
Integrin αv and Vitronectin Prime Macrophage-Related Inflammation and Contribute the Development of Dry Eye Disease

Abstract
:1. Introduction
2. Results
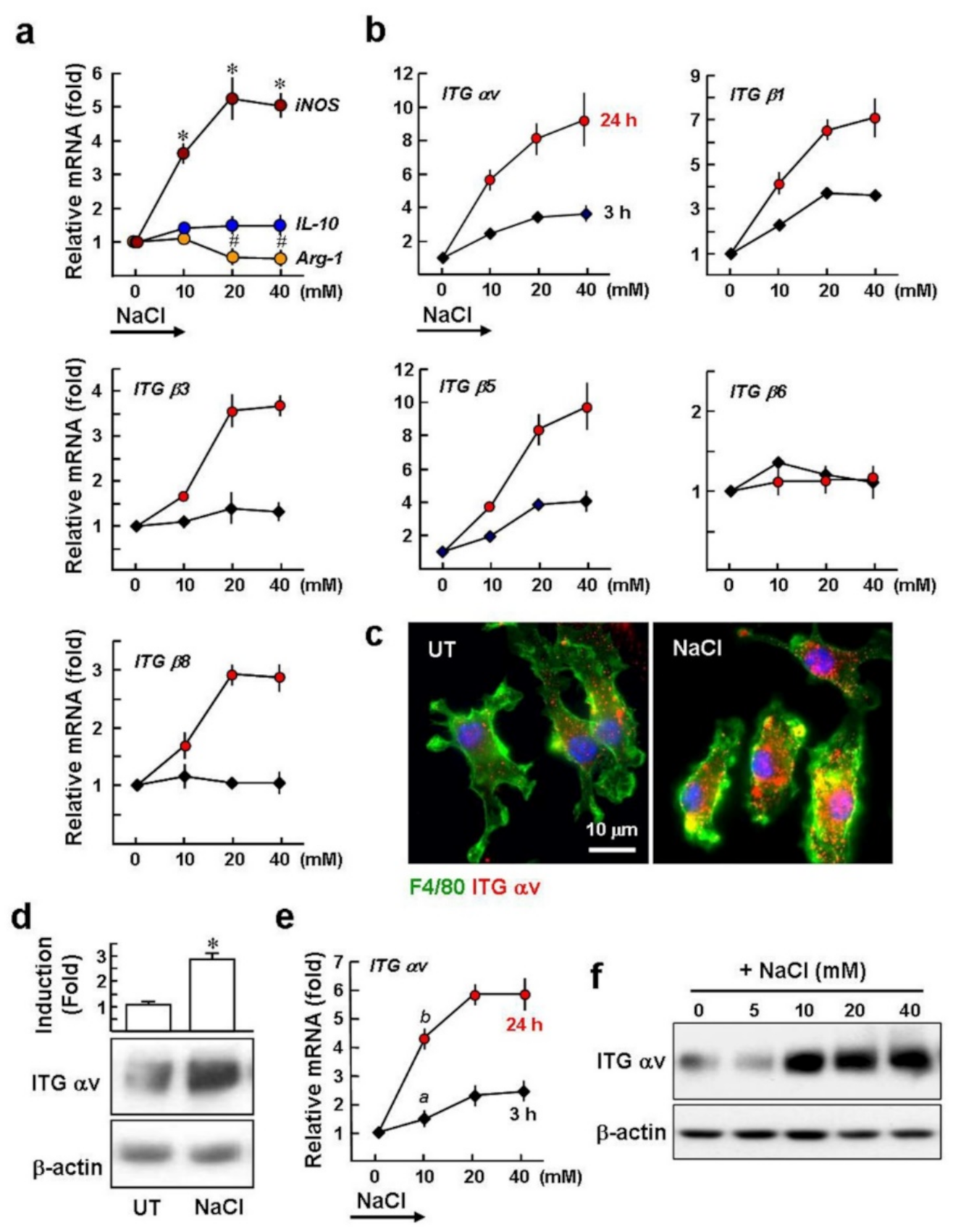
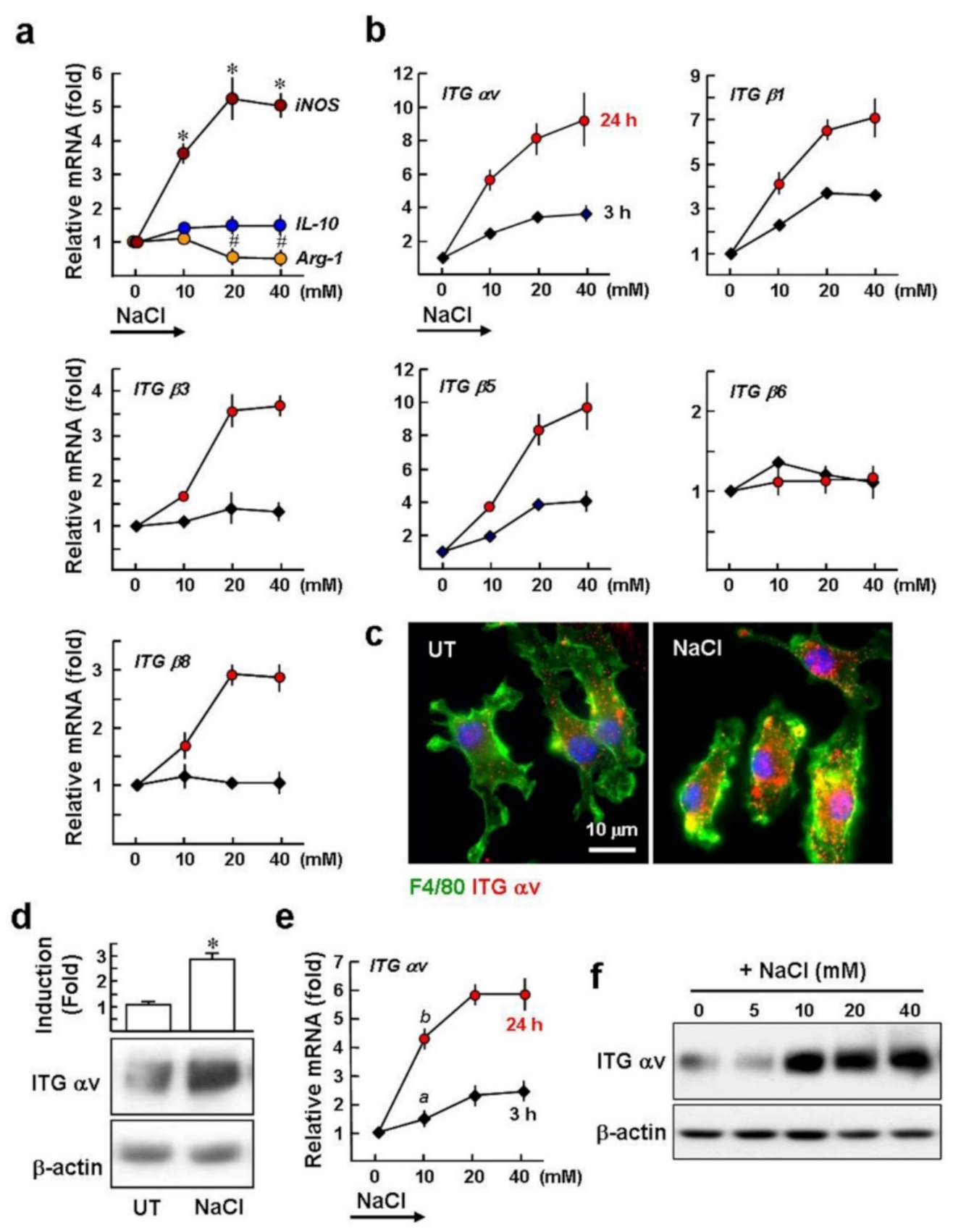
2.1. High Salt Upregulates the Gene Expression of αv Integrins in Macrophages
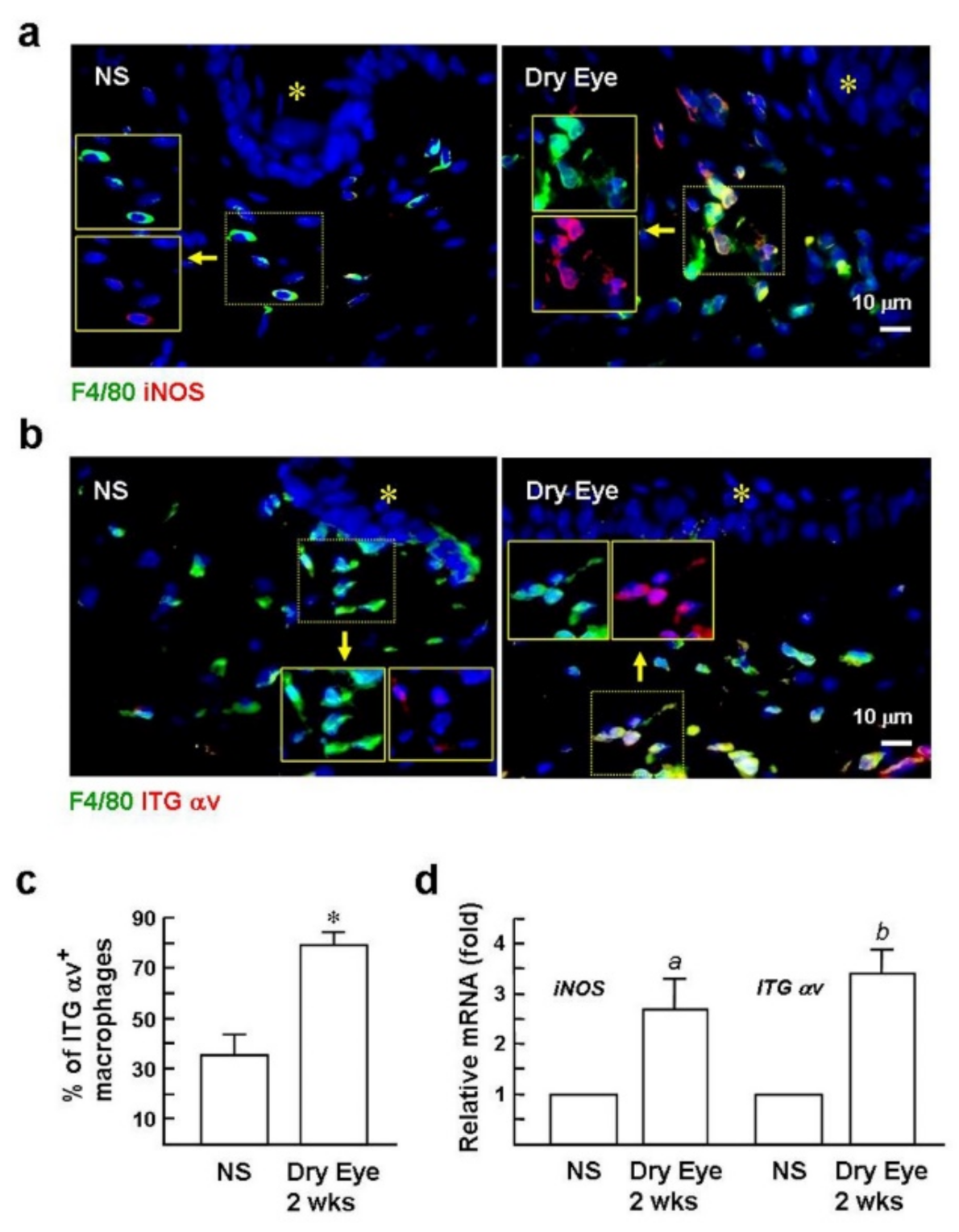
2.2. Expression of the αv Integrin in Macrophages Is Induced in Dry Eyes
2.3. VTN Is a Proinflammatory Stimulator of BMDMs
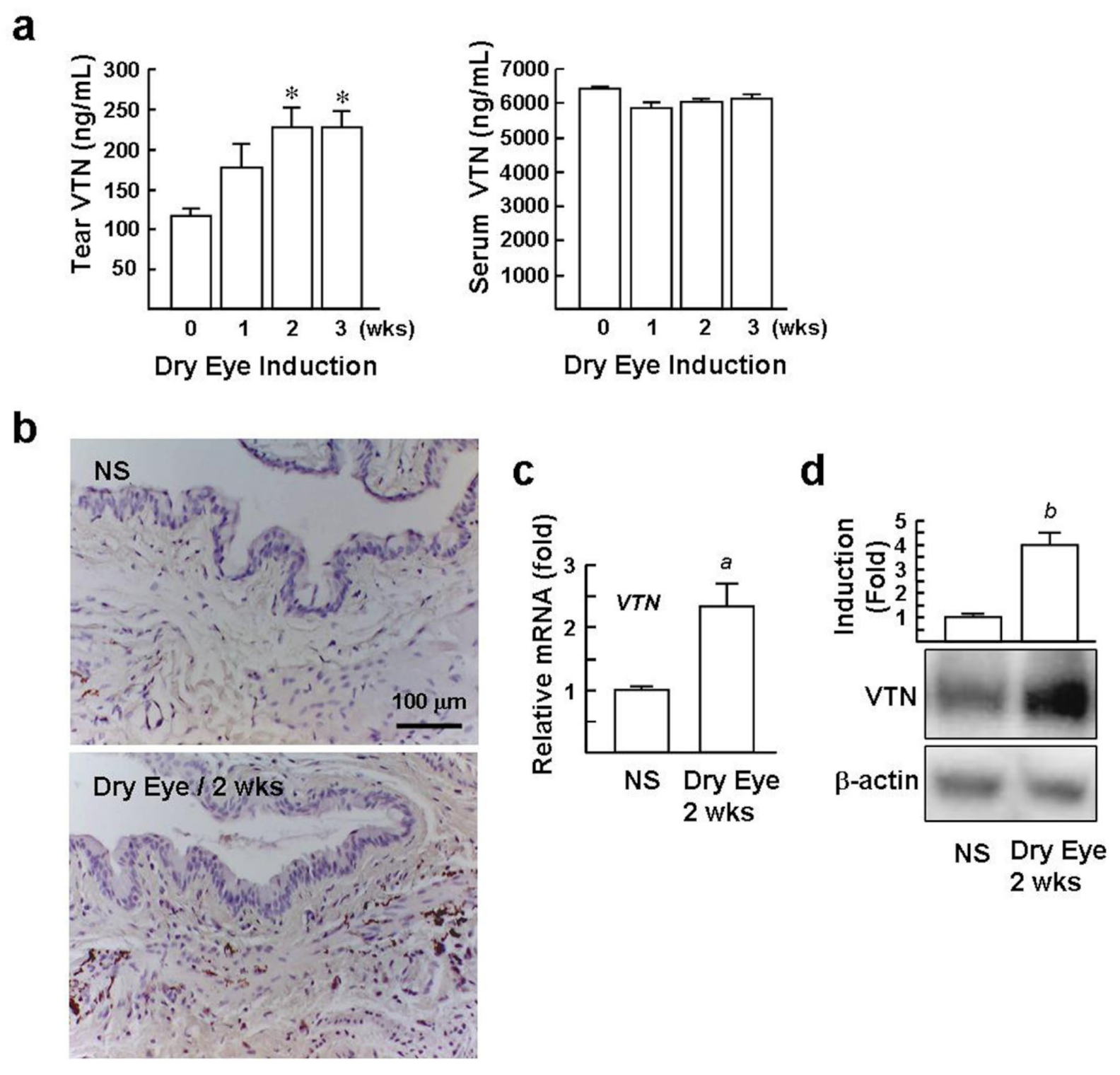
2.4. Levels of VTN in the Tear and Conjunctival StromaareElevated in DED Mice
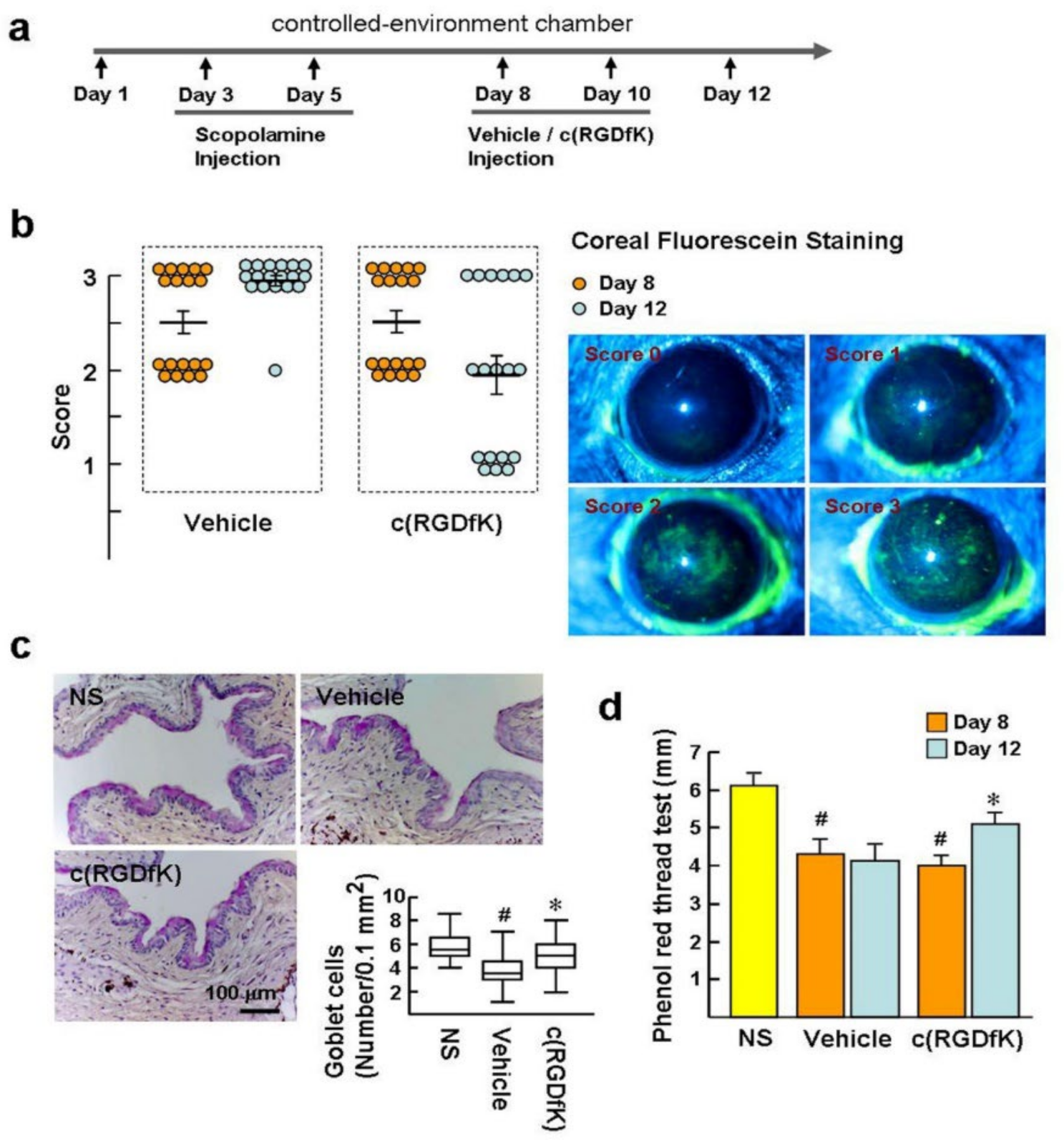
2.5. c(RGDfK) Benefitsthe Recovery of Experimental Dry Eye
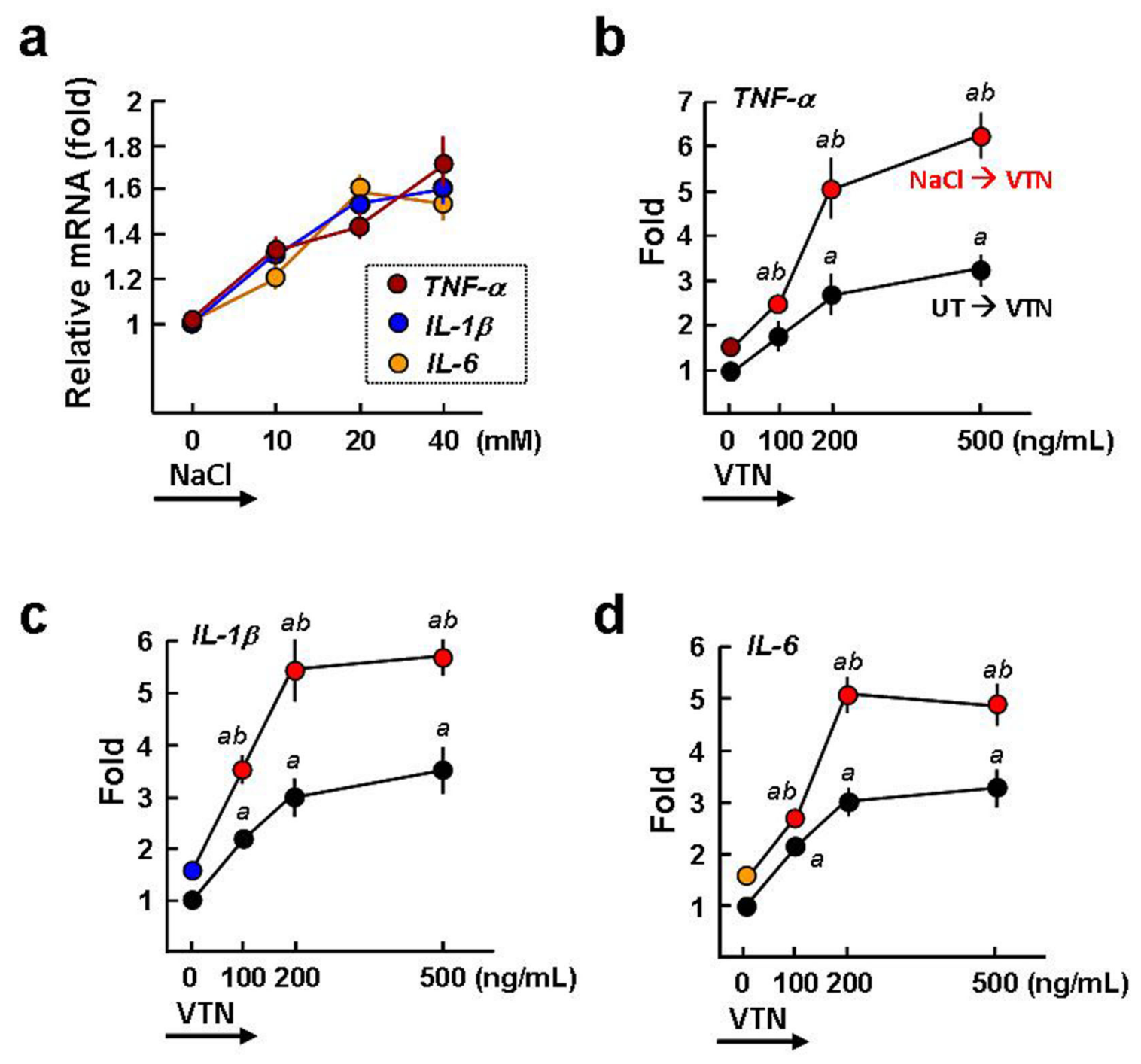
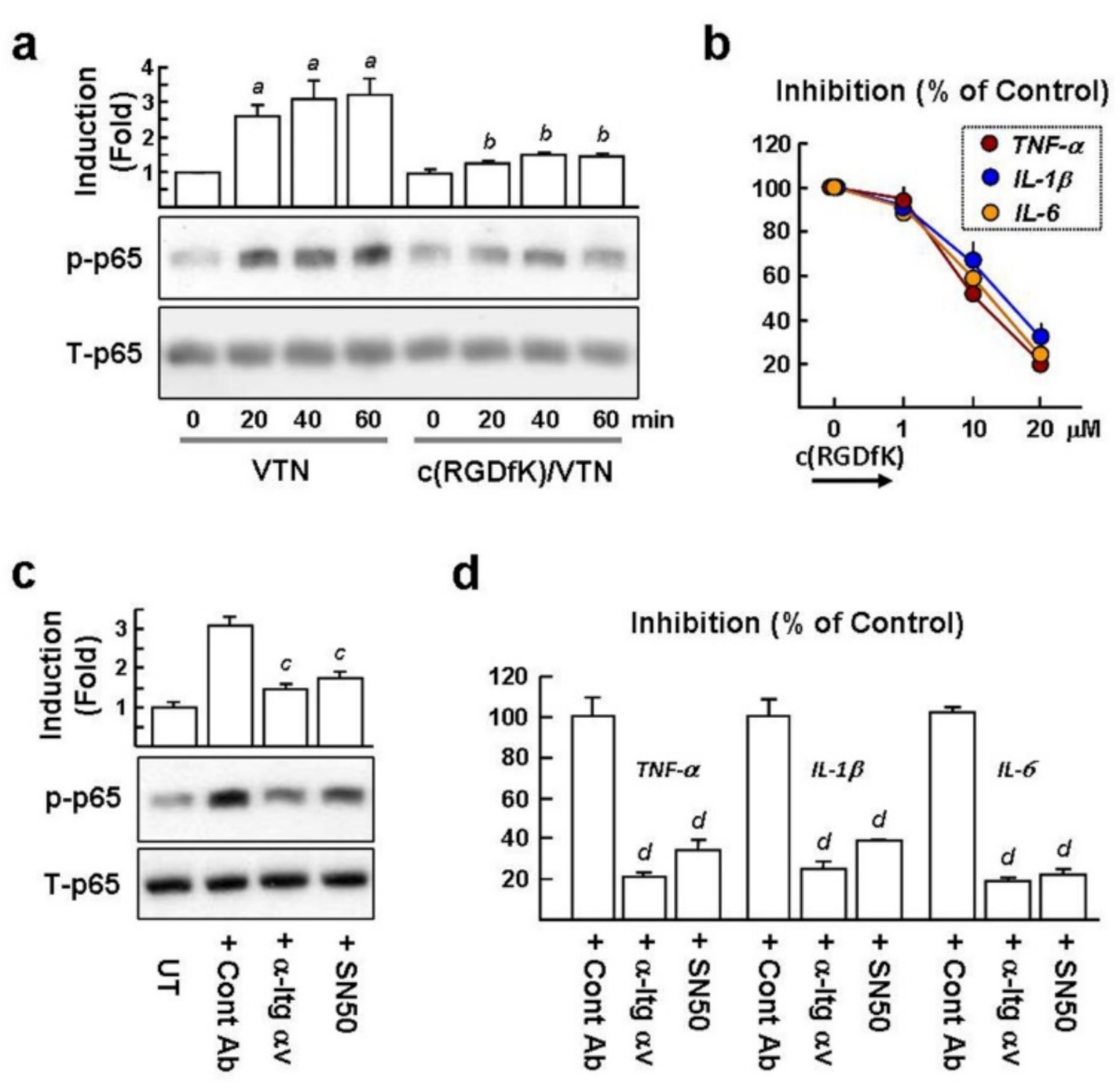
2.6. VTN Interacts with αv Integrins to Induce NF-κB-Mediated Proinflammatory Cytokine GeneExpression in NaCl-Treated Macrophages
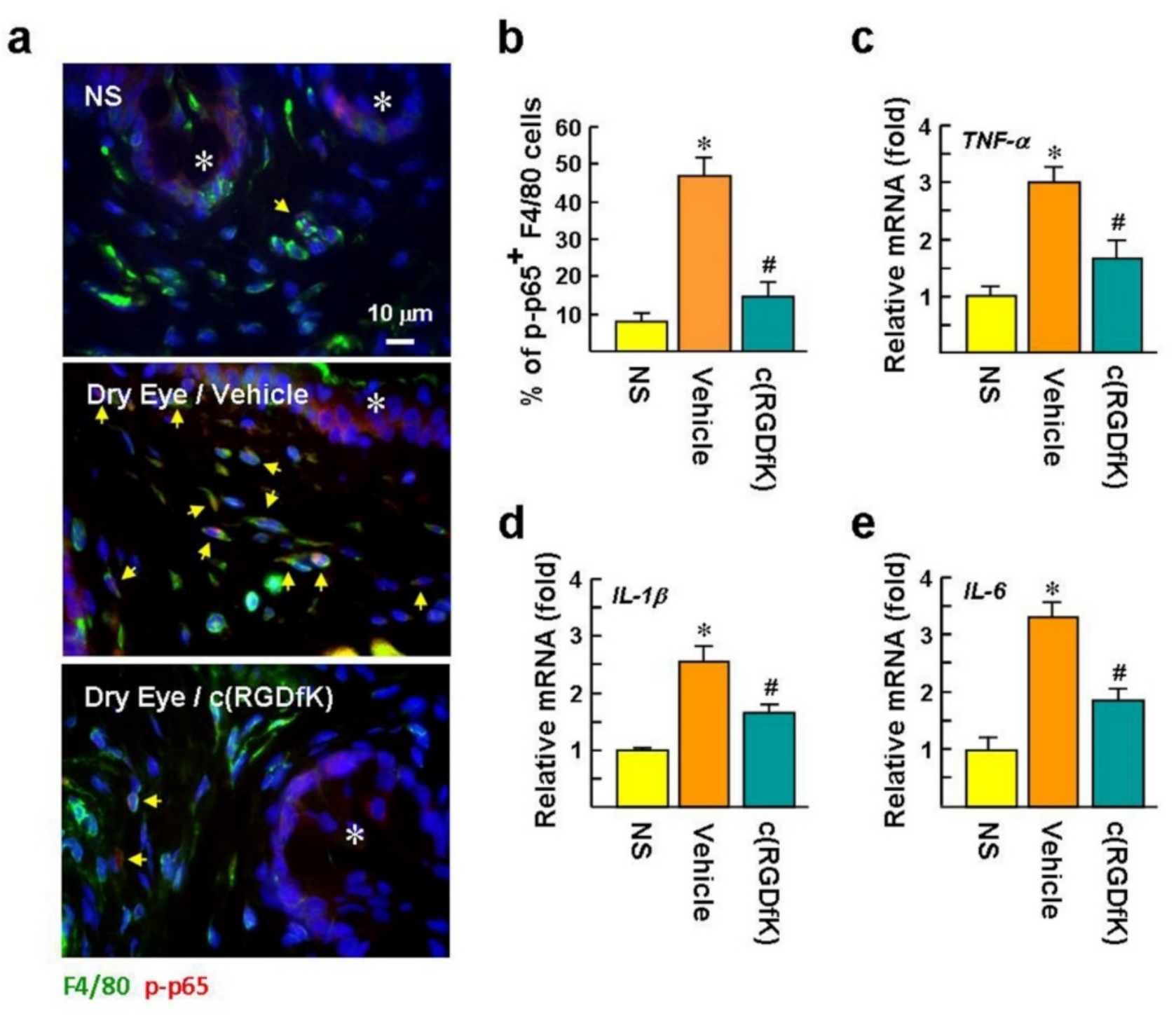
2.7. c(RGDfK) Suppresses NF-κB Activation in Macrophages and Proinflammatory Cytokine Expression in the Dry Eye
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and NaCl Treatment
4.3. Induction of DED
4.4. Clinical Examination of DED
4.5. PAS Staining of Goblet Cells
4.6. Quantitative Real-Time PCR
4.7. Western Blot Analysis
4.8. Immunohistochemistry
4.9. Immunofluorescence
4.10. Mouse VTN ELISA
4.11. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Listed, N. The definition and classification of dry eye disease: Report of the Definition and Classification Subcommittee of the International Dry Eye Workshop. Ocul. Surf. 2007, 5, 75–92. [Google Scholar]
