
Rhodamine 6G-Ligand Influencing G-Quadruplex Stability and Topology




Abstract
1. Introduction
2. Results and Discussion
2.1. The Spectral Properties of DNA-Ligand Complexes
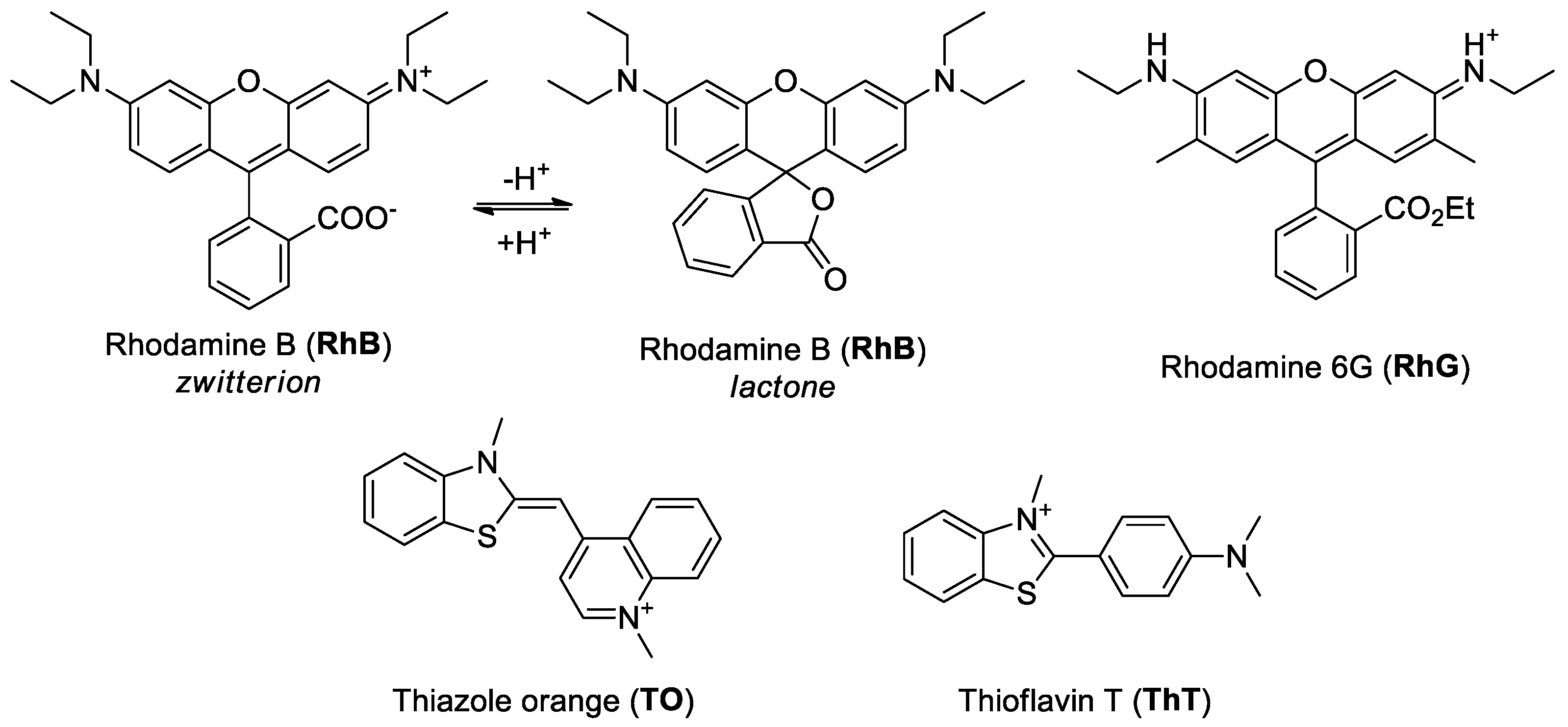
RhG versus RhB
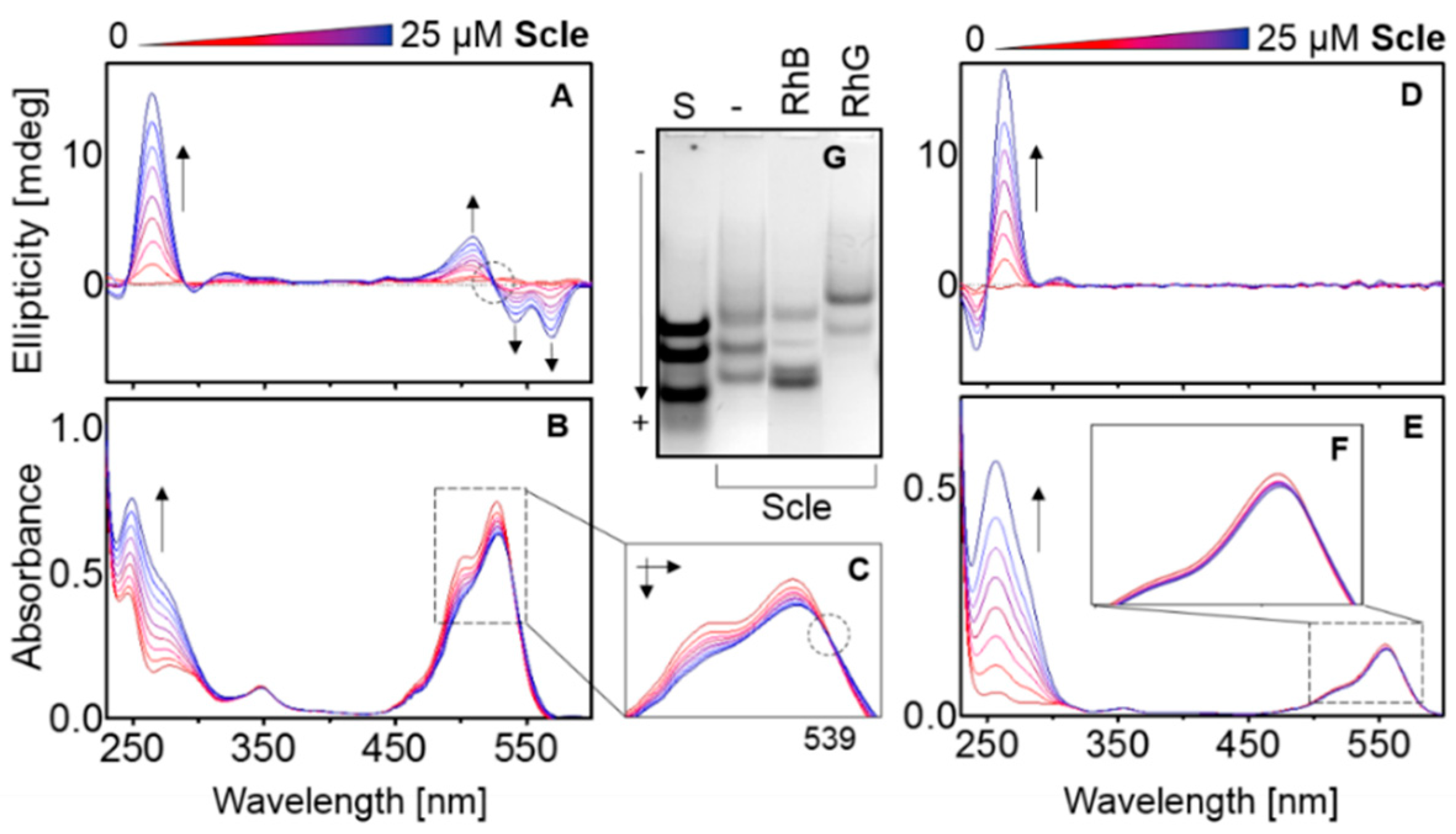
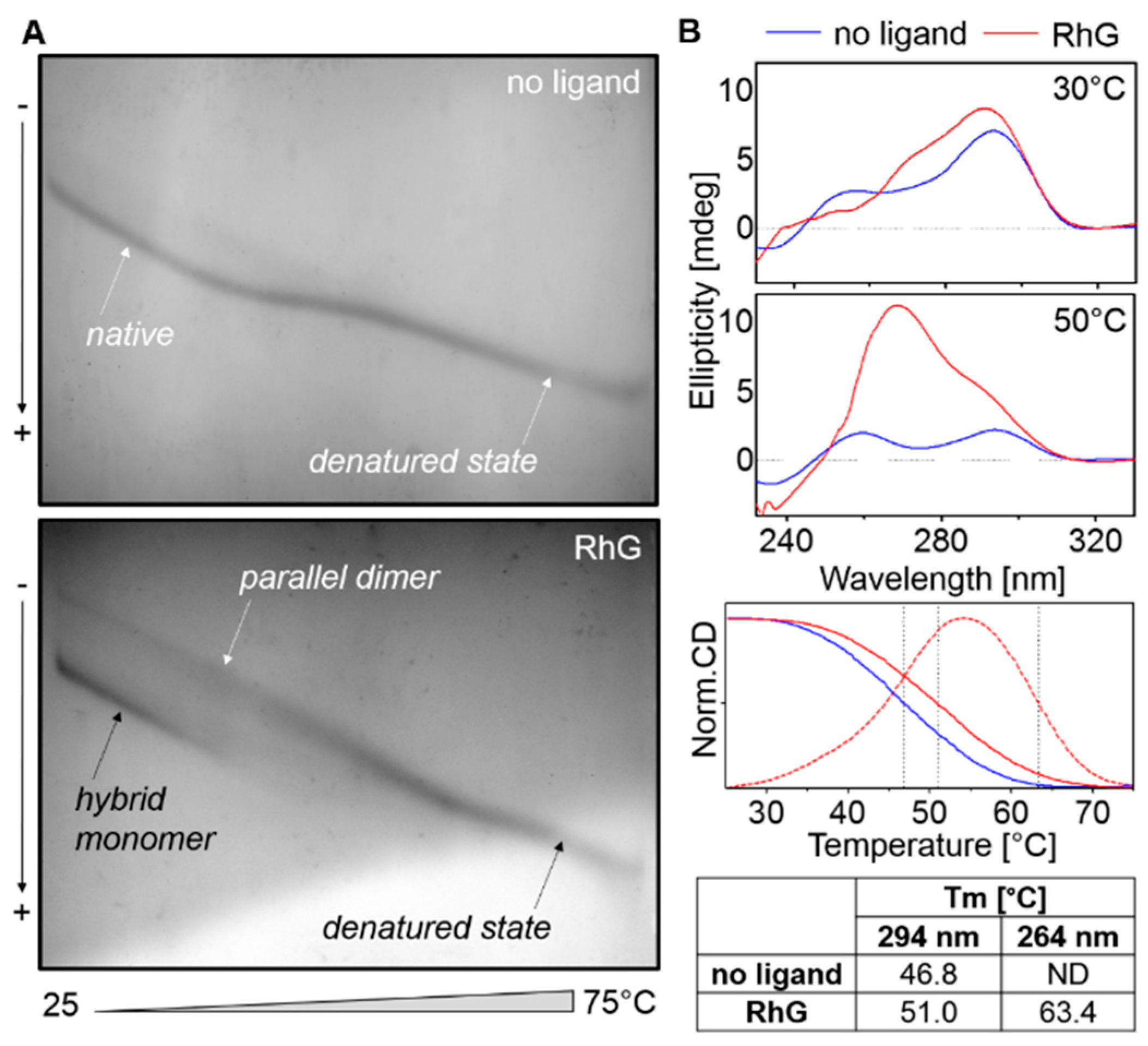
2.2. RhG: Influence on Polymorphism and Stability
2.3. Temperature and Concentration Measurements
2.4. Fluorescence Spectroscopic Properties of RhB and RhG
2.5. Molecular Modeling of Ligand-G4 Interactions
3. Materials and Methods
3.1. Circular Dichroism Spectroscopy
3.2. CD Melting Curves
3.3. Electrophoresis
3.4. Fluorescence Spectroscopy
3.5. Docking Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Bohálová, N.; Cantara, A.; Bartas, M.; Kaura, P.; Šťastný, J.; Pečinka, P.; Fojta, M.; Mergny, J.L.; Brázda, V. Analyses of viral genomes for G-quadruplex forming sequences reveal their correlation with the type of infection. Biochimie 2021, 186, 13–27. [Google Scholar] [CrossRef]
- Ruggiero, E.; Richter, S.N. G-quadruplexes and G-quadruplex ligands: Targets and tools in antiviral therapy. Nucleic Acids Res. 2018, 46, 3270–3283. [Google Scholar] [CrossRef] [PubMed]
- Viglasky, V.; Hianik, T. Potential uses of G-quadruplex-forming aptamers. Gen. Physiol. Biophys. 2013, 32, 149–172. [Google Scholar] [CrossRef]
- Roxo, C.; Kotkowiak, W.; Pasternak, A. G-Quadruplex-Forming Aptamers-Characteristics, Applications, and Perspectives. Molecules 2019, 24, 3781. [Google Scholar] [CrossRef] [PubMed]
- Banco, M.T.; Ferré-D’Amaré, A.R. The emerging structural complexity of G-quadruplex RNAs. RNA 2021, 27, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Mitra, J.; Ha, T. Nanomechanics and co-transcriptional folding of Spinach and Mango. Nat. Commun. 2019, 10, 4318. [Google Scholar] [CrossRef] [PubMed]
- Eddy, J.; Maizels, N. Gene function correlates with potential for G4 DNA formation in the human genome. Nucleic Acids Res. 2006, 34, 3887–3896. [Google Scholar] [CrossRef]
