
Anthocyanin Biosynthesis Genes as Model Genes for Genome Editing in Plants


Abstract
:1. CRISPR/Cas Technology in Plant Science
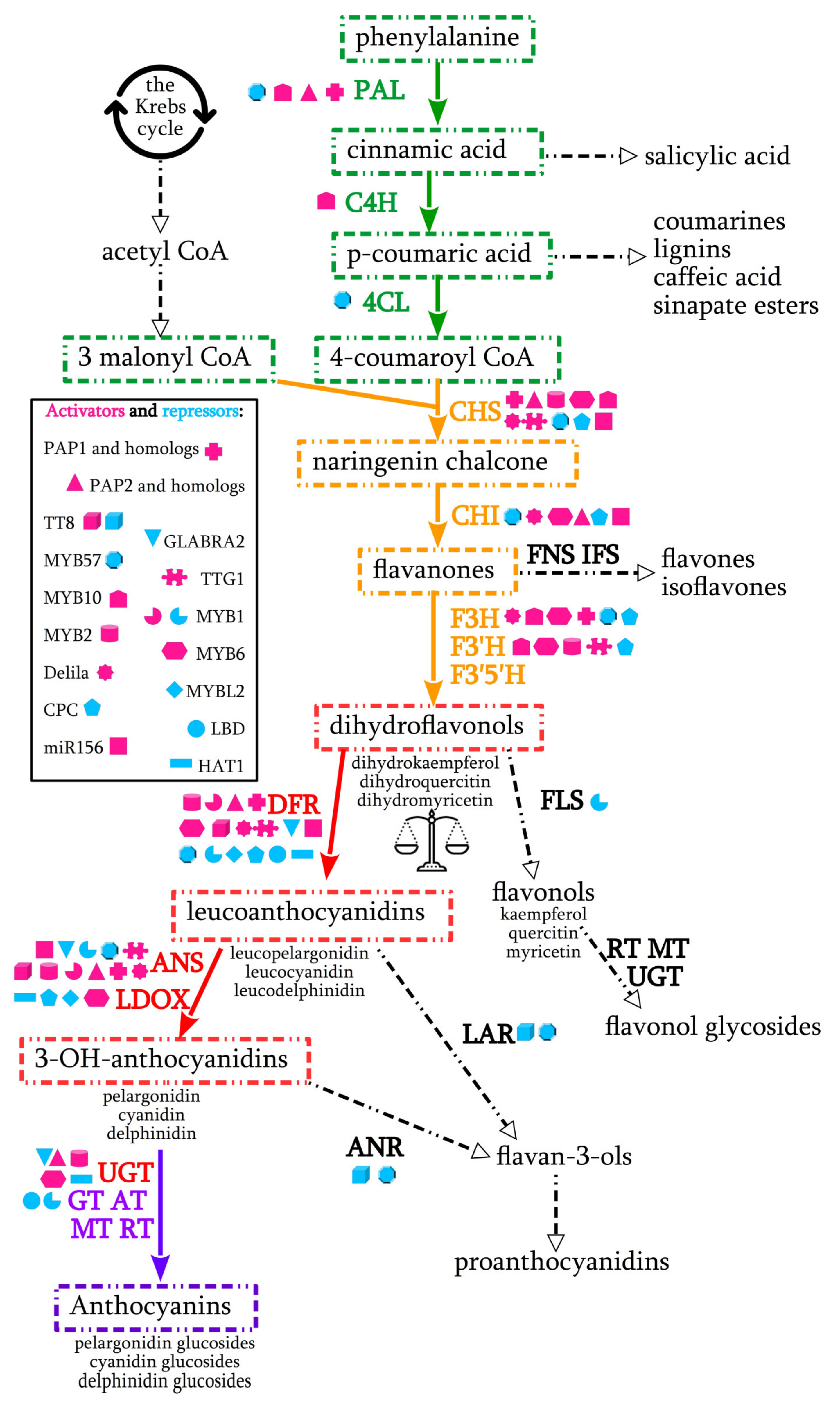
2. Anthocyanin Biosynthesis Pathway in Plants
3. R2R3MYB Transcriptional Activators
4. bHLH Transcriptional Activators
5. WD40 Transcriptional Activators
6. Transcriptional Repressors
6.1. R2R3 MYB
6.2. R3 MYB
6.3. Other Proteins
7. microRNAs
8. Structural Genes of Anthocyanin Biosynthesis
9. Conclusions
Funding
Conflicts of Interest
References
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-guided human genome engineering via cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Zhou, H.; Bi, H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic. Acids. Res. 2013, 41, e188. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, V.; Staskawicz, B.; Weigel, D.; Jones, J.D.G.; Kamoun, S. Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 691–693. [Google Scholar] [CrossRef]
- Kuluev, B.R.; Gumerova, G.R.; Mikhaylova, E.V.; Gerashchenkov, G.A.; Rozhnova, N.A.; Vershinina, Z.R.; Khyazev, A.V.; Matniyazov, R.T.; Baymiev, A.K.; Chemeris, A.V. Delivery of CRISPR/Cas components into higher plant cells for genome editing. Russ. J. Plant. Physiol. 2019, 66, 694–706. [Google Scholar] [CrossRef]
- Hahn, F.; Korolev, A.; Sanjurjo, L.L.; Nekrasov, V. A modular cloning toolkit for genome editing in plants. BMC Plant. Biol. 2020, 20, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.K.; Puchta, H. Novel CRISPR/Cas applications in plants: From prime editing to chromosome engineering. Transgenic Res. 2021, 30, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Malzahn, A.A.; Tang, X.; Lee, K.; Ren, Q.; Sretenovic, S.; Zhang, Y.; Qi, Y. Application of CRISPR-Cas12a temperature sensitivity for improved genome editing in rice, maize, and Arabidopsis. BMC Bio. 2019, 17, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Lowder, L.G.; Zhang, T.; Malzahn, A.A.; Zheng, X.; Voytas, D.F.; Qi, Y. A CRISPR–Cpf1 system for efficient genome editing and transcriptional repression in plants. Nat. Plants 2017, 3, 1–5. [Google Scholar] [CrossRef]
- Begemann, M.B.; Gray, B.N.; January, E.; Singer, A.; Kesler, D.C.; He, Y.; Oufattole, M. Characterization and validation of a novel group of type V, class 2 nucleases for in vivo genome editing. bioRxiv 2017. [Google Scholar] [CrossRef] [Green Version]
- Chavez, A.; Scheiman, J.; Vora, S.; Pruitt B., W.; Tuttle, M.; Iyer E., P.; Church G., M. Highly efficient Cas9-mediated transcriptional programming. Nat. Methods 2015, 12, 326–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.J.; Dempewolf, E.; Zhang, W.; Wang, Z.Y. RNA-guided transcriptional activation via CRISPR/dCas9 mimics overexpression phenotypes in Arabidopsis. PLoS ONE 2017, 12, e0179410. [Google Scholar] [CrossRef] [Green Version]
- Mishra, R.; Joshi, R.K.; Zhao, K. Base editing in crops: Current advances, limitations and future implications. Plant. Biotechnol. J. 2020, 18, 20–31. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Liu, D.R. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Schiml, S.; Fauser, F.; Puchta, H. The CRISPR/C as system can be used as nuclease for in planta gene targeting and as paired nickases for directed mutagenesis in Arabidopsis resulting in heritable progeny. Plant. J. 2014, 80, 1039–1050. [Google Scholar] [CrossRef]
