
Unique Aspects of Human Placentation


Abstract
1. Introduction
Mammalian Evolution and Phylogeny
2. Early Development
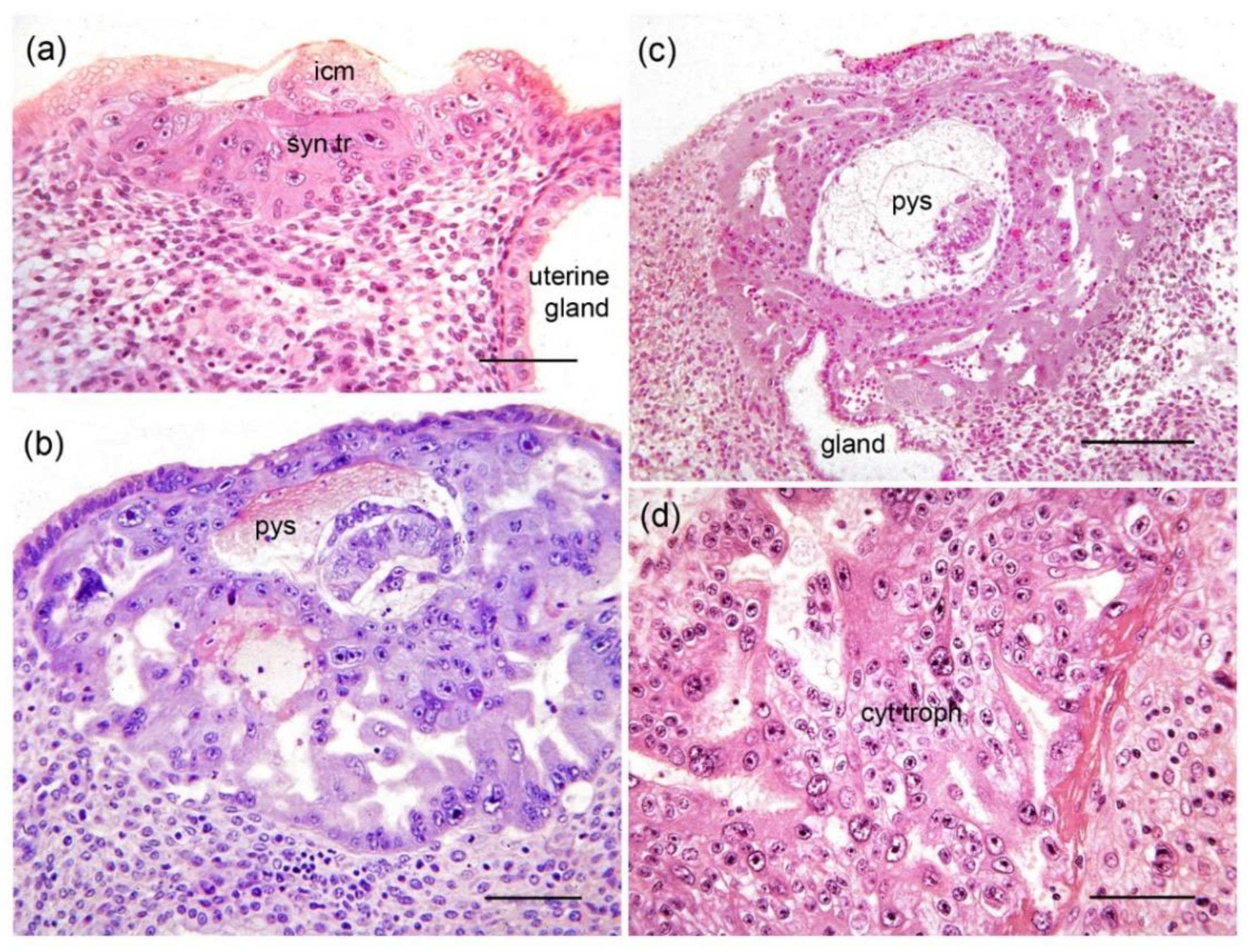
2.1. Interstitial Implantation
2.2. Initial Decidual Reaction
2.3. Early Differentiation of Mesoderm and Secondary Yolk Sac
2.4. Allantoic Stalk
3. Placentation
3.1. Haemochorial Placentation
3.2. Villous rather Than Labyrinthine Placentation
3.3. Uterine Spiral Artery Transformation
3.4. Trophoblast Invasion by Interstitial and Intravascular Routes
4. Immunology of Decidua and Trophoblast
Interplay of uNK Cell Receptors and HLA Antigens
5. Endocrinology of the Placenta
5.1. Chorionic Gonadotrophins
5.2. Placental Lactogens and Growth Hormones
5.3. Progesterone and Its Receptors
6. Pregnancy Duration and Newborn State
7. Discussion
7.1. Placental Evolution
7.2. Pregnancy Complications
8. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- Brosens, I.; Pijnenborg, R.; Vercruysse, L.; Romero, R. The “Great Obstetrical Syndromes” are associated with disorders of deep placentation. Am. J. Obstet. Gynecol. 2011, 204, 193–201. [Google Scholar] [CrossRef]
- Carter, A.M. Animal models of human pregnancy and placentation: Alternatives to the mouse. Reproduction 2020, 160, R129–R143. [Google Scholar] [CrossRef]
- Carter, A.M. Animal models of human placentation—A review. Placenta 2007, 28, S41–S47. [Google Scholar] [CrossRef]
- Carter, A.M. Evolution of Placental Function in Mammals: The Molecular Basis of Gas and Nutrient Transfer, Hormone Secretion, and Immune Responses. Physiol. Rev. 2012, 92, 1543–1576. [Google Scholar] [CrossRef]
- Huhn, O.; Zhao, X.; Esposito, L.; Moffett, A.; Colucci, F.; Sharkey, A.M. How Do Uterine Natural Killer and Innate Lymphoid Cells Contribute to Successful Pregnancy? Front. Immunol. 2021, 12, 607669. [Google Scholar] [CrossRef] [PubMed]
- Prabhudas, M.; Bonney, E.; Caron, K.; Dey, S.; Erlebacher, A.; Fazleabas, A.; Fisher, S.; Golos, T.; Matzuk, M.; McCune, J.M.; et al. Immune mechanisms at the maternal-fetal interface: Perspectives and challenges. Nat. Immunol. 2015, 16, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Meredith, R.W.; Janečka, J.E.; Gatesy, J.; Ryder, O.A.; Fisher, C.A.; Teeling, E.C.; Goodbla, A.; Eizirik, E.; Simão, T.L.L.; Stadler, T.; et al. Impacts of the Cretaceous Terrestrial Revolution and KPg Extinction on Mammal Diversification. Science 2011, 334, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Murphy, W.J.; Pringle, T.H.; Crider, T.A.; Springer, M.S.; Miller, W. Using genomic data to unravel the root of the placental mammal phylogeny. Genome Res. 2007, 17, 413–421. [Google Scholar] [CrossRef]
- Janečka, J.E.; Miller, W.; Pringle, T.H.; Wiens, F.; Zitzmann, A.; Helgen, K.M.; Springer, M.S.; Murphy, W.J. Molecular and Genomic Data Identify the Closest Living Relative of Primates. Science 2007, 318, 792–794. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.M.; Enders, A.C. The Evolution of Epitheliochorial Placentation. Annu. Rev. Anim. Biosci. 2013, 1, 443–467. [Google Scholar] [CrossRef]
- Hubrecht, A.A.W. Relations of tarsius to the lemurs and apes. Science 1897, 5, 550–551. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Perelman, P.; Johnson, W.; Roos, C.; Seuánez, H.N.; Horvath, J.E.; Moreira, M.A.M.; Kessing, B.; Pontius, J.; Roelke, M.; Rumpler, Y.; et al. A Molecular Phylogeny of Living Primates. PLoS Genet. 2011, 7, e1001342. [Google Scholar] [CrossRef] [PubMed]
- James, J.; Carter, A.; Chamley, L. Human placentation from nidation to 5 weeks of gestation. Part I: What do we know about formative placental development following implantation? Placenta 2012, 33, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.P., II. Croonian lecture—The developmental history of the primates. Philos. Trans. R. Soc. Lond. Ser. B Contain. Pap. A Biol. Character 1932, 221, 45–178. [Google Scholar] [CrossRef]
