Abstract
Metal oxide nanoparticles (MONPs) are inorganic materials that have become a valuable tool for many industrial sectors, especially in healthcare, due to their versatility, unique intrinsic properties, and relatively inexpensive production cost. As a consequence of their wide applications, human exposure to MONPs has increased dramatically. More recently, their use has become somehow controversial. On one hand, MONPs can interact with cellular macromolecules, which makes them useful platforms for diagnostic and therapeutic interventions. On the other hand, research suggests that these MONPs can cross the blood–testis barrier and accumulate in the testis. Although it has been demonstrated that some MONPs have protective effects on male germ cells, contradictory reports suggest that these nanoparticles compromise male fertility by interfering with spermatogenesis. In fact, in vitro and in vivo studies indicate that exposure to MONPs could induce the overproduction of reactive oxygen species, resulting in oxidative stress, which is the main suggested molecular mechanism that leads to germ cells’ toxicity. The latter results in subsequent damage to proteins, cell membranes, and DNA, which ultimately may lead to the impairment of the male reproductive system. The present manuscript overviews the therapeutic potential of MONPs and their biomedical applications, followed by a critical view of their potential risks in mammalian male fertility, as suggested by recent scientific literature.
1. Introduction
Nanotechnology is a field of science that studies the properties, design, manipulation, production, and applications of structures and devices at the nanoscale level (10−9 m). Objects on this scale, such as nanoparticles (NPs), have properties and functions that differ from those with a larger scale [1]. European and other International Committees have defined NPs, as particles of matter in which at least one of their phases has one dimension (length, width, or thickness) within the range of 1 to 100 nanometers (nm) [2,3].
Among the several types of NPs reported in the literature, metal oxide NPs (MONPs) stand out as the category of versatile materials. Being a type of metallic NPs that have controllable features and small size, making them able to easily cross cells and tissues within the body to reach a target location [4,5]. This makes MONPs a valuable tool for biomedical applications, such as anticancer, antidiabetic, antimicrobial purposes, as well as imaging applications, drug delivery, and even in reproductive medicine [6].
Most of these inorganic materials that make up MONPs are typically classified as biocompatible since their metallic precursors are already present in human tissues, whose vital role in body functions was reported [7,8]. Because they are essential to the body, they will be more readily accepted by the organism [6]. Manganese (Mn), molybdenum (Mo), magnesium (Mg), iron (Fe), cobalt (Co), chromium (Cr), copper (Cu), zinc (Zn), and selenium (Se) are among some of the elements considered essential for humans [6,7]. Some of these metals are closely related to male fertility. Zinc transduces a sign that induces sperm to become motile [9,10]. Selenium deficiency was previously associated with a decline in sperm motility [11]. Copper-dependent enzymes are present at all stages of spermatogenesis, as well as in somatic cells of the testis and epididymis [12]. However, in high concentrations, these physiologically compatible metals have toxic effects on mammalian cells and can even cause cell death [13]. Depending on how many metal ions are readily available, they can be beneficial or harmful, making their use a double-edged sword [6,12,14]. This may be part of the reason why there are so many controversial reports on the reproductive toxicity of MONPs [15]. In fact, it has been proven that MONPs can cross the blood–testis barrier (BTB), a structural and physiological compartment that protects spermatogenesis [16]. This raises concerns about male fertility, especially as spermatogenesis is a highly vulnerable process that is sensitive to exogenous materials, such as NPs [17,18]. Thus, addressing the effects of MONPs on the male reproductive system is crucial.
This review summarizes in vitro and in vivo studies that describe the potential reproductive toxicity of MONPs, to clarify the accurate effects of these NPs on the male reproductive system. Gaps in knowledge and ideas for future research are highlighted.
2. Classification of Nanoparticles and MONP Synthesis
NPs are versatile nanosized structures and, therefore, can be classified according to their dimensions, morphology, materials properties, origin, and synthesis process (Figure 1) [19]. Regarding their classification, all NPs share some aspects: they are known to have reduced size, which is related to their high surface area to volume ratio, they have chemically alterable physical properties, easy surface functionalization, and they all have different physical properties in respect to the bulk material [5,20,21].
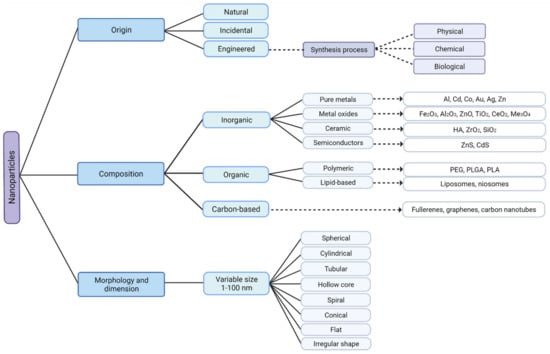
Figure 1.
Classification of nanoparticles according to their origin, composition, morphology, and dimension with some examples. Metal oxide nanoparticles are engineered, inorganic nanoparticles, that can be synthesized by physical, chemical, or biological techniques, created with Biorender.com (accessed on 27 June 2021).
Based on morphology and dimensions, NPs are typically spherical, but they can have many other shapes, such as cylindrical, tubular, conical, hollow core, spiral, flat, or irregular in shape with variable size [22,23].
Nowadays, NPs can be produced incidentally because of human activities, as a by-product of industrial and domestic endeavors that result in the unintentional release of NPs into the environment. On the other hand, engineered NPs with new properties may be synthesized by rearranging atoms of an object. However, NPs are not entirely a product of modern technology. Some exist in the natural world and can be found everywhere on earth, that is, in the hydrosphere, atmosphere, lithosphere, and biosphere. Therefore, regarding their origin, NPs can be classified as incidental, synthetic/engineered, or natural [24]. This emphasizes the idea that nanotechnology has become even more pervasive, and that NPs are ubiquitous in the environment, becoming more deeply embedded in today’s life.
According to properties of their materials, engineered NPs can be classified as carbon-based if they are made completely of carbon (e.g., fullerenes, graphene, carbon nanotubes), metal-based if NPs are made purely from metal precursors (e.g., Al, Cd, Co, Au Ag, Zn), metal oxides based if they have been synthesized to modify the properties of their respective metal based NPs (e.g., Fe2O3, Al2O3, ZnO), ceramic NPs if they are nonmetallic solids (e.g., HA, ZrO2, SiO2) and semiconductor NPs if they have properties between metals and nonmetals (e.g., ZnS, CdS) [22]. Polymeric NPs (e.g., PEG, PLGA, PLA) and lipid-based NPs (e.g., liposomes, niosomes), unlike those just mentioned, are generally organic [3,19,25].
There is a broad variety of techniques that can be used to synthesize MONPs, each with its own advantages and disadvantages. Generally, they can be arranged into physical, chemical, and biological (green synthesis) methods [26]. Biologically synthesized NPs are preferred in biomedical applications since they are safer than those produced by traditional physicochemical approaches. This can be attributed to the fact that the metallic core of NPs is coated with non-toxic biomolecules, making them biocompatible [27]. Additionally, in this method, the use of dangerous substances, such as organic solvents and inorganic salts—which are commonly used in physical and chemical methods—is minimized [28].
However, unlike other methods, this green approach has the drawback of being unable to control the size, shape, and yield of NPs [29]. Essentially, no single technique is ideal in all aspects or for all applications. Therefore, the desired application should be considered to select the most appropriate method.
The exact physical and chemical properties of NPs depend on the different ways in which they are produced, namely the synthesis process, external factors (reaction temperature, concentration of reagents and type of capping agents used), and internal factors (morphology, size, concentration) during their production [30]. These parameters, in turn, will determine the interaction of NPs with biological systems [29].
3. Biomedical Applications of MONPs
MONPs are inorganic materials made to modify the properties of metallic elements. These have been subjected to intense biomedical research, mainly due to their unique intrinsic properties, such as good optical, electrical, catalytic, and magnetic behavior, chemical and mechanical stability, simple preparation process, easily engineered for the desired size, shape and porosity, and large surface area for reactions [4,5,31]. In addition, these materials can easily have their surface modified with several chemical functional groups, allowing their conjugation with antibodies, ligands and drugs of interest, which further enhances their potential in the biomedical field [5]. Although there is a wide spectrum of metals available, their use in the medical field is limited to those tolerated by the organism [32]. The fact that some metals exist in appreciable amounts in the body makes most MONPs biocompatible. For example, in the human body, iron (3–4 g) is mainly found associated with hemoglobin, making it the most abundant metal [33,34]. Followed by iron, zinc (~2 g) [35], and copper (~0.1 g) [36] are the second and third most common metals in the human body, and they are essential constituents of several enzymes. Unlike the previous metals, manganese is present in very small amounts in the body (~12 mg). However, it is one of the most important nutrients for human health as it assists in the development of connective tissue, bones, blood-clotting factors, and sex hormones [33].
The use of MONPs to treat cancer, diabetes, and even to eradicate infectious diseases has been extensively studied, which proves the effort that has been made to create a symbiosis between nanoscience and medical science [31,37].
The common biomedical applications of MONPs and their main mechanisms of action are summarized in Figure 2.
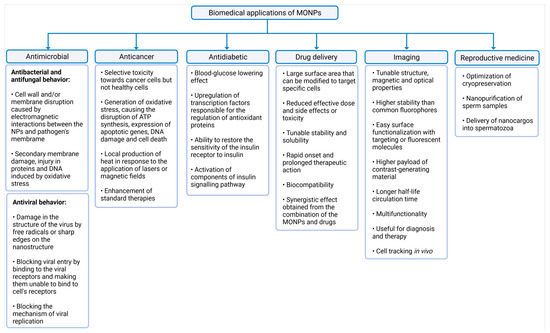
Figure 2.
Summary of the biomedical applications of MONPs. The latter were divided into six categories, namely antimicrobial activity, anticancer activity, antidiabetic activity, drug delivery, imaging, and reproductive medicine, created with Biorender.com (accessed on 27 June 2021).
3.1. Antimicrobial, Anticancer, and Antidiabetic Activity
Although in excessive doses many metals are toxic to all cell types, in lower concentrations, MONPs may be able to selectively target bacteria, since their metal transport system and metalloproteins are different from those existing in mammalian eukaryotic cells [38,39]. To exert this microbial function, MONPs need to be in contact with microbial cells. This interaction increases microbes’ membrane permeability, and allows the entry of NPs into the cytoplasm [38,40], where NPs induce damage to cellular macromolecules (Figure 2) [41]. This antimicrobial activity is enhanced for higher concentrations and smaller MONPs sizes [42,43], since smaller sizes allow a closer contact between NPs and the microbial membrane [4].
A wide range of MONPs seem to have antimicrobial abilities, including titanium dioxide (TiO2) [44], magnesium oxide (MgO) [45], zinc oxide (ZnO) [46], copper oxide (CuO) [47], iron oxide (Fe3O4) [48], cerium oxide (CeO) [49], and silver oxide (Ag2O) [50]. The molecular mechanisms of the antifungal activity of MONPs have been less studied because most studies have focused on antibacterial activity. Nonetheless, recent research suggests that these MONPs have similar mechanisms for bacteria and fungi [51].
Besides presenting antibacterial and antifungal activities, some MONPs also exert antiviral properties (Figure 2). MONPs can adhere to the virus envelope, causing its destruction [52], or they can block their mechanism of viral replication [53] or viral entry into a cell [54]. Metal oxides, such as TiO2 [52] and Cu2O [55], have already been shown to be effective antiviral agents against influenza A virus subtype H3N2 and Hepatitis C, respectively. These findings open a new perspective to prevent and treat viral diseases using MONPs.
MONPs can also selectively target cancer cells [56] and exert their anticancer activity mainly through the generation of oxidative stress [57]. This property can be further enhanced with the application of external stimuli such as magnetic fields or lasers, which induce the local production of heat in tumor sites [58]. Additionally, these NPs can also be used as enhancers of standard therapies, acting as co-adjuvants to improve the effect of radiation on radiotherapy, or to facilitate the action of conventional anticancer drugs, reducing the required dose and side effects of such drugs [59]. Therefore, different strategies take advantage of MONPs in the treatment of cancer: alone, conjugated with biological molecules, ligands, and anticancer drugs, or in combination with other conventional therapies to potentiate their therapeutic efficacy [60].
In addition, other MONPs such as MgO, MnO [61], CeO2 [62], ZnO [63,64], and Fe2O3 [65] have been explored as possible antidiabetic agents, since recent studies have shown promising results. Essentially, the antioxidant ability of MONPs contributes to a decrease in oxidative stress, which is the main cause of β-cell damage [66]. However, concentration determines whether NPs elicit oxidative stress or increase the cell antioxidant capacity. Generally, small doses seem to be related to the antidiabetic potential [14,65].