- Bedrat, A.; Lacroix, L.; Mergny, J.L. Re-evaluation of G-quadruplex propensity with G4Hunter. Nucleic Acids Res. 2016, 44, 1746–1759. [Google Scholar] [CrossRef] [PubMed]
- Huppert, J.L.; Balasubramanian, S. Prevalence of quadruplexes in the human genome. Nucleic Acids Res. 2005, 33, 2908–2916. [Google Scholar] [CrossRef]
- Huppert, J.L. Four-stranded nucleic acids: Structure, function and targeting of G-quadruplexes. Chem. Soc. Rev. 2008, 37, 1375–1384. [Google Scholar] [CrossRef]
- Bidzinska, J.; Cimino-Reale, G.; Zaffaroni, N.; Folini, M. G-quadruplex structures in the human genome as novel therapeutic targets. Molecules 2013, 18, 12368–12395. [Google Scholar] [CrossRef] [PubMed]
- Neidle, S. Quadruplex nucleic acids as targets for anticancer therapeutics. Nat. Rev. Chem. 2017, 1, 95. [Google Scholar] [CrossRef]
- Ou, T.M.; Lu, Y.J.; Tan, J.H.; Huang, Z.S.; Wong, K.Y.; Gu, L.Q. G-quadruplexes: Targets in anticancer drug design. ChemMedChem 2008, 3, 690–713. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Zheng, Y.; Zhai, Q.; Wei, D. Recent advances in the development of small molecules targeting RNA G-quadruplexes for drug discovery. Bioorg. Chem. 2021, 110, 804. [Google Scholar] [CrossRef]
- Berrones Reyes, J.; Kuimova, M.K.; Vilar, R. Metal complexes as optical probes for DNA sensing and imaging. Curr. Opin. Chem. Biol. 2021, 61, 179–190. [Google Scholar] [CrossRef]
- Guan, L.; Zhao, J.; Sun, W.; Deng, W.; Wang, L. Meso-Substituted Thiazole Orange for Selective Fluorescence Detection to G-Quadruplex DNA and Molecular Docking Simulation. ACS Omega 2020, 5, 26056–26062. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, L.; Jiang, W. Label free DNA detection based on gold nanoparticles quenching fluorescence of Rhodamine B. Talanta 2011, 85, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, Y.; Wang, L.; Huang, Y.; Liu, D.; Sun, R.; Luo, J.; Sun, C. A facile aptamer-based sensing strategy for dopamine through the fluorescence resonance energy transfer between rhodamine B and gold nanoparticles. Dyes Pigments 2015, 123, 55–63. [Google Scholar] [CrossRef]