- Cermak, T.; Baltes, N.J.; Cegan, R.; Zhang, Y.; Voytas, D.F. High-frequency, precise modification of the tomato genome. Genome Biol. 2015, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Butler, N.M.; Baltes, N.J.; Voytas, D.F.; Douches, D.S. Geminivirus-mediated genome editing in potato (Solanum tuberosum L.) using sequence-specific nucleases. Front. Plant Sci. 2016, 7, 1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cermak, T.; Curtin, S.J.; Gil-Humanes, J.; Čegan, R.; Kono, T.J.; Konečná, E.; Voytas, D.F. A multipurpose toolkit to enable advanced genome engineering in plants. Plant. Cell 2017, 29, 1196–1217. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Zhang, Q.; Zhu, Q.; Liu, W.; Chen, Y.; Qiu, R.; Liu, Y.G. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gao, Y.; Gao, Y.; Zhang, Q. Targeted deletion of floral development genes in Arabidopsis with CRISPR/Cas9 using the RNA endoribonuclease Csy4 processing system. Hort. Res. 2019, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Xie, K.; Minkenberg, B.; Yang, Y. Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system. Proc. Natl. Acad. Sci. USA 2015, 112, 3570–3575. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Zhao, Y. Self-processing of ribozyme-flanked RNAs into guide RNAs in vitro and in vivo for CRISPR-mediated genome editing. J. Integr. Plant. Biol. 2014, 56, 343–349. [Google Scholar] [CrossRef]
- Hajiahmadi, Z.; Movahedi, A.; Wei, H.; Li, D.; Orooji, Y.; Ruan, H.; Zhuge, Q. Strategies to increase on-target and reduce off-target effects of the CRISPR/Cas9 system in plants. Int. J. Mol. Sci. 2019, 20, 3719. [Google Scholar] [CrossRef] [Green Version]
- Hahn, F.; Nekrasov, V. CRISPR/Cas precision: Do we need to worry about off-targeting in plants? Plant. Cell Rep. 2019, 38, 437–441. [Google Scholar] [CrossRef] [Green Version]
- Waltz, E. Gene-edited CRISPR mushroom escapes US regulation. Nature 2016, 532, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menz, J.; Modrzejewski, D.; Hartung, F.; Wilhelm, R.; Sprink, T. Genome edited crops touch the market: A view on the global development and regulatory environment. Front. Plant. Sci. 2020, 11, 586027. [Google Scholar] [CrossRef] [PubMed]
- Waltz, E. Appetite grows for biotech foods with health benefits. Nat. Biotechnol. 2019, 37, 573–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonaka, S.; Arai, C.; Takayama, M.; Matsukura, C.; Ezura, H. Efficient increase of ɣ-aminobutyric acid (GABA) content in tomato fruits by targeted mutagenesis. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lowder, L.G.; Zhang, D.; Baltes, N.J.; Paul, J.W.; Tang, X.; Zheng, X.; Qi, Y. A CRISPR/Cas9 toolbox for multiplexed plant genome editing and transcriptional regulation. Plant. Physiol. 2015, 169, 971–985. [Google Scholar] [CrossRef] [Green Version]