3.2. Drug Delivery Platforms and Imaging
Medical imaging is essential for medical diagnosis. MONPs have been used as nanoparticle-based contrast agents in multiple modern imaging modalities that allow the visualization of abnormalities, such as tumor lesions or other regions of interest [67]. Of all the plethora of available NPs, metal oxides have advantages in imaging applications due to their diverse size- and shape-dependent optoelectronic properties [27,68] and high stability, which are not achievable with traditional lipid or polymer-based nanoparticles [69]. In addition, compared to molecular probes, MONPs are virtually inert, which means that they hardly interact with other cellular molecules and, therefore, their optical properties remain unaffected [70]. Their surface can also be easily functionalized with drugs, targeting or fluorescent molecules, or other components [71,72]. Therefore, these contrast agents can deliver therapeutic agents simultaneously, allowing for a dual diagnostic and therapeutic effect [73].
Considering all this, MONPs are attractive imaging agents. As a result, they have been exploited for different imaging modalities, such as magnetic resonance imaging (MRI) [74], photoacoustic imaging (PA) [75], positron emission tomography (PET) [76], computed tomography (CT) [77], fluorescent imaging [78], among many others. In addition, NPs can be multifunctional and, therefore, can provide contrast for more than one imaging modality [23].
3.3. An Asset for Reproductive Medicine
Although the detrimental effects of NPs on male fertility and sperm cell function have been suggested [16], some research teams have been exploring the properties of these materials to improve assisted reproductive techniques. Falchi et al. reported that the incubation of ram semen with CeO2 NPs during cryopreservation improved sperm quantity and quality [79]. This study suggests that CeO2 NPs can have beneficial effects on sperm preservation. Other research teams have functionalized Fe2O3 NPs with lectins and antibodies, to selectively bind to glycans expressed in acrosome reaction, or to ubiquitin, which is present on the surface of defective spermatozoa [79,80]. Then, aberrant spermatozoa can be removed from a sample using a magnetic force. This method of sperm purification may be used to increase conception rates following artificial insemination [80]. Nanoplatforms for the delivery of biological compounds to spermatozoa are another nanotechnology that has been investigated in the field of reproductive medicine [15].
Makhluf et al. described the spontaneous penetration of polyvinyl alcohol (PVA)-Fe3O4 NPs in bovine sperm, without affecting their motility and ability to undergo the acrosome reaction [81]. These interesting results suggest that, in the future, NPs may be conjugated with target nutrients or treatments for direct nutrient supplementation to sperm.
These and other research teams have presented interesting results that highlight the usefulness of MONPs. However, despite these promising results, uncertainty remains about the safety of MONPs. Therefore, it is crucial to investigate in more detail how MONPs interact with the male reproductive system and the consequences of this exposure.
4. The Impact of MONPs on Male Fertility
MONPs have received a lot of attention, especially in the biomedical field, due to their biological usefulness, as discussed in previous sections. In addition, due to their unique properties and versatility, the application of NPs extends to many other fields, making them ubiquitous in the environment. Consequently, human exposure to nanomaterials has increased dramatically. However, in recent years, the use of NPs of any material has become controversial [82]. On one hand, MONPs can interact with cellular macromolecules, leading to therapeutic effects [83]. On the other hand, cytotoxic effects were found in some tissues, presenting a health hazard [84].
Many studies suggest that human male infertility has increased significantly over the past few decades [85,86,87]. Due to this alarming trend, it has been hypothesized that environmental, dietary, and/or lifestyle changes are interfering with men’s ability to produce spermatozoa with a consequent impact on male fertility [88,89]. In addition, the male reproductive system is known to be susceptible to environmental stress, as toxicants, vehicular pollutants, and even NPs [90]. As a result, the impact of MONPs on male reproductive health has become an important subject of study. While several reports suggest that some NPs might have protective effects on sperm cells [91], other reports suggest that they compromise male fertility by interfering with spermatogenesis [92]. In fact, spermatogenesis is prone to errors. Defects in any of its steps can result in the failure of the entire process and, in some cases, can lead to testicular diseases or male infertility [93,94].
Since spermatogenesis is a highly vulnerable process, it occurs in a protected environment, controlled by the BTB, whose purpose is to protect the developing germ cells from external insults [17]. It is formed by tight junctions between Sertoli cells that divide the epithelium of the seminiferous tubules (ST) into two different compartments: basal and adluminal (Figure 3). Although it is one of the tightest blood–tissue barriers in the mammalian body [95], it was previously reported that NPs could cross this biological barrier due to their ultra-small size [16]. In fact, in mice treated with TiO2 [96] and Fe2O3 [97], both NPs were able to penetrate the testis, despite the BTB. Takeda et al. even reported that TiO2 NPs accumulated in the testis of male offspring from pregnant mice who were treated with these NPs [98]. Other animal studies have also demonstrated that NPs can move from the initial absorption site, for example, the lungs and skin, to secondary organs, such as the testis [99]. The integrity of BTB is a concern since NPs can easily permeate cells and their nuclei. This creates favorable circumstances for mutations appearance, which in germ cells may interfere with fertilization, embryogenesis [100], or even generate congenital defects in the offspring [101].
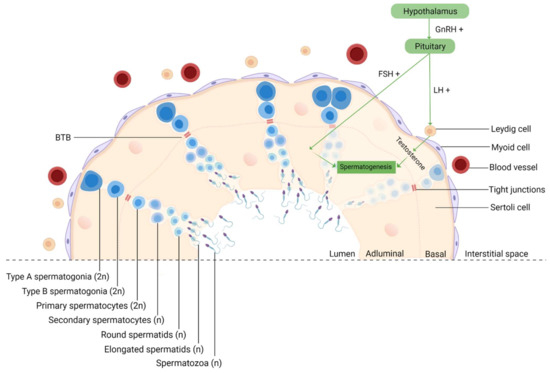
Figure 3.
Schematic representation of spermatogenesis in the cross-section of a seminiferous tubule. Spermatogenesis is initiated at puberty by the hypothalamus, which produces GnRH, which, in turn, stimulates the release of FSH and LH at the reproductive tract. LH stimulates Leydig cells to produce testosterone and FSH stimulates Sertoli cells that provide support and nutrition for sperm survival, proliferation, and differentiation [102]. Sertoli cells then initiate the functional responses required for spermatogenesis. Spermatogenesis starts when type A spermatogonia (2n) commit to differentiating into type B spermatogonia. Then, through mitosis, B-spermatogonia (2n) give rise to primary spermatocytes (2n). The latter undergo a long meiotic phase that originates the secondary spermatocytes (n), which ends with spermatids (n) generation [103]. The round spermatids then go through substantial morphological changes during spermiogenesis originating highly specialized spermatozoa through the reorganization of the entire cell, where the nuclear envelope seems to be crucially involved [104,105]. The next event is spermiation, in which mature spermatids are released from the supporting Sertoli cells into the lumen of the seminiferous tubule, and the remainder of the spermatid cytoplasm, known as the residual body, is phagocytosed by the Sertoli cells [106]. However, at this stage, spermatozoa still lack motility. Immotile spermatozoa are then transported into the epididymis where the final steps of maturation occur [107]. GnRH, gonadotropin-releasing hormone; LH, luteinizing hormone; FSH, follicle-stimulating hormone; BTB, blood–testis-barrier; 2n, diploid cell; n, haploid cell, created with Biorender.com (accessed on 2 July 2021).
Therefore, a clear understanding of the impact of MONPs on reproductive health is fundamental. Table 1 and Table 2 summarize the adverse effects of different MONPs on the male reproductive system, both in vitro and in vivo. However, it is important to keep in mind that these effects depend on several factors, such as dosage, duration of exposure, administration route, chemical nature of the compound (e.g., method of synthesis, size, shape, surface charge), as well as the biological system involved (e.g., strain and age of animal/cell, cell variability) [15].

Table 1.
In vitro studies of adverse effects of MONPs on mammalian male germ cells. The conditions where the main findings were observed are indicated in brackets.
Table 2.
In vivo studies of adverse effects of MONPs on the mammalian male reproductive system. The conditions where the main findings were observed are indicated in brackets.
4.1. In Vitro Studies
Few studies have focused on the adverse effects of NPs on male germ cells in vitro (Table 1).
The summary studies provide valuable information on the outcome of the interaction between MONPs and germ cells, which is useful for establishing the mechanisms of MONP toxicity. Parameters such as cell viability, oxidative stress, DNA damage, nanoparticle internalization, and mechanisms of cell death were assessed.
The in vitro studies reported in Table 1 were carried out with NPs made from Cerium (Ce), Iron (Fe), Manganese (Mn), Titanium (Ti), and Zinc (Zn) oxides. However, TiO2 and ZnO NPs are, by far, the most explored NPs.
The studies listed were conducted in different reproductive cells at three stages of maturation: spermatogonia, spermatocyte, and spermatozoa. Additionally, the cells responsible for testicular architecture and function, namely Sertoli and Leydig cells, were also used in the listed studies. In addition, most studies have carried out the extensive chemical and physical characterization of NPs, which is crucial for a better understanding of the toxicity mechanisms of NPs on reproductive cells.
A wide range of concentrations of MONPs has been studied, from very low (0.04 µg/mL) to high concentrations (1000 µg/mL). It is crucial to evaluate different concentrations of MONPs to establish their cytotoxic effect. However, the results still were conflicting. Préaubert et al. reported that the lowest concentrations of CeO2 NPs (0.01 µg/mL) were associated with higher levels of DNA damage in human spermatozoa [108]. ZnO NPs were also highly cytotoxic to mouse Leydig cells, even at low concentrations and incubation times [117]. Those are the exceptions since most studies indicate that MONP cytotoxicity is dose and time-dependent. Other authors even reported that lower MONPs concentrations were inefficient to cause genotoxicity [92,111].
The periods of incubation were also variable, ranging from 15 min to 24 h. From the results summarized in Table 1, it can be deduced that the reproductive toxicity of MONPs depends mainly on the concentration used and on the time of incubation.
The size of the NPs used ranges from ultrafine particles (7 nm) to much larger NPs (177 nm). Previous studies reported that even a small difference in size can make particles up to six times more harmful [119]. Gromadzka-ostrowska et al. also found that the toxicity of NPs is not only dependent on dose and time, but also depends on size, which seems to be inversely proportional to the cytotoxicity of NPs [120]. However, none of the studies reported in Table 1 evaluated the effect of the size of NPs on male germ cells.
The most studied parameters were oxidative stress indexes, cell viability, apoptosis, and genotoxicity. The principal suggested mechanism by which MONPs may exert that their toxic and genotoxic effect is oxidative stress [113,117]. In fact, increased oxidative stress was observed in almost all studies where this parameter was tested, except one [117]. Bara and Kaul reported an increase in the levels of antioxidant enzymes SOD and CAT in Leydig cells after exposure to ZnO NPs [117]. However, it has also been reported by other studies that NPs initially induce antioxidant enzyme activities in response to stress, as a defense mechanism, but, eventually, ROS production overcomes the capacity of the antioxidant response mechanisms [121].
Both exogenous stimuli and endogenous physiological stress can induce ROS production [117]. Oxidative stress is known to induce DNA damage through the oxidation of DNA bases [108] (Figure 4). However, it can also induce injury to biomolecules and organelles in other cells, mainly mitochondria [117]. In addition, under stress conditions, cells activate different cellular processes important for cell adaption to adverse conditions or to activate cell mechanisms of cell death, such as apoptosis or necrosis [117]. Pinho et al. reported an increase in the number of spermatogonia in necrosis (but not apoptosis) after ZnO NP exposure [92], while other studies have reported apoptosis as the preferred mechanism of cell death [110,117,118]. Autophagy is an example of an adaptive mechanism under stress conditions, and it was reported in Leydig cells after ZnO NPs exposure [118].
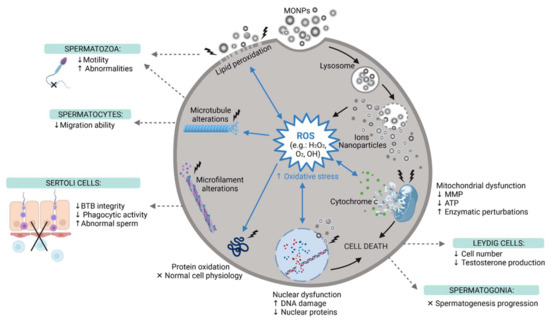
Figure 4.
The main reproductive toxic events induced by MONPs at the cellular level. MONPs, Metal Oxide Nanoparticles; ROS, Reactive Oxygen Species; MMP, Mitochondria Membrane Potential; ATP, Adenosine Triphosphate; BTB, Blood-Testis-Barrier; ↑, increase; ↓, decrease; x, impaired, created with Biorender.com (accessed on 30 May 2021).
The mechanism of MONPs internalization by cells was explored in some studies. Pawar and Kaul, using Scanning Electron Microscopy (SEM) and Transmission Electron Microscopy (TEM) images, reported that TiO2 in both agglomerated and single forms can remain attached to the spermatozoon surface (head and tail) after the addition of NPs to the sperm suspension, even after washing [111]. This indicates that NPs can attach and remain intact on the cell membrane immediately after mixing the NPs with the cell suspension. When in direct contact with cells, NPs cause mechanical damage to the membrane and destabilization of the plasma membrane, allowing NP entrance. The latter will exert pro-oxidant effects. In fact, Mao et al. monitored the internalization of TiO2 NPs by spermatocytes and Sertoli cells, both by flow cytometry and by TEM [112]. Bara and Kaul TEM results also revealed that ZnO NPs can enter Leydig cells and cross their nuclear membranes [117]. Moreover, Préaubert et al. also found an accumulation of CeO2 NPs at the spermatozoon plasma membrane [108]. However, in this case, the NPs were not internalized by the cells, but genotoxicity was still present. These authors proposed that MONPs do not need to be internalized to induce cell damage. To date, the exact mechanism by which the NPs induce cell damage is far to be elucidated, and, therefore, more comprehensive studies are needed.