- Kennedy, J.; Larrañeta, E.; McCrudden, M.T.C.; McCrudden, C.M.; Brady, A.J.; Fallows, S.J.; McCarthy, H.O.; Kissenpfennig, A.; Donnelly, R.F. In vivo studies investigating biodistribution of nanoparticle-encapsulated rhodamine B delivered via dissolving microneedles. J. Control. Release 2017, 265, 57–65. [Google Scholar] [CrossRef]
- Jbeily, N.; Claus, R.A.; Dahlke, K.; Neugebauer, U.; Bauer, M.; Gonnert, F.A. Comparative suitability of CFDA-SE and rhodamine 6G for in vivo assessment of leukocyte-endothelium interactions. J. Biophotonics 2014, 7, 369–375. [Google Scholar] [CrossRef]
- Thaler, S.; Haritoglou, C.; Choragiewicz, T.J.; Messias, A.; Baryluk, A.; May, C.A.; Rejdak, R.; Fiedorowicz, M.; Zrenner, E.; Schuettauf, F. In vivo toxicity study of rhodamine 6G in the rat retina. Investig. Ophthalmol. Vis. Sci. 2008, 49, 2120–2126. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Li, Q.; Xiang, J.; Yang, Q.; Sun, H.; Guan, A.; Wang, L.; Liu, Y.; Yu, L.; Shi, Y.; et al. Thioflavin T as an efficient fluorescence sensor for selective recognition of RNA G-quadruplexes. Sci. Rep. 2016, 6, 24793. [Google Scholar] [CrossRef]
- Largy, E.; Granzhan, A.; Hamon, F.; Verga, D.; Teulade-Fichou, M.P. Visualizing the quadruplex: From fluorescent ligands to light-upprobes. Top. Curr. Chem. 2013, 330, 111–177. [Google Scholar]
- Vorlíčková, M.; Kejnovská, I.; Bednářová, K.; Renčiuk, D.; Kypr, J. Circular dichroism spectroscopy of DNA: From duplexes to quadruplexes. Chirality 2012, 24, 691–698. [Google Scholar] [CrossRef]
- Hud, N.V.; Smith, F.W.; Anet, F.A.; Feigon, J. The selectivity for K+ versus Na+ in DNA quadruplexes is dominated by relative free energies of hydration: A thermodynamic analysis by 1H NMR. Biochemistry 1996, 35, 15383–15390. [Google Scholar] [CrossRef]