- Butler, N.M.; Atkins, P.A.; Voytas, D.F.; Douches, D.S. Generation and inheritance of targeted mutations in potato (Solanum tuberosum L.) using the CRISPR/Cas system. PLoS ONE 2015, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Dahan-Meir, T.; Filler-Hayut, S.; Melamed-Bessudo, C.; Bocobza, S.; Czosnek, H.; Aharoni, A.; Levy, A.A. Efficient in planta gene targeting in tomato using geminiviral replicons and the CRISPR/Cas9 system. Plant. J. 2018, 95, 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hummel, A.W.; Chauhan, R.D.; Čermák, T.; Mutka, A.M.; Vijayaraghavan, A.; Boyher, A.; Starker, C.G. Allele exchange at the EPSPS locus confers glyphosate tolerance in cassava. Plant. Biotechnol. J. 2018, 16, 1275–1282. [Google Scholar] [CrossRef] [Green Version]
- Vu, T.V.; Sivankalyani, V.; Kim, E.J.; Doan, D.T.H.; Tran, M.T.; Kim, J.; Kim, J.Y. Highly efficient homology-directed repair using CRISPR/Cpf1-geminiviral replicon in tomato. Plant. Biotechnol. J. 2020, 18, 2133–2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malone, L.A.; Barraclough, E.I.; Lin-Wang, K.; Stevenson, D.E.; Allan, A.C. Effects of red-leaved transgenic tobacco expressing a MYB transcription factor on two herbivorous insects, Spodoptera litura and Helicoverpa armigera. Entomol. Exp. Appl. 2009, 133, 117–127. [Google Scholar] [CrossRef]
- Mitsunami, T.; Nishihara, M.; Galis, I.; Alamgir, K.M.; Hojo, Y.; Fujita, K.; Arimura, G.I. Overexpression of the PAP1 transcription factor reveals a complex regulation of flavonoid and phenylpropanoid metabolism in Nicotiana tabacum plants attacked by Spodoptera litura. PLoS ONE 2014, 9, e108849. [Google Scholar] [CrossRef]
- Gandikota, M.; de Kochko, A.; Chen, L.; Ithal, N.; Fauquet, C.; Reddy, A.R. Development of transgenic rice plants expressing maize anthocyanin genes and increased blast resistance. Mol. Breed. 2001, 7, 73–83. [Google Scholar] [CrossRef]
- Kangatharalingam, N.; Pierce, M.L.; Bayles, M.B.; Essenberg, M. Epidermal anthocyanin production as an indicator of bacterial blight resistance in cotton. Physiol. Mol. Plant. Pathol. 2002, 61, 189–195. [Google Scholar] [CrossRef]
- Shoeva, O.Y.; Glagoleva, A.Y.; Khlestkina, E.K. The factors affecting the evolution of the anthocyanin biosynthesis pathway genes in monocot and dicot plant species. BMC Plant. Biol. 2017, 17, 5–14. [Google Scholar] [CrossRef]
- Springob, K.; Nakajima, J.I.; Yamazaki, M.; Saito, K. Recent advances in the biosynthesis and accumulation of anthocyanins. Nat. Prod. Rep. 2003, 20, 288–303. [Google Scholar] [CrossRef]
- Cominelli, E.; Gusmaroli, G.; Allegra, D.; Galbiati, M.; Wade, H.K.; Jenkins, G.I.; Tonelli, C. Expression analysis of anthocyanin regulatory genes in response to different light qualities in Arabidopsis thaliana. J. Plant. Physiol. 2008, 165, 886–894. [Google Scholar] [CrossRef]
