
Exercise Pills for Drug Addiction: Forced Moderate Endurance Exercise Inhibits Methamphetamine-Induced Hyperactivity through the Striatal Glutamatergic Signaling Pathway in Male Sprague Dawley Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. METH Administration and Exercise Training
2.2.1. METH Administration
2.2.2. Exercise Training
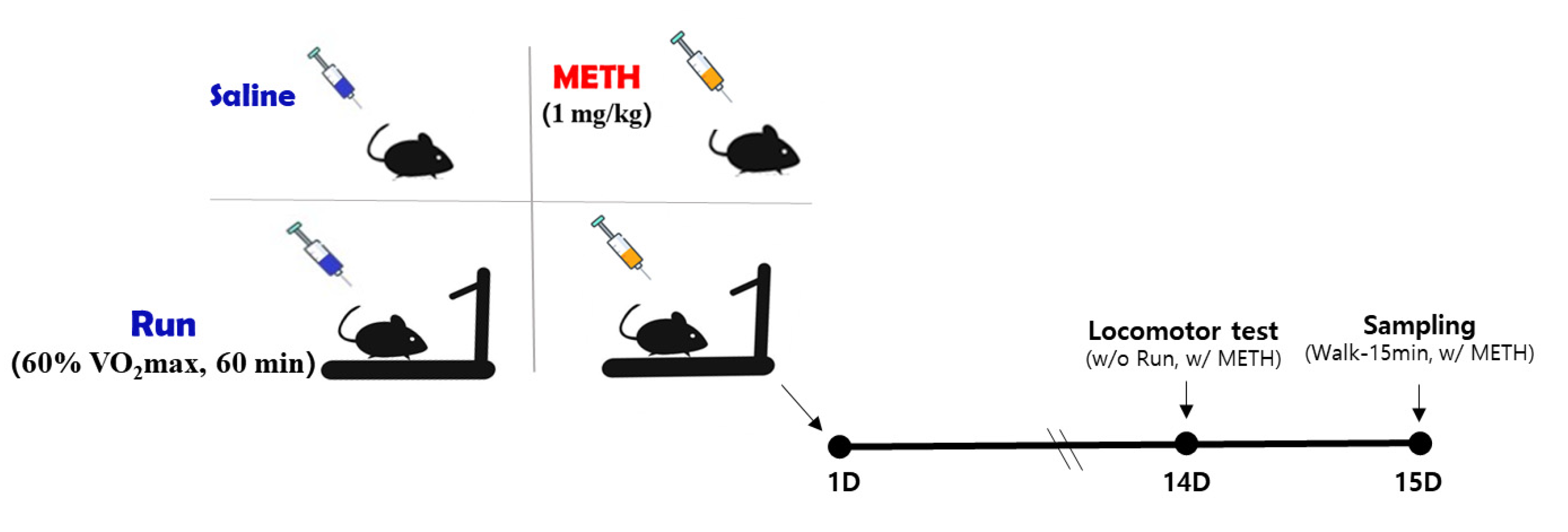
2.3. Experimental Design
2.4. Locomotor Activity
2.5. Sample Preparation
2.6. Western Blotting
2.7. Striatal Dopamine Levels
2.8. Statistical Analysis
3. Results
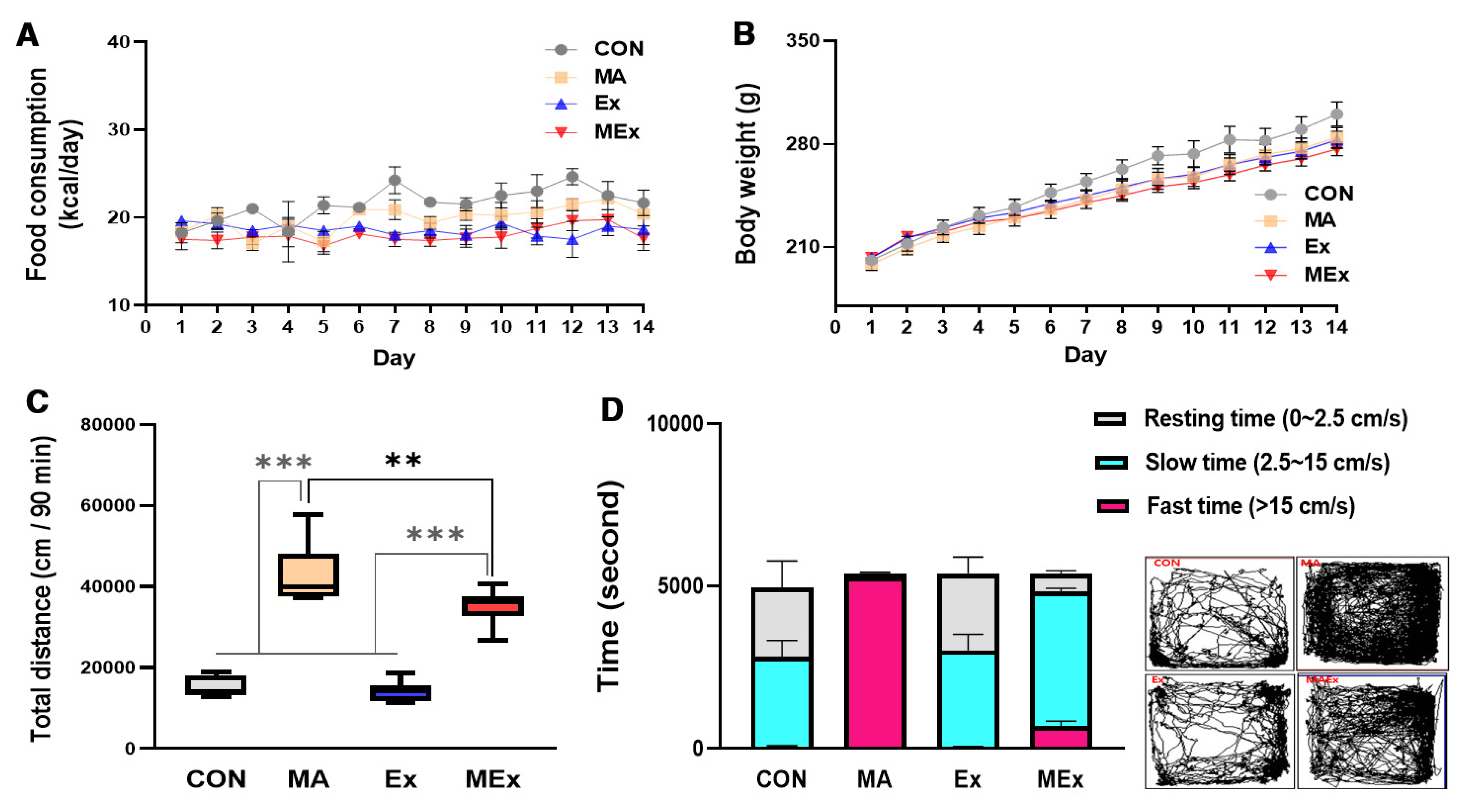
3.1. Forced Moderate Endurance Exercise Significantly Reduced METH-Induced Hyperactivity
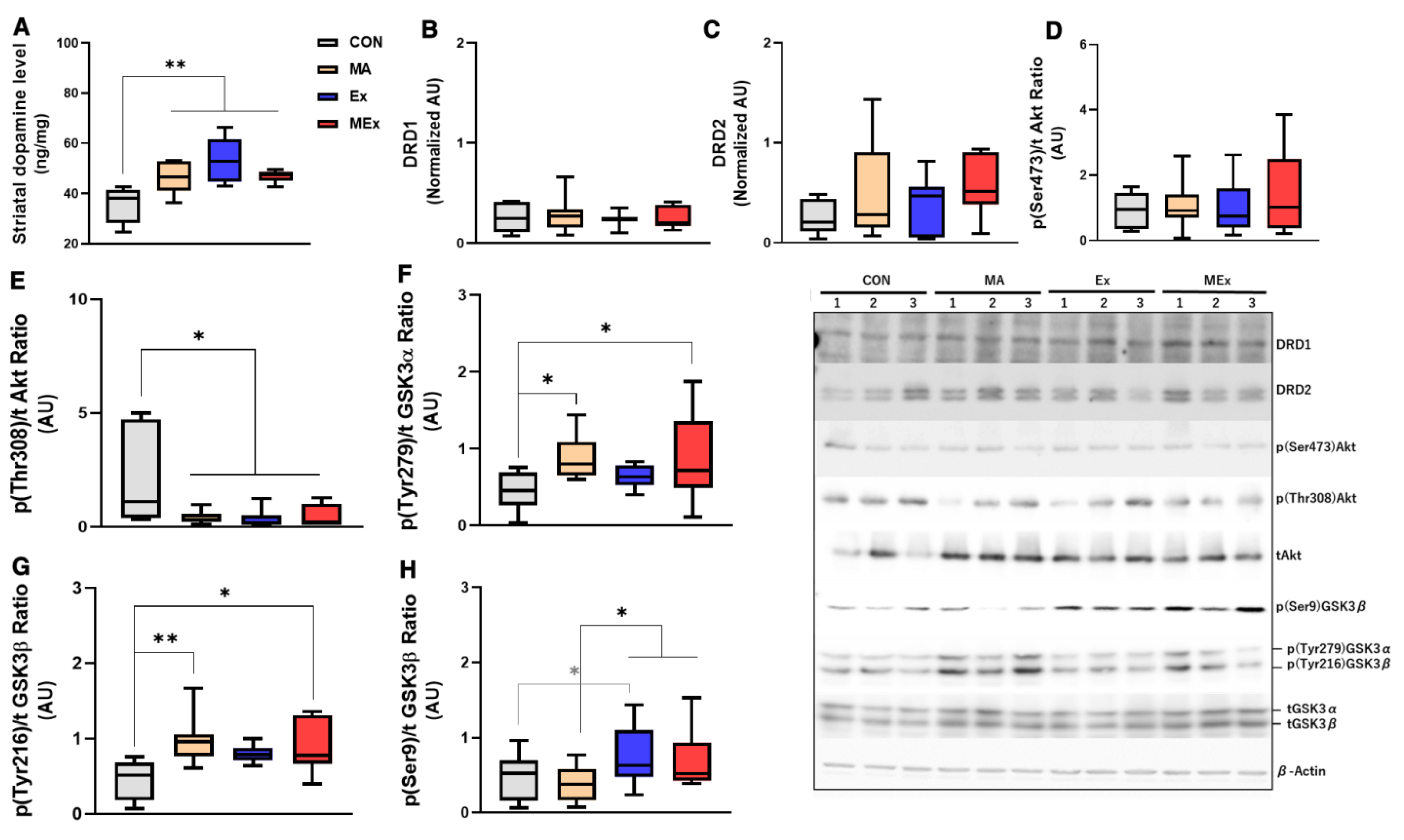
3.2. Forced Moderate Endurance Exercise Did Not Affect the METH-Induced Dopaminergic Signaling Pathway
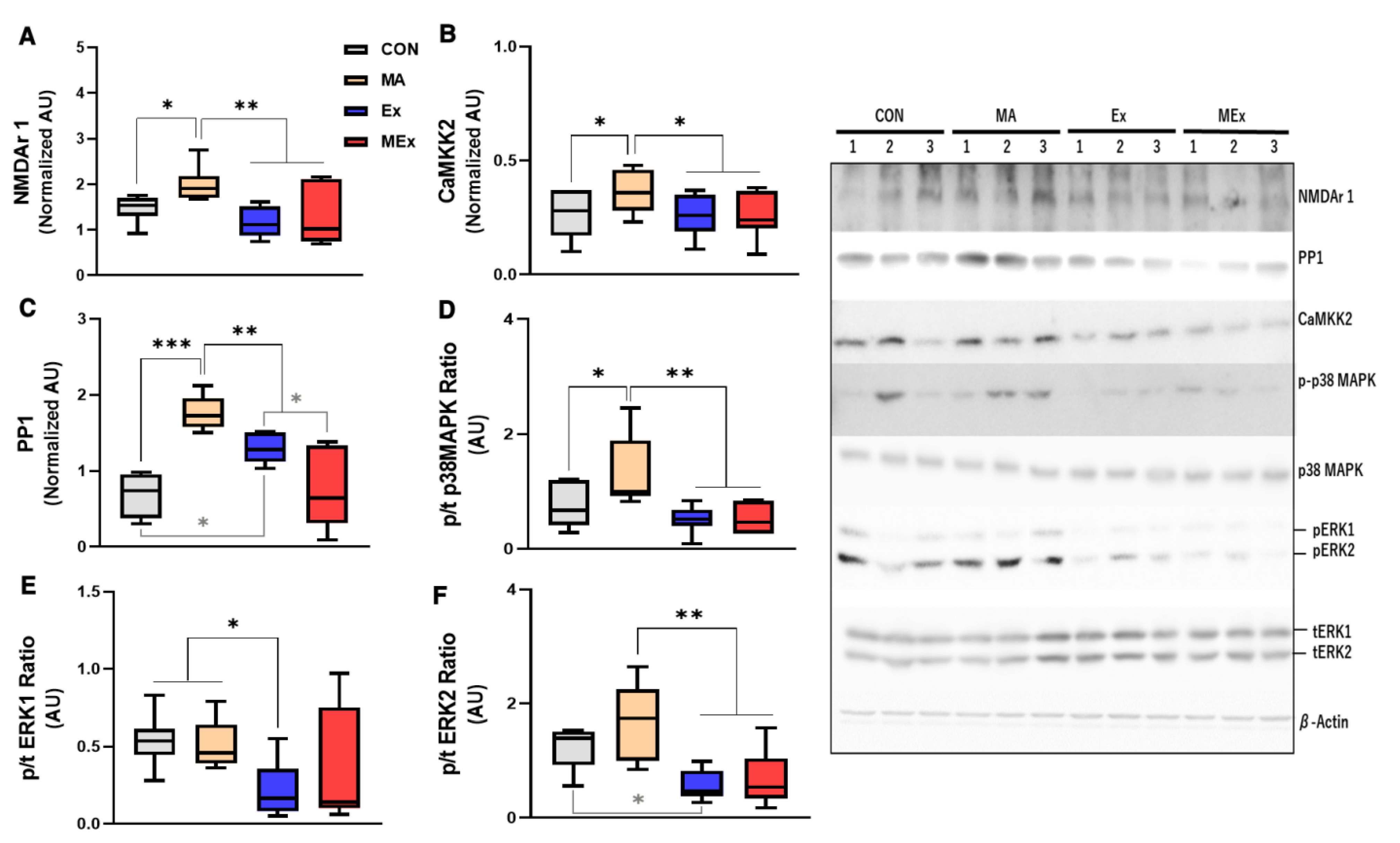
3.3. Forced Moderate Endurance Exercise Affected the METH-Induced Glutamatergic Signaling Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- United Nations Office on Drugs and Crime. World Drug Report-2016; United Nations: New York, NY, USA, 2016. [Google Scholar]
- Courtney, K.E.; Ray, L.A. Methamphetamine: An update on epidemiology, pharmacology, clinical phenomenology, and treatment literature. Drug Alcohol Depend. 2014, 143, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawson, R.A. Current research on the epidemiology, medical and psychiatric effects, and treatment of methamphetamine use. J. Food Drug Anal. 2013, 21, S77–S81. [Google Scholar] [CrossRef] [Green Version]
- Morais, A.P.D.; Pita, I.R.; Fontes-Ribeiro, C.A.; Pereira, F.C. The neurobiological mechanisms of physical exercise in methamphetamine addiction. CNS Neurosci. Ther. 2018, 24, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.S.; Cha, E.H.; Park, B.; Ha, E.; Seo, J.H. PBN inhibits a detrimental effect of methamphetamine on brain endothelial cells by alleviating the generation of reactive oxygen species. Arch. Pharm. Res. 2020, 43, 1347–1355. [Google Scholar] [CrossRef]
- Kirkpatrick, M.G.; Haney, M.; Vosburg, S.K.; Comer, S.D.; Foltin, R.W.; Hart, C.L. Methamphetamine self-administration by humans subjected to abrupt shift and sleep schedule changes. Psychopharmacology 2009, 203, 771–780. [Google Scholar] [CrossRef]
- Mendelson, J.; Rawson, R.; Newton, T.; Galloway, G.; de Wit, H.; Dewey, S.L.; Hart, C.L.; Epstein, D.H. Treatment of methamphetamine dependence. Mayo Clin. Proc. 2008, 83, 369–370. [Google Scholar] [CrossRef] [Green Version]
- Perez, A.Y.; Kirkpatrick, M.G.; Gunderson, E.W.; Marrone, G.; Silver, R.; Foltin, R.W.; Hart, C.L. Residual effects of intranasal methamphetamine on sleep, mood, and performance. Drug Alcohol Depend. 2008, 94, 258–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, M.D.; Tangalakis, K.; Antonipillai, J.; Stojanovska, L.; Nurgali, K.; Apostolopoulos, V. Methamphetamine: Effects on the brain, gut and immune system. Pharmacol. Res. 2017, 120, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Halpin, L.E.; Collins, S.A.; Yamamoto, B.K. Neurotoxicity of methamphetamine and 3,4-methylenedioxymethamphetamine. Life Sci. 2014, 97, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, A.C.; Groman, S.M.; Morales, A.M.; London, E.D. An evaluation of the evidence that methamphetamine abuse causes cognitive decline in humans. Neuropsychopharmacology 2013, 38, 259–274. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.J.; Song, S.H.; Kim, S.; Jang, W.J.; Jeong, C.H.; Lee, S. Characteristics of Korean patients with methamphetamine use disorder based on the quantitative analysis of methamphetamine and amphetamine in hair. Arch. Pharm. Res. 2020, 43, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Somkuwar, S.S.; Staples, M.C.; Fannon, M.J.; Ghofranian, A.; Mandyam, C.D. Evaluating Exercise as a Therapeutic Intervention for Methamphetamine Addiction-Like Behavior. Brain Plast. 2015, 1, 63–81. [Google Scholar] [CrossRef] [Green Version]