- Luu, K.N.; Phan, A.T.; Kuryavyi, V.; Lacroix, L.; Patel, D.J. Structure of the human telomere in K+ solution: An intramolecular (3 + 1) G-quadruplex scaffold. J. Am. Chem. Soc. 2006, 128, 9963–9970. [Google Scholar] [CrossRef]
- Tóthová, P.; Krafčíková, P.; Víglaský, V. Formation of highly ordered multimers in G-quadruplexes. Biochemistry 2014, 53, 7013–7027. [Google Scholar] [CrossRef]
- Tlučková, K.; Marusič, M.; Tóthová, P.; Bauer, L.; Sket, P.; Plavec, J.; Víglasky, V. Human papillomavirus G-quadruplexes. Biochemistry 2013, 52, 7207–7216. [Google Scholar] [CrossRef] [PubMed]
- Kocman, V.; Plavec, J. A tetrahelical DNA fold adopted by tandem repeats of alternating GGG and GCG tracts. J. Nat. Commun. 2014, 5, 5831. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Iaccarino, N.; Di Porzio, A.; Amato, J.; Pagano, B.; Brancaccio, D.; Novellino, E.; Leardi, R.; Randazzo, A. Assessing the influence of pH and cationic strength on i-motif DNA structure. Anal. Bioanal. Chem. 2019, 411, 7473–7479. [Google Scholar] [CrossRef] [PubMed]
- Demkovičová, E.; Bauer, Ľ.; Krafčíková, P.; Tlučková, K.; Tóthova, P.; Halaganová, A.; Valušová, E.; Víglaský, V. Telomeric G-Quadruplexes: From Human to Tetrahymena Repeats. J. Nucleic Acids 2017, 2017, 9170371. [Google Scholar] [CrossRef] [PubMed]
- Krafčíková, P.; Demkovičová, E.; Víglaský, V. Ebola virus derived G-quadruplexes: Thiazole orange interaction. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 1321–1328. [Google Scholar] [CrossRef] [PubMed]
- Helttunen, K.; Prus, P.; Luostarinen, M.; Nissinen, M. Interaction of aminomethylated resorcinarenes with rhodamine B. New J. Chem. 2009, 33, 1148–1154. [Google Scholar] [CrossRef]