- Guo, N.; Cheng, F.; Wu, J.; Liu, B.; Zheng, S.; Liang, J.; Wang, X. Anthocyanin biosynthetic genes in Brassica rapa. BMC Genom. 2014, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Goswami, G.; Nath, U.K.; Park, J.I.; Hossain, M.R.; Biswas, M.K.; Kim, H.T.; Kim, H.R.; Nou, I.S. Transcriptional regulation of anthocyanin biosynthesis in a high-anthocyanin resynthesized Brassica napus cultivar. J. Biol. Res.-Thessaloniki 2018, 25, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Borevitz, J.O.; Xia, Y.; Blount, J.; Dixon, R.A.; Lamb, C. Activation tagging identifies a conserved MYB regulator of phenylpropanoid biosynthesis. Plant. Cell 2000, 12, 2383–2393. [Google Scholar] [CrossRef] [Green Version]
- Davies, K.M.; Schwinn, K.E.; Deroles, S.C.; Manson, D.G.; Lewis, D.H.; Bloor, S.J.; Bradley, J.M. Enhancing anthocyanin production by altering competition for substrate between flavonol synthase and dihydroflavonol 4-reductase. Euphytica 2003, 131, 259–268. [Google Scholar] [CrossRef]
- Hughes, N.M.; Connors, M.K.; Grace, M.H.; Lila, M.A.; Willans, B.N.; Wommack, A.J. The same anthocyanins served four different ways: Insights into anthocyanin structure-function relationships from the wintergreen orchid, Tipularia discolor. Plant. Sci. 2021, 303, 110793. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuka, T.; Sasaki, N.; Nishihara, M. Transcriptional regulators of flavonoid biosynthesis and their application to flower color modification in Japanese gentians. Plant. Biotechnol. 2014, 14, 0731. [Google Scholar] [CrossRef] [Green Version]
- Tasaki, K.; Higuchi, A.; Watanabe, A.; Sasaki, N.; Nishihara, M. Effects of knocking out three anthocyanin modification genes on the blue pigmentation of gentian flowers. Scientific Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.H.; Tagami, T.; Matsunaga, K.; Okuyama, M.; Suzuki, T.; Noda, N.; Shimura, H. Functional characterization of UDP-rhamnose-dependent rhamnosyltransferase involved in anthocyanin modification, a key enzyme determining blue coloration in Lobelia erinus. Plant. J. 2017, 89, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Yonekura-Sakakibara, K.; Tohge, T.; Niida, R.; Saito, K. Identification of a Flavonol 7-O-Rhamnosyltransferase Gene Determining Flavonoid Pattern in Arabidopsis by Transcriptome Coexpression Analysis and Reverse Genetics. J. Biol. Chem. 2007, 282, 14932–14941. [Google Scholar] [CrossRef] [Green Version]
- Dubos, C.; Le Gourrierec, J.; Baudry, A.; Huep, G.; Lanet, E.; Debeaujon, I.; Routaboul, J.M.; Alboresi, A.; Weisshaar, B.; Lepiniec, L. MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. Plant. J. 2008, 55, 940–953. [Google Scholar] [CrossRef]
- Matsui, K.; Umemura, Y.; Ohme-Takagi, M. AtMYBL2, a protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis. Plant. J. 2008, 55, 954–967. [Google Scholar] [CrossRef]
- Zhu, H.F.; Fitzsimmons, K.; Khandelwaland, A.; Kranz, R.G. CPC, a single-repeat R3 MYB, is a negative regulator of anthocyanin biosynthesis in Arabidopsis. Mol. Plant. 2009, 2, 790–802. [Google Scholar] [CrossRef]
