
Interaction between Angiotensinase Activities in Pituitary and Adrenal Glands of Wistar–Kyoto and Spontaneously Hypertensive Rats under Hypotensive or Hypertensive Treatments

 , , , ,
, , , ,
Abstract
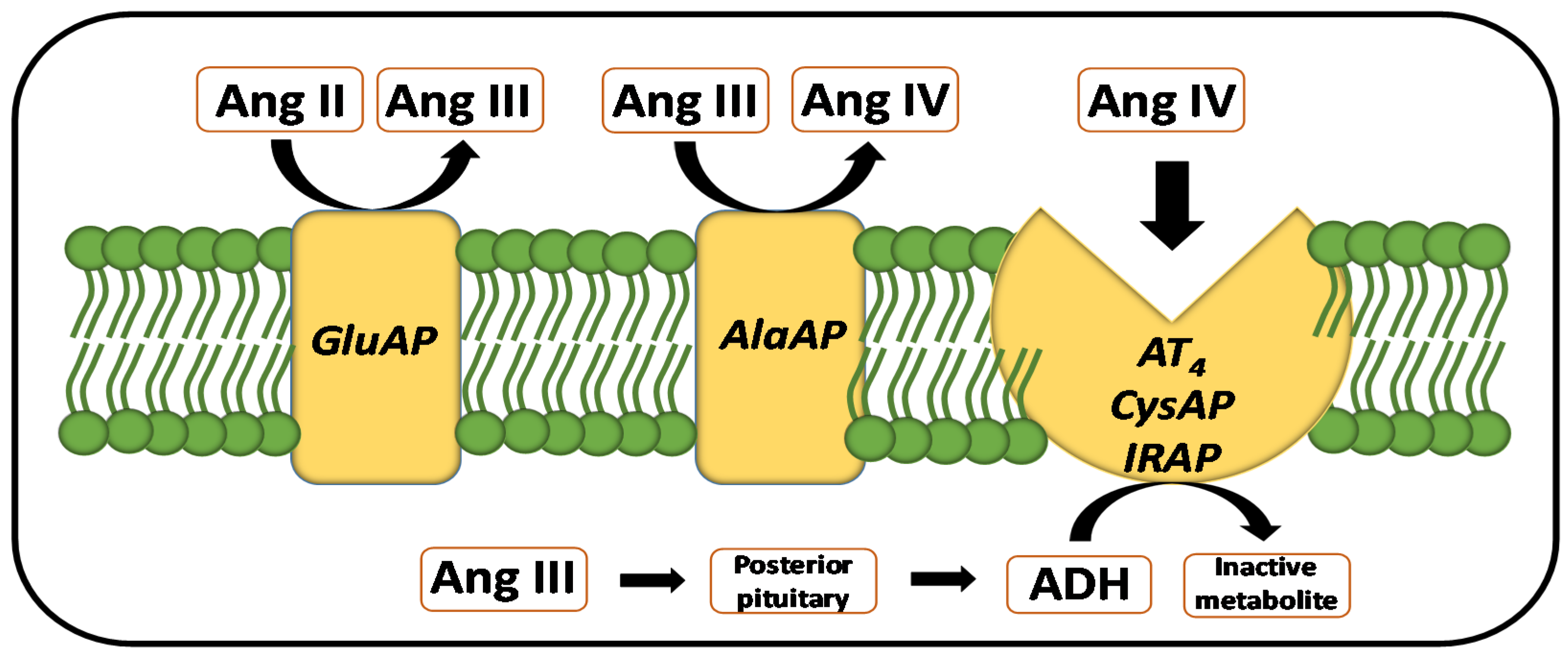
:1. Introduction
2. Results
2.1. Regional Distribution
2.2. Influence of Treatments
2.3. Intra- and Inter-Gland Correlations
2.4. Correlations between SBP and Aminopeptidase Activities
3. Discussion
4. Materials and Methods
4.1. Animals, Ethical Approval, and Drug Treatments
4.2. Blood Pressure Measurement
4.3. Surgery and Tissue Samples
4.4. Procedure for Protein and Enzymatic Assays
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | adrenals |
| ADH | antidiuretic hormone |
| AlaA | alanine aminopeptidase |
| Ang | angiotensin |
| AT4 | AT4 receptor |
| CAP | captopril |
| CysAP | cystine aminopeptidase |
| CT | control |
| GluAP | glutamate aminopeptidase |
| IRAP | insulin-regulated aminopeptidase |
| LN | L-NAME |
| PRO | propanolol |
| PT | pituitary |
| RAS | Renin-angiotensin system |
| SHR | spontaneously hypertensive rats |
| WKY | Wistar–Kyoto |
References
- Ramírez-Sánchez, M.; Prieto, I.; Wangensteen, R.; Banegas, I.; Segarra, A.B.; Villarejo, A.B.; Vives, F.; Cobo, J.; de Gasparo, M. The renin-angiotensin system: New insight into old therapies. Curr. Med. Chem. 2013, 20, 1313–1322. [Google Scholar] [CrossRef]
- de Gasparo, M.; Speth, R.C.; Baltatu, O.C.; Vanderheyden, P. Brain RAS: Hypertension and beyond. Int. J. Hypertens. 2013, 2013, 157180. [Google Scholar] [CrossRef]
- Arendse, L.B.; Danser, A.H.J.; Poglitsch, M.; Touyz, R.M.; Burnett, J.C., Jr.; Llorens-Cortes, C.; Ehlers, M.R.; Sturrock, E.D. Novel therapeutic approaches targeting the renin-angiotensin system and associated peptides in hypertension and heart failure. Pharmacol. Rev. 2019, 71, 539–570. [Google Scholar] [CrossRef] [Green Version]
- Llorens-Cortes, C.; Touyz, R.M. Evolution of a new class of antihypertensive drugs: Targeting the brain renin-angiotensin system. Hypertension 2020, 75, 6–15. [Google Scholar] [CrossRef]
- Ramírez, M.; Prieto, I.; Alba, F.; Vives, F.; Banegas, I.; de Gasparo, M. Role of central and peripheral aminopeptidase activities in the control of blood pressure: A working hypothesis. Heart Fail. Rev. 2008, 13, 339–353. [Google Scholar] [CrossRef]
- Albiston, A.L.; McDowall, S.G.; Matsacos, D.; Sim, P.; Clune, E.; Mustafa, T.; Lee, J.; Mendelsohn, F.A.; Simpson, R.J.; Connolly, L.M.; et al. Evidence that the angiotensin IV (AT(4)) receptor is the enzyme insulin-regulated aminopeptidase. J. Biol. Chem. 2001, 276, 48623–48626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prieto, I.; Villarejo, A.B.; Segarra, A.B.; Wangensteen, R.; Banegas, I.; de Gasparo, M.; Vanderheyden, P.; Zorad, S.; Vives, F.; Ramírez-Sánchez, M. Tissue distribution of CysAP activity and its relationship to blood pressure and water balance. Life Sci. 2015, 134, 73–78. [Google Scholar] [CrossRef]
- Stragier, B.; De Bundel, D.; Sarre, S.; Smolders, I.; Vauquelin, G.; Dupont, A.; Michotte, Y.; Vanderheyden, P. Involvement of insulin-regulated aminopeptidase in the effects of the renin-angiotensin fragment angiotensin IV: A review. Heart Fail. Rev. 2008, 13, 321–337. [Google Scholar] [CrossRef] [PubMed]
- Marc, Y.; Llorens-Cortes, C. The role of the brain renin-angiotensin system in hypertension: Implications for new treatment. Prog. Neurobiol. 2011, 95, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Edwards, D.G.; Berecek, K.H. Effects of early captopril treatment and its removal on plasma angiotensin converting enzyme (ACE) activity and arginine vasopressin in hypertensive rats (SHR) and normotensive rats (WKY). Clin. Exp. Hypertens. 1996, 18, 201–226. [Google Scholar] [CrossRef]
- Prieto, I.; Segarra, A.B.; de Gasparo, M.; Martínez-Cañamero, M.; Ramírez-Sánchez, M. Divergent profile between hypothalamic and plasmatic aminopeptidase activities in WKY and SHR. Influence of beta-adrenergic blockade. Life Sci. 2018, 192, 9–17. [Google Scholar] [CrossRef]
- Prieto, I.; Segarra, A.B.; Villarejo, A.B.; de Gasparo, M.; Martínez-Cañamero, M.M.; Ramírez-Sánchez, M. Neuropeptidase activity in the frontal cortex of Wistar-Kyoto and spontaneously hypertensive rats treated with vasoactive drugs: A bilateral study. J. Hypertens. 2019, 37, 612–628. [Google Scholar] [CrossRef]
- Segarra, A.B.; Prieto-Gomez, I.; Banegas, I.; Martínez-Cañamero, M.; Luna, J.D.; de Gasparo, M.; Ramírez-Sánchez, M. Functional and neurometabolic asymmetry in SHR and WKY rats following vasoactive treatments. Sci. Rep. 2019, 9, 16098. [Google Scholar] [CrossRef]
- Prieto, I.; Villarejo, A.B.; Segarra, A.B.; Banegas, I.; Wangensteen, R.; Martinez-Cañamero, M.; de Gasparo, M.; Vives, F.; Ramírez-Sánchez, M. Brain, heart and kidney correlate for the control of blood pressure and water balance: Role of angiotensinases. Neuroendocrinology 2014, 100, 198–208. [Google Scholar] [CrossRef]
- Segarra, A.B.; Prieto, I.; Banegas, I.; Martínez-Cañamero, M.; de Gasparo, M.; Ramírez-Sánchez, M. Blood pressure correlates asymmetrically with neuropeptidase activities of the left and right frontal cortices. Symmetry 2021, 13, 105. [Google Scholar] [CrossRef]
- Clough, D.P.; Hatton, R.; Keddie, J.R.; Collis, M.G. Hypotensive action of captopril in spontaneously hypertensive and normotensive rats. Interference with neurogenic vasoconstriction. Hypertension 1982, 4, 764–772. [Google Scholar] [CrossRef] [Green Version]
- Bhagat, B.D. Mechanism of the antihypertensive effect of propranolol. Gen. Pharmacol. 1979, 10, 291–296. [Google Scholar] [CrossRef]
- Pechanova, O.; Vrankova, S.; Cebova, M. Chronic L-Name-treatment produces hypertension by different mechanisms in peripheral tissues and brain: Role of central eNOS. Pathophysiology 2020, 27, 46–54. [Google Scholar] [CrossRef]
- Eshima, K.; Hirooka, Y.; Shigematsu, H.; Matsuo, I.; Koike, G.; Sakai, K.; Takeshita, A. Angiotensin in the nucleus tractus solitarii contributes to neurogenic hypertension caused by chronic nitric oxide synthase inhibition. Hypertension 2000, 36, 259–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priviero, F.B.; Teixeira, C.E.; Claudino, M.A.; De Nucci, G.; Zanesco, A.; Antunes, E. Vascular effects of long-term propranolol administration after chronic nitric oxide blockade. Eur. J. Pharmacol. 2007, 571, 189–196. [Google Scholar] [CrossRef]
- Domínguez-Vías, G.; Aretxaga-Maza, G.; Prieto, I.; Luna, J.D.; de Gasparo, M.; Ramírez-Sánchez, M. Diurnal opposite variation between angiotensinase activities in photo-neuro-endocrine tissues of rats. Chronobiol. Int. 2017, 34, 1180–1186. [Google Scholar] [CrossRef] [PubMed]
- Slaiby, J.M.; Ricci, M.A.; Gadowski, G.R.; Hendley, E.D.; Pilcher, D.B. Expansion of aortic aneurysms is reduced by propranolol in a hypertensive rat model. J. Vasc. Surg. 1994, 20, 178–183. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, M.; Prieto, I.; Banegas, I.; Segarra, A.B.; Alba, F. Neuropeptidases. Methods Mol. Biol. 2011, 789, 287–294. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WKY | SHR | ||||
|---|---|---|---|---|---|
| Correlation | r | p | Correlation | r | p |
| Control | |||||
| PT AlaAP vs.PT CysAP | +0.879 | 0.0007 | PT CysAP vs. AD GluAP | −0.751 | 0.03 |
| AD AlaAP vs. AD CysAP | +0.885 | 0.003 | |||
| Captopril | |||||
| PT AlaAP vs. PT CysAP | +0.744 | 0.01 | PT CysAP vs. AD AlaAP | +0.817 | 0.01 |
| AD CysAP vs. AD GluAP | −0.726 | 0.01 | PT CysAP vs. AD CysAP | +0.812 | 0.01 |
| PT CysAP vs. AD GluAP | +0.737 | 0.03 | |||
| AD AlaAP vs. AD CysAP | +0.949 | 0.0003 | |||
| AD AlaAP vs. AD GluAP | +0.830 | 0.01 | |||
| AD CysAP vs. AD GluAP | +0.742 | 0.03 | |||
| Propranolol | |||||
| PT AlaAP vs. PT CysAP | +0.631 | 0.03 | PT AlaAP vs. PT CysAP | +0.785 | 0.02 |
| PT AlaAP vs. PT GluAP | +0.768 | 0.005 | AD AlaAP vs. AD CysAP | +0.847 | 0.007 |
| PT AlaAP vs. AD CysAP | +0.624 | 0.03 | AD CysAP vs. AD GluAP | +0.712 | 0.04 |
| AD AlaAP vs. AD CysAP | +0.665 | 0.02 | |||
| AD AlaAP vs. AD GluAP | +0.876 | 0.0004 | |||
| AD CysAP vs. AD GluAP | +0.689 | 0.01 | |||
| L-NAME | |||||
| PT AlaAP vs. PT CysAP | +0.721 | 0.02 | AD AlaAP vs. AD CysAP | +0.909 | 0.001 |
| PT AlaAP vs. PT GluAP | +0.834 | 0.005 | |||
| PT CysAP vs. AD GluAP | −0.731 | 0.02 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Segarra, A.B.; Prieto, I.; Banegas, I.; Martínez-Cañamero, M.; Villarejo, A.B.; Domínguez-Vías, G.; de Gasparo, M.; Ramírez-Sánchez, M. Interaction between Angiotensinase Activities in Pituitary and Adrenal Glands of Wistar–Kyoto and Spontaneously Hypertensive Rats under Hypotensive or Hypertensive Treatments. Int. J. Mol. Sci. 2021, 22, 7823. https://doi.org/10.3390/ijms22157823
Segarra AB, Prieto I, Banegas I, Martínez-Cañamero M, Villarejo AB, Domínguez-Vías G, de Gasparo M, Ramírez-Sánchez M. Interaction between Angiotensinase Activities in Pituitary and Adrenal Glands of Wistar–Kyoto and Spontaneously Hypertensive Rats under Hypotensive or Hypertensive Treatments. International Journal of Molecular Sciences. 2021; 22(15):7823. https://doi.org/10.3390/ijms22157823
Chicago/Turabian StyleSegarra, Ana B., Isabel Prieto, Inmaculada Banegas, Magdalena Martínez-Cañamero, Ana B. Villarejo, Germán Domínguez-Vías, Marc de Gasparo, and Manuel Ramírez-Sánchez. 2021. "Interaction between Angiotensinase Activities in Pituitary and Adrenal Glands of Wistar–Kyoto and Spontaneously Hypertensive Rats under Hypotensive or Hypertensive Treatments" International Journal of Molecular Sciences 22, no. 15: 7823. https://doi.org/10.3390/ijms22157823
APA StyleSegarra, A. B., Prieto, I., Banegas, I., Martínez-Cañamero, M., Villarejo, A. B., Domínguez-Vías, G., de Gasparo, M., & Ramírez-Sánchez, M. (2021). Interaction between Angiotensinase Activities in Pituitary and Adrenal Glands of Wistar–Kyoto and Spontaneously Hypertensive Rats under Hypotensive or Hypertensive Treatments. International Journal of Molecular Sciences, 22(15), 7823. https://doi.org/10.3390/ijms22157823