- Shum, K.T.; Chan, C.; Leung, C.M.; Tanner, J.A. Identification of a DNA aptamer that inhibits sclerostin’s antagonistic effect on Wnt signalling. Biochem. J. 2011, 434, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Víglaský, V.; Antalík, M.; Bagel’ová, J.; Tomori, Z.; Podhradský, D. Heat-induced conformational transition of cytochrome c observed by temperature gradient gel electrophoresis at acidic pH. Electrophoresis 2000, 21, 850–858. [Google Scholar] [CrossRef]
- Víglaský, V.; Bauer, L.; Tlucková, K. Structural features of intra- and intermolecular G-quadruplexes derived from telomeric repeats. Biochemistry 2010, 49, 2110–2120. [Google Scholar] [CrossRef]
- Krafčíková, P.; Demkovičová, E.; Halaganová, A.; Víglaský, V. Putative HIV and SIV G-Quadruplex Sequences in Coding and Noncoding Regions Can Form G-Quadruplexes. J. Nucleic Acids 2017, 2017, 6513720. [Google Scholar] [CrossRef]
- Gabelica, V.; Maeda, R.; Fujimoto, T.; Yaku, H.; Murashima, T.; Sugimoto, N.; Miyoshi, D. Multiple and cooperative binding of fluorescence light-up probe thioflavin T with human telomere DNA G-quadruplex. Biochemistry 2013, 52, 5620–5628. [Google Scholar] [CrossRef]
- Lubitz, I.; Zikich, D.; Kotlyar, A. Specific high-affinity binding of thiazole orange to triplex and G-quadruplex DNA. Biochemistry 2010, 49, 3567–3574. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, J.; Barooah, N.; Dhamodharan, V.; Harikrishna, S.; Pradeepkumar, P.I.; Bhasikuttan, A.C. Thioflavin T as an efficient inducer and selective fluorescent sensor for the human telomeric G-quadruplex DNA. J. Am. Chem. Soc. 2013, 135, 367–376. [Google Scholar] [CrossRef]
- Sjekloca, L.; Ferre-D’Amare, A.R. Binding between G Quadruplexes at the Homodimer Interface of the Corn RNA Aptamer Strongly Activates Thioflavin T Fluorescence. Cell Chem. Biol. 2019, 26, 1159. [Google Scholar] [CrossRef]