- Rubin, G.; Tohge, T.; Matsuda, F.; Saito, K.; Scheible, W.-R. Members of the LBD family of transcription factors repress anthocyanin synthesis and affect additional nitrogen responses in Arabidopsis. Plant. Cell 2009, 21, 3567–3584. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Dubos, C.; Lepiniec, L. Transcriptional control of avonoid biosynthesis by MYB-bHLH-WDR complexes. Trends Plant. Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Baudry, A.; Heim, M.A.; Dubreucq, B.; Caboche, M.; Weisshaar, B.; Lepiniec, L. TT2, TT8, and TTG1 synergistically specify the expression of BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana. Plant. J. 2004, 39, 366–380. [Google Scholar] [CrossRef] [PubMed]
- Hichri, I.; Barrieu, F.; Bogs, J.; Kappel, C.; Delrot, S.; Lauvergeat, V. Recent advances in the transcriptional regulation of the flavonoid biosynthetic pathway. J. Exp. Bot. 2011, 62, 2465–2483. [Google Scholar] [CrossRef] [Green Version]
- Luo, Q.J.; Mittal, A.; Jia, F. An autoregulatory feedback loop involving PAP1 and TAS4 in response to sugars in Arabidopsis. Plant. Mol. Biol 2012, 80, 117–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Cai, J.; Yang, Y. Overexpression of microRNA828 reduces anthocyanin accumulation in Arabidopsis. Plant. Cell Tiss. Organ. Cult. 2013, 115, 159–167. [Google Scholar] [CrossRef]
- Sunitha, S.; Rock, C.D. CRISPR/Cas9-mediated targeted mutagenesis of TAS4 and MYBA7 loci in grapevine rootstock 101-14. Transgenic Res. 2020, 29, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Chiu, L.W.; Li, L. Transcriptional regulation of anthocyanin biosynthesis in red cabbage. Planta 2009, 230, 1141. [Google Scholar] [CrossRef]
- He, Q.; Ren, Y.; Zhao, W.; Li, R.; Zhang, L. Low temperature promotes anthocyanin biosynthesis and related gene expression in the seedlings of purple head chinese cabbage (Brassica rapa L.). Genes 2020, 11, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Lu, W.; Ran, L.; Dou, L.; Yao, S.; Hu, J.; Luo, K. R2R3-MYB transcription factor MYB6 promotes anthocyanin and proanthocyanidin biosynthesis but inhibits secondary cell wall formation in Populus tomentosa. Plant. J. 2019, 99, 733–751. [Google Scholar] [CrossRef]
- Riaz, B.; Chen, H.; Wang, J.; Du, L.; Wang, K.; Ye, X. Overexpression of maize ZmC1 and ZmR transcription factors in wheat regulates anthocyanin biosynthesis in a tissue-specific manner. Int. J. Mol. Sci. 2019, 20, 5806. [Google Scholar] [CrossRef] [Green Version]
- Ververidis, F.; Trantas, E.; Douglas, C.; Vollmer, G.; Kretzschmar, G.; Panopoulos, N. Biotechnology of flavonoids and other phenylpropanoid-derived natural products. Part II: Reconstruction of multienzyme pathways in plants and microbes. J. Biotechnol. 2007, 2, 1235–1249. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Chen, N.; Huang, Z.; Li, D.; Zhi, J.; Yu, B.; Qiu, Z. Anthocyanin Fruit encodes an R2R3-MYB transcription factor, SlAN2-like, activating the transcription of SlMYBATV to fine-tune anthocyanin content in tomato fruit. New Phytol. 2020, 225, 2048–2063. [Google Scholar] [CrossRef]
- Zhi, J.; Liu, X.; Li, D.; Huang, Y.; Yan, S.; Cao, B.; Qiu, Z. CRISPR/Cas9-mediated SlAN2 mutants reveal various regulatory models of anthocyanin biosynthesis in tomato plant. Plant. Cell Rep. 2020, 39, 799–809. [Google Scholar] [CrossRef]
- Xu, Z.S.; Feng, K.; Xiong, A.S. CRISPR/cas9-mediated multiply targeted mutagenesis in orange and purple carrot plants. Mol. Biotechnol. 2019, 61, 191–199. [Google Scholar] [CrossRef]
- Zhai, Y.; Yu, K.; Cai, S.; Hu, L.; Amoo, O.; Xu, L.; Zhou, Y. Targeted mutagenesis of BnTT8 homologs controls yellow seed coat development for effective oil production in Brassica napus L. Plant. Biotechnol. J. 2020, 18, 1153–1168. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Liu, X.; Li, T.; Qin, H.; Li, X.; Chen, K.; Xu, Y. CRISPR-Cas9 Mediated Knockout of NtAn1 to Enhance the Lipid Accumulation in Tobacco Seed for Biodiesel Production. 2020. Available online: https://www.researchsquare.com/article/rs-37669/v1 (accessed on 16 August 2021). [CrossRef]
- Ryder, P.; McHale, M.; Fort, A.; Spillane, C. Generation of stable nulliplex autopolyploid lines of Arabidopsis thaliana using CRISPR/Cas9 genome editing. Plant. Cell Rep. 2017, 36, 1005–1008. [Google Scholar] [CrossRef]
- Yang, X.; Wang, J.; Xia, X.; Zhang, Z.; He, J.; Nong, B.; Deng, G. OsTTG1, a WD40 repeat gene, regulates anthocyanin biosynthesis in rice. Plant. J. 2021, 151, 1114–1129. [Google Scholar] [CrossRef]
- Wan, S.; Li, C.; Ma, X. PtrMYB57 contributes to the negative regulation of anthocyanin and proanthocyanidin biosynthesis in poplar. Plant. Cell Rep. 2017, 36, 1263–1276. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, Z.; Feng, Z.; Wei, P.; Zhang, H.; Botella, J.R.; Zhu, J.K. Development of germ-line-specific CRISPR-Cas9 systems to improve the production of heritable gene modifications in Arabidopsis. Plant. Biotechnol. J. 2016, 14, 519–532. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.; Sinkevicius, K.W.; Selinger, D.A.; Tarczynski, M.C. The homeobox gene GLABRA2 affects seed oil content in Arabidopsis. Plant. Mol. Biol 2006, 60, 377–387. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Hu, Q.; Dai, X.; Tian, H.; Zheng, K.; Wang, X.; Mao, T.; Chen, J.G.; Wang, S. Characterization of an activation-tagged mutant uncovers a role of GLABRA2 in anthocyanin biosynthesis in Arabidopsis. Plant. J. 2015, 83, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Kobayashi, A.; Endo, M. CRISPR/Cas9-mediated mutagenesis of the dihydroflavonol-4-reductase-B (DFR-B) locus in the Japanese morning glory Ipomoea (Pharbitis) nil. Sci. Rep. 2017, 7, 10028. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.J.; Lee, H.J.; Kim, J.H. CRISPR/Cas9-targeted mutagenesis of F3′H, DFR and LDOX, genes related to anthocyanin biosynthesis in black rice (Oryza sativa L.). Plant. Biotechnol. Rep. 2019, 13, 521–531. [Google Scholar] [CrossRef]
- Danilo, B.; Perrot, L.; Botton, E.; Nogué, F.; Mazier, M. The DFR locus: A smart landing pad for targeted transgene insertion in tomato. PLoS ONE 2018, 13, e0208395. [Google Scholar] [CrossRef] [PubMed]
- Selma, S.; Bernabé-Orts, J.M.; Vazquez-Vilar, M.; Diego-Martin, B.; Ajenjo, M.; Garcia-Carpintero, V.; Orzaez, D. Strong gene activation in plants with genome-wide specificity using a new orthogonal CRISPR/Cas9-based programmable transcriptional activator. Plant Biotechnol. J. 2019, 17, 1703. [Google Scholar] [CrossRef] [Green Version]
- Char, S.N.; Neelakandan, A.K.; Nahampun, H.; Frame, B.; Main, M.; Spalding, M.H.; Yang, B. An Agrobacterium-delivered CRISPR/Cas9 system for high-frequency targeted mutagenesis in maize. Plant. Biotechnol. J. 2017, 15, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, Y.J.; Zhang, F.J.; Zhang, G.Z.; Jiang, X.Y.; Yu, H.M.; Hou, B.K. The Arabidopsis UDP-glycosyltransferases UGT79B2 and UGT79B3, contribute to cold, salt and drought stress tolerance via modulating anthocyanin accumulation. Plant. J. 2017, 89, 85–103. [Google Scholar] [CrossRef] [Green Version]
- Nitarska, D.; Boehm, R.; Debener, T.; Lucaciu, R.C.; Halbwirth, H. First genome edited poinsettias: Targeted mutagenesis of flavonoid 3′-hydroxylase using CRISPR/Cas9 results in a colour shift. Plant. Cell Tissue Organ. Culture (PCTOC) 2021, 1–12. [Google Scholar] [CrossRef]
- Nishihara, M.; Higuchi, A.; Watanabe, A.; Tasaki, K. Application of the CRISPR/Cas9 system for modification of flower color in Torenia fournieri. BMC Plant. Biol. 2018, 18, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Klimek-Chodacka, M.; Oleszkiewicz, T.; Baranski, R. Visual Assay for Gene Editing Using a CRISPR/Cas9 System in Carrot Cells. In Plant Genome Editing with CRISPR Systems; Humana Press Inc: New York, NY, USA, 2019; Volume 1917, pp. 203–2015. [Google Scholar] [CrossRef]
- Liu, J.; Wang, S.; Wang, H. Rapid generation of tomato male-sterile lines with a marker use for hybrid seed production by CRISPR/Cas9 system. Mol. Breeding 2021, 41, 25. [Google Scholar] [CrossRef]
- Lowder, L.G.; Paul, J.W.; Qi, Y. Multiplexed transcriptional activation or repression in plants using CRISPR-dCas9 based systems. Methods Mol. Biol. 2017, 1629, 167–184. [Google Scholar] [CrossRef]
- Hirohata, A.; Sato, I.; Kaino, K. CRISPR/Cas9-mediated homologous recombination in tobacco. Plant. Cell Rep. 2019, 38, 463–473. [Google Scholar] [CrossRef]
- Xu, Z.S.; Yang, Q.Q.; Feng, K.; Yu, X.; Xiong, A.S. DcMYB113, a root-specific R2R3-MYB, conditions anthocyanin biosynthesis and modification in carrot. Plant. Biotechnol. J. 2020, 18, 1585–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Hu, Z.; Zhang, Y. A putative functional MYB transcription factor induced by low temperature regulates anthocyanin biosynthesis in purple kale (Brassica Oleracea var. acephala f. tricolor). Plant. Cell Rep. 2012, 31, 281–289. [Google Scholar] [CrossRef]
- He, Y.; Zhu, M.; Wu, J.; Ouyang, L.; Wang, R.; Sun, H.; Zhao, Y. Repurposing of anthocyanin biosynthesis for plant transformation and genome editing. Front. Genome Ed. 2020, 2, 19. [Google Scholar] [CrossRef]
- Li, W.; Ding, Z.; Ruan, M. Kiwifruit R2R3-MYB transcription factors and contribution of the novel AcMYB75 to red kiwifruit anthocyanin biosynthesis. Sci Rep. 2017, 7, 16861. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.H.; Choi, M.G.; Kang, C.S.; Park, C.S.; Choi, S.B.; Park, Y.I. A wheat R2R3-MYB protein PURPLE PLANT1 (TaPL1) functions as a positive regulator of anthocyanin biosynthesis. Biochem. Biophys. Res. Commun. 2016, 469, 686–691. [Google Scholar] [CrossRef]
- Li, Y.; Shan, X.; Tong, L.; Wei, C.; Lu, K.; Li, S.; Gao, X. The Conserved and Particular Roles of the R2R3-MYB Regulator FhPAP1 from Freesia hybrida in flower anthocyanin biosynthesis. Plant. Cell Physiol. 2020, 61, 1365–1380. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Liu, H.; Lou, Q.; & Liu, Y. Ectopic expression of the grape hyacinth (Muscari armeniacum) R2R3-MYB transcription factor gene, MaAN2, induces anthocyanin accumulation in tobacco. Front. Plant. Sci. 2017, 8, 965. [Google Scholar] [CrossRef] [Green Version]
- Zong, Y.; Zhu, X.; Liu, Z. Functional MYB transcription factor encoding gene AN2 is associated with anthocyanin biosynthesis in Lycium ruthenicum Murray. BMC Plant. Biol. 2019, 19, 169. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, X.; Li, H.; Liu, S.; Jin, L.; Lyu, Y.; Lyu, S. Anthocyanin, a novel and user-friendly reporter for convenient, non-destructive, low cost, directly visual selection of transgenic hairy roots in the study of rhizobia-legume symbiosis. Plant. Methods 2020, 16, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhang, K.X.; Muhammad, K.; Li, J.B.; Zhou, M.L. MYB transcription repressors regulate plant secondary metabolism. Crit. Rev. Plant. Sci. 2019, 38, 159–170. [Google Scholar] [CrossRef]
- Li, N.; Wu, H.; Ding, Q. The heterologous expression of Arabidopsis PAP2 induces anthocyanin accumulation and inhibits plant growth in tomato. Funct. Integr. Genomics 2018, 18, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Gu, C.; Wang, X.; Gao, S.; Li, C.; Zhao, C.; Li, C.; Ma, C.; Zhang, Q. Identification of the Eutrema salsugineum EsMYB90 gene important for anthocyanin biosynthesis. BMC Plant. Biol. 2020, 20, 186. [Google Scholar] [CrossRef]
- Schwinn, K.E.; Ngo, H.; Kenel, F.; Brummell, D.A.; Albert, N.W.; McCallum, J.A.; Davies, K.M. The onion (Allium cepa L.) R2R3-MYB gene MYB1 regulates anthocyanin biosynthesis. Front. Plant. Sci. 2016, 7, 1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, G.; Kim, J.S.; Park, J.E.; Shin, H.; Yu, S.H.; Park, S.; Huh, J.H. MYB1 transcription factor is a candidate responsible for red root skin in radish (Raphanus sativus L.). PLoS ONE 2018, 13, e0204241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takos, A.M.; Jaffé, F.W.; Jacob, S.R.; Bogs, J.; Robinson, S.P.; Walker, A.R. Light-induced expression of a MYB gene regulates anthocyanin biosynthesis in red apples. Plant. Physiol. 2006, 142, 1216–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aharoni, A.; De Vos, C.R.; Wein, M.; Sun, Z.; Greco, R.; Kroon, A.; O’Connell, A.P. The strawberry FaMYB1 transcription factor suppresses anthocyanin and flavonol accumulation in transgenic tobacco. Plant. J. 2001, 28, 319–332. [Google Scholar] [CrossRef]
- Lai, B.; Cheng, Y.; Liu, H. Differential anthocyanin accumulation in radish taproot: Importance of RsMYB1 gene structure. Plant. Cell Rep. 2020, 39, 217–226. [Google Scholar] [CrossRef]
- Kanzaki, S.; Ichihi, A.; Tanaka, Y.; Fujishige, S.; Koeda, S.; Shimizu, K. The R2R3-MYB transcription factor MiMYB1 regulates light dependent red coloration of ‘Irwin’mango fruit skin. Sci. Hortic. 2020, 272, 109567. [Google Scholar] [CrossRef]
- Paolocci, F.; