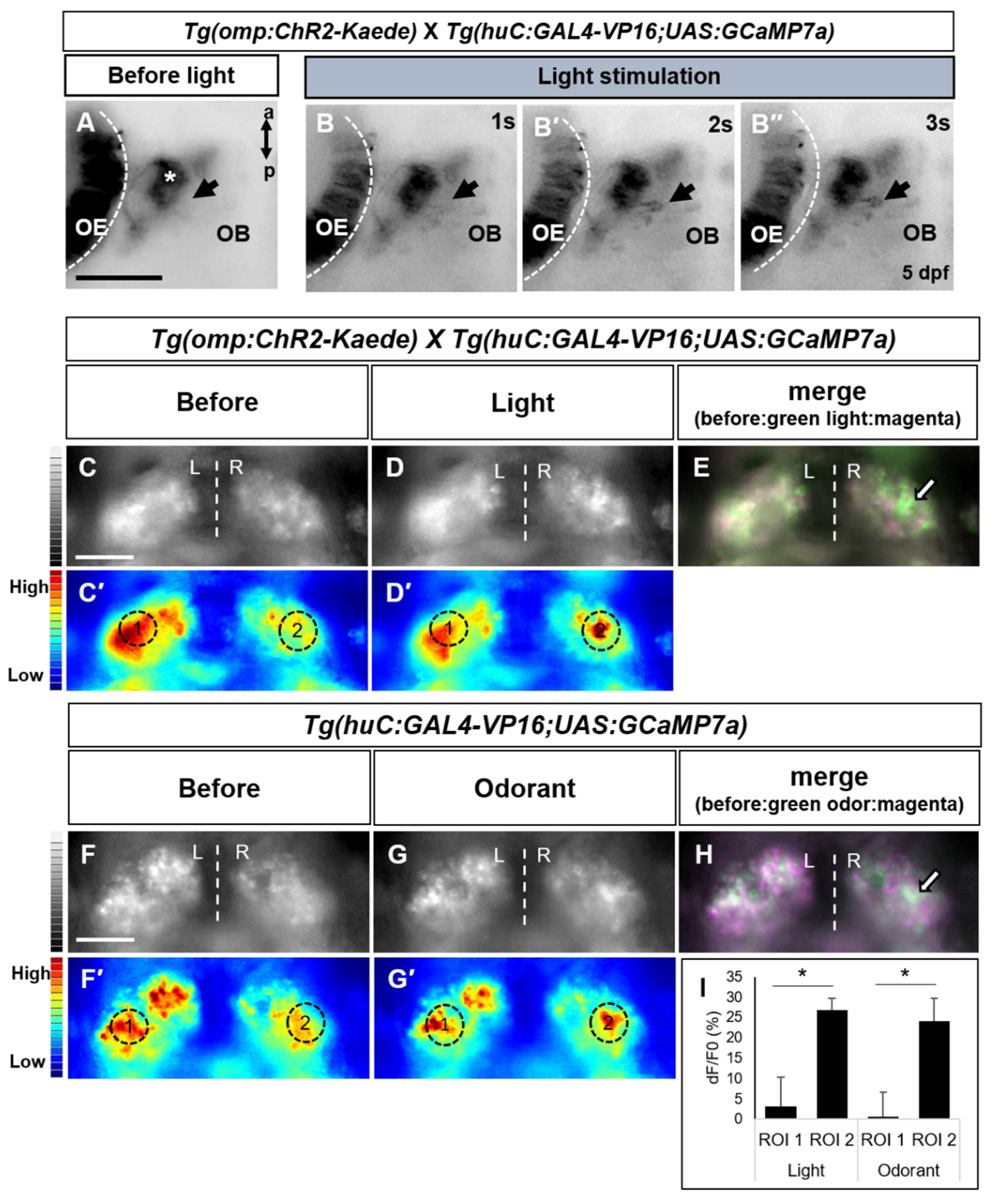
2.4. Light-Driven Neuronal Activity Analysis in the Olfactory Brain Area Using a Genetically Encoded Calcium Indicator
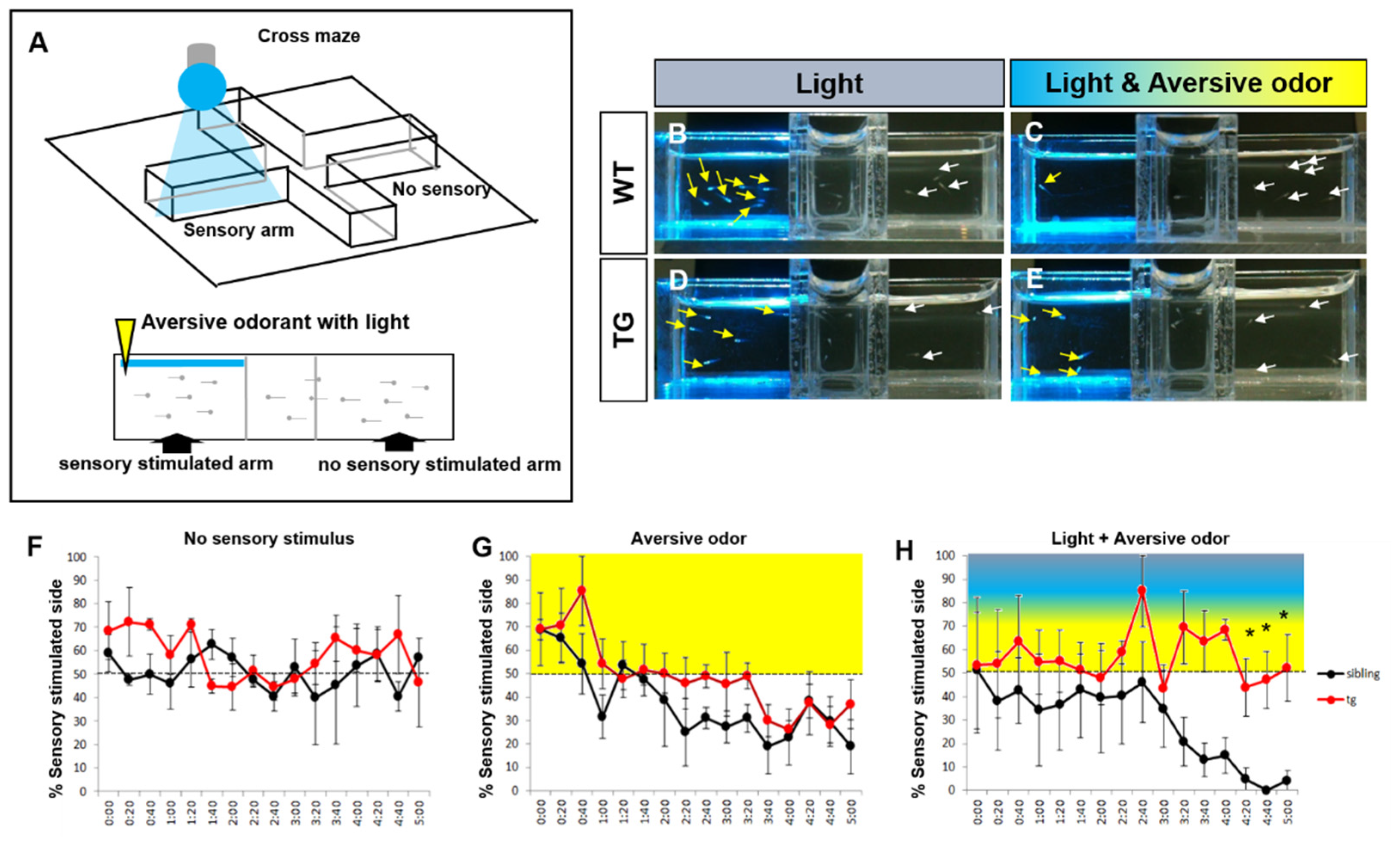
2.5. Behavioral Adaptation to Light Evoked Odor Response in the Transgenic Zebrafish
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Establishment Transgenic Zebrafish for the Omp-Driven ChR2-Kaede
4.2.1. Construction of Expression Vector
4.2.2. Microinjection and Generation of Transgenic Zebrafish Lines
4.3. Whole-Mount In Situ Hybridization
4.4. Light Stimulation and Calcium Imaging
4.5. Behavioral Assay Odorant Treatment
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ChR2 | Channelrhodpsin |
| omp | olfactory marker protein |
| GABA | gamma-Aminobutyric acid |
| GECI | genetically encoded calcium indicator |
| OSNs | olfactory sensory neurons |
References
- Song, Y.H.; Kim, J.H.; Jeong, H.W.; Choi, I.; Jeong, D.; Kim, K.; Lee, S.H. A Neural Circuit for Auditory Dominance over Visual Perception. Neuron 2017, 93, 1236–1237. [Google Scholar] [CrossRef] [PubMed]
- Horberg, T.; Larsson, M.; Ekstrom, I.; Sandoy, C.; Lunden, P.; Olofsson, J.K. Olfactory Influences on Visual Categorization: Behavioral and ERP Evidence. Cereb. Cortex 2020, 30, 4220–4237. [Google Scholar] [CrossRef]
- Yoshihara, Y. Molecular genetic dissection of the zebrafish olfactory system. Results Probl. Cell Differ. 2009, 47, 97–120. [Google Scholar]
- Whitlock, K.E. The sense of scents: Olfactory behaviors in the zebrafish. Zebrafish 2006, 3, 203–213. [Google Scholar] [CrossRef]
- Kermen, F.; Darnet, L.; Wiest, C.; Palumbo, F.; Bechert, J.; Uslu, O.; Yaksi, E. Stimulus-specific behavioral responses of zebrafish to a large range of odors exhibit individual variability. BMC Biol. 2020, 18, 66. [Google Scholar] [CrossRef]
- Burgess, H.A.; Schoch, H.; Granato, M. Distinct retinal pathways drive spatial orientation behaviors in zebrafish navigation. Curr. Biol. 2010, 20, 381–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, K.P.; Neuhauss, S.C.F. Behavioral neurobiology: How larval fish orient towards the light. Curr. Biol. 2010, 20, R159–R161. [Google Scholar] [CrossRef] [Green Version]
- Lau, B.Y.; Mathur, P.; Gould, G.G.; Guo, S. Identification of a brain center whose activity discriminates a choice behavior in zebrafish. Proc. Natl. Acad. Sci. USA 2011, 108, 2581–2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, A.M.; Fero, K.; Arrenberg, A.B.; Bergeron, S.A.; Driever, W.; Burgess, H.A. Deep brain photoreceptors control light-seeking behavior in zebrafish larvae. Curr. Biol. 2012, 22, 2042–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Chen, S.; Liu, S.; Zhang, C.; Peng, G. Effects of lorazepam and WAY-200070 in larval zebrafish light/dark choice test. Neuropharmacology 2015, 95, 226–233. [Google Scholar] [CrossRef]
- Facciol, A.; Iqbal, M.; Eada, A.; Tran, S.; Gerlai, R. The light-dark task in zebrafish confuses two distinct factors: Interaction between background shade and illumination level preference. Pharmacol. Biochem. Behav. 2019, 179, 9–21. [Google Scholar] [CrossRef]
- Facciol, A.; Tran, S.; Gerlai, R. Re-examining the factors affecting choice in the light-dark preference test in zebrafish. Behav. Brain Res. 2017, 327, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Bogli, S.Y.; Huang, M.Y. Spontaneous alternation behavior in larval zebrafish. J. Exp. Biol. 2017, 220, 171–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, A.C.; Bill, B.R.; Glanzman, D.L. Learning and memory in zebrafish larvae. Front. Neural Circuits 2013, 7, 126. [Google Scholar] [CrossRef] [Green Version]
- Deisseroth, K. Optogenetics. Nat. Methods 2011, 8, 26–29. [Google Scholar] [CrossRef]
- Portugues, R.; Severi, K.E.; Wyart, C.; Ahrens, M.B. Optogenetics in a transparent animal: Circuit function in the larval zebrafish. Curr. Opin. Neurobiol. 2013, 23, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Vierock, J.; Yizhar, O.; Fenno, L.E.; Tsunoda, S.; Kianianmomeni, A.; Prigge, M.; Berndt, A.; Cushman, J.; Polle, J.; et al. The microbial opsin family of optogenetic tools. Cell 2011, 147, 1446–1457. [Google Scholar] [CrossRef] [Green Version]
- Nagel, G.; Brauner, M.; Liewald, J.F.; Adeishvili, N.; Bamberg, E.; Gottschalk, A. Light activation of channelrhodopsin-2 in excitable cells of Caenorhabditis elegans triggers rapid behavioral responses. Curr. Biol. 2005, 15, 2279–2284. [Google Scholar] [CrossRef] [Green Version]
- Douglass, A.D.; Kraves, S.; Deisseroth, K.; Schier, A.F.; Engert, F. Escape behavior elicited by single, channelrhodopsin-2-evoked spikes in zebrafish somatosensory neurons. Curr. Biol. 2008, 18, 1133–1137. [Google Scholar] [CrossRef] [Green Version]
- Kravitz, A.V.; Freeze, B.S.; Parker, P.R.; Kay, K.; Thwin, M.T.; Deisseroth, K.; Kreitzer, A.C. Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry. Nature 2010, 466, 622–626. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Miyasaka, N.; Yoshihara, Y. Mutually exclusive glomerular innervation by two distinct types of olfactory sensory neurons revealed in transgenic zebrafish. J. Neurosci. 2005, 25, 4889–4897. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Ito, A.; Matsuda, N.; Mishina, M. Regulation by protein kinase A switching of axonal pathfinding of zebrafish olfactory sensory neurons through the olfactory placode–olfactory bulb boundary. J. Neurosci. 2002, 22, 4964–4972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ando, R.; Hama, H.; Yamamoto-Hino, M.; Mizuno, H.; Miyawaki, A. An optical marker based on the UV-induced green-to-red photoconversion of a fluorescent protein. Proc. Natl. Acad. Sci. USA 2002, 99, 12651–12656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, H.Y.; Kim, O.H.; Kim, H.T.; Choi, J.H.; Yeo, S.Y.; Kim, N.S.; Park, D.S.; Oh, H.W.; You, K.H.; De Zoysa, M.; et al. Establishment of a transgenic zebrafish EF1alpha:Kaede for monitoring cell proliferation during regeneration. Fish. Shellfish Immunol. 2013, 34, 1390–1394. [Google Scholar] [CrossRef]
- Arrenberg, A.B.; Del Bene, F.; Baier, H. Optical control of zebrafish behavior with halorhodopsin. Proc. Natl. Acad. Sci. USA 2009, 106, 17968–17973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curran, T.; Morgan, J.I. Fos: An immediate-early transcription factor in neurons. J. Neurobiol. 1995, 26, 403–412. [Google Scholar] [CrossRef]
- Kovacs, K.J. Measurement of immediate-early gene activation- c-fos and beyond. J. Neuroendocrinol. 2008, 20, 665–672. [Google Scholar] [CrossRef]
- Okuno, H. Regulation and function of immediate-early genes in the brain: Beyond neuronal activity markers. Neurosci. Res. 2011, 69, 175–186. [Google Scholar] [CrossRef]
- Bowen, M.T.; Kevin, R.C.; May, M.; Staples, L.G.; Hunt, G.E.; McGregor, I.S. Defensive aggregation (huddling) in Rattus norvegicus toward predator odor: Individual differences, social buffering effects and neural correlates. PLoS ONE 2013, 8, e68483. [Google Scholar] [CrossRef]
- DeCarvalho, T.N.; Akitake, C.M.; Thisse, C.; Thisse, B.; Halpern, M.E. Aversive cues fail to activate fos expression in the asymmetric olfactory-habenula pathway of zebrafish. Front. Neural Circuits 2013, 7, 98. [Google Scholar] [CrossRef] [Green Version]
- Mack-Bucher, J.A.; Li, J.; Friedrich, R.W. Early functional development of interneurons in the zebrafish olfactory bulb. Eur. J. Neurosci. 2007, 25, 460–470. [Google Scholar] [CrossRef]
- Tabor, R.; Yaksi, E.; Friedrich, R.W. Multiple functions of GABAA and GABAB receptors during pattern processing in the zebrafish olfactory bulb. Eur. J. Neurosci. 2008, 28, 117–127. [Google Scholar] [CrossRef]
- Kimura, Y.; Satou, C.; Higashijima, S. V2a and V2b neurons are generated by the final divisions of pair-producing progenitors in the zebrafish spinal cord. Development 2008, 135, 3001–3005. [Google Scholar] [CrossRef] [Green Version]
- Muto, A.; Ohkura, M.; Abe, G.; Nakai, J.; Kawakami, K. Real-time visualization of neuronal activity during perception. Curr. Biol. 2013, 23, 307–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyasaka, N.; Morimoto, K.; Tsubokawa, T.; Higashijima, S.; Okamoto, H.; Yoshihara, Y. From the olfactory bulb to higher brain centers: Genetic visualization of secondary olfactory pathways in zebrafish. J. Neurosci. 2009, 29, 4756–4767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreosti, E.; Llopis, N.V.; Carl, M.; Yaksi, E.; Wilson, S.W. Left-right asymmetry is required for the habenulae to respond to both visual and olfactory stimuli. Curr. Biol. 2014, 24, 440–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amo, R.; Fredes, F.; Kinoshita, M.; Aoki, R.; Aizawa, H.; Agetsuma, M.; Aoki, T.; Shiraki, T.; Kakinuma, H.; Matsuda, M.; et al. The habenulo-raphe serotonergic circuit encodes an aversive expectation value essential for adaptive active avoidance of danger. Neuron 2014, 84, 1034–1048. [Google Scholar] [CrossRef] [Green Version]
- Viswanath, H.; Carter, A.Q.; Baldwin, P.R.; Molfese, D.L.; Salas, R. The medial habenula: Still neglected. Front. Hum. Neurosci. 2013, 7, 931. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Mathuru, A.S.; Kibat, C.; Rahman, M.; Lupton, C.E.; Stewart, J.; Claridge-Chang, A.; Yen, S.C.; Jesuthasan, S. The right dorsal habenula limits attraction to an odor in zebrafish. Curr. Biol. 2014, 24, 1167–1175. [Google Scholar] [CrossRef] [Green Version]
- Jetti, S.K.; Vendrell-Llopis, N.; Yaksi, E. Spontaneous activity governs olfactory representations in spatially organized habenular microcircuits. Curr. Biol. 2014, 24, 434–439. [Google Scholar] [CrossRef] [Green Version]
- Esposti, F.; Johnston, J.; Rosa, J.M.; Leung, K.M.; Lagnado, L. Olfactory stimulation selectively modulates the OFF pathway in the retina of zebrafish. Neuron 2013, 79, 97–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zufall, F.; Leinders-Zufall, T. The cellular and molecular basis of odor adaptation. Chem. Senses 2000, 25, 473–481. [Google Scholar] [CrossRef]
- Stortkuhl, K.F.; Fiala, A. The Smell of Blue Light: A New Approach toward Understanding an Olfactory Neuronal Network. Front. Neurosci. 2011, 5, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishimoto, N.; Asakawa, K.; Madelaine, R.; Blader, P.; Kawakami, K.; Sawamoto, K. Interhemispheric asymmetry of olfactory input-dependent neuronal specification in the adult brain. Nat. Neurosci. 2013, 16, 884–888. [Google Scholar] [CrossRef]
- Milbrandt, J. Nerve growth factor rapidly induces c-fos mRNA in PC12 rat pheochromocytoma cells. Proc. Natl. Acad. Sci. USA 1986, 83, 4789–4793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecoq, J.; Tiret, P.; Charpak, S. Peripheral adaptation codes for high odor concentration in glomeruli. J. Neurosci. 2009, 29, 3067–3072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Maaswinkel, H.; Li, L. Olfactoretinal centrifugal input modulates zebrafish retinal ganglion cell activity: A possible role for dopamine-mediated Ca2+ signalling pathways. J. Physiol. 2005, 569, 939–948. [Google Scholar] [CrossRef] [PubMed]
- DeMaria, S.; Ngai, J. The cell biology of smell. J. Cell Biol. 2010, 191, 443–452. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Lee, H.; Choi, M.-K.; Lee, D.; Kim, H.-S.; Hwang, H.; Kim, H.; Park, S.; Paik, Y.-K.; Lee, J. Nictation, a dispersal behavior of the nematode Caenorhabditis elegans, is regulated by IL2 neurons. Nat. Neurosci. 2012, 15, 107. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.T.; Kim, E.H.; Yoo, K.W.; Lee, M.S.; Choi, J.H.; Park, H.C.; Yeo, S.Y.; Lee, D.S.; Kim, C.H. Isolation and expression analysis of Alzheimer’s disease-related gene xb51 in zebrafish. Dev. Dyn. 2008, 237, 3921–3926. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, Y.-M.; Choi, T.-I.; Hwang, K.-S.; Lee, J.-S.; Gerlai, R.; Kim, C.-H. Optogenetic Manipulation of Olfactory Responses in Transgenic Zebrafish: A Neurobiological and Behavioral Study. Int. J. Mol. Sci. 2021, 22, 7191. https://doi.org/10.3390/ijms22137191
Jeong Y-M, Choi T-I, Hwang K-S, Lee J-S, Gerlai R, Kim C-H. Optogenetic Manipulation of Olfactory Responses in Transgenic Zebrafish: A Neurobiological and Behavioral Study. International Journal of Molecular Sciences. 2021; 22(13):7191. https://doi.org/10.3390/ijms22137191
Chicago/Turabian StyleJeong, Yun-Mi, Tae-Ik Choi, Kyu-Seok Hwang, Jeong-Soo Lee, Robert Gerlai, and Cheol-Hee Kim. 2021. "Optogenetic Manipulation of Olfactory Responses in Transgenic Zebrafish: A Neurobiological and Behavioral Study" International Journal of Molecular Sciences 22, no. 13: 7191. https://doi.org/10.3390/ijms22137191