- Campbell, N.H.; Parkinson, G.N.; Reszka, A.P.; Neidle, S. Structural basis of DNA quadruplex recognition by an acridine drug. J. Am. Chem. Soc. 2008, 130, 6722–6724. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Bauer, L.; Tlučková, K.; Tóhová, P.; Viglaský, V. G-quadruplex motifs arranged in tandem occurring in telomeric repeats and the insulin-linked polymorphic region. Biochemistry 2011, 50, 7484–7492. [Google Scholar] [CrossRef] [PubMed]
- Poniková, S.; Tlučková, K.; Antalík, M.; Víglaský, V.; Hianik, T. The circular dichroism and differential scanning calorimetry study of the properties of DNA aptamer dimers. Biophys. Chem. 2011, 155, 29–35. [Google Scholar] [CrossRef]
- Stewart, J.J.P. Optimization of parameters for semiempirical methods I. Method. J. Comput. Chem. 1989, 10, 209–220. [Google Scholar] [CrossRef]
- Dai, J.; Carver, M.; Punchihewa, C.; Jones, R.A.; Yang, D. Structure of the hybrid-2 type intramolecular human telomeric G-quadruplex in K+ solution: Insights into structure polymorphism of the human telomeric sequence. Nucleic Acids Res. 2007, 35, 4927–4940. [Google Scholar] [CrossRef]
- Do, N.Q.; Lim, K.W.; Teo, M.H.; Heddi, B.; Phan, A.T. Stacking of G-quadruplexes: NMR structure of a G-rich oligonucleotide with potential anti-HIV and anticancer activity. Nucleic Acids Res. 2011, 39, 9448–9457. [Google Scholar] [CrossRef]
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graphics Mod. 1999, 17, 57–61. [Google Scholar]
- Morris, G.M.; Ruth, H.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. Software news and updates AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Melting Temperature [°C] | |||||
|---|---|---|---|---|---|
| Oligo | Wavelength [nm] | 50 mM NaCl | 50 mM KCl | ||
| No Ligand | RhG | No Ligand | RhG | ||
| HTR | 294 | 52.0 | 52.0 | 63.5 | 64.0 |
| Scle | 264 | 62.0 | 77.0 | 81.5 | >100 |
| TBA | 294 | 20.0 | 26.8 | 46.5 | 48.5 |
| Hema | 264 | ND | 54.3 | 72.0 | 86.0 |
| STAT | 264 | 54.5 | 76.5 | 92.8 | >100 |
| HCV | 264 | 44.5 | 60.0 | 72.6 | 86.0 |
| ionK | 294 | 46.2 | 48.5 | 59.1 | 58.0 |
| VEGF | 264 | 47.5 | 82.0 | 85.6 | 87.9 |
| no ligand a | RhG a | ||||
| C3A2T b | 286 | 28.3 | 24.3 | ||
| TFO1 | 282 | 20.4 | 17.7 | ||
| No. | Name | Sequence in 5′→3′ Direction | Category and Preferred Motif | |
|---|---|---|---|---|
| 1 | G3A | GGGAGGGAGGGAGGGA | G3Nn [31] | G-quadruplex |
| 2 | G3C | GGGCGGGCGGGCGGGC | ||
| 3 | G3T | GGGTGGGTGGGTGGGT | ||
| 4 | G3T2 | GGGTTGGGTTGGGTTGGG | ||
| 5 | G3T3 | GGGTTTGGGTTTGGGTTTGGG | ||
| 6 | G3T4 | GGGTTTTGGGTTTTGGGTTTTGGG | ||
| 7 | HTR | GGGTTAGGGTTAGGGTTAGGG | ||
| 8 | HTR2 | AGGGTTAGGGTTAGGGTTAGGGT | ||
| 9 | HTR-T | GGGTTAGGGTTAGGGTTAGGGT | ||
| 10 | G3T2C | GGGTTCGGGTTCGGGTTCGGG | ||
| 11 | 8G3 | GGGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGG | ||
| 12 | 8G3T2 | GGGTTGGGTTGGGTTGGGTTGGGTTGGGTTGGGTTGGG | ||
| 13 | 8G3T3 | GGGTTTGGGTTTGGGTTTGGGTTTGGGTTTGGGTTTGGGTTTGGG | ||
| 14 | G3-3-A20 | GGGTTAGGGTTAGGGTTAGGGAAAAAAAAAAAAAAAAAAAA | ||
| 15 | G3-5-T20 | TTTTTTTTTTTTTTTTTTTTGGGTTAGGGTTAGGGTTAGGG | ||
| 16 | G4T | GGGGTGGGGTGGGGTGGGG | G4Nn [31] | |
| 17 | G4T2 | GGGGTTGGGGTTGGGGTTGGGG | ||
| 18 | G4T3 | GGGGTTTGGGGTTTGGGGTTTGGGG | ||
| 19 | G4T4 | GGGGTTTTGGGGTTTTGGGGTTTTGGGG | ||
| 20 | G4T2A | GGGGTTAGGGGTTAGGGGTTAGGGG | ||
| 21 | HCV | GGGCGTGGTGGGTGGGGT | Aptamers [45] | |
| 22 | Hema | GGGGTCGGGCGGGCCGGGTG | ||
| 23 | HIV | GGGGTGGGAGGAGGGT | ||
| 24 | Insu | GGTGGTGGGGGGGGTTGGTAGGGT | ||
| 25 | ionK | GGGTTAGGGTTAGGGTAGGG | ||
| 26 | OCH-A | CGGGTGTGGGTGGCGTAAAGGGA | ||
| 27 | Scle | TGGGGGGGTGGGTGGGT | ||
| 28 | STAT | GGGCGGGCGGGCGGG | ||
| 29 | TBA | GGTTGGTGTGGTTGG | ||
| 30 | TBA-5T | GGTTGGTGTGGTTGGTTTTTGGTTGGTGTGGTTGG | ||
| 31 | VEGF | GGGGCGGGCCGGGGGCGGG | ||
| 32 | HIV1-K02 | GTGGCCTGGGCGGGACTGGGGA | HIV [37] | |
| 33 | HIV1-K03 | CGGGGTTGGGAGGTGGGT | ||
| 34 | HIV1-L20 | TGGGAGGGATAAGGGGCGGTTCGGGGA | ||
| 35 | HIV1-M27 | GTGGCCTGGGCGGGACTTGGGA | ||
| 36 | E-Cote2 | TGGGGAGGGTGGGGAGGGTGGGGAAGG | Ebola virus [32] | |
| 37 | E-Cote4 | TGGGATGGGTGGGGTGCTTGTCTGGGGC | ||
| 38 | MarRavn | GTGGTCGGCGTGGGGGGGAGGGT | ||
| 39 | c-myc | TGGGGAGGGTGGGGAGGGTGGGGAAGG | Others | |
| 40 | N-myc | TAGGGCGGGAGGGAGGGAA | ||
| 41 | pUC-G1 | GGGGTGTTGGCGGGTGTCGGGGC | ||
| 42 | RAN | TGGGGGTGGGGTTGGGTGGTGT | ||
| 43 | RAN-del | TGGGGGTGGGGTTGGGTGGT | ||
| 44 | Z-G4 | TGGTGGTGGTGTGGTGGTGGTGGTGTT | ||
| 45 | i-HTR | CCCAATCCCAATCCCAATCCC | i-motif | |
| 46 | i-HTR2 | TCCCAATCCCAATCCCAATCCCA | ||
| 47 | C3-Msl1 | CCCTAACCCTAAACCCTAACCC | ||
| 48 | AC9 | ACACACACA | ssDNA | |
| 49 | AC12 | ACACACACACAC | ||
| 50 | AC18 | ACACACACACACACACAC | ||
| 51 | AC28 | ACACACACACACACACACACACACACAC | ||
| 52 | AT-alt | ATATATATATATCCCATATATATATAT | dsDNA | |
| 53 | GC-alt | GCGCGCGCGCGCTTTGCGCGCGCGCGC | ||
| 54 | ctDNA | Unspecified calf thymus DNA | ||
| 55 | TFO1 | AAAAAAAACCCCTTTTTTTTCCCCTTTTTTTT | triplex | |
| 56 | TFO2 | AGAGAGAACCCCTTCTCTCTTATATCTCTCTT | ||
| 57 | VK1 | GGGAGCGAGGGAGCG | AG-tetraplex [29] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trizna, L.; Janovec, L.; Halaganová, A.; Víglaský, V. Rhodamine 6G-Ligand Influencing G-Quadruplex Stability and Topology. Int. J. Mol. Sci. 2021, 22, 7639. https://doi.org/10.3390/ijms22147639
Trizna L, Janovec L, Halaganová A, Víglaský V. Rhodamine 6G-Ligand Influencing G-Quadruplex Stability and Topology. International Journal of Molecular Sciences. 2021; 22(14):7639. https://doi.org/10.3390/ijms22147639
Chicago/Turabian StyleTrizna, Lukáš, Ladislav Janovec, Andrea Halaganová, and Viktor Víglaský. 2021. "Rhodamine 6G-Ligand Influencing G-Quadruplex Stability and Topology" International Journal of Molecular Sciences 22, no. 14: 7639. https://doi.org/10.3390/ijms22147639
APA StyleTrizna, L., Janovec, L., Halaganová, A., & Víglaský, V. (2021). Rhodamine 6G-Ligand Influencing G-Quadruplex Stability and Topology. International Journal of Molecular Sciences, 22(14), 7639. https://doi.org/10.3390/ijms22147639