Changes in the cytoskeleton were assessed only by Mao et al. and Pinho et al., using TiO2 NPs and ZnO NPs, respectively [92,112]. The latter reported disturbances in both microtubules and microfilament networks in spermatogonia cells [92]. Mao et al. also studied the effect of TiO2 NPs on the cytoskeleton of two different germ cells, spermatocytes, and Sertoli cells. TiO2 NPs interfered with microtubules of spermatocytes, but Sertoli cells only had their microfilaments altered [112]. These studies indicate that different germ cells respond differently to MONP insults. Additional studies should investigate alterations in the cytoskeleton since changes in the microtubule dynamics affect the formation of sperm flagella and migration abilities, and changes in the microfilament dynamics can affect the formation of tight junctions of Sertoli cells, which altogether interfere with spermatogenesis [112]. Although Liu et al. did not study cytoskeleton dynamics, their results indicate downregulation of tight junction proteins in Sertoli cells, leading to BTB impairment [116]. In addition, the disturbed microfilament arrangement interferes with the phagocytic capacity of Sertoli cells, which makes cells unable to properly phagocytose abnormal sperm cells [112].
Besides studying the cytoskeleton, Pinho et al. also reported, for the first time, the impact of ZnO NP exposure in the nucleoskeleton [92]. These authors reported several nuclear alterations in spermatogonia that may affect the progression of spermatogenesis.
Bara and Kaul was the only in vitro study to investigate the effect of NPs on steroidogenesis and testosterone biosynthesis in male reproductive cells. Interestingly, they found that a low concentration treatment with ZnO NPs for short incubation periods enhanced the steroidogenic ability of Leydig cells [117]. However, the exact mechanism is still unclear and therefore should be explored in future studies.
Overall, the interesting data collected indicate that the reproductive toxicity of NPs is not simply a matter of the NP material type, size, concentration, and exposure time, but also the result of intricate interactions at the nano-bio interface, which is influenced by many factors [13].
Since in vitro studies cannot consider tissue distribution, organs accumulation, clearance, or diffusion across biological barriers, such as the BTB, in vivo studies must be considered [122] and are of paramount importance to understand NP cytotoxicity.
4.2. In Vivo Studies
Table 2 lists the biochemical, molecular, and histopathological evidence of reproductive toxicity of MONPs. All MONPs that have been used in previous in vitro studies were also applied in in vivo studies. Considering the aluminum oxide (Al2O3) NPs, they have not been evaluated under cell culture conditions, only in vivo.
Table 2 clearly shows that animal models used for the in vivo experiments were mice and rats of different strains. Most of the studies listed have addressed the toxicity of MONPs at concentrations that are far from real-life conditions, even though there is no information available on the concentration of MONPs to which humans are exposed. Lauvås et al. used a lower and more realistic intratracheal dose of TiO2 NPs (63 µg/week for seven weeks), based on the estimated lung deposition of titanium at the Danish occupational exposure limit [137].
The exposure times used for the studies were highly variable. In some studies, male mammals received MONPs for very short periods, like 4 days [126], and, in other studies, the MONPs were used for much longer periods, namely six months [131]. These studies with these differences in exposure times are crucial since they help to create a better understanding of the acute and long-term effects of MONP administration. Additionally, many experiments have established the duration of treatment at around four weeks, to accomplish the duration of complete spermatogenesis in mice and rats [149].
Different routes of MONPS administration were used in animal experiments, namely, oral, intragastric, intratracheal, intraperitoneal, intravenous, and subcutaneous administration. It has been previously reported that there is very low absorption of MONPs through inhalation or oral administration in animals [62]. This aspect was confirmed by Lauvås et al., which was the only study included in Table 2, which administered MONPs intratracheally, and found that sperm cells are not susceptible to MONP exposure via the airways, at low doses [137]. On the other hand, in oral exposure, MONPs release more ions in the stomach due to the acidic environment. Therefore, this dissolution may be a reason for the cytotoxicity reported in the studies that used this administration approach, although fewer amounts of NPs are absorbed [145]. In contrast, intraperitoneal injections ensure proper absorption of the tested MONPs due to the highly vascularized peritoneal cavity [140]. The intravenous administration of nanomaterials ensures a much higher direct testicular exposure since NPs are administered directly into the bloodstream.
Regarding the parameters observed, most studies measured the weight of the male reproductive organs. Only Tang et al., Yousef et al. and Radhi et al. reported its increase after the oral administration of NPs, which may be attributed to the inflammation and hypertrophy or even accumulation of NPs in those tissues [90,123,147]. In fact, all studies that evaluated the content of MONPs in the testis and epididymis confirmed their presence in these organs. This was the case for cerium [124], iron [97], manganese [110], titanium [131,134,138], and zinc [90] NPs. The only exception was reported by Miura et al. studies, in which TiO2 NPs administered intravenously were found in the testis, but not in significant amounts [134,138]. This deposition of NPs in the reproductive tissues triggers the harmful events that will be described throughout this section. In fact, the damage has been reported in the testis and epididymis. Al2O3 [123], F2O3 [125], Fe3O4 [126], Mn3O4 [110,128], MnO2 [129], TiO2 [131,132,135,136,139,140], and ZnO [123,140,141,143,144,146] NPs all caused histopathological changes in the testis, mainly due to degeneration of the seminiferous tubules. Furthermore, Morgan et al. studied the histopathological changes induced by TiO2 NPs in the prostate and seminal vesicle, and reported that these reproductive organs were also affected by NPs, since they caused congestion, hyperplasia, and desquamation of the prostate’s epithelial, lining, and congestion in the seminal vesicle [133]. Salman also reported that ZnO NPs caused mild damage in seminal vesicles but severe damage to the prostate [148]. The reduction in the testis cell population has also been commonly reported, which is an indicator of a lack of active spermatogenesis in the testis [150].
The translocation of MONPs from their site of administration to the testicular tissue confirms that these NPs can cross and enter the BTB, where they interfere with normal physiological processes. Then, when in contact with reproductive tissues, these NPs can permeate cell membranes, inducing the overproduction of ROS, which leads to oxidative stress (Figure 4). This interferes with the balance between the oxidant and antioxidant systems, which causes oxidative damage in biomolecules, such as lipids, proteins, and nucleic acids [97]. To confirm the oxidative damage caused by MONPs, different studies evaluated ROS production and the levels of other oxidant markers, such as Malondialdehyde (MDA), Nitric Oxide (NO), Protein Carbonyl Content (PC), Lipid Peroxidation (LPO), and Total Oxidant Status (TOS). Antioxidant parameters such as Superoxide Dismutase (SOD), Glutathione Peroxidase (GPx), Reduced Glutathione (GSH), Catalase (CAT), and Total Antioxidant Capacity (TAC), were also evaluated. These parameters of oxidative stress were assessed on all types of MONPs, except CeO2 NPs [124]. The results reported an increase in oxidant markers and a decrease in intracellular antioxidant defenses and TAC. This confirms that MONPs suppress the antioxidant machinery and induce oxidative stress, which can lead to various cellular damages and, consequently, interfere with male fertility. In fact, according to previous studies, 30–80% of male infertility cases can be attributed to oxidative stress-mediated injury to the male reproductive system [110,151,152]. Persistent oxidative stress leads to the downregulation of Bcl-2 and upregulation of Bax, which results in the leakage of cytochrome c from dysfunctional mitochondria, ultimately resulting in apoptosis (Figure 4), through the activation of caspase molecules, as confirmed by Sundarraj et al. Meena et al., Shen et al. and Morgan et al. [97,118,132,135]. MONPs not only induce apoptosis, but some have also proven to be autophagy activators and inducers of autophagic cell death [118].
The levels of endocrine and reproductive hormones were also evaluated, and the results also suggest an imbalance in reproductive hormones (Testosterone, FSH, LH, GnRH, E2) and thyroid hormones (TSH, T3, T4) that can be attributed to the increase of ROS and the concomitant reduction of antioxidant enzymes. The exceptions were Lauvås et al. and Ogunsuyi et al., who reported that TiO2 NPs did not trigger alterations in testosterone levels [137,140]. Contrarily, Miura et al. reported that TiO2 NPs affected testosterone levels, but not FSH, LH, and GnRH [134].
Additionally, some authors explored the influence of MONPs on the expression of genes related to steroidogenesis. Testosterone is produced mainly in Leydig cells by a series of enzymatic reactions. First, the StAR protein transfers cholesterol to mitochondria. Then, the mitochondrial cytochrome P450scc transforms cholesterol into pregnenolone. Subsequently, other enzymes (3β-HSD, P450c17, 17β-HSD) convert the pregnenolone into testosterone [124]. Interestingly, Nr5A1, a transcription factor that regulates the expression of steroidogenic genes in Leydig cells (such as 3β-HSD), was downregulated after exposure to ZnO NP [144]. The StAR protein was also downregulated by CeO2 [124] and ZnO NPs [90], which can manifest in their inability to transfer cholesterol to the inner mitochondrial membrane, which stops steroidogenesis and justifies the decline in testosterone levels in most of the results listed. However, Bara and Kaul reported the conflicting results of increased testosterone production and StAR upregulation, but this was only related to small concentrations of ZnO NPs [117]. Ogunsuyi et al. did not report alterations in testosterone levels after intraperitoneal administration of TiO2 NPs; however, these levels were increased in the same study, under the same conditions, by ZnO NPs [140]. Likewise, Lauvås et al. found no significant alterations in testosterone levels after intratracheal administration of TiO2 NPs [137].
Sperm parameters, such as sperm number, viability, abnormalities, and motility, have been extensively studied. All studies that analyzed sperm count observed its decline with increasing concentrations of NPs, except for Varzeghani et al., Lauvås et al. and Song et al., who did not report significant alterations [126,136,137]. The results listed in Table 2 also indicate a reduction in motile spermatozoa, which affects their fertilizing potential. This decrease in sperm motility may have been a result of lipid peroxidation [140] (Figure 4). In addition, Morgan et al., Hussein et al., Srivastav et al. and Abbasalipourkabir et al. were the only research teams that evaluated sperm viability, having reported its decline [133,135,142,144,145]. An increase in sperm abnormalities, such as small head, double head, formless head, and double tails, has also been reported, which may be the result of oxidative damage [140] (Figure 4). These results are in agreement with those reported under in vitro conditions (Table 1).
Hong et al. evaluated the activity of metabolism-related enzymes—LDH, SDH, and SODH—that play key roles in the growth and development of testicular cells [130]. The results suggest that there was a decline in their activity, which may be associated with the disturbance of energy metabolism in germ cells. It was also the only study to evaluate the testicular activity of G-6PD, testis-marker enzymes ACP and AKP, and the activity of Ca2+-ATPase, Ca2+/Mg2+-ATPase and Na+/K+-ATPase. G-6PD is associated with androgen biogenesis, and its reduction implies that TiO2 NPs interfered with androgen secretion. In this study, ACP and AKP were used as markers of impaired spermatogenesis. Since ACP is related to the degeneration of the seminiferous epithelium and AKP is related to the activity of division of germ cells, their increase suggests testicular degeneration. Reductions in ATPases suggest an imbalance in the concentrations of intracellular ions, which could promote spermatogenesis dysfunctions [130].
Due to their small size, MONPs can reach the nucleus and interact directly with DNA, which causes the generation of ROS that further damages DNA (Figure 4) [146]. Not all studies tested the genotoxicity of NPs, but all studies that evaluated DNA damage later confirmed it. Mesallam et al. detected DNA fragmentation in the testis and prostate of rats treated with 422 mg/kg ZnO NPs daily for four weeks [146]. Meena et al. also found DNA strand breaks in spermatozoa of rats treated with 25 and 50 mg/kg TiO2 NPs weekly for 30 days [132].
Results also indicate elevated levels of TNF-α [123,146], and pro-inflammatory IL-6 cytokine [123], and a decrease in anti-inflammatory IL-4 cytokine [146] in reproductive tissues, which indicates a cellular inflammatory response to the NP exposure.
Zhang et al. evaluated male fertility by assessing the offspring of rats treated with Mn3O4 NPs [110]. The obtained results confirmed that this treatment decreased rats’ fertility and reduced the survival rate of their offspring in a time-dependent manner. For these authors, these results are attributed to changes in reproductive hormones and the decline in sperm quality [110].
In summary, most biochemical and molecular results were concomitant with histological findings. Therefore, despite the many benefits of MONPs, the results of the listed in vivo studies confirm the in vitro studies, emphasizing the possibility that exposure to these NPs could have a detrimental impact on male fertility.
4.3. MONPs in Human Reproductive Medicine
The recent approval of MONPs-based technologies in clinical medicine allowed an increase in human living standards and an improvement in mankind’s healthcare conditions through the prevention, early detection, diagnosis, treatment, and follow-up of multiple diseases [153]. However, their usefulness in human reproductive medicine has yet to be proved.
Considering that 50% of infertile couples, the male partner is affected by aberrations in sperm properties, number, vitality, and morphology [154], there is a clear need to develop novel methodologies for the early identification of infertility causes and its treatment. Some research teams have already developed MONP-based approaches that were tested in vitro and in vivo, with promising results. These include methods to reduce oxidative stress induced by cryopreservation [155], improve the proportion of healthy spermatozoa in semen prior to insemination [156], provide movement to sperm with motility deficit [157], protect the fertility of men who are exposed to fertility disrupters [158], and even treat other male associated disorders, such as erectile dysfunction [159].
Although these and other approaches have shown promising results, most of the literature still suggests uncertainty regarding the risk of MONPs in fertility, which may be one of the main reasons why, to date, there are no trials involving this type of engineered NPs for fertility regulation and treatment of male reproductive diseases. Another limiting factor is that only a few studies tried to identify the exact mechanism and pathways induced by MONPs. Current animal experiments also fail to assess pregnancy rates, and the health of offspring, which is the most relevant outcome parameter of fertility [160]. This gap in literature allows the speculation around the hazard posed by MONPs, which could prevent the translation of the results from the lab to the clinical applications [161]. NPs represent a valuable tool to alleviate much of the suffering arising from many reproductive difficulties and disorders, but further work is required to determine if these NPs can fulfill the needs in reproductive health. Human clinical reproductive trials may help accelerate the commercial availability of these new alternatives.
5. Conclusions and Future Perspectives
The increased application of MONPs in many industries and scientific fields has made these materials highly present in the environment, resulting in an increased risk of human exposure. Additionally, evidence that keeps emerging suggests that MONPs interfere with the male reproductive system at many biological levels.
The results presented in this review from both in vitro and in vivo studies prove that MONPs can interfere with the male reproductive system, and these results should not be ignored. The collected data show that this reproductive toxicity is achieved due to the MONPs’ ability to interfere with cell molecules and reproductive hormones, which often results in DNA damage and altered gene expression. It was also reported that MONPs induce oxidative stress in germ cells, which affects their number, quality, morphology, and activity. At the organ level, MONPs can cross the BTB and accumulate in the testis, resulting in many histological alterations in tissues of the reproductive system. Since the normal physiological processes that occur in the male reproductive system are highly complex and vulnerable, the interference of MONPs at any level can be deleterious and impair male fertility. Whether these harmful effects are reversible or not is still unclear and should be investigated in further research. How these alterations affect pregnancy and offspring is still an unresolved issue and should be addressed in future studies.
In the studies presented, the only conditions considered to evaluate the reproductive toxicity of MONPs were concentration and duration of exposure. However, the size and surface area are two crucial physical properties that affect how MONPs interact with cells and thus greatly determine the cytotoxicity of NPs. In addition, current studies generally focus on individual alterations but fail to establish a relationship between them. This may be partly the reason why the exact mechanism of nanotoxicity is not yet fully elucidated. Therefore, future studies should make a more in-depth examination of the molecular mechanisms of NPs and MONPs, in particular in reproductive toxicity and the interaction between each reported alteration. In addition, the in vivo studies are of significant heterogeneity, mainly due to the difference in the route of administration and the highly variable administered doses and exposure times. All of these factors can potentially be a source of toxicity that may influence the outcome of the studies. In some cases, unrealistically high concentrations of MONPs were used in cell culture and animal studies, which obviously results in cytotoxicity. Those studies lead to discouraging results that affect the accurate estimation of the reproductive health risks and hinder clinical translation.
It is reasonable to conclude that there are still difficulties in evaluating the reproductive toxicity of MONPs and in understanding exactly how they interact with the male reproductive system. The results summarized in this review reinforce the need for further studies with uniform protocols to obtain solid results with real implications in humans.
Author Contributions
Conceptualization, S.R. and M.d.L.P.; methodology, M.V.; software, M.V.; formal analysis, M.V.; investigation, M.V.; resources, S.R. and M.d.L.P.; writing—original draft preparation, M.V.; writing—review and editing, S.R., M.d.L.P., and M.V.; supervision, S.R. and M.d.L.P.; project administration, S.R. and M.d.L.P.; funding acquisition, S.R. and M.d.L.P.; All authors have read and agreed to the published version of the manuscript.
Funding
This work was sponsored by Project CICECO-Aveiro Institute of Materials, UIDB/50011/2020 and UIDP/50011/2020, national funds by the Portuguese Foundation for Science and Technology/MCTES. This research was also funded by the Institute of Biomedicine (iBiMED) UID/BIM/04501/2020.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McNeil, S.E. Nanotechnology for the biologist. J. Leukoc. Biol. 2005, 78, 585–594. [Google Scholar] [CrossRef]
- Lövestam, G.; Rauscher, H.; Roebben, G.; Klüttgen, B.; Gibson, N.; Putaud, J.-P.; Stamm, H. Considerations on a Definition of Nanomaterial for Regulatory Purposes; JRC Reference Reports; Publications Office of the European Union: Luxembourg, 2010. [Google Scholar]
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, applications and toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Singla, R.; Guliani, A.; Kumari, A.; Yadav, S.K. Metallic Nanoparticles, Toxicity Issues and Applications in Medicine; Springer: Singapore, 2016; ISBN 9789811008184. [Google Scholar]
- Nikolova, M.P.; Chavali, M.S. Metal oxide nanoparticles as biomedical materials. Biomimetics 2020, 5, 27. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, J.; Ghosh, S.; Datta, P.; Gomes, A.; Gomes, A. Physiologically important metal nanoparticles and their toxicity. J. Nanosci. Nanotechnol. 2014, 14, 990–1006. [Google Scholar] [CrossRef] [PubMed]
- Zoroddu, M.A.; Aaseth, J.; Crisponi, G.; Medici, S.; Peana, M.; Nurchi, V.M. The essential metals for humans: A brief overview. J. Inorg. Biochem. 2019, 195, 120–129. [Google Scholar] [CrossRef]
- Das, R.K.; Brar, S.K.; Verma, M. Checking the Biocompatibility of Plant-Derived Metallic Nanoparticles: Molecular Perspectives. Trends Biotechnol. 2016, 34, 440–449. [Google Scholar] [CrossRef]
- Zhao, J.; Dong, X.; Hu, X.; Long, Z.; Wang, L.; Liu, Q.; Sun, B.; Wang, Q.; Wu, Q.; Li, L. Zinc levels in seminal plasma and their correlation with male infertility: A systematic review and meta-analysis. Sci. Rep. 2016, 6, 22386. [Google Scholar] [CrossRef] [PubMed]
- Fallah, A.; Mohammad-Hasani, A.; Colagar, A. Zinc is an Essential Element for Male Fertility: A Review of Zn Roles in Men’s Health, Germination, Sperm Quality, and Fertilization. J. Reprod. Infertil. 2018, 19, 69–81. [Google Scholar] [PubMed]
- Rezaeian, Z.; Yazdekhasti, H.; Nasri, S.; Rajabi, Z.; Fallahi, P.; Amidi, F. Effect of selenium on human sperm parameters after freezing and thawing procedures. Asian Pac. J. Reprod. 2016, 5, 462–466. [Google Scholar] [CrossRef]
- Herman, S.; Lipinski, P.; Ogórek, M.; Starzynski, R.; Grzmil, P.; Bednarz, A.; Lenartowicz, M. Molecular Regulation of Copper Homeostasis in the Male Gonad during the Process of Spermatogenesis. Int. J. Mol. Sci. 2020, 21, 9053. [Google Scholar] [CrossRef] [PubMed]
- Taylor, U.; Barchanski, A.; Kues, W.; Barcikowski, S.; Rath, D. Impact of Metal Nanoparticles on Germ Cell Viability and Functionality Production of Metal Nanoparticles. Reprod. Domest. Anim. 2012, 47, 359–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umrani, R.D.; Paknikar, K.M. Zinc oxide nanoparticles show antidiabetic activity in streptozotocin- induced Type 1 and 2 diabetic rats. Nanomedicine 2014, 9, 89–104. [Google Scholar] [CrossRef]
- Falchi, L.; Khalil, W.A.; Hassan, M.; Marei, W.F.A. Perspectives of nanotechnology in male fertility and sperm function. Int. J. Vet. Sci. Med. 2018, 6, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, A.H.; Possebon, L.; Costa, S.D.S.; Souza, H.; Girol, A.; Pereira, M.d.L. Adverse effects of Metal-based Nanoparticles on Male Reproductive Cells. In Top 10 Contributions on Environmental Healt; Avid Science: Berlin, Germany, 2018; pp. 1–19. [Google Scholar]
- Matsumoto, A.M.; Bremner, W.J. Testicular Disorders, 13th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Zhou, Q.; Yue, Z.; Li, Q.; Zhou, R.; Liu, L. Exposure to PbSe Nanoparticles and Male Reproductive Damage in a Rat Model. Environ. Sci. Technol. 2019, 53, 13408–13416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ijaz, I.; Gilani, E.; Nazir, A.; Bukhari, A. Detail review on chemical, physical and green synthesis, classification, characterizations and applications of nanoparticles. Green Chem. Lett. Rev. 2020, 13, 59–81. [Google Scholar] [CrossRef]
- Vaseem, M.; Umar, A.; Hahn, Y. ZnO Nanoparticles: Growth, Properties, and Applications; American Scientific Publishers: Los Angeles, CA, USA, 2010; Volume 5, ISBN 1588831701. [Google Scholar]
- Reverberi, A.P.; Kuznetsov, N.T.; Meshalkin, V.P.; Salerno, M.; Fabiano, B. Systematical analysis of chemical methods in metal nanoparticles synthesis. Theor. Found. Chem. Eng. 2016, 50, 59–66. [Google Scholar] [CrossRef]
- Ealias, A.M.; Saravanakumar, M.P. A review on the classification, characterisation, synthesis of nanoparticles and their application. IOP Conf. Ser. Mater. Sci. Eng. 2017, 263, 032019. [Google Scholar] [CrossRef]
- Cormode, D.; Naha, P.; Fayad, Z. Nanoparticle contrast agents for computed tomography: A focus on micelles. Contrast Media Mol. Imaging 2014, 9, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Jeevanandam, J.; Barhoum, A.; Chan, Y.S.; Dufresne, A.; Danquah, M.K. Review on nanoparticles and nanostructured materials: History, sources, toxicity and regulations. Beilstein J. Nanotechnol. 2018, 9, 1050–1074. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Vega, A.I.; Gómez-Quintero, T.; Nuñez-Anita, R.E.; Acosta-Torres, L.S.; Castaño, V. Polymeric and ceramic nanoparticles in biomedical applications. J. Nanotechnol. 2012, 2012, 936041. [Google Scholar] [CrossRef] [Green Version]
- Chavali, M.S.; Nikolova, M.P. Metal oxide nanoparticles and their applications in nanotechnology. SN Appl. Sci. 2019, 1, 607. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, F.; Ashraf, N.; Ashraf, T.; Zhou, R.B.; Yin, D.C. Biological synthesis of metallic nanoparticles (MNPs) by plants and microbes: Their cellular uptake, biocompatibility, and biomedical applications. Appl. Microbiol. Biotechnol. 2019, 103, 2913–2935. [Google Scholar] [CrossRef] [PubMed]
- Das, R.K.; Laxman, V.; Linson, P. Biological synthesis of metallic nanoparticles: Plants, animals and microbial aspects. Nanotechnol. Environ. Eng. 2017, 2, 18. [Google Scholar] [CrossRef] [Green Version]
- Stankic, S.; Suman, S.; Haque, F.; Vidic, J. Pure and multi metal oxide nanoparticles: Synthesis, antibacterial and cytotoxic properties. J. Nanobiotechnology 2016, 14, 73. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Rajput, J.; Kaith, B.S.; Kaur, M.; Sharma, S. Synthesis of ZnO nanoparticles and study of their antibacterial and antifungal properties. Thin Solid Films 2010, 519, 1224–1229. [Google Scholar] [CrossRef]
- Pinho, A.; Rebelo, S.; Pereira, M. The Impact of Zinc Oxide Nanoparticles on Male (In)Fertility. Materials 2020, 13, 849. [Google Scholar] [CrossRef] [Green Version]
- Augustine, R.; Mathew, A.P.; Sosnik, A. Metal Oxide Nanoparticles as Versatile Therapeutic Agents Modulating Cell Signaling Pathways: Linking Nanotechnology with Molecular Medicine. Appl. Mater. Today 2017, 7, 91–103. [Google Scholar] [CrossRef]
- Al-fartusie, F.S.; Mohssan, S.N. Trace Elements and Their Vital Roles in Human Body. Indian J. Adv. Chem. Sci. 2017, 5, 127–136. [Google Scholar] [CrossRef]
- Huxford, R.C.; Della Rocca, J.; Lin, W. Metal-organic frameworks as potential drug carriers. Curr. Opin. Chem. Biol. 2010, 14, 262–268. [Google Scholar] [CrossRef] [Green Version]
- Raven, E.; Le Brun, N.E.; McMaster, J.; Reedijk, J.; Robinson, N.J. Bioinorganic Chemistry. Dalton Trans. 2013, 42, 3027–3028. [Google Scholar] [CrossRef]
- Bost, M.; Houdart, S.; Oberli, M.; Kalonji, E.; Huneau, J.F.; Margaritis, I. Dietary copper and human health: Current evidence and unresolved issues. J. Trace Elem. Med. Biol. 2016, 35, 107–115. [Google Scholar] [CrossRef]
- Venkatachalam, M.; Govindaraju, K.; Mohamed Sadiq, A.; Tamilselvan, S.; Ganesh Kumar, V.; Singaravelu, G. Functionalization of gold nanoparticles as antidiabetic nanomaterial. Spectrochim. Acta-Part A Mol. Biomol. Spectrosc. 2013, 116, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Gold, K.; Slay, B.; Knackstedt, M.; Gaharwar, A.K. Antimicrobial Activity of Metal and Metal-Oxide Based Nanoparticles. Adv. Ther. 2018, 1, 1700033. [Google Scholar] [CrossRef]
- Lemire, J.A.; Harrison, J.J.; Turner, R.J. Antimicrobial activity of metals: Mechanisms, molecular targets and applications. Nat. Rev. Microbiol. 2013, 11, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Shahzadi, S.; Zafar, N.; Sharif, R. Antibacterial Activity of Metallic Nanoparticles. In Bacterial Pathogenesis and Antibacterial Control; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Beyth, N.; Houri-Haddad, Y.; Domb, A.; Khan, W.; Hazan, R. Alternative antimicrobial approach: Nano-antimicrobial materials. Evid.-Based Complement. Altern. Med. 2015, 2015, 246012. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Hu, C.; Shao, L. The-antimicrobial-activity-of-nanoparticles—Present-situati. Int. J. Nanomed. 2017, 12, 1227–1249. [Google Scholar] [CrossRef] [Green Version]
- Talebian, N.; Amininezhad, S.M.; Doudi, M. Controllable synthesis of ZnO nanoparticles and their morphology-dependent antibacterial and optical properties. J. Photochem. Photobiol. B Biol. 2013, 120, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Jesline, A.; John, N.P.; Narayanan, P.M.; Vani, C.; Murugan, S. Antimicrobial activity of zinc and titanium dioxide nanoparticles against biofilm-producing methicillin-resistant Staphylococcus aureus. Appl. Nanosci. 2015, 5, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.Y.T.; Grelling, N.; Wetteland, C.L.; Rosario, R.; Liu, H. Antimicrobial Activities and Mechanisms of Magnesium Oxide Nanoparticles (nMgO) against Pathogenic Bacteria, Yeasts, and Biofilms. Sci. Rep. 2018, 8, 16260. [Google Scholar] [CrossRef] [Green Version]
- Navale, G.R.; Thripuranthaka, M.; Late, D.J.; Shinde, S.S. Antimicrobial Activity of ZnO Nanoparticles against Pathogenic Bacteria and Fungi. JSM Nanotechnol. Nanomed. 2015, 3, 1033. [Google Scholar]
- Mageshwari, K.; Sathyamoorthy, R. Flower-shaped CuO Nanostructures: Synthesis, Characterization andAntimicrobial Activity. J. Mater. Sci. Technol. 2013, 29, 909–914. [Google Scholar] [CrossRef]
- Anghel, I.; Grumezescu, A.M.; Holban, A.M.; Ficai, A.; Anghel, A.G.; Chifiriuc, M.C. Biohybrid nanostructured iron oxide nanoparticles and Satureja hortensis to prevent fungal biofilm development. Int. J. Mol. Sci. 2013, 14, 18110–18123. [Google Scholar] [CrossRef]
- Farias, I.; Santos, C.; Sampaio, F. Antimicrobial Activity of Cerium Oxide Nanoparticles on Opportunistic Microorganisms: A Systematic Review. BioMed Res. Int. 2018, 2018, 1923606. [Google Scholar] [CrossRef] [Green Version]
- Allahverdiyev, A.M.; Abamor, E.S.; Bagirova, M.; Rafailovich, M. Antimicrobial effects of TiO2 and Ag2O nanoparticles against drug-resistant bacteria and leishmania parasites. Future Microbiol. 2011, 6, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Parveen, S.; Wani, A.H.; Shah, M.A.; Devi, H.S.; Bhat, M.Y.; Koka, J.A. Preparation, characterization and antifungal activity of iron oxide nanoparticles. Microb. Pathog. 2018, 115, 287–292. [Google Scholar] [CrossRef]
- Mazurkova, N.A.; Spitsyna, Y.E.; Shikina, N.V.; Ismagilov, Z.R.; Zagrebel’nyi, S.N.; Ryabchikova, E.I. Interaction of titanium dioxide nanoparticles with influenza virus. Nanotechnologies Russ. 2010, 5, 417–420. [Google Scholar] [CrossRef]
- Brandelli, A.; Ritter, A.C.; Veras, F. Antimicrobial Activities of Metal Nanoparticles. In Metal Nanoparticles in Pharma; Springer: Berlin/Heidelberg, Germany, 2017; ISBN 9783319637907. [Google Scholar]
- Meléndez-Villanueva, M.A.; Morán-Santibañez, K.; Martínez-Sanmiguel, J.J.; Rangel-López, R.; Garza-Navarro, M.A.; Rodríguez-Padilla, C.; Zarate-Triviño, D.G.; Trejo-Ávila, L.M. Virucidal activity of gold nanoparticles synthesized by green chemistry using garlic extract. Viruses 2019, 11, 1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hang, X.; Peng, H.; Song, H.; Qi, Z.; Miao, X.; Xu, W. Antiviral activity of cuprous oxide nanoparticles against Hepatitis C Virus in vitro. J. Virol. Methods 2015, 222, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, J.W.; Martinez, E.; Louka, P.; Wingett, D.G. Zinc oxide nanoparticles for selective destruction of tumor cells and potential for drug delivery applications. Expert Opin. Drug Deliv. 2010, 7, 1063–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loutfy, S.A.; Al-Ansary, N.A.; Abdel-Ghani, N.T.; Hamed, A.R.; Mohamed, M.B.; Craik, J.D.; Salah Eldin, T.A.; Abdellah, A.M.; Hussein, Y.; Hasanin, M.T.M.; et al. Anti-proliferative activities of metallic nanoparticles in an in vitro breast cancer model. Asian Pac. J. Cancer Prev. 2015, 16, 6039–6046. [Google Scholar] [CrossRef] [Green Version]
- Vinardell, M.P.; Mitjans, M. Antitumor activities of metal oxide nanoparticles. Nanomaterials 2015, 5, 1004–1021. [Google Scholar] [CrossRef] [Green Version]
- Alphandéry, E. Natural metallic nanoparticles for application in nano-oncology. Int. J. Mol. Sci. 2020, 21, 4412. [Google Scholar] [CrossRef]
- Bai Aswathanarayan, J.; Rai Vittal, R.; Muddegowda, U. Anticancer activity of metal nanoparticles and their peptide conjugates against human colon adenorectal carcinoma cells. Artif. Cells Nanomed. Biotechnol. 2018, 46, 1444–1451. [Google Scholar] [CrossRef] [PubMed]
- Shaukat, A.; Anwar, H.; Mahmood, A.; Hussain, G.; Rasul, A.; Umar, M.; Naeem, M.; Ibrahim, M.; Ali, A. Synthesis cum characterization of MgO and MnO nanoparticles and their assessment as antidiabetic and antioxidative agents in diabetic rat model. Phys. B Phys. Condens. Matter 2020, 602, 412570. [Google Scholar] [CrossRef]
- Artimani, T.; Asl, S.; Saidijam, M.; Hasanvand, D.; Afshar, S. Amelioration of diabetes--induced testicular and sperm damage in rats by cerium oxide nanoparticle treatment. Andrologia 2018, 50, e13089. [Google Scholar] [CrossRef]
- El-gharbawy, R.M.; Mahmoud, A.; Abu-risha, S.E. Zinc oxide nanoparticles and a standard antidiabetic drug restore the function and structure of beta cells in Type-2 diabetes. Biomed. Pharmacother. 2016, 84, 810–820. [Google Scholar] [CrossRef]
- Alkaladi, A.; Abdelazim, A.M.; Afifi, M. Antidiabetic activity of zinc oxide and silver nanoparticles on streptozotocin-induced diabetic rats. Int. J. Mol. Sci. 2014, 15, 2015–2023. [Google Scholar] [CrossRef] [Green Version]
- Ali, L.; Shaker, S.; Pinol, R.; Millan, A.; Hanafy, M.; Helmy, M.; Kamel, M.; Mahmoud, S. Effect of superparamagnetic iron oxide nanoparticles on glucose homeostasis on type 2 diabetes experimental model. Life Sci. 2020, 245, 117361. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, S.; Vinodhini, S.; Elanchezhiyan, C.; Rajeswari, D. Evaluation of antidiabetic activity of biologically synthesized silver nanoparticles using Pouteria sapota in streptozotocin-induced diabetic rats. J. Diabetes 2018, 10, 28–42. [Google Scholar] [CrossRef]
- Anderson, D.; Anderson, T.; Fahmi, F. Advances in Applications of Metal Oxide Nanomaterials as Imaging Contrast Agents. Phys. Status Solidi 2019, 216, 1801008. [Google Scholar] [CrossRef]
- Thurn, K.T.; Brown, E.M.B.; Wu, A.; Vogt, S.; Lai, B.; Maser, J.; Paunesku, T.; Woloschak, G.E. Nanoparticles for applications in cellular imaging. Nanoscale Res. Lett. 2007, 2, 430–441. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.C.; Barua, S.; Sharma, G.; Dey, S.K.; Rege, K. Inorganic nanoparticles for cancer imaging and therapy. J. Control. Release 2011, 155, 344–357. [Google Scholar] [CrossRef]
- Wolfbeis, O.S. An overview of nanoparticles commonly used in fluorescent bioimaging. Chem. Soc. Rev. 2015, 44, 4743–4768. [Google Scholar] [CrossRef] [Green Version]
- Naseri, N.; Ajorlou, E.; Asghari, F.; Pilehvar-Soltanahmadi, Y. An update on nanoparticle-based contrast agents in medical imaging. Artif. Cells, Nanomed. Biotechnol. 2018, 46, 1111–1121. [Google Scholar] [CrossRef]
- Arifin, D.; Long, C.; Gilad, A.; Alric, C.; Roux, S.; Tillement, O.; Link, T.; Arepally, A.; Bulte, J. Trimodal Gadolinium-Gold Pancreatic Islet Cells Restore Normoglycemia in Diabetic Mice and Can Be Tracked by Using US, Purpose: Methods: Results. Radiology 2011, 260, 790–798. [Google Scholar] [CrossRef] [Green Version]
- Forte, E.; Fiorenza, D.; Torino, E.; Costagliola di Polidoro, A.; Cavaliere, C.; Netti, P.A.; Salvatore, M.; Aiello, M. Radiolabeled PET/MRI Nanoparticles for Tumor Imaging. J. Clin. Med. 2019, 9, 89. [Google Scholar] [CrossRef] [Green Version]
- Jeon, M.; Halbert, M.V.; Stephen, Z.R.; Zhang, M. Iron Oxide Nanoparticles as T1 Contrast Agents for Magnetic Resonance Imaging: Fundamentals, Challenges, Applications, and Prospectives. Adv. Mater. 2020, 33, 1906539. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Wang, F.; Ding, D.; Song, C.; Guo, C.; Liu, S. TiO 2-x Based Nano-platform for Bimodal Cancer Imaging and NIR-Triggered Chem/Photodynamic/Photothermal Combination Therapy. Chem. Mater. 2017, 29, 9262–9274. [Google Scholar] [CrossRef]
- Zhan, Y.; Shi, S.; Ehlerding, E.B.; Graves, S.A.; Goel, S.; Engle, J.W.; Liang, J.; Tian, J.; Cai, W. Radiolabeled, Antibody-Conjugated Manganese Oxide Nanoparticles for Tumor Vasculature Targeted Positron Emission Tomography and Magnetic Resonance Imaging. ACS Appl. Mater. Interfaces 2017, 9, 38304–38312. [Google Scholar] [CrossRef]
- Xue, S.; Wang, Y.; Wang, M.; Du, X.; Gu, H.; Zhang, C. Iodinated oil-loaded, fluorescent mesoporous silica-coated iron oxide nanoparticles for magnetic resonance imaging/computed tomography/fluorescence trimodal imaging. Int. J. Nanomed. 2014, 9, 2527–2538. [Google Scholar]
- Guo, Z.; Zhang, P.; Luo, Y.; Xie, H.Q.; Chakraborty, S.; Monikh, F.A.; Bu, L.; Liu, Y.; Ma, Y.; Zhang, Z.; et al. Intranasal exposure to ZnO nanoparticles induces alterations in cholinergic neurotransmission in rat brain. Nano Today 2020, 35, 100977. [Google Scholar] [CrossRef]
- Falchi, L.; Galleri, G.; Dore, G.M.; Zedda, M.T.; Pau, S.; Bogliolo, L.; Ariu, F.; Pinna, A.; Nieddu, S.; Innocenzi, P.; et al. Effect of exposure to CeO 2 nanoparticles on ram spermatozoa during storage at 4 ° C for 96 hours. Reprod. Biol. Endocrinol. 2018, 16, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odhiambo, J.; Dejarnette, J.; Geary, T.; Kennedy, C.; Susan, S.; Sutovsky, M.; Sutovsky, P. Increased Conception Rates in Beef Cattle Inseminated with Nanopurified Bull Semen. Biol. Reprod. 2014, 91, 97–101. [Google Scholar] [CrossRef]
- Makhluf, S.; Qasem, R.; Rubinstein, S.; Gedanken, A. Loading Magnetic Nanoparticles into Sperm Cells Does Not Affect Their Functionality. Langmuir 2006, 22, 9480–9482. [Google Scholar] [CrossRef]
- Stern, S.T.; McNeil, S.E. Nanotechnology safety concerns revisited. Toxicol. Sci. 2008, 101, 4–21. [Google Scholar] [CrossRef]
- Chenthamara, D.; Subramaniam, S.; Ramakrishnan, S.G.; Krishnaswamy, S.; Essa, M.M.; Lin, F.H.; Qoronfleh, M.W. Therapeutic efficacy of nanoparticles and routes of administration. Biomater. Res. 2019, 23, 20. [Google Scholar] [CrossRef]
- Song, B.; Zhang, Y.L.; Liu, J.; Feng, X.L.; Zhou, T.; Shao, L.Q. Is Neurotoxicity of Metallic Nanoparticles the Cascades of Oxidative Stress? Nanoscale Res. Lett. 2016, 11, 291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jørgensen, N.; Asklund, C.; Carlsen, E.; Skakkebæk, N.E. Coordinated European investigations of semen quality: Results from studies of Scandinavian young men is a matter of concern. Int. J. Androl. 2006, 29, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Levine, H.; Jørgensen, N.; Martino-, A.; Mendiola, J.; Weksler-derri, D.; Mindlis, I.; Pinotti, R.; Swan, S.H. Temporal trends in sperm count: A systematic review and meta-regression analysis. Hum. Reprod. Update 2017, 23, 646–659. [Google Scholar] [CrossRef]
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandel, M.; Jain, G. Toxic effects of transition metals on male reproductive system: A review. J. Environ. Occup. Sci. 2014, 3, 204. [Google Scholar] [CrossRef] [Green Version]
- Benatta, M.; Kettache, R.; Buchholz, N.; Trinchieri, A. The impact of nutrition and lifestyle on male fertility. Arch. Ital. Urol. Androl. 2020, 92, 121–131. [Google Scholar] [CrossRef]