- Koob, G.F.; Volkow, N.D. Neurocircuitry of addiction. Neuropsychopharmacology 2010, 35, 217–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalivas, P.W.; Volkow, N.D. The neural basis of addiction: A pathology of motivation and choice. Am. J. Psychiatry 2005, 162, 1403–1413. [Google Scholar] [CrossRef]
- Brown, R.A.; Abrantes, A.M.; Read, J.P.; Marcus, B.H.; Jakicic, J.; Strong, D.R.; Oakley, J.R.; Ramsey, S.E.; Kahler, C.W.; Stuart, G.G.; et al. A Pilot Study of Aerobic Exercise as an Adjunctive Treatment for Drug Dependence. Ment. Health Phys. Act. 2010, 3, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, W.J.; Peterson, A.B.; Sanchez, V.; Abel, J.; Smith, M.A. Exercise as a novel treatment for drug addiction: A neurobiological and stage-dependent hypothesis. Neurosci. Biobehav. Rev. 2013, 37, 1622–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawson, R.A.; Chudzynski, J.; Mooney, L.; Gonzales, R.; Ang, A.; Dickerson, D.; Penate, J.; Salem, B.A.; Dolezal, B.; Cooper, C.B. Impact of an exercise intervention on methamphetamine use outcomes post-residential treatment care. Drug Alcohol Depend. 2015, 156, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Rawson, R.A.; Chudzynski, J.; Gonzales, R.; Mooney, L.; Dickerson, D.; Ang, A.; Dolezal, B.; Cooper, C.B. The Impact of Exercise On Depression and Anxiety Symptoms Among Abstinent Methamphetamine-Dependent Individuals in A Residential Treatment Setting. J. Subst. Abus. Treat. 2015, 57, 36–40. [Google Scholar] [CrossRef] [Green Version]
- Haglund, M.; Ang, A.; Mooney, L.; Gonzales, R.; Chudzynski, J.; Cooper, C.B.; Dolezal, B.A.; Gitlin, M.; Rawson, R.A. Predictors of depression outcomes among abstinent methamphetamine-dependent individuals exposed to an exercise intervention. Am. J. Addict. 2015, 24, 246–251. [Google Scholar] [CrossRef]
- Muller, A.E.; Clausen, T. Group exercise to improve quality of life among substance use disorder patients. Scand. J. Public Health 2015, 43, 146–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koob, G.F. Addiction is a Reward Deficit and Stress Surfeit Disorder. Front. Psychiatry 2013, 4, 72. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.M.; Knutson, D.; Haines, D. Common problems in patients recovering from chemical dependency. Am. Fam. Phys. 2003, 68, 1971–1978. [Google Scholar]
- Polter, A.M.; Kauer, J.A. Stress and VTA synapses: Implications for addiction and depression. Eur. J. Neurosci. 2014, 39, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Dolezal, B.A.; Chudzynski, J.; Dickerson, D.; Mooney, L.; Rawson, R.A.; Garfinkel, A.; Cooper, C.B. Exercise training improves heart rate variability after methamphetamine dependency. Med. Sci. Sports Exerc. 2014, 46, 1057–1066. [Google Scholar] [CrossRef] [Green Version]
- Dolezal, B.A.; Chudzynski, J.; Storer, T.W.; Abrazado, M.; Penate, J.; Mooney, L.; Dickerson, D.; Rawson, R.A.; Cooper, C.B. Eight weeks of exercise training improves fitness measures in methamphetamine-dependent individuals in residential treatment. J. Addict. Med. 2013, 7, 122–128. [Google Scholar] [CrossRef]
- Miller, M.L.; Vaillancourt, B.D.; Wright, M.J., Jr.; Aarde, S.M.; Vandewater, S.A.; Creehan, K.M.; Taffe, M.A. Reciprocal inhibitory effects of intravenous d-methamphetamine self-administration and wheel activity in rats. Drug Alcohol Depend. 2012, 121, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Aarde, S.M.; Miller, M.L.; Creehan, K.M.; Vandewater, S.A.; Taffe, M.A. One day access to a running wheel reduces self-administration of D-methamphetamine, MDMA and methylone. Drug Alcohol Depend. 2015, 151, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Engelmann, A.J.; Aparicio, M.B.; Kim, A.; Sobieraj, J.C.; Yuan, C.J.; Grant, Y.; Mandyam, C.D. Chronic wheel running reduces maladaptive patterns of methamphetamine intake: Regulation by attenuation of methamphetamine-induced neuronal nitric oxide synthase. Brain Struct. Funct. 2014, 219, 657–672. [Google Scholar] [CrossRef] [Green Version]
- O’Dell, S.J.; Galvez, B.A.; Ball, A.J.; Marshall, J.F. Running wheel exercise ameliorates methamphetamine-induced damage to dopamine and serotonin terminals. Synapse 2012, 66, 71–80. [Google Scholar] [CrossRef]