- Zhang, X.; M., V.J.; Qu, Y.; He, X.; Ou, S.; Bu, J.; Jia, C.; Wang, J.; Wu, H.; Liu, Z.; et al. Dry Eye Management: Targeting the Ocular Surface Microenvironment. Int. J. Mol. Sci. 2017, 18, 1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 1801. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.C.; Zheng, X.J.; Du, L.J.; Sun, J.Y.; Shen, Z.X.; Shi, C.; Sun, S.; Zhang, Z.; Chen, X.Q.; Qin, M.; et al. High salt primes a specific activation state of macrophages, M(Na). Cell Res. 2015, 25, 893–910. [Google Scholar] [CrossRef]
- Hucke, S.; Eschborn, M.; Liebmann, M.; Herold, M.; Freise, N.; Engbers, A.; Ehling, P.; Meuth, S.G.; Roth, J.; Kuhlmann, T.; et al. Sodium chloride promotes pro-inflammatory macrophage polarization thereby aggravating CNS autoimmunity. J. Autoimmun. 2016, 67, 90–101. [Google Scholar] [CrossRef] [PubMed]
- You, I.C.; Coursey, T.G.; Bian, F.; Barbosa, F.L.; de Paiva, C.S.; Pflugfelder, S.C. Macrophage Phenotype in the Ocular Surface of Experimental Murine Dry Eye Disease. Arch. Immunol. Ther. Exp. 2015, 63, 299–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pflugfelder, S.C.; Bian, F.; Gumus, K.; Farley, W.; Stern, M.E.; De Paiva, C.S. Severity of Sjögren’s Syndrome Keratoconjunctivitis Sicca Increases with Increased Percentage of Conjunctival Antigen-Presenting Cells. Int. J. Mol. Sci. 2018, 19, 2760. [Google Scholar] [CrossRef] [Green Version]
- Alam, J.; de Paiva, C.S.; Pflugfelder, S.C. Desiccation Induced Conjunctival Monocyte Recruitment and Activation-Implications for Keratoconjunctivitis. Front. Immunol. 2021, 12, 701415. [Google Scholar] [CrossRef] [PubMed]
- Schaumburg, C.S.; Siemasko, K.F.; De Paiva, C.S.; Wheeler, L.A.; Niederkorn, J.Y.; Pflugfelder, S.C.; Stern, M.E. Ocular surface APCs are necessary for autoreactive T cell-mediated experimental autoimmune lacrimal keratoconjunctivitis. J. Immunol. 2011, 187, 3653–3662. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Chen, Y.T.; Chen, F.; Gallup, M.; Vijmasi, T.; Bahrami, A.F.; Noble, L.B.; van Rooijen, N.; McNamara, N.A. Critical involvement of macrophage infiltration in the development of Sjögren’s syndrome-associated dry eye. Am. J. Pathol. 2012, 181, 753–760. [Google Scholar] [CrossRef] [Green Version]
- Felding-Habermann, B.; Cheresh, D.A. Vitronectin and its receptors. Curr. Opin. Cell Biol. 1993, 5, 864–868. [Google Scholar] [CrossRef]
- Sheppard, D. Modulation of acute lung injury by integrins. Proc. Am. Thorac. Soc. 2012, 9, 126–129. [Google Scholar] [CrossRef]
- Laitinen, I.; Saraste, A.; Weidl, E.; Poethko, T.; Weber, A.W.; Nekolla, S.G.; Leppänen, P.; Ylä-Herttuala, S.; Hölzlwimmer., G.; Walch, A.; et al. Evaluation of alphavbeta3integrin-targeted positron emission tomography tracer 18F-galacto-RGD for imaging of vascular inflammation in atherosclerotic mice. Circ. Cardiovasc. Imaging 2009, 2, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Henderson, N.C.; Arnold, T.D.; Katamura, Y.; Giacomini, M.M.; Rodriguez, J.D.; McCarty, J.H.; Pellicoro, A.; Raschperger, E.; Betsholtz, C.; Ruminski, P.G.; et al. Targeting of αv integrin identifies a core molecular pathway that regulates fibrosis in several organs. Nat. Med. 2013, 19, 1617–1624. [Google Scholar] [CrossRef] [Green Version]
- Welser-Alves, J.V.; Boroujerdi, A.; Tigges, U.; Milner, R. Microglia use multiple mechanisms to mediate interactions with vitronectin; non-essential roles for the highly-expressed αvβ3 and αvβ5 integrins. J. Neuroinflamm. 2011, 8, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cursiefen, C.; Cordeiro, F.; Cunha-Vaz, J.; Wheeler-Schilling, T.; Scholl, H.P.N.; EVI Steering Board. Unmet Needs in Ophthalmology: A European Vision Institute-Consensus Roadmap 2019–2025. Ophthalmic. Res. 2019, 62, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Seiffert, D.; Keeton, M.; Eguchi, Y.; Sawdey, M.; Loskutoff, D.J. Detection of vitronectin mRNA in tissues and cells of the mouse. Proc. Natl. Acad. Sci. USA 1991, 88, 9402–9406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackwell, T.S.; Christman, J.W. The role of nuclear factor-kappa B in cytokine gene regulation. Am. J. Respir. Cell Mol. Biol. 1997, 17, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lu, Q.; Bottero, V.; Estepa, G.; Morrison, L.; Mercurio, F.; Verma, I.M. Enhanced NF-kappaB activation and cellular function in macrophages lacking IkappaB kinase 1 (IKK1). Proc. Natl. Acad. Sci. USA 2005, 102, 12425–12430. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.W.; Byun, Y.J.; Choi, W.; Jeong, E.; Kim, J.S.; Noh, H.; Kim, E.S.; Song, Y.J.; Park, S.K.; Lee, H.K. Neutralization of ocular surface TNF-α reduces ocular surface and lacrimal gland inflammationinduced by in vivo dry eye. Invest. Ophthalmol. Vis. Sci. 2013, 54, 7557–7566. [Google Scholar] [CrossRef] [Green Version]
- Okanobo, A.; Chauhan, S.K.; Dastjerdi, M.H.; Kodati, S.; Dana, R. Efficacy of topical blockade of interleukin-1 in experimental dry eye disease. Am. J. Ophthalmol. 2012, 154, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Antonov, A.S.; Kolodgie, F.D.; Munn, D.H.; Gerrity, R.G. Regulation of macrophage foam cell formation by alphaVbeta3 integrin: Potential role in human atherosclerosis. Am. J. Pathol. 2004, 165, 247–258. [Google Scholar] [CrossRef]
- Cowden Dahl, K.D.; Robertson, S.E.; Weaver, V.M.; Simon, M.C. Hypoxia-inducible factor regulates alphavbeta3 integrin cell surface expression. Mol. Biol. Cell 2005, 16, 1901–1912. [Google Scholar] [CrossRef] [PubMed]
- Kapp, T.G.; Rechenmacher, F.; Neubauer, S.; Maltsev, O.V.; Cavalcanti-Adam, E.A.; Zarka, R.; Reuning, U.; Notni, J.; Wester, H.J.; Mas-Moruno, C.; et al. A Comprehensive Evaluation of the Activity and Selectivity Profile of Ligands for RGD-binding Integrins. Sci. Rep. 2017, 7, 39805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lishko, V.K.; Yakubenko, V.P.; Ugarova, T.P. The interplay between integrins alphaMbeta2 and alpha5beta1during cell migration to fibronectin. Exp. Cell Res. 2003, 283, 116–126. [Google Scholar] [CrossRef]
- Ho, T.C.; Yeh, S.I.; Chen, S.L.; Tsao, Y.P. The Psoriasis Therapeutic Potential of a Novel Short Laminin Peptide C16. Int. J. Mol. Sci. 2019, 20, 3144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Li, M.; Li, B.; Wang, W.; Lin, A.; Sheng, M. Effect of reactive oxygen species generation in rabbit corneal epithelial cells on inflammatory and apoptotic signaling pathways in the presence of high osmotic pressure. PLoS ONE 2013, 8, e72900. [Google Scholar] [CrossRef] [Green Version]
- Storm, R.J.; Persson, B.D.; Skalman, L.N.; Frängsmyr, L.; Lindström, M.; Rankin, G.; Lundmark, R.; Domellöf, F.P.; Arnberg, N. Human Adenovirus Type 37 Uses αVβ1 and α3β1 Integrins for Infection of Human Corneal Cells. J. Virol. 2017, 91, e02019. [Google Scholar] [CrossRef] [Green Version]
- Du, F.; Garg, A.V.; Kosar, K.; Majumder, S.; Kugler, D.G.; Mir, G.H.; Maggio, M.; Henkel, M.; Lacy-Hulbert, A.; McGeachy, M.J. Inflammatory Th17 Cells Express Integrin αvβ3 for Pathogenic Function. Cell Rep. 2016, 16, 1339–1351. [Google Scholar] [CrossRef] [Green Version]
- Pflugfelder, S.C.; Corrales, R.M.; de Paiva, C.S. T helper cytokines in dry eye disease. Exp. Eye Res. 2013, 117, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Perez, V.L.; Pflugfelder, S.C.; Zhang, S.; Shojaei, A.; Haque, R. Lifitegrast, a Novel Integrin Antagonist for Treatment of Dry Eye Disease. Ocul. Surf. 2016, 14, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Fu, Y.C.; Becerra, C.R. Dissecting role of regulatory factors in NFkappaBpathway with siRNA. Acta Pharmacol. Sin. 2005, 26, 780–788. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Green, J.; Yurdagul, A., Jr.; Albert, P.; McInnis, M.C.; Orr, A.W. αvβ3 Integrins Mediate Flow-Induced NF-κB Activation, Proinflammatory Gene Expression, and Early Atherogenic Inflammation. Am. J. Pathol. 2015, 185, 2575–2589. [Google Scholar] [CrossRef] [Green Version]
- Antonov, A.S.; Antonova, G.N.; Munn, D.H.; Mivechi, N.; Lucas, R.; Catravas, J.D.; Verin, A.D. αVβ3 integrin regulates macrophage inflammatory responses via PI3 kinase/Akt-dependent NF-κB activation. J. Cell. Physiol. 2011, 226, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.M.; Jeong, K.; Lim, S.S. FAK Family Kinases in Vascular Diseases. Int. J. Mol. Sci. 2020, 21, 3630. [Google Scholar] [CrossRef]
- Li, R.; Ren, M.; Chen, N.; Luo, M.; Zhang, Z.; Wu, J. Vitronectin increases vascular permeability by promoting VE-cadherin internalization at cell junctions. PLoS ONE 2012, 7, e37195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hageman, G.S.; Mullins, R.F.; Russell, S.R.; Johnson, L.V.; Anderson, D.H. Vitronectin is a constituent of ocular drusen and the vitronectin gene is expressed in human retinal pigmented epithelial cells. FASEB J. 1999, 13, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.C.; Chen, S.L.; Shih, S.C.; Chang, S.J.; Yang, S.L.; Hsieh, J.W.; Cheng, H.C.; Chen, L.J.; Tsao, Y.P. Pigment epithelium-derived factor (PEDF) promotes tumor cell death by inducing macrophage membrane tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). J. Biol. Chem. 2011, 286, 35943–35954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dursun, D.; Wang, M.; Monroy, D.; Li, D.Q.; Lokeshwar, B.L.; Stern, M.E.; Pflugfelder, S.C. A mouse model of keratoconjunctivitis sicca. Invest. Ophthalmol. Vis. Sci. 2002, 43, 632–638. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer (Sense) | Primer (Antisense) | Accession No. |
|---|---|---|---|
| moItgAV | 5′-TGTTCACACTTTGGGCTGTG | 5′-GCCTCTATCCAGTCGACCAA | NM_008402.3 |
| moItgB1 | 5′-GGTCAGCAACGCATATCTGG | 5′-AATCAGCGATCCACAAACCG | NM_010578.2 |
| moItgB3 | 5′-GCTCATTGGCCTTGCTACTC | 5′-GTTGTTTGCTGTGTCCCACT | NM_016780.2 |
| moItgB5 | 5′-GACCTTTCTGCGAGTGTGAC | 5′-CTCTCCATGGCCTGAGCATA | NM_001145884.1 |
| moItgB6 | 5′-GCATTTGGAAGCTGCTGGTA | 5′-ATCTGAGGAAAGGCCTGCTT | NM_001159564.1 |
| moItgB8 | 5′-CTGGGCCAAAGTGAACACAA | 5′-CAACTGGACAGCCTTTGCTT | NM_177290.3 |
| moVTN | 5′-CATACTAGCCCTGGTGGCAT | 5′-CCATGAAACCCTGAGTGCAG | NM_011707.2 |
| moNOS2 | 5′-CCTTGTTCAGCTACGCCTTC | 5′-CTTCAGAGTCTGCCCATTGC | NM_010927.4 |
| moIL-10 | 5′-AGCTGAAGACCCTCAGGATG | 5′-CACTCTTCACCTGCTCCACT | NM_010548.2 |
| moArg1 | 5′-AAGACAGGGCTCCTTTCAGG | 5′-AGCAAGCCAAGGTTAAAGCC | NM_007482.3 |
| moTNF-α | 5′-CCAAATGGCCTCCCTCTCAT | 5′-CACTTGGTGGTTTGCTACGA | NM_013693.3 |
| moIL-1β | 5′-GGCTCATCTGGGATCCTCTC | 5′-GTTTGGAAGCAGCCCTTCAT | NM_008361.4 |
| moIL-6 | 5′-GGAGCCCACCAAGAACGATA | 5′-ACCAGCATCAGTCCCAAGAA | NM_031168.2 |
| mGAPDH | 5′-AACGGATTTGGCCGTATTGG | 5′-CATTCTCGGCCTTGACTGTG | NM_001289726.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, T.-C.; Yeh, S.-I.; Chen, S.-L.; Tsao, Y.-P. Integrin αv and Vitronectin Prime Macrophage-Related Inflammation and Contribute the Development of Dry Eye Disease. Int. J. Mol. Sci. 2021, 22, 8410. https://doi.org/10.3390/ijms22168410
Ho T-C, Yeh S-I, Chen S-L, Tsao Y-P. Integrin αv and Vitronectin Prime Macrophage-Related Inflammation and Contribute the Development of Dry Eye Disease. International Journal of Molecular Sciences. 2021; 22(16):8410. https://doi.org/10.3390/ijms22168410
Chicago/Turabian StyleHo, Tsung-Chuan, Shu-I Yeh, Show-Li Chen, and Yeou-Ping Tsao. 2021. "Integrin αv and Vitronectin Prime Macrophage-Related Inflammation and Contribute the Development of Dry Eye Disease" International Journal of Molecular Sciences 22, no. 16: 8410. https://doi.org/10.3390/ijms22168410
APA StyleHo, T.-C., Yeh, S.-I., Chen, S.-L., & Tsao, Y.-P. (2021). Integrin αv and Vitronectin Prime Macrophage-Related Inflammation and Contribute the Development of Dry Eye Disease. International Journal of Molecular Sciences, 22(16), 8410. https://doi.org/10.3390/ijms22168410