- Tang, Y.; Chen, B.; Hong, W.; Chen, L.; Yao, L.; Zhao, Y.; Aguilar, Z.P.; Xu, H. ZnO nanoparticles induced male reproductive toxicity based on the effects on the endoplasmic reticulum stress signaling pathway. Int. J. Nanomed. 2019, 14, 9563–9576. [Google Scholar] [CrossRef] [Green Version]
- Afifi, M.; Almaghrabi, O.A.; Kadasa, N.M. Ameliorative Effect of Zinc Oxide Nanoparticles on Antioxidants and Sperm Characteristics in Streptozotocin-Induced Diabetic Rat Testes. BioMed Res. Int. 2015, 2015, 153573. [Google Scholar] [CrossRef]
- Pinho, A.R.; Martins, F.; Costa, M.E.; Senos, A.M.; da Cruz e Silva, O.A.; Pereira, M.D.; Rebelo, S. In Vitro Cytotoxicity Effects of Zinc Oxide Nanoparticles on Spermatogonia Cells. Cells 2020, 9, 1081. [Google Scholar] [CrossRef]
- Mäkelä, J.-A.; Toppari, J. Spermatogenesis. In Endocrinology; Simoni, M., Huhtaniemi, I.T., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 417–455. ISBN 9783319444413. [Google Scholar]
- de Kretser, D.M.; Loveland, K.L.; Meinhardt, A.; Simorangkir, D.; Wreford, N. Spermatogenesis. Hum. Reprod. 1998, 13, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.Y.; Mruk, D.D. Biochemistry of Sertoli cell/germ cell junctions, germ cell transport, and spermiation in the seminiferous epithelium. In Sertoli Cell Biology; Elsevier Inc.: Amsterdam, The Netherlands, 2015; ISBN 9780124170476. [Google Scholar]
- Gao, G.; Ze, Y.; Zhao, X.; Sang, X.; Zheng, L.; Ze, X.; Gui, S.; Sheng, L.; Sun, Q.; Hong, J.; et al. Titanium dioxide nanoparticle-induced testicular damage, spermatogenesis suppression, and gene expression alterations in male mice. J. Hazard. Mater. 2013, 258–259, 133–143. [Google Scholar] [CrossRef]
- Sundarraj, K.; Manickam, V.; Raghunath, A.; Periyasamy, M.; Viswanathan, M.; Perumal, E. Repeated Exposure to Iron Oxide Nanoparticles Causes Testicular Toxicity in Mice. Environ. Toxicol. 2016, 32, 594–608. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Suzuki, K.I.; Ishihara, A.; Kubo-Irie, M.; Fujimoto, R.; Tabata, M.; Oshio, S.; Nihei, Y.; Ihara, T.; Sugamata, M. Nanoparticles transferred from pregnant mice to their offspring can damage the genital and cranial nerve systems. J. Heal. Sci. 2009, 55, 95–102. [Google Scholar] [CrossRef] [Green Version]
- McAuliffe, M.E.; Perry, M.J. Are nanoparticles potential male reproductive toxicants? A literature review. Nanotoxicology 2007, 1, 204–210. [Google Scholar] [CrossRef]
- Gallo, A.; Boni, R.; Buttino, I.; Tosti, E. Spermiotoxicity of nickel nanoparticles in the marine invertebrate Ciona intestinalis (ascidians). Nanotoxicology 2016, 10, 1096–1104. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, X.; Xiao, S.; Liu, X.; Chen, X.; Xia, Q.; Lei, S.; Li, H.; Zhong, Z.; Xiao, K. The Effects of Gold Nanoparticles on Leydig Cells and Male Reproductive Function in Mice. Int. J. Nanomed. 2020, 15, 9499–9514. [Google Scholar] [CrossRef]
- Sharma, R.; Agarwal, A. Sperm Chromatin: Biological and Clinical Applications in Male Infertility and Assisted Reproduction, 1st ed.; Zini, A., Agarwal, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; ISBN 9781441968579. [Google Scholar]
- Manku, G.; Culty, M. Mammalian gonocyte and spermatogonia differentiation: Recent advances and remaining challenges. Reproduction 2015, 149, R139–R157. [Google Scholar] [CrossRef] [Green Version]
- Hess, R.A.; De Franca, L.R. Spermatogenesis and cycle of the seminiferous epithelium. Adv. Exp. Med. Biol. 2008, 636, 1–15. [Google Scholar] [CrossRef]
- Pereira, C.; Serrano, J.; Martins, F.; Silva, O.; Rebelo, S. Nuclear envelope dynamics during mammalian spermatogenesis: New insights on male fertility. Biol. Rev. 2019, 94, 1195–1219. [Google Scholar] [CrossRef]
- Donnell, L.; Nicholls, P.; Bryan, M.; Mclachlan, R.; Stanton, P. Spermiation: The process of sperm release. Spermatogenesis 2011, 1, 14–35. [Google Scholar] [CrossRef]
- Tapia, J.; Peña, F. Apoptotic Events in Male Germ Cells and in Mature Mammalian Spermatozoa; Salido, G.M., Rosado, J.A., Eds.; Springer: Dordrecht, The Netherlands, 2009; ISBN 9781402098734. [Google Scholar]
- Préaubert, L.; Tassistro, V.; Au, M.; Sari-minodier, I.; Rose, J.; Courbiere, B.; Perrin, J. Very low concentration of cerium dioxide nanoparticles induce DNA damage, but no loss of vitality, in human spermatozoa. Toxicol. Vitr. 2018, 50, 236–241. [Google Scholar] [CrossRef]
- Basioura, A.; Michos, I.; Ntemka, A.; Karagiannis, I.; Boscos, M. Effect of iron oxide and silver nanoparticles on boar semen CASA motility and kinetics. J. Hell. Vet. Med. Soc. 2020, 71, 2331–2338. [Google Scholar] [CrossRef]
- Zhang, X.; Yue, Z.; Zhang, H.; Liu, L.; Zhou, X. Repeated administrations of Mn3O4 nanoparticles cause testis damage and fertility decrease through PPAR-signaling pathway. Nanotoxicology 2020, 14, 326–340. [Google Scholar] [CrossRef]
- Pawar, K.; Kaul, G. Toxicity of titanium oxide nanoparticles causes functionality and DNA damage in buffalo (Bubalus bubalis) sperm in vitro. Toxicol. Ind. Health 2014, 30, 520–533. [Google Scholar] [CrossRef]
- Mao, Z.; Yao, M.; Xu, B.; Ji, X.; Jiang, H.; Han, X.; Tang, Q.; Zhou, Z.; Chen, R.; Li, X.; et al. Cytoskeletons of two reproductive germ cell lines response differently to titanium dioxide nanoparticles mediating vary reproductive toxicity. J. Biomed. Nanotechnol. 2017, 13, 409–416. [Google Scholar] [CrossRef]
- Santonastaso, M.; Mottola, F.; Colacurci, N.; Iovine, C.; Pacifico, S.; Cammarota, M.; Cesaroni, F.; Rocco, L. In vitro genotoxic effects of titanium dioxide nanoparticles (n-TiO2) in human sperm cells. Mol. Reprod. Dev. 2019, 86, 1369–1377. [Google Scholar] [CrossRef]
- Barkhoradi, A.; Hekmatimoghaddam, S.; Jebali, A.; Khalili, M.; Talebi, A.; Noorani, M. Effect of zinc oxide nanoparticles on viability of human spermatozoa. Iran. J. Reprod. Med. 2013, 11, 767–771. [Google Scholar]
- Han, Z.; Yan, Q.; Ge, W.; Liu, Z.-G.; Gurunathan, S.; Felici, M.; Shen, W.; Zang, X.-F. Cytotoxic effects of ZnO nanoparticles on mouse testicular cells. Int. J. Nanomed. 2016, 11, 5187–5203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Xu, C.; Ji, G.; Liu, H.; Mo, Y.; Tollerud, D.J.; Gu, A.; Zhang, Q. Sublethal effects of zinc oxide nanoparticles on male reproductive cells. Toxicol. Vitr. 2016, 35, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Bara, N.; Kaul, G. Enhanced steroidogenic and altered antioxidant response by ZnO nanoparticles in mouse testis Leydig cells. Toxicol. Ind. Health 2018, 34, 571–588. [Google Scholar] [CrossRef]
- Shen, J.; Yang, D.; Zhou, X.; Wang, Y.; Tang, S.; Yin, H. Role of Autophagy in Zinc Oxide Nanoparticles-Induced Apoptosis of Mouse LEYDIG Cells. Int. J. Mol. 2019, 20, 4042. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Neuss, S.; Leifert, A.; Fischler, M.; Wen, F.; Simon, U.; Schmid, G.; Brandau, W.; Jahnen-dechent, W. Size-Dependent Cytotoxicity of Gold Nanoparticles. Small 2007, 3, 1941–1949. [Google Scholar] [CrossRef]
- Gromadzka-ostrowska, J.; Dziendzikowska, K.; Lankoff, A.; Radzikowska, J.; Wojewódzka, M.; Kruszewski, M. Silver nanoparticles effects on epididymal sperm in rats. Toxicol. Lett. 2012, 214, 251–258. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Castellini, C.; Ruggeri, S.; Mattioli, S.; Bernardini, G.; Macchioni, L.; Moretti, E.; Collodel, G. Long-term effects of silver nanoparticles on reproductive activity of rabbit buck. Syst. Biol. Reprod. Med. 2014, 60, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Yousef, M.; Al-hamadani, M.; Kamel, M. Reproductive Toxicity of Aluminum Oxide Nanoparticles and Zinc Oxide Nanoparticles in Male Rats. Nanoparticle 2019, 1, 3. [Google Scholar] [CrossRef]
- Qin, F.; Shen, T.; Li, J.; Qian, J.; Zhang, J.; Zhou, G.; Tong, J. SF-1 mediates reproductive toxicity induced by Cerium oxide nanoparticles in male mice. J. Nanobiotechnology 2019, 17, 41. [Google Scholar] [CrossRef]
- Nasri, S.; Rezai-zarchi, S.; Kerishchi, P.; Sadeghi, S. The Effect of Iron Oxide Nanoparticles on Sperm Numbers and Mobility in Male Mice. Zahedan J. Res. Med. Sci. 2015, 17, 10–12. [Google Scholar] [CrossRef]
- Varzeghani, S.M.; Parivar, K.; Abdollahifar, M.-A.; Karamian, A. Effects of Iron Oxide Nanoparticles on Mouse Sperm Parameters and Testicular Tissue. Iran. J. Toxicol. 2018, 12, 39–44. [Google Scholar] [CrossRef]
- Younus, A.I.; Yousef, M.I.; Abdel-NabiKamel, M.; Alrawi, R.; Abdulrahman, J.M. Changes in semen characteristics and sex hormones of rats treated with iron oxide nanoparticles, silver nanoparticles and their mixture. GSC Biol. Pharm. Sci. 2020, 12, 229–237. [Google Scholar] [CrossRef]
- Negahdary, M.; Arefian, Z.; Dastjerdi, H.A.; Ajdary, M. Toxic effects of Mn2O3 nanoparticles on rat testis and sex hormone. J. Nat. Sci. Biol. Med. 2015, 6, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Yousefalizadegan, N.; Mousavi, Z.; Rastegar, T.; Razavi, Y.; Najafizadeh, P. Reproductive toxicity of manganese dioxide in forms of micro-and nanoparticles in male rats. Int. J. Reprod. Biomed. 2019, 17, 361–370. [Google Scholar] [CrossRef]
- Hong, F.; Si, W.; Zhao, X.; Wang, L.; Zhou, Y.; Chen, M.; Ge, Y.; Zhang, Q.; Wang, Y.; Zhang, J. TiO2 Nanoparticle Exposure Decreases Spermatogenesis via Biochemical Dysfunctions in the Testis of Male Mice. J. Agric. Food Chem. 2015, 63, 7084–7092. [Google Scholar] [CrossRef]
- Hong, F.; Zhao, X.; Si, W.; Ze, Y.; Wang, L. Decreased spermatogenesis led to alterations of testis-specific gene expression in male mice following nano-TiO 2 exposure. J. Hazard. Mater. 2015, 300, 718–728. [Google Scholar] [CrossRef] [PubMed]
- Meena, R.; Kajal, K.; Paulraj, R. Cytotoxic and Genotoxic Effects of Titanium Dioxide Nanoparticles in Testicular Cells of Male Wistar Rat. Appl. Biochem. Biotechnol. 2015, 175, 825–840. [Google Scholar] [CrossRef] [PubMed]
- Morgan, A.; El-hamid, M.; Noshy, P. Reproductive Toxicity Investigation of Titanium Dioxide Nanoparticles in Male Albino Rats. World J. Pharm. Pharm. Sci. 2015, 4, 34–49. [Google Scholar]
- Miura, N.; Ohtani, K.; Hasegawa, T.; Yoshioka, H.; Hwang, G.W. High sensitivity of testicular function to titanium nanoparticles. J. Toxicol. Sci. 2017, 42, 359–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, A.M.; Ibrahim, M.A.; Noshy, P.A. Reproductive toxicity provoked by titanium dioxide nanoparticles and the ameliorative role of Tiron in adult male rats. Biochem. Biophys. Res. Commun. 2017, 486, 595–600. [Google Scholar] [CrossRef]
- Song, G.; Lin, L.; Liu, L.; Wang, K.; Ding, Y.; Niu, Q.; Mu, L.; Wang, H.; Shen, H.; Guo, S. Toxic effects of anatase titanium dioxide nanoparticles on spermatogenesis and testicles in male mice. Polish J. Environ. Stud. 2017, 26, 2739–2746. [Google Scholar] [CrossRef]
- Lauvås, A.J.; Skovmand, A.; Poulsen, M.S.; Kyjovska, Z.O.; Roursgaard, M.; Goericke-Pesch, S.; Vogel, U.; Hougaard, K.S. Airway exposure to TiO2 nanoparticles and quartz and effects on sperm counts and testosterone levels in male mice. Reprod. Toxicol. 2019, 90, 134–140. [Google Scholar] [CrossRef]
- Miura, N.; Ohtani, K.; Hasegawa, T.; Yoshioka, H.; Hwang, G.W. Biphasic adverse effect of titanium nanoparticles on testicular function in mice. Sci. Rep. 2019, 9, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Jafari, A.; Karimipour, M.; Khaksar, M.R.; Ghasemnejad-Berenji, M. Protective effects of orally administered thymol against titanium dioxide nanoparticle–induced testicular damage. Environ. Sci. Pollut. Res. 2020, 27, 2353–2360. [Google Scholar] [CrossRef]
- Ogunsuyi, O.M.; Ogunsuyi, O.I.; Akanni, O.; Alabi, O.A.; Alimba, C.G.; Adaramoye, O.A.; Cambier, S.; Eswara, S.; Gutleb, A.C.; Bakare, A.A. Alteration of sperm parameters and reproductive hormones in Swiss mice via oxidative stress after co-exposure to titanium dioxide and zinc oxide nanoparticles. Andrologia 2020, 52, e13758. [Google Scholar] [CrossRef]
- Talebi, A.R.; Khorsandi, L.; Moridian, M. The effect of zinc oxide nanoparticles on mouse spermatogenesis. J. Assist. Reprod. Genet. 2013, 30, 1203–1209. [Google Scholar] [CrossRef] [Green Version]
- Abbasalipourkabir, R.; Moradi, H.; Zarei, S.; Asadi, S.; Salehzadeh, A.; Ghafourikhosroshahi, A.; Mortazavi, M.; Ziamajidi, N. Toxicity of zinc oxide nanoparticles on adult male Wistar rats. Food Chem. Toxicol. 2015, 84, 154–160. [Google Scholar] [CrossRef]
- Mozaffari, Z.; Parivar, K.; Roodbari, N.H.; Irani, S. Histopathological Evaluation of the Toxic Effects of Zinc Oxide (ZnO) Nanoparticles on Testicular Tissue of NMRI Adult Mice. Adv. Stud. Biol. 2015, 7, 275–291. [Google Scholar] [CrossRef] [Green Version]
- Hussein, M.; Ali, H.; Saadeldin, I.; Ahmed, M. Querectin Alleviates Zinc Oxide Nanoreprotoxicity in Male Albino Rats. J. Biochem. Mol. Toxicol. 2016, 30, 489–496. [Google Scholar] [CrossRef]
- Srivastav, A.K.; Kumar, A.; Prakash, J.; Singh, D.; Jagdale, P.; Shankar, J.; Kumar, M. Genotoxicity evaluation of zinc oxide nanoparticles in Swiss mice after oral administration using chromosomal aberration, micronuclei, semen analysis, and RAPD profile. Toxicol. Ind. Health 2017, 33, 821–834. [Google Scholar] [CrossRef]
- Mesallam, D.; Deraz, R.; Abdel Aal, S.; Ahmed, S. Toxicity of Subacute Oral Zinc Oxide Nanoparticles on Testes and Prostate of Adult Albino Rats and Role of Recovery. J. Histol. Histopathol. 2019, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Radhi, M.J.; Adnan, G.; Latef, A. Effect of Zinc oxide nanoparticles (ZnO-NPs) on weights of some reproductive organs and sperm abnormalities in the tail of epididymis of albino mice. J. Pharm. Sci. Res. 2019, 11, 243–246. [Google Scholar]
- Salman, R.A. The Influence of ZnO NPs on Reproductive System Tissues of Albino Male Mice. Histopathological Study. Int. J. Sci. Res. 2021, 6, 2021–2025. [Google Scholar] [CrossRef]
- Hess, R.; Chen, P. Computer tracking of germ cells in the cycle of the seminiferous epithelium and prediction of changes in cycle duration in animals commonly used in reproductive biology and toxicology. J. Androl. 1992, 13, 185–190. [Google Scholar]
- Millaku, L.; Imeri, R.; Trebicka, A. Histopathological changes in testes of house sparrow (Passer domesticus). J. Mater. Environ. Sci. 2015, 6, 1292–1296. [Google Scholar]
- Saleh, R.; Agarwal, A. Oxidative Stress and Male Infertility: From Research Bench to Clinical Practice. J. Androl. 2002, 23, 737–752. [Google Scholar]
- Tremellen, K. Oxidative stress and male infertility—A clinical perspective. Hum. Reprod. Update 2008, 14, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Feng, W.; Chen, Y.; Shi, J. Inorganic nanoparticles in clinical trials and translations. Nano Today 2020, 35, 100972. [Google Scholar] [CrossRef]
- Shandilya, R.; Mishra, P.K.; Pathak, N.; Lohiya, N.K.; Sharma, R.S. Nanotechnology in reproductive medicine: Opportunities for clinical translation. Clin. Exp. Reprod. Med. 2020, 47, 245–262. [Google Scholar] [CrossRef] [PubMed]
- Isaac, A.V.; Kumari, S.; Nair, R.; Urs, D.R.; Salian, S.R.; Kalthur, G.; Adiga, S.K.; Manikkath, J.; Mutalik, S.; Sachdev, D.; et al. Supplementing zinc oxide nanoparticles to cryopreservation medium minimizes the freeze-thaw-induced damage to spermatozoa. Biochem. Biophys. Res. Commun. 2017, 494, 656–662. [Google Scholar] [CrossRef]
- Durfey, C.L.; Swistek, S.E.; Liao, S.F.; Crenshaw, M.A.; Clemente, H.J.; Thirumalai, R.V.K.G.; Steadman, C.S.; Ryan, P.L.; Willard, S.T.; Feugang, J.M. Nanotechnology-based approach for safer enrichment of semen with best spermatozoa. J. Anim. Sci. Biotechnol. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Chang, M.; Chang, Y.-J.; Wang, T.-Y.; Yu, Q. Sperm Movement Control Utilizing Surface Charged Magnetic Nanoparticles. J. Nanosci. Nanotechnol. 2019, 19, 5713–5722. [Google Scholar] [CrossRef] [PubMed]
- Moridi, H.; Hosseini, S.A.; Shateri, H.; Kheiripour, N.; Kaki, A.; Hatami, M.; Ranjbar, A. Protective effect of cerium oxide nanoparticle on sperm quality and oxidative damage in malathioninduced testicular toxicity in rats: An experimental study. Int. J. Reprod. Biomed. 2018, 16, 261–266. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, H.J.; Doo, S.H.; Yang, W.J.; Choi, D.; Kim, J.H.; Won, J.H.; Song, Y.S. Use of nanoparticles to monitor human mesenchymal stem cells transplanted into penile cavernosum of rats with erectile dysfunction. Korean J. Urol. 2015, 56, 280–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobyliak, N.M.; Falalyeyeva, T.M.; Kuryk, O.G.; Beregova, T.V.; Bodnar, P.M.; Zholobak, N.M.; Shcherbakov, O.B.; Bubnov, R.V.; Spivak, Y.M. Antioxidative effects of cerium dioxide nanoparticles ameliorate age-related male infertility: Optimistic results in rats and the review of clinical clues for integrative concept of men health and fertility. EPMA J. 2015, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitounis, D.; Klein, J.P.; Mery, L.; El-Merhie, A.; Forest, V.; Boudard, D.; Pourchez, J.; Cottier, M. Ex vivo detection and quantification of gold nanoparticles in human seminal and follicular fluids. Analyst 2018, 143, 475–486. [Google Scholar] [CrossRef]
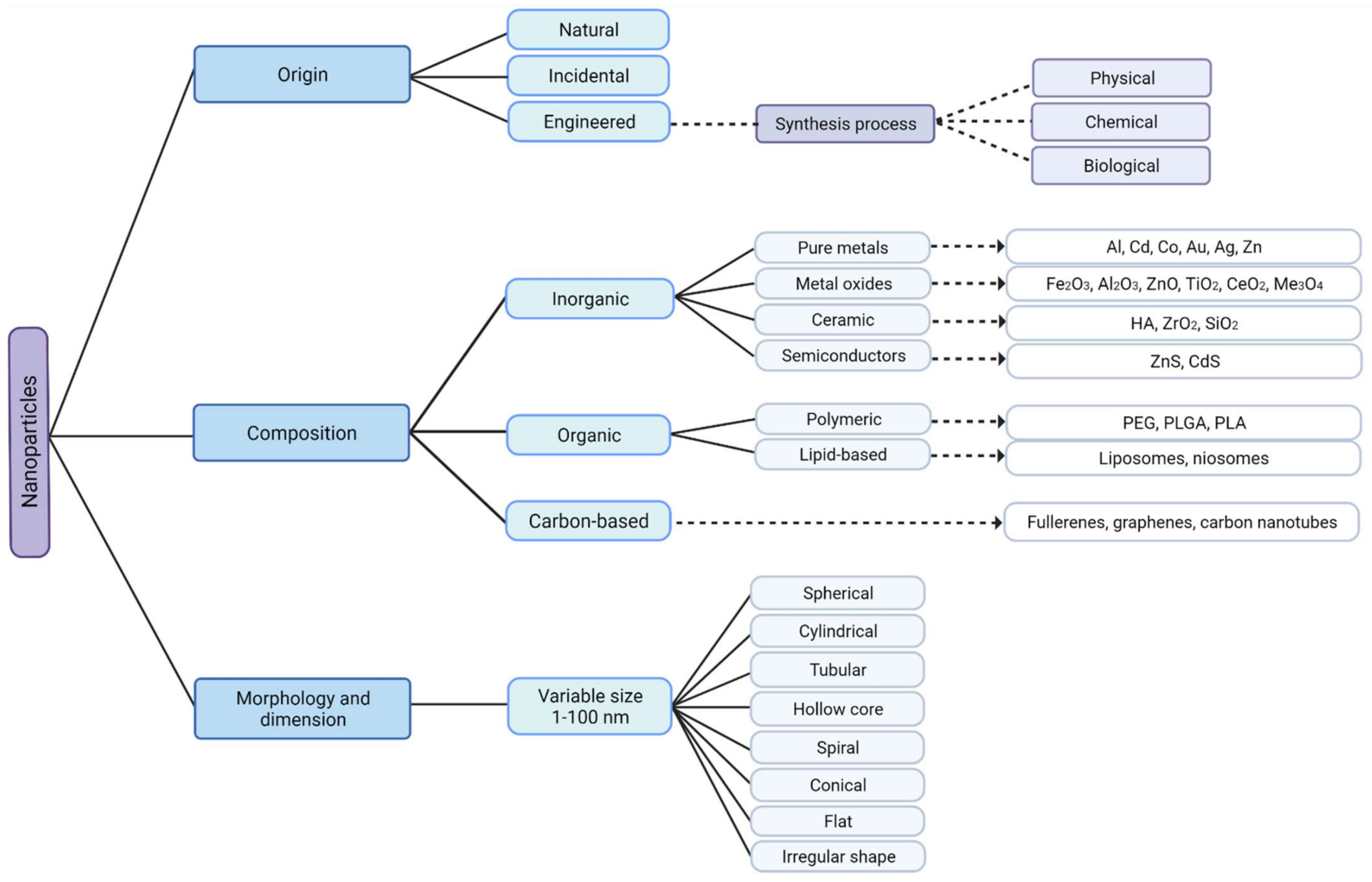
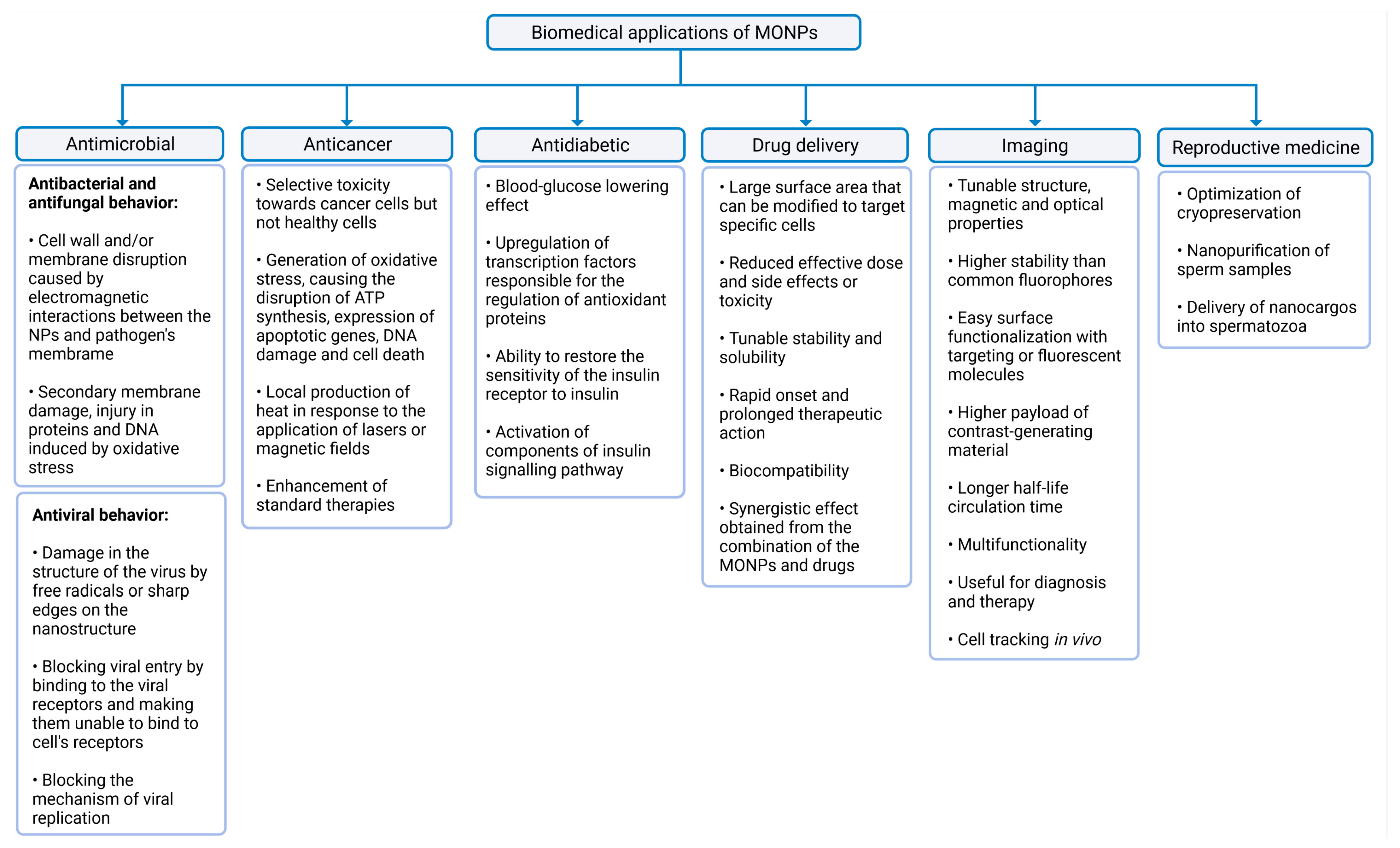
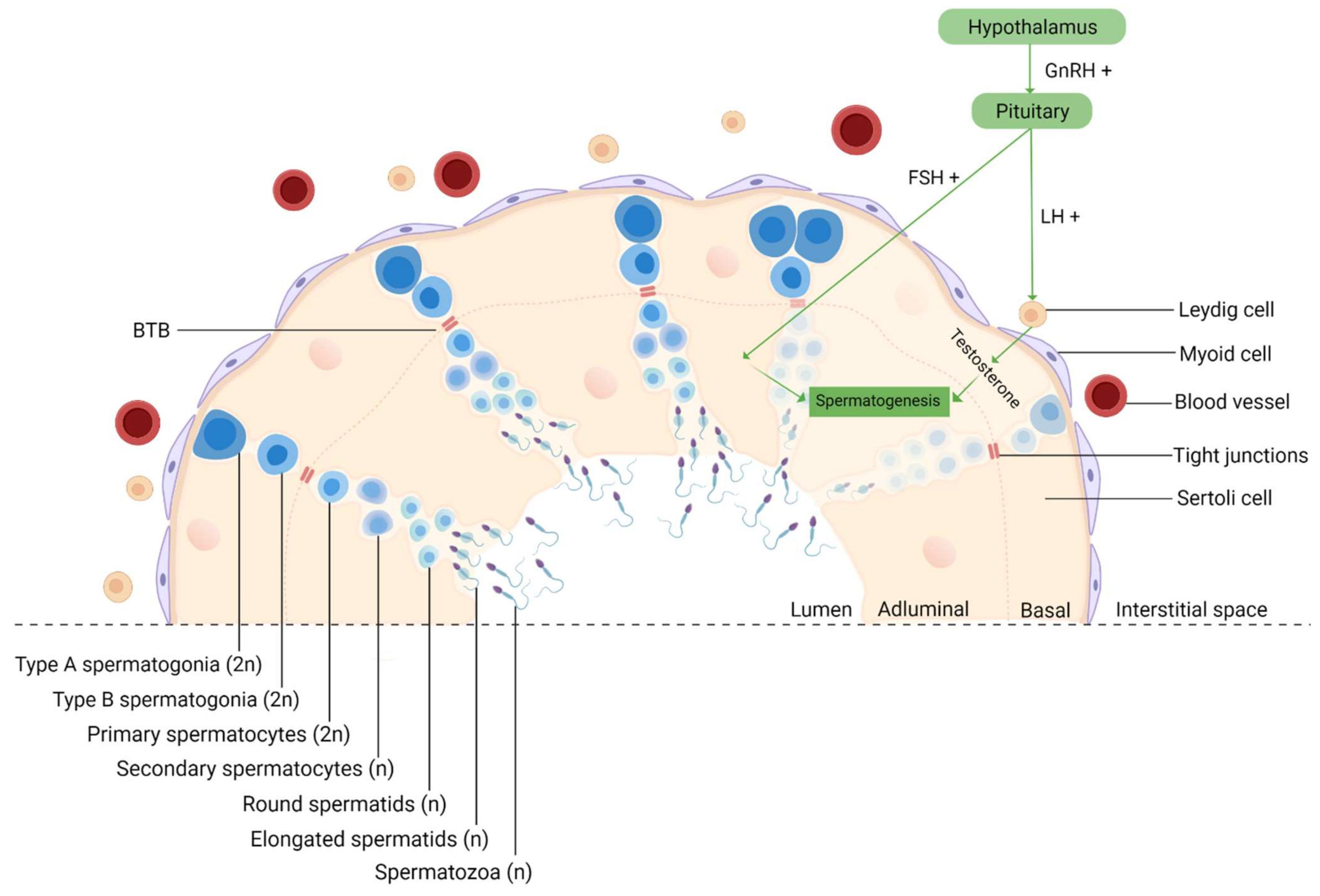
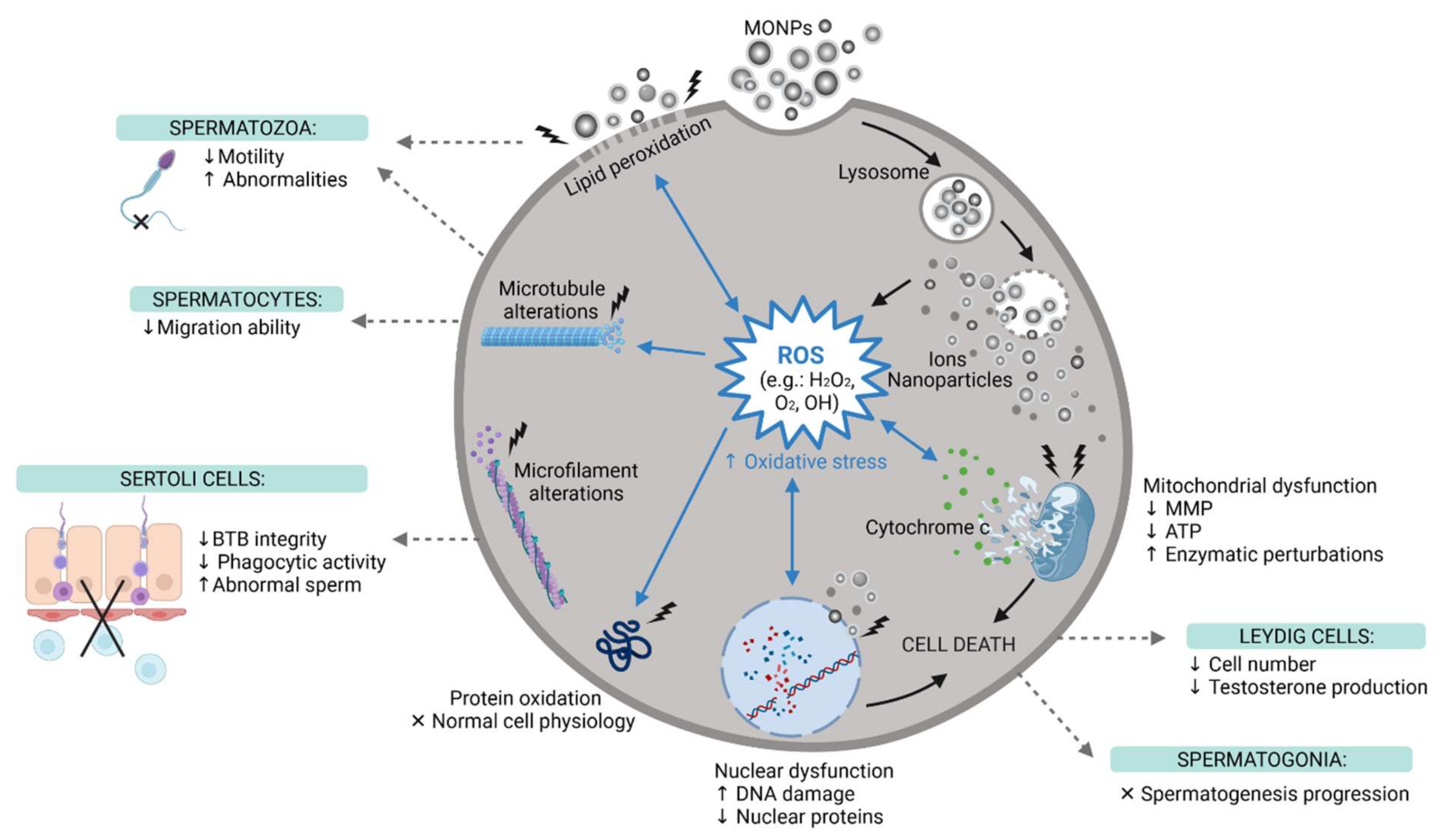
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).