- Radak, Z.; Chung, H.Y.; Goto, S. Systemic adaptation to oxidative challenge induced by regular exercise. Free Radic Biol. Med. 2008, 44, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, A.S., Jr.; Tuon, T.; Soares, F.S.; da Rocha, L.G.; Silveira, P.C.; Pinho, R.A. The effect of n-acetylcysteine and deferoxamine on exercise-induced oxidative damage in striatum and hippocampus of mice. Neurochem. Res. 2008, 33, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Camiletti-Moirón, D.; Aparicio, V.A.; Aranda, P.; Radak, Z. Does exercise reduce brain oxidative stress? A systematic review. Scand. J. Med. Sci. Sports 2013, 23, e202–e212. [Google Scholar] [CrossRef] [Green Version]
- Speck, A.E.; Tromm, C.B.; Pozzi, B.G.; Paganini, C.S.; Tuon, T.; Silveira, P.C.; Aguiar, A.S., Jr.; Pinho, R.A. The dose-dependent antioxidant effects of physical exercise in the hippocampus of mice. Neurochem. Res. 2014, 39, 1496–1501. [Google Scholar] [CrossRef]
- Sobieraj, J.C.; Kim, A.; Fannon, M.J.; Mandyam, C.D. Chronic wheel running-induced reduction of extinction and reinstatement of methamphetamine seeking in methamphetamine dependent rats is associated with reduced number of periaqueductal gray dopamine neurons. Brain Struct. Funct. 2016, 221, 261–276. [Google Scholar] [CrossRef] [Green Version]
- Mandyam, C.D.; Wee, S.; Eisch, A.J.; Richardson, H.N.; Koob, G.F. Methamphetamine self-administration and voluntary exercise have opposing effects on medial prefrontal cortex gliogenesis. J. Neurosci. 2007, 27, 11442–11450. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Levine, H.; Toborek, M. Exercise protects against methamphetamine-induced aberrant neurogenesis. Sci. Rep. 2016, 6, 34111. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Wang, Y.; Wang, Y.; Li, R.; Zhou, C. Impact of physical exercise on substance use disorders: A meta-analysis. PLoS ONE 2014, 9, e110728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filip, M.; Frankowska, M.; Gołda, A.; Zaniewska, M.; Vetulani, J.; Przegaliński, E. Various GABA-mimetic drugs differently affect cocaine-evoked hyperlocomotion and sensitization. Eur. J. Pharmacol. 2006, 541, 163–170. [Google Scholar] [CrossRef]
- Fukushima, S.; Shen, H.; Hata, H.; Ohara, A.; Ohmi, K.; Ikeda, K.; Numachi, Y.; Kobayashi, H.; Hall, F.S.; Uhl, G.R.; et al. Methamphetamine-induced locomotor activity and sensitization in dopamine transporter and vesicular monoamine transporter 2 double mutant mice. Psychopharmacology 2007, 193, 55–62. [Google Scholar] [CrossRef]
- Shimosato, K.; Ohkuma, S. Simultaneous monitoring of conditioned place preference and locomotor sensitization following repeated administration of cocaine and methamphetamine. Pharmacol. Biochem. Behav. 2000, 66, 285–292. [Google Scholar] [CrossRef]
- Hooks, M.S.; Jones, G.H.; Hemby, S.E.; Justice, J.B., Jr. Environmental and pharmacological sensitization: Effects of repeated administration of systemic or intra-nucleus accumbens cocaine. Psychopharmacology 1993, 111, 109–116. [Google Scholar] [CrossRef]
- Mattingly, B.A.; Rice, L.L.; Langfels, M.; Fields, S.E. Repeated treatments with 7-OH-DPAT: Context-independent behavioral sensitization and conditioned hyperactivity. Pharmacol. Biochem. Behav. 2000, 65, 241–246. [Google Scholar] [CrossRef]
- Shen, F.; Meredith, G.E.; Napier, T.C. Amphetamine-induced place preference and conditioned motor sensitization requires activation of tyrosine kinase receptors in the hippocampus. J. Neurosci. 2006, 26, 11041–11051. [Google Scholar] [CrossRef]
- Bello, E.P.; Mateo, Y.; Gelman, D.M.; Noaín, D.; Shin, J.H.; Low, M.J.; Alvarez, V.A.; Lovinger, D.M.; Rubinstein, M. Cocaine supersensitivity and enhanced motivation for reward in mice lacking dopamine D2 autoreceptors. Nat. Neurosci. 2011, 14, 1033–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pak, A.C.; Ashby, C.R., Jr.; Heidbreder, C.A.; Pilla, M.; Gilbert, J.; Xi, Z.X.; Gardner, E.L. The selective dopamine D3 receptor antagonist SB-277011A reduces nicotine-enhanced brain reward and nicotine-paired environmental cue functions. Int. J. Neuropsychopharmacol. 2006, 9, 585–602. [Google Scholar] [CrossRef]
- Bisagno, V.; González, B.; Urbano, F.J. Cognitive enhancers versus addictive psychostimulants: The good and bad side of dopamine on prefrontal cortical circuits. Pharmacol. Res. 2016, 109, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Nestler, H.P. International Proteolysis Society—Drug discovery and application. iDrugs 2001, 4, 1365–1367. [Google Scholar] [PubMed]
- Barr, J.L.; Unterwald, E.M. Glycogen synthase kinase-3 signaling in cellular and behavioral responses to psychostimulant drugs. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118746. [Google Scholar] [CrossRef]
- Jope, R.S.; Johnson, G.V. The glamour and gloom of glycogen synthase kinase-3. Trends Biochem. Sci. 2004, 29, 95–102. [Google Scholar] [CrossRef]
- Woodgett, J.R. Judging a protein by more than its name: GSK-3. Sci. STKE 2001, 2001, re12. [Google Scholar] [CrossRef]
- Wang, Q.M.; Fiol, C.J.; DePaoli-Roach, A.A.; Roach, P.J. Glycogen synthase kinase-3 beta is a dual specificity kinase differentially regulated by tyrosine and serine/threonine phosphorylation. J. Biol. Chem. 1994, 269, 14566–14574. [Google Scholar] [CrossRef]
- Hughes, K.; Nikolakaki, E.; Plyte, S.E.; Totty, N.F.; Woodgett, J.R. Modulation of the glycogen synthase kinase-3 family by tyrosine phosphorylation. EMBO J. 1993, 12, 803–808. [Google Scholar] [CrossRef]
- Hur, E.M.; Zhou, F.Q. GSK3 signalling in neural development. Nat. Rev. Neurosci. 2010, 11, 539–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beurel, E.; Grieco, S.F.; Jope, R.S. Glycogen synthase kinase-3 (GSK3): Regulation, actions, and diseases. Pharmacol. Ther. 2015, 148, 114–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.M.; Wang, J.; Wu, P.; Xue, Y.X.; Zhu, W.L.; Li, Q.Q.; Zhai, H.F.; Shi, J.; Lu, L. Glycogen synthase kinase 3β in the nucleus accumbens core is critical for methamphetamine-induced behavioral sensitization. J. Neurochem. 2011, 118, 126–139. [Google Scholar] [CrossRef]
- Duda, P.; Hajka, D.; Wójcicka, O.; Rakus, D.; Gizak, A. GSK3β: A Master Player in Depressive Disorder Pathogenesis and Treatment Responsiveness. Cells 2020, 9, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, M.; Lee, V.M. Insulin and insulin-like growth factor-1 regulate tau phosphorylation in cultured human neurons. J. Biol. Chem. 1997, 272, 19547–19553. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, J.M.; Sotnikova, T.D.; Yao, W.D.; Kockeritz, L.; Woodgett, J.R.; Gainetdinov, R.R.; Caron, M.G. Lithium antagonizes dopamine-dependent behaviors mediated by an AKT/glycogen synthase kinase 3 signaling cascade. Proc. Natl. Acad. Sci. USA 2004, 101, 5099–5104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, T.D.; Manji, H.K. Glycogen synthase kinase-3: A putative molecular target for lithium mimetic drugs. Neuropsychopharmacology 2005, 30, 1223–1237. [Google Scholar] [CrossRef] [Green Version]
- Koros, E.; Dorner-Ciossek, C. The role of glycogen synthase kinase-3beta in schizophrenia. Drug News Perspect. 2007, 20, 437–445. [Google Scholar] [CrossRef]
- Lei, G.; Xia, Y.; Johnson, K.M. The role of Akt-GSK-3beta signaling and synaptic strength in phencyclidine-induced neurodegeneration. Neuropsychopharmacology 2008, 33, 1343–1353. [Google Scholar] [CrossRef] [Green Version]
- Solís, O.; García-Sanz, P.; Martín, A.B.; Granado, N.; Sanz-Magro, A.; Podlesniy, P.; Trullas, R.; Murer, M.G.; Maldonado, R.; Moratalla, R. Behavioral sensitization and cellular responses to psychostimulants are reduced in D2R knockout mice. Addict. Biol. 2021, 26, e12840. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.M.; Pierce, R.C. Cocaine-induced alterations in dopamine receptor signaling: Implications for reinforcement and reinstatement. Pharmacol. Ther. 2005, 106, 389–403. [Google Scholar] [CrossRef] [PubMed]
- Cervo, L.; Samanin, R. Effects of dopaminergic and glutamatergic receptor antagonists on the acquisition and expression of cocaine conditioning place preference. Brain Res. 1995, 673, 242–250. [Google Scholar] [CrossRef]
- Kim, H.S.; Park, W.K.; Jang, C.G.; Oh, S. Inhibition by MK-801 of cocaine-induced sensitization, conditioned place preference, and dopamine-receptor supersensitivity in mice. Brain Res. Bull. 1996, 40, 201–207. [Google Scholar] [CrossRef]
- Luo, H.R.; Hattori, H.; Hossain, M.A.; Hester, L.; Huang, Y.; Lee-Kwon, W.; Donowitz, M.; Nagata, E.; Snyder, S.H. Akt as a mediator of cell death. Proc. Natl. Acad. Sci. USA 2003, 100, 11712–11717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linke, S.E.; Ussher, M. Exercise-based treatments for substance use disorders: Evidence, theory, and practicality. Am. J. Drug Alcohol Abus. 2015, 41, 7–15. [Google Scholar] [CrossRef]
- Kim, S.; Jang, E.Y.; Song, S.H.; Kim, J.S.; Ryu, I.S.; Jeong, C.H.; Lee, S. Brain Microdialysis Coupled to LC-MS/MS Revealed That CVT-10216, a Selective Inhibitor of Aldehyde Dehydrogenase 2, Alters the Neurochemical and Behavioral Effects of Methamphetamine. ACS Chem. Neurosci. 2021, 12, 1552–1562. [Google Scholar] [CrossRef]
- Gu, S.M.; Kim, S.Y.; Lamichhane, S.; Hong, J.T.; Yun, J. Limonene Inhibits Methamphetamine-Induced Sensitizations via the Regulation of Dopamine Receptor Supersensitivity. Biomol. Ther. 2019, 27, 357–362. [Google Scholar] [CrossRef]
- Wang, D.; Zhou, C.; Zhao, M.; Wu, X.; Chang, Y.K. Dose-response relationships between exercise intensity, cravings, and inhibitory control in methamphetamine dependence: An ERPs study. Drug Alcohol Depend. 2016, 161, 331–339. [Google Scholar] [CrossRef]
- Kemi, O.J.; Loennechen, J.P.; Wisløff, U.; Ellingsen, Ø. Intensity-controlled treadmill running in mice: Cardiac and skeletal muscle hypertrophy. J. Appl. Physiol. 2002, 93, 1301–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, M.J.; Mulia, G.E.; van Rijn, R.M. Commonly Used Anesthesia/Euthanasia Methods for Brain Collection Differentially Impact MAPK Activity in Male and Female C57BL/6 Mice. Front. Cell Neurosci. 2019, 13, 96. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.E.; Lang, Q.H.; Yu, Q.Y.; Tang, X.Y.; Liu, Q.Q.; Li, X.Y.; Feng, X.Z. Behavioral Assessments of Spontaneous Locomotion in a Murine MPTP-induced Parkinson’s Disease Model. J. Vis. Exp. 2019, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brudzynski, S.M.; Krol, S. Analysis of locomotor activity in the rat: Parallelism index, a new measure of locomotor exploratory pattern. Physiol. Behav. 1997, 62, 635–642. [Google Scholar] [CrossRef]
- Crombag, H.S.; Bossert, J.M.; Koya, E.; Shaham, Y. Context-induced relapse to drug seeking: A review. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 3233–3243. [Google Scholar] [CrossRef]
- Koob, G.F. Stress, corticotropin-releasing factor, and drug addiction. Ann. N. Y. Acad. Sci. 1999, 897, 27–45. [Google Scholar] [CrossRef]
- Knackstedt, L.A.; Kalivas, P.W. Glutamate and reinstatement. Curr. Opin. Pharmacol. 2009, 9, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karler, R.; Calder, L.D.; Thai, L.H.; Bedingfield, J.B. A dopaminergic-glutamatergic basis for the action of amphetamine and cocaine. Brain Res. 1994, 658, 8–14. [Google Scholar] [CrossRef]
- McCreary, A.C.; Marsden, C.A. Cocaine-induced behaviour: Dopamine D1 receptor antagonism by SCH 23390 prevents expression of conditioned sensitisation following repeated administration of cocaine. Neuropharmacology 1993, 32, 387–391. [Google Scholar] [CrossRef]
- Choe, E.S.; Chung, K.T.; Mao, L.; Wang, J.Q. Amphetamine increases phosphorylation of extracellular signal-regulated kinase and transcription factors in the rat striatum via group I metabotropic glutamate receptors. Neuropsychopharmacology 2002, 27, 565–575. [Google Scholar] [CrossRef] [Green Version]
- Lett, B.T.; Grant, V.L.; Koh, M.T.; Flynn, G. Prior experience with wheel running produces cross-tolerance to the rewarding effect of morphine. Pharmacol. Biochem. Behav. 2002, 72, 101–105. [Google Scholar] [CrossRef]
- Smith, M.A.; Gergans, S.R.; Iordanou, J.C.; Lyle, M.A. Chronic exercise increases sensitivity to the conditioned rewarding effects of cocaine. Pharmacol. Rep. 2008, 60, 561–565. [Google Scholar]
- Epling, W.F.; Pierce, W.D. Solving the Anorexia Puzzle: A Scientific Approach; Hogrefe & Huber Pub.: Toronto, ON, Canada, 1992. [Google Scholar]
- Egorov, A.Y.; Szabo, A. The exercise paradox: An interactional model for a clearer conceptualization of exercise addiction. J. Behav. Addict. 2013, 2, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolb, E.M.; Kelly, S.A.; Garland, T., Jr. Mice from lines selectively bred for high voluntary wheel running exhibit lower blood pressure during withdrawal from wheel access. Physiol. Behav. 2013, 112–113, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Perrine, S.A.; Sheikh, I.S.; Nwaneshiudu, C.A.; Schroeder, J.A.; Unterwald, E.M. Withdrawal from chronic administration of cocaine decreases delta opioid receptor signaling and increases anxiety- and depression-like behaviors in the rat. Neuropharmacology 2008, 54, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Anraku, T.; Ikegaya, Y.; Matsuki, N.; Nishiyama, N. Withdrawal from chronic morphine administration causes prolonged enhancement of immobility in rat forced swimming test. Psychopharmacology 2001, 157, 217–220. [Google Scholar] [CrossRef]
- Malisch, J.L.; Breuner, C.W.; Kolb, E.M.; Wada, H.; Hannon, R.M.; Chappell, M.A.; Middleton, K.M.; Garland, T., Jr. Behavioral despair and home-cage activity in mice with chronically elevated baseline corticosterone concentrations. Behav. Genet. 2009, 39, 192–201. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-5, 5th ed.; American Psychiatric Association: Washingotn, DC, USA, 2013; p. 947. [Google Scholar]
- Mark, K.A.; Soghomonian, J.J.; Yamamoto, B.K. High-dose methamphetamine acutely activates the striatonigral pathway to increase striatal glutamate and mediate long-term dopamine toxicity. J. Neurosci. 2004, 24, 11449–11456. [Google Scholar] [CrossRef] [Green Version]
- Nash, J.F.; Yamamoto, B.K. Methamphetamine neurotoxicity and striatal glutamate release: Comparison to 3,4-methylenedioxymethamphetamine. Brain Res. 1992, 581, 237–243. [Google Scholar] [CrossRef]
- Krasnova, I.N.; Cadet, J.L. Methamphetamine toxicity and messengers of death. Brain Res. Rev. 2009, 60, 379–407. [Google Scholar] [CrossRef] [Green Version]
- Randall, R.D.; Thayer, S.A. Glutamate-induced calcium transient triggers delayed calcium overload and neurotoxicity in rat hippocampal neurons. J. Neurosci. 1992, 12, 1882–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szatmari, E.; Habas, A.; Yang, P.; Zheng, J.J.; Hagg, T.; Hetman, M. A positive feedback loop between glycogen synthase kinase 3beta and protein phosphatase 1 after stimulation of NR2B NMDA receptors in forebrain neurons. J. Biol. Chem. 2005, 280, 37526–37535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulkey, R.M.; Endo, S.; Shenolikar, S.; Malenka, R.C. Involvement of a calcineurin/inhibitor-1 phosphatase cascade in hippocampal long-term depression. Nature 1994, 369, 486–488. [Google Scholar] [CrossRef]
- Peineau, S.; Taghibiglou, C.; Bradley, C.; Wong, T.P.; Liu, L.; Lu, J.; Lo, E.; Wu, D.; Saule, E.; Bouschet, T.; et al. LTP inhibits LTD in the hippocampus via regulation of GSK3beta. Neuron 2007, 53, 703–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, F.; Langa, E.; Cuadros, R.; Avila, J.; Villanueva, N. Regulation of GSK3 isoforms by phosphatases PP1 and PP2A. Mol. Cell Biochem. 2010, 344, 211–215. [Google Scholar] [CrossRef]
- Nagini, S.; Sophia, J.; Mishra, R. Glycogen synthase kinases: Moonlighting proteins with theranostic potential in cancer. Semin. Cancer Biol. 2019, 56, 25–36. [Google Scholar] [CrossRef]
- El Rawas, R.; Amaral, I.M.; Hofer, A. Is p38 MAPK Associated to Drugs of Abuse-Induced Abnormal Behaviors? Int. J. Mol. Sci. 2020, 21, 4833. [Google Scholar] [CrossRef]
- Valjent, E.; Corvol, J.C.; Pages, C.; Besson, M.J.; Maldonado, R.; Caboche, J. Involvement of the extracellular signal-regulated kinase cascade for cocaine-rewarding properties. J. Neurosci. 2000, 20, 8701–8709. [Google Scholar] [CrossRef]
- Choe, E.S.; Wang, J.Q. CaMKII regulates amphetamine-induced ERK1/2 phosphorylation in striatal neurons. Neuroreport 2002, 13, 1013–1016. [Google Scholar] [CrossRef]
- Berhow, M.T.; Hiroi, N.; Nestler, E.J. Regulation of ERK (extracellular signal regulated kinase), part of the neurotrophin signal transduction cascade, in the rat mesolimbic dopamine system by chronic exposure to morphine or cocaine. J. Neurosci. 1996, 16, 4707–4715. [Google Scholar] [CrossRef] [Green Version]
- Freeman, W.M.; Nader, M.A.; Nader, S.H.; Robertson, D.J.; Gioia, L.; Mitchell, S.M.; Daunais, J.B.; Porrino, L.J.; Friedman, D.P.; Vrana, K.E. Chronic cocaine-mediated changes in non-human primate nucleus accumbens gene expression. J. Neurochem. 2001, 77, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Gerdjikov, T.V.; Ross, G.M.; Beninger, R.J. Place preference induced by nucleus accumbens amphetamine is impaired by antagonists of ERK or p38 MAP kinases in rats. Behav. Neurosci. 2004, 118, 740–750. [Google Scholar] [CrossRef] [Green Version]
- Pierce, R.C.; Pierce-Bancroft, A.F.; Prasad, B.M. Neurotrophin-3 contributes to the initiation of behavioral sensitization to cocaine by activating the Ras/Mitogen-activated protein kinase signal transduction cascade. J. Neurosci. 1999, 19, 8685–8695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkadhi, K.A. Exercise as a Positive Modulator of Brain Function. Mol. Neurobiol. 2018, 55, 3112–3130. [Google Scholar] [CrossRef] [PubMed]
- Damghani, F.; Bigdeli, I.; Miladi-Gorji, H.; Fadaei, A. Swimming exercise attenuates psychological dependence and voluntary methamphetamine consumption in methamphetamine withdrawn rats. Iran. J. Basic Med. Sci. 2016, 19, 594–600. [Google Scholar]
- Fontes-Ribeiro, C.A.; Marques, E.; Pereira, F.C.; Silva, A.P.; Macedo, T.R. May exercise prevent addiction? Curr. Neuropharmacol. 2011, 9, 45–48. [Google Scholar] [CrossRef] [Green Version]
- Greenwood, B.N.; Spence, K.G.; Crevling, D.M.; Clark, P.J.; Craig, W.C.; Fleshner, M. Exercise-induced stress resistance is independent of exercise controllability and the medial prefrontal cortex. Eur. J. Neurosci. 2013, 37, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.M.; Dunsiger, S.; Whiteley, J.A.; Ussher, M.H.; Ciccolo, J.T.; Jennings, E.G. Acute effects of moderate intensity aerobic exercise on affective withdrawal symptoms and cravings among women smokers. Addict. Behav. 2011, 36, 894–897. [Google Scholar] [CrossRef] [Green Version]
- Abrantes, A.M.; Battle, C.L.; Strong, D.R.; Ing, E.; Dubreuil, M.E.; Gordon, A.; Brown, R.A. Exercise preferences of patients in substance abuse treatment. Ment. Health Phys. Act. 2011, 4, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.M.; Whiteley, J.A.; Dunsiger, S.; Jennings, E.G.; Albrecht, A.E.; Ussher, M.H.; Ciccolo, J.T.; Parisi, A.F.; Marcus, B.H. Moderate intensity exercise as an adjunct to standard smoking cessation treatment for women: A pilot study. Psychol. Addict. Behav. 2010, 24, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Aaron, D.J.; Dearwater, S.R.; Anderson, R.; Olsen, T.; Kriska, A.M.; Laporte, R.E. Physical activity and the initiation of high-risk health behaviors in adolescents. Med. Sci. Sports Exerc. 1995, 27, 1639–1645. [Google Scholar] [CrossRef]
- Pate, R.R.; Trost, S.G.; Levin, S.; Dowda, M. Sports participation and health-related behaviors among US youth. Arch. Pediatr. Adolesc. Med. 2000, 154, 904–911. [Google Scholar] [CrossRef] [PubMed]
- Kulig, K.; Brener, N.D.; McManus, T. Sexual activity and substance use among adolescents by category of physical activity plus team sports participation. Arch. Pediatr. Adolesc. Med. 2003, 157, 905–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, M.J.; Werch, C.E. Sport and physical activity participation and substance use among adolescents. J. Adolesc. Health 2005, 36, 486–493. [Google Scholar] [CrossRef]
- Ströhle, A.; Höfler, M.; Pfister, H.; Müller, A.G.; Hoyer, J.; Wittchen, H.U.; Lieb, R. Physical activity and prevalence and incidence of mental disorders in adolescents and young adults. Psychol. Med. 2007, 37, 1657–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korhonen, T.; Kujala, U.M.; Rose, R.J.; Kaprio, J. Physical activity in adolescence as a predictor of alcohol and illicit drug use in early adulthood: A longitudinal population-based twin study. Twin Res. Hum. Genet. 2009, 12, 261–268. [Google Scholar] [CrossRef]
- Ehringer, M.A.; Hoft, N.R.; Zunhammer, M. Reduced alcohol consumption in mice with access to a running wheel. Alcohol 2009, 43, 443–452. [Google Scholar] [CrossRef]
- Cosgrove, K.P.; Hunter, R.G.; Carroll, M.E. Wheel-running attenuates intravenous cocaine self-administration in rats: Sex differences. Pharmacol. Biochem. Behav. 2002, 73, 663–671. [Google Scholar] [CrossRef]
- Brocardo, P.S.; Boehme, F.; Patten, A.; Cox, A.; Gil-Mohapel, J.; Christie, B.R. Anxiety- and depression-like behaviors are accompanied by an increase in oxidative stress in a rat model of fetal alcohol spectrum disorders: Protective effects of voluntary physical exercise. Neuropharmacology 2012, 62, 1607–1618. [Google Scholar] [CrossRef]
- Peterson, A.B.; Hivick, D.P.; Lynch, W.J. Dose-dependent effectiveness of wheel running to attenuate cocaine-seeking: Impact of sex and estrous cycle in rats. Psychopharmacology 2014, 231, 2661–2670. [Google Scholar] [CrossRef] [PubMed]
- Zlebnik, N.E.; Saykao, A.T.; Carroll, M.E. Effects of combined exercise and progesterone treatments on cocaine seeking in male and female rats. Psychopharmacology 2014, 231, 3787–3798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thanos, P.K.; Tucci, A.; Stamos, J.; Robison, L.; Wang, G.J.; Anderson, B.J.; Volkow, N.D. Chronic forced exercise during adolescence decreases cocaine conditioned place preference in Lewis rats. Behav. Brain. Res. 2010, 215, 77–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.A.; Pitts, E.G. Access to a running wheel inhibits the acquisition of cocaine self-administration. Pharmacol. Biochem. Behav. 2011, 100, 237–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.A.; Pitts, E.G. Wheel running decreases the positive reinforcing effects of heroin. Pharmacol. Rep. 2012, 64, 960–964. [Google Scholar] [CrossRef] [Green Version]
- Berchtold, N.C.; Kesslak, J.P.; Pike, C.J.; Adlard, P.A.; Cotman, C.W. Estrogen and exercise interact to regulate brain-derived neurotrophic factor mRNA and protein expression in the hippocampus. Eur. J. Neurosci. 2001, 14, 1992–2002. [Google Scholar] [CrossRef] [PubMed]
- Lynch, W.J.; Roth, M.E.; Carroll, M.E. Biological basis of sex differences in drug abuse: Preclinical and clinical studies. Psychopharmacology 2002, 164, 121–137. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.B.; Hu, M. Sex differences in drug abuse. Front. Neuroendocr. 2008, 29, 36–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, S.; Kim, Y.; Kim, M.; Seo, M.; Kim, S.; Kim, S.; Lee, S. Exercise Pills for Drug Addiction: Forced Moderate Endurance Exercise Inhibits Methamphetamine-Induced Hyperactivity through the Striatal Glutamatergic Signaling Pathway in Male Sprague Dawley Rats. Int. J. Mol. Sci. 2021, 22, 8203. https://doi.org/10.3390/ijms22158203
Jung S, Kim Y, Kim M, Seo M, Kim S, Kim S, Lee S. Exercise Pills for Drug Addiction: Forced Moderate Endurance Exercise Inhibits Methamphetamine-Induced Hyperactivity through the Striatal Glutamatergic Signaling Pathway in Male Sprague Dawley Rats. International Journal of Molecular Sciences. 2021; 22(15):8203. https://doi.org/10.3390/ijms22158203
Chicago/Turabian StyleJung, Suryun, Youjeong Kim, Mingyu Kim, Minjae Seo, Suji Kim, Seungju Kim, and Sooyeun Lee. 2021. "Exercise Pills for Drug Addiction: Forced Moderate Endurance Exercise Inhibits Methamphetamine-Induced Hyperactivity through the Striatal Glutamatergic Signaling Pathway in Male Sprague Dawley Rats" International Journal of Molecular Sciences 22, no. 15: 8203. https://doi.org/10.3390/ijms22158203
APA StyleJung, S., Kim, Y., Kim, M., Seo, M., Kim, S., Kim, S., & Lee, S. (2021). Exercise Pills for Drug Addiction: Forced Moderate Endurance Exercise Inhibits Methamphetamine-Induced Hyperactivity through the Striatal Glutamatergic Signaling Pathway in Male Sprague Dawley Rats. International Journal of Molecular Sciences, 22(15), 8203. https://doi.org/10.3390/ijms22158203