- Selenka, E. Entwickelung des Gibbon (Hylobates und Siamanga). Stud. Über Enwickelungsgeschichte Tiere 1899, 8, 163–208. [Google Scholar]
- Carter, A.M.; Enders, A.C.; Pijnenborg, R. The role of invasive trophoblast in implantation and placentation of primates. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140070. [Google Scholar] [CrossRef]
- Dupressoir, A.; Lavialle, C.; Heidmann, T. From ancestral infectious retroviruses to bona fide cellular genes: Role of the captured syncytins in placentation. Placenta 2012, 33, 663–671. [Google Scholar] [CrossRef]
- Lavialle, C.; Cornelis, G.; Dupressoir, A.; Esnault, C.; Heidmann, O.; Vernochet, C.; Heidmann, T. Paleovirology of ‘syncytins’, retroviral env genes exapted for a role in placentation. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120507. [Google Scholar] [CrossRef]
- Chavan, A.R.; Bhullar, B.-A.S.; Wagner, G.P. What was the ancestral function of decidual stromal cells? A model for the evolution of eutherian pregnancy. Placenta 2016, 40, 40–51. [Google Scholar] [CrossRef]
- Mossman, H.W. Vertebrate Fetal Membranes: Comparative Ontogeny and Morphology; Evolution; Phylogenetic Significance; Basic Functions; Research Opportunities; Rutgers University Press: New Brunswick, NJ, USA, 1987. [Google Scholar]
- Kin, K.; Maziarz, J.; Chavan, A.R.; Kamat, M.; Vasudevan, S.; Birt, A.; Wagner, G.P. The transcriptomic evolution of mammalian pregnancy: Gene expression innovations in endometrial stromal fibroblasts. Genome Biol. Evol. 2016, 8, 2459–2473. [Google Scholar] [CrossRef]
- Griffith, O.W.; Chavan, A.R.; Pavlicev, M.; Protopapas, S.; Callahan, R.; Maziarz, J.; Wagner, G.P. Endometrial recognition of pregnancy occurs in the grey short-tailed opossum (Monodelphis domestica). Proc. Biol. Sci. 2019, 286, 20190691. [Google Scholar] [CrossRef]
- Griffith, O.W.; Chavan, A.R.; Protopapas, S.; Maziarz, J.; Romero, R.; Wagner, G.P. Embryo implantation evolved from an ancestral inflammatory attachment reaction. Proc. Natl. Acad. Sci. USA 2017, 114, E6566–E6575. [Google Scholar] [CrossRef]
- Erkenbrack, E.M.; Maziarz, J.D.; Griffith, O.; Liang, C.; Chavan, A.R.; Nnamani, M.C.; Wagner, G.P. The mammalian decidual cell evolved from a cellular stress response. PLoS Biol. 2018, 16, e2005594. [Google Scholar] [CrossRef]
- Enders, A.C.; King, B.F. Development of the human yolk sac. In The Human Yolk Sac and Yolk Sac Tumors; Nogales, F.F., Ed.; Springer: Berlin, Germany, 1993; pp. 33–47. [Google Scholar]
- Burton, G.J.; Cindrova-Davies, T.; Turco, M.Y. Review: Histotrophic nutrition and the placental-endometrial dialogue during human early pregnancy. Placenta 2020, 102, 21–26. [Google Scholar] [CrossRef]
- Boroviak, T.; Stirparo, G.; Dietmann, S.; Hernando-Herraez, I.; Mohammed, H.; Reik, W.; Smith, A.; Sasaki, E.; Nichols, J.; Bertone, P. Single cell transcriptome analysis of human, marmoset and mouse embryos reveals common and divergent features of preimplantation development. Development 2018, 145, 27. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.; Boroviak, T.E. Origin and function of the yolk sac in primate embryogenesis. Nat. Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Carter, A. IFPA Senior Award Lecture: Mammalian fetal membranes. Placenta 2016, 48, S21–S30. [Google Scholar] [CrossRef] [PubMed]
- Mess, A.; Carter, A.M. Evolutionary transformations of fetal membrane characters in Eutheria with special reference to Afrotheria. J. Exp. Zool. Part B Mol. Dev. Evol. 2006, 306B, 140–163. [Google Scholar] [CrossRef] [PubMed]
- Bongso, T.A.; Basrur, P.K. Foetal fluids in cattle. Can. Vet. J. 1976, 17, 38–41. [Google Scholar] [PubMed]
- Wildman, D.E.; Chen, C.; Erez, O.; Grossman, L.I.; Goodman, M.; Romero, R. Evolution of the mammalian placenta revealed by phylogenetic analysis. Proc. Natl. Acad. Sci. USA 2006, 103, 3203–3208. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.; Jauniaux, E. Ultrastructure of the materno-embryonic interface in the first trimester of pregnancy. Micron 1995, 26, 145–173. [Google Scholar] [CrossRef]
- Enders, A.C. A comparative study of the fine structure of the trophoblast in several hemochorial placentas. Am. J. Anat. 1965, 116, 29–67. [Google Scholar] [CrossRef]
- Carter, A.M. Placental Gas Exchange and the Oxygen Supply to the Fetus. Compr. Physiol. 2015, 5, 1381–1403. [Google Scholar] [CrossRef] [PubMed]
- Mossman, H.W. The rabbit placenta and the problem of placental transmission. Am. J. Anat. 1926, 37, 433–497. [Google Scholar] [CrossRef]
- Metcalfe, J.; Bartels, H.; Moll, W. Gas exchange in the pregnant uterus. Physiol. Rev. 1967, 47, 782–838. [Google Scholar] [CrossRef]
- Adamson, S.L.; Lu, Y.; Whiteley, K.J.; Holmyard, D.; Hemberger, M.; Pfarrer, C.; Cross, J.C. Interactions between trophoblast cells and the maternal and fetal circulation in the mouse placenta. Dev. Biol. 2002, 250, 358–373. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, E.M.; Harris, J.W.S. Comparison of utero-placental vasculature and circulation in the rhesus monkey and man. Contrib. Embryol. Carnegie Inst. 1966, 38, 61–70. [Google Scholar]
- Harris, L. IFPA Gabor Than Award lecture: Transformation of the spiral arteries in human pregnancy: Key events in the remodelling timeline. Placenta 2011, 32, S154–S158. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pijnenborg, R.; Vercruysse, L.; Hanssens, M. The Uterine Spiral Arteries in Human Pregnancy: Facts and Controversies. Placenta 2006, 27, 939–958. [Google Scholar] [CrossRef] [PubMed]
- Lash, G.E.; Bulmer, J.N. Do uterine natural killer (uNK) cells contribute to female reproductive disorders? J. Reprod. Immunol. 2011, 88, 156–164. [Google Scholar] [CrossRef]
- Burton, G.; Woods, A.; Jauniaux, E.; Kingdom, J. Rheological and Physiological Consequences of Conversion of the Maternal Spiral Arteries for Uteroplacental Blood Flow during Human Pregnancy. Placenta 2009, 30, 473–482. [Google Scholar] [CrossRef]
- Pankiewicz, K.; Fijałkowska, A.; Issat, T.; Maciejewski, T. Insight into the Key Points of Preeclampsia Pathophysiology: Uterine Artery Remodeling and the Role of MicroRNAs. Int. J. Mol. Sci. 2021, 22, 3132. [Google Scholar] [CrossRef] [PubMed]
- Zybina, T.G.; Frank, H.-G.; Biesterfeld, S.; Kaufmann, P. Genome multiplication of extravillous trophoblast cells in human placenta in the course of differentiation and invasion into endometrium and myometrium. II. Mech. Polyploidization Tsitologiya 2004, 46, 640–648. [Google Scholar]
- Pijnenborg, R.; Bland, J.M.; Robertson, W.B.; Dixon, G.; Brosens, I. The pattern of interstitial trophoblastic invasion of the myometrium in early human pregnancy. Placenta 1981, 2, 303–316. [Google Scholar] [CrossRef]
- Pijnenborg, R.; Vercruysse, L.; Carter, A.M. Deep trophoblast invasion and spiral artery remodelling in the placental bed of the lowland gorilla. Placenta 2011, 32, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Pijnenborg, R.; Vercruysse, L.; Carter, A.M. Deep trophoblast invasion and spiral artery remodelling in the placental bed of the chimpanzee. Placenta 2011, 32, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Pijnenborg, R.; D’Hooghe, T.; Vercruysse, L.; Bambra, C. Evaluation of trophoblast invasion in placental bed biopsies of the baboon, with immunohistochemical localisation of cytokeratin, fibronectin, and laminin. J. Med. Primatol. 1996, 25, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Blankenship, T.N.; Enders, A.C.; King, B.F. Trophoblastic invasion and the development of uteroplacental arteries in the macaque: Immunohistochemical localization of cytokeratins, desmin, type IV collagen, laminin, and fibronectin. Cell Tissue Res. 1993, 272, 227–236. [Google Scholar] [CrossRef]
- Carter, A.M. Comparative studies of placentation and immunology in non-human primates suggest a scenario for the evolution of deep trophoblast invasion and an explanation for human pregnancy disorders. Reproduction 2011, 141, 391–396. [Google Scholar] [CrossRef]
- Verkeste, C.; Slangen, B.; Daemen, M.; Van Straaten, H.; Kohnen, G.; Kaufmann, P.; Peeters, L. The extent of trophoblast invasion in the preplacental vasculature of the guinea-pig. Placenta 1998, 19, 49–54. [Google Scholar] [CrossRef]
- Redline, R.W.; Lu, C.Y. Localization of fetal major histocompatibility complex antigens and maternal leukocytes in murine placenta. Implications for maternal-fetal immunological relationship. Lab. Investig. 1989, 61, 27–36. [Google Scholar] [PubMed]
- Hu, D.; Cross, J.C. Development and function of trophoblast giant cells in the rodent placenta. Int. J. Dev. Biol. 2010, 54, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Ain, R.; Canham, L.N.; Soares, M.J. Gestation stage-dependent intrauterine trophoblast cell invasion in the rat and mouse: Novel endocrine phenotype and regulation. Dev. Biol. 2003, 260, 176–190. [Google Scholar] [CrossRef]
- Geusens, N.; Hering, L.; Verlohren, S.; Luyten, C.; Drijkoningen, K.; Taube, M.; Vercruysse, L.; Hanssens, M.; Dechend, R.; Pijnenborg, R. Changes in endovascular trophoblast invasion and spiral artery remodelling at term in a transgenic preeclamptic rat model. Placenta 2010, 31, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Medawar, P.B. Some immunological and endocrinological problems raised by the evolution of viviparity in vertebrates. Symp. Soc. Exp. Biol. 1953, 7, 320–338. [Google Scholar]
- Lash, G.E.; Robson, S.C.; Bulmer, J.N. Review: Functional role of uterine natural killer (uNK) cells in human early pregnancy decidua. Placenta 2010, 31, S87–S92. [Google Scholar] [CrossRef]
- Abi-Rached, L.; Kuhl, H.; Roos, C.; Ten Hallers, B.; Zhu, B.; Carbone, L.; Walter, L. A small, variable, and irregular killer cell Ig-like receptor locus accompanies the absence of MHC-C and MHC-G in gibbons. J. Immunol. 2010, 184, 1379–1391. [Google Scholar] [CrossRef]
- Golos, T.G.; Bondarenko, G.I.; Dambaeva, S.V.; Breburda, E.E.; Durning, M. On the role of placental Major Histocompatibility Complex and decidual leukocytes in implantation and pregnancy success using non-human primate models. Int. J. Dev. Biol. 2010, 54, 431–443. [Google Scholar] [CrossRef]
- Thomson, G.; Adams, E.J.; Parham, P. Evidence for an HLA-C -like locus in the orangutan Pongo pygmaeus. Immunogenetics 1999, 49, 865–871. [Google Scholar] [CrossRef]
- Aguilar, A.M.O.; Guethlein, L.A.; Adams, E.J.; Abi-Rached, L.; Moesta, A.; Parham, P. Coevolution of Killer Cell Ig-Like Receptors with HLA-C To Become the Major Variable Regulators of Human NK Cells. J. Immunol. 2010, 185, 4238–4251. [Google Scholar] [CrossRef]
- Penman, B.S.; Moffett, A.; Chazara, O.; Gupta, S.; Parham, P. Reproduction, infection and killer-cell immunoglobulin-like receptor haplotype evolution. Immunogenetics 2016, 68, 755–764. [Google Scholar] [CrossRef]
- Hiby, S.E.; Apps, R.; Sharkey, A.; Farrell, L.E.; Gardner, L.; Mulder, A.; Claas, F.H.; Walker, J.; Redman, C.C.; Morgan, L.; et al. Maternal activating KIRs protect against human reproductive failure mediated by fetal HLA-C2. J. Clin. Investig. 2010, 120, 4102–4110. [Google Scholar] [CrossRef]
- Hiby, S.E.; Walker, J.; O’Shaughnessy, K.M.; Redman, C.W.; Carrington, M.; Trowsdale, J.; Moffett, A. Combinations of Maternal KIR and Fetal HLA-C Genes Influence the Risk of Preeclampsia and Reproductive Success. J. Exp. Med. 2004, 200, 957–965. [Google Scholar] [CrossRef]
- Khakoo, S.; Rajalingam, R.; Shum, B.P.; Weidenbach, K.; Flodin, L.; Muir, D.G.; Canavez, F.; Cooper, S.L.; Valiante, N.M.; Lanier, L.L.; et al. Rapid Evolution of NK Cell Receptor Systems Demonstrated by Comparison of Chimpanzees and Humans. Immunity 2000, 12, 687–698. [Google Scholar] [CrossRef]
- Wroblewski, E.E.; Parham, P.; Guethlein, L.A. Two to Tango: Co-evolution of Hominid Natural Killer Cell Receptors and MHC. Front. Immunol. 2019, 10, 177. [Google Scholar] [CrossRef]
- Le Bouteiller, P. HLA-G in the human placenta: Expression and potential functions. Biochem. Soc. Trans. 2000, 28, 208–212. [Google Scholar] [CrossRef]
- Hunt, J.S.; Langat, D.L. HLA-G: A human pregnancy-related immunomodulator. Curr. Opin. Pharmacol. 2009, 9, 462–469. [Google Scholar] [CrossRef]
- Castro, M.J.; Morales, P.; Fernández-Soria, V.; Suarez, B.; Recio, M.J.; Alvarez, M.; Arnaiz-Villena, A. Allelic diversity at the primate MHC-G locus: Exon 3 bears stop codons in all Cercopithecinae sequences. Immunogenetics 1996, 43, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Bondarenko, G.I.; Dambaeva, S.V.; Grendell, R.L.; Hughes, A.L.; Durning, M.; Garthwaite, M.A.; Golos, T.G. Characterization of cynomolgus and vervet monkey placental MHC class I expression: Diversity of the nonhuman primate AG locus. Immunogenetics 2009, 61, 431–442. [Google Scholar] [CrossRef]
- Croy, B.A.; Luross, J.A.; Guimond, M.J.; Hunt, J.S. Uterine natural killer cells: Insights into lineage relationships and functions from studies of pregnancies in mutant and transgenic mice. Nat. Immun. 1996, 15, 22–33. [Google Scholar]
- Burke, S.; Barrette, V.F.; Gravel, J.; Carter, A.L.I.; Hatta, K.; Zhang, J.; Chen, Z.; Leno-Durán, E.; Bianco, J.; Leonard, S.; et al. Uterine NK Cells, Spiral Artery Modification and the Regulation of Blood Pressure During Mouse Pregnancy. Am. J. Reprod. Immunol. 2010, 63, 472–481. [Google Scholar] [CrossRef]
- Croy, B.; Esadeg, S.; Chantakru, S.; Heuvel, M.V.D.; Paffaro, V.A.; He, H.; Black, G.P.; Ashkar, A.; Kiso, Y.; Zhang, J. Update on pathways regulating the activation of uterine Natural Killer cells, their interactions with decidual spiral arteries and homing of their precursors to the uterus. J. Reprod. Immunol. 2003, 59, 175–191. [Google Scholar] [CrossRef]
- Nguyen-Lefebvre, A.T.; Ajith, A.; Portik-Dobos, V.; Horuzsko, D.D.; Mulloy, L.L.; Horuzsko, A. Mouse models for studies of HLA-G functions in basic science and pre-clinical research. Hum. Immunol. 2016, 77, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Napso, T.; Yong, H.E.J.; Lopez-Tello, J.; Sferruzzi-Perri, A. The Role of Placental Hormones in Mediating Maternal Adaptations to Support Pregnancy and Lactation. Front. Physiol. 2018, 9, 1091. [Google Scholar] [CrossRef] [PubMed]
- Maston, G.A.; Ruvolo, M. Chorionic gonadotropin has a recent origin within primates and an evolutionary history of selection. Mol. Biol. Evol. 2002, 19, 320–335. [Google Scholar] [CrossRef] [PubMed]
- Chopineau, M.; Stewart, F.; Allen, W.R. Cloning and analysis of the cDNA encoding the horse and donkey luteinizing hormone beta-subunits. Gene 1995, 160, 253–256. [Google Scholar] [CrossRef]
- Chen, E.Y.; Liao, Y.-C.; Smith, D.H.; Barrera-Saldaña, H.A.; Gelinas, R.E.; Seeburg, P.H. The human growth hormone locus: Nucleotide sequence, biology, and evolution. Genomics 1989, 4, 479–497. [Google Scholar] [CrossRef]
- Frankenne, F.; Closset, J.; Gomez, F.; Scippo, M.L.; Smal, J.; Hennen, G. The Physiology of Growth Hormones (GHs) in Pregnant Women and Partial Characterization of the Placental GH Variant. J. Clin. Endocrinol. Metab. 1988, 66, 1171–1180. [Google Scholar] [CrossRef] [PubMed]
- de Mendoza, A.R.; Escobedo, D.E.; Dávila, I.M.; Saldaña, H. Expansion and divergence of the GH locus between spider monkey and chimpanzee. Gene 2004, 336, 185–193. [Google Scholar] [CrossRef]
- Wallis, O.C.; Wallis, M. Evolution of growth hormone in primates: The GH gene clusters of the New World monkeys marmoset (Callithrix jacchus) and white-fronted capuchin (Cebus albifrons). J. Mol. Evol. 2006, 63, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Papper, Z.; Jameson, N.M.; Romero, R.; Weckle, A.L.; Mittal, P.; Benirschke, K.; Santolaya-Forgas, J.; Uddin, M.; Haig, D.; Goodman, M.; et al. Ancient origin of placental expression in the growth hormone genes of anthropoid primates. Proc. Natl. Acad. Sci. USA 2009, 106, 17083–17088. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.J. The prolactin and growth hormone families: Pregnancy-specific hormones/cytokines at the maternal-fetal interface. Reprod. Biol. Endocrinol. 2004, 2, 51. [Google Scholar] [CrossRef] [PubMed]
- Csapo, A. Progesterone block. Am. J. Anat. 1956, 98, 273–291. [Google Scholar] [CrossRef]
- Thorburn, G.D.; Challis, J.R.G.; Robinson, J.S. Endocrine Control of Parturition; Springer Science and Business Media LLC: Tokyo, Japan, 1977; pp. 653–732. [Google Scholar]
- Merlino, A.A.; Welsh, T.N.; Tan, H.; Yi, L.J.; Cannon, V.; Mercer, B.M.; Mesiano, S. Nuclear Progesterone Receptors in the Human Pregnancy Myometrium: Evidence that Parturition Involves Functional Progesterone Withdrawal Mediated by Increased Expression of Progesterone Receptor-A. J. Clin. Endocrinol. Metab. 2007, 92, 1927–1933. [Google Scholar] [CrossRef] [PubMed]
- Shynlova, O.; Nadeem, L.; Zhang, J.; Dunk, C.; Lye, S. Myometrial activation: Novel concepts underlying labor. Placenta 2020, 92, 28–36. [Google Scholar] [CrossRef]
- Marinić, M.; Lynch, V.J. Relaxed constraint and functional divergence of the progesterone receptor (PGR) in the human stem-lineage. PLoS Genet. 2020, 16, e1008666. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Opazo, J.C.; Erez, O.; Uddin, M.; Santolaya-Forgas, J.; Goodman, M.; Grossman, L.I.; Romero, R.; Wildman, D.E. The human progesterone receptor shows evidence of adaptive evolution associated with its ability to act as a transcription factor. Mol. Phylogenet. Evol. 2008, 47, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Blanks, A.; Brosens, J. Progesterone Action in the Myometrium and Decidua in Preterm Birth. Facts Views Vis. ObGyn 2012, 4, 188–194. [Google Scholar]
- Challis, J.R.; John Davies, I.; Benirschke, K.; Hendrickx, A.G.; Ryan, K.J. The concentrations of progesterone, estrone and estradiol-17 beta in the peripheral plasma of the rhesus monkey during the final third of gestation, and after the induction of abortion with PGF 2 alpha. Endocrinology 1974, 95, 547–553. [Google Scholar] [CrossRef]
- Mitchell, B.F.; Taggart, M.J. Are animal models relevant to key aspects of human parturition? Am. J. Physiol. Integr. Comp. Physiol. 2009, 297, R525–R545. [Google Scholar] [CrossRef]
- Martin, R.D. The evolution of human reproduction: A primatological perspective. Am. J. Phys. Anthr. 2007, 134, 59–84. [Google Scholar] [CrossRef]
- Roberts, M. Growth, development, and parental care in the western tarsier (Tarsius bancanus) in captivity: Evidence for a “slow” life-history and nonmonogamous mating system. Int. J. Primatol. 1994, 15, 1–28. [Google Scholar] [CrossRef]
- Abbott, D.H.; Barnett, D.K.; Colman, R.J.; Yamamoto, M.E.; Schultz-Darken, N.J. Aspects of common marmoset basic biology and life history important for biomedical research. Comp. Med. 2003, 53, 339–350. [Google Scholar]
- Tardif, S.D.; Smucny, D.A.; Abbott, D.H.; Mansfield, K.; Schultz-Darken, N.; Yamamoto, M.E. Reproduction in captive common marmosets (Callithrix jacchus). Comp. Med. 2003, 53, 364–368. [Google Scholar]
- Altmann, J.; Samuels, A. Costs of maternal care: Infant-carrying in baboons. Behav. Ecol. Sociobiol. 1992, 29, 391–398. [Google Scholar] [CrossRef]
- Rhine, R.J.; Norton, G.W.; Wynn, G.M.; Wynn, R.D. Weaning of free-ranging infant baboons (Papio cynocephalus) as indicated by one-zero and instantaneous sampling of feeding. Int. J. Primatol. 1985, 6, 491–499. [Google Scholar] [CrossRef]
- Gesquiere, L.R.; Altmann, J.; Archie, E.A.; Alberts, S.C. Interbirth intervals in wild baboons: Environmental predictors and hormonal correlates. Am. J. Phys. Anthr. 2018, 166, 107–126. [Google Scholar] [CrossRef] [PubMed]
- Lappan, S. Patterns of Infant Care in Wild Siamangs (Symphalangus syndactylus) in Southern Sumatra. In The Gibbons; Springer Science and Business Media LLC: Tokyo, Japan, 2009; pp. 327–345. [Google Scholar]
- McConkey, K. Bornean Orangutan (Pongo pygmaeus). In World Atlas of Great Apes and Their Conservation; Caldecott, J.O., Miles, L., Eds.; University of California Press, in Association with UNEP-WCMC: Berkeley, CA, USA, 2005; pp. 161–183. [Google Scholar]
- Tullner, W.W. Comparative Aspects of Primate Chorionic Gonadotropins. In Reproductive Biology of the Primates; Luckett, W.P., Ed.; Karger: Basel, Switzerland, 1974; pp. 235–257. [Google Scholar]
- Ferriss, S.; Robbins, M.M.; Williamson, E.A. Eastern gorilla (Gorilla beringei). In World Atlas of Great Apes and Their Conservation; Caldecott, J.O., Miles, L., Eds.; University of California Press, in Association with UNEP-WCMC: Berkeley, CA, USA, 2005; pp. 129–152. [Google Scholar]
- Canington, S.L. Gorilla beringei (Primates: Hominidae). Mamm. Species 2018, 967, 119–133. [Google Scholar] [CrossRef]
- Inskipp, T. Chimpanzee (Pan troglodytes). In World Atlas of Great Apes and their Conservation; Caldecott, J.O., Miles, L., Eds.; University of California Press, in Association with UNEP-WCMC: Berkeley, CA, USA, 2005; pp. 53–81. [Google Scholar]
- Jones, C.; Jones, C.A.; Knox Jones, J.; Wilson, D.E. Pan troglodytes. Mamm. Species 1996, 529, 1–9. [Google Scholar] [CrossRef]
- Martin, R.D.; MacLarnon, A. Gestation period, neonatal size and maternal investment in placental mammals. Nature 1985, 313, 220–223. [Google Scholar] [CrossRef]
- Trevathan, W. Primate pelvic anatomy and implications for birth. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140065. [Google Scholar] [CrossRef] [PubMed]
- Sakai, T.; Hirata, S.; Fuwa, K.; Sugama, K.; Kusunoki, K.; Makishima, H.; Eguchi, T.; Yamada, S.; Ogihara, N.; Takeshita, H. Fetal brain development in chimpanzees versus humans. Curr. Biol. 2012, 22, R791–R792. [Google Scholar] [CrossRef]
- Martin, R.D. How We Do It: The Evolution and Future of Human Reproduction, xii; Basic Books: New York, NY, USA, 2013; p. 304. [Google Scholar]
- Chatterjee, H.J.; Ho, S.Y.W.; Barnes, I.; Groves, C. Estimating the phylogeny and divergence times of primates using a supermatrix approach. BMC Evol. Biol. 2009, 9, 259. [Google Scholar] [CrossRef] [PubMed]
- Stout, C.; Lemmon, W. Glomerular capillary endothelial swelling in a pregnant chimpanzee. Am. J. Obstet. Gynecol. 1969, 105, 212–215. [Google Scholar] [CrossRef]
- Makris, A.; Thornton, C.; Thompson, J.; Thomson, S.; Martin, R.; Ogle, R.; Waugh, R.; McKenzie, P.; Kirwan, P.; Hennessy, A. Uteroplacental ischemia results in proteinuric hypertension and elevated sFLT-1. Kidney Int. 2007, 71, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, M.K.; Goleva, S.B.; Beierwaltes, W.H.; Osborn, J.L. Renal vascular and glomerular pathologies associated with spontaneous hypertension in the nonhuman primate Chlorocebus aethiops sabaeus. Am. J. Physiol. Integr. Comp. Physiol. 2017, 313, R211–R218. [Google Scholar] [CrossRef]
- Palmer, A.E.; London, W.T.; Sly, D.L.; Rice, J.M. Spontaneous preeclamptic toxemia of pregnancy in the patas monkey (Erythrocebus patas). Lab. Anim. Sci. 1979, 29, 102–106. [Google Scholar] [PubMed]
- Rutherford, J.N.; Tardif, S.D. Placental efficiency and intrauterine resource allocation strategies in the common marmoset pregnancy. Am. J. Phys. Anthr. 2008, 137, 60–68. [Google Scholar] [CrossRef]
- Mansfield, K. Marmoset models commonly used in biomedical research. Comp. Med. 2003, 53, 383–392. [Google Scholar]
- Schmidt, A.; Prieto, D.M.M.; Pastuschek, J.; Fröhlich, K.; Markert, U.R. Only humans have human placentas: Molecular differences between mice and humans. J. Reprod. Immunol. 2015, 108, 65–71. [Google Scholar] [CrossRef]
- Malassine, A.; Frendo, J.-L.; Evain-Brion, D. A comparison of placental development and endocrine functions between the human and mouse model. Hum. Reprod. Updat 2003, 9, 531–539. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clade | Species | Common Name | Length of Gestation | Parental Care | References |
|---|---|---|---|---|---|
| Tarsiers | Cephalopacus bancanus | Western tarsier | 178 days | Nutritional and social independence by 60 days | [95] |
| New World monkeys | Callithrix jacchus | Common marmoset | 143–144 days | Independent movement by 3 weeks; weaning by 3 months | [96,97] |
| Old World monkeys | Papio cynocephalus | Yellow baboon | 178 ± 6 days | Milk supplemented early with plant foods; fully weaned after about a year; carried for 8 months | [98,99,100] |
| Lesser apes | Symphalangus syndactylus | Siamang | 230–235 days | Partial weaning at 6 months; travel independently by 1 year | [101] |
| Great apes | Pongo pygmaeus | Bornean orangutan | 275 days | Partial weaning by 11 months; fully independent at 7–10 years | [102,103] |
| Gorilla beringei | Eastern gorilla | 255 days | Weaning at 3–4 years | [103,104,105] | |
| Pan troglodytes | Chimpanzee | 196–260 days | Weaning at 10 months; dependent on mother for 5 years | [103,106,107] |
| Character | Taxonomic Clade | Branching Point | Geological Period or Epoch | Comments |
|---|---|---|---|---|
| Invasive placentation | Eutheria | 98.5 Mya | Late Cretaceous | |
| Decidual reaction | Eutheria | 98.5 Mya | Late Cretaceous | An inflammatory response in marsupials |
| Persistence of decidual stromal cells | Euarchontoglires (includes rodents and primates) | 91.8 Mya | Late Cretaceous | |
| Precocious extraembryonic mesoderm | Haplorrhini | 44.8 Mya | Middle Eocene | |
| Secondary yolk sac | Haplorrhini | 44.8 Mya | Middle Eocene | |
| Allantoic stalk | Haplorrhini | 44.8 Mya | Middle Eocene | Many mammals have an allantoic sac |
| Haemomonochorial placentation | Haplorrhini | 44.8 Mya | Middle Eocene | |
| Syncytin-2 env gene | Haplorrhini | 44.8 Mya | Middle Eocene | |
| Chorionic gonadotropin | Haplorrhini | 44.8 Mya | Middle Eocene | |
| Placental lactogens and growth hormone | Haplorrhini | 44.8 Mya | Middle Eocene | Vary between primate lineages |
| Trophoblast invasion by intravascular route | Old World monkeys and apes | 29.8 Mya | Oligocene | |
| Villous placentation with an intervillous space | Old World monkeys and apes | 29.8 Mya | Oligocene | Trabecular placentation in tarsiers and NW monkeys |
| Interstitial implantation | Lesser and greater apes | 20.2 Mya | Early Miocene | |
| Syncytin-1 env gene | Lesser and greater apes | 20.2 Mya | Early Miocene | |
| Trophoblast invasion by interstitial route | Great apes | 15.1 Mya | Middle Miocene | |
| HLA-C | Great apes | 15.1 Mya | Middle Miocene |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carter, A.M. Unique Aspects of Human Placentation. Int. J. Mol. Sci. 2021, 22, 8099. https://doi.org/10.3390/ijms22158099
Carter AM. Unique Aspects of Human Placentation. International Journal of Molecular Sciences. 2021; 22(15):8099. https://doi.org/10.3390/ijms22158099
Chicago/Turabian StyleCarter, Anthony M. 2021. "Unique Aspects of Human Placentation" International Journal of Molecular Sciences 22, no. 15: 8099. https://doi.org/10.3390/ijms22158099
APA StyleCarter, A. M. (2021). Unique Aspects of Human Placentation. International Journal of Molecular Sciences, 22(15), 8099. https://doi.org/10.3390/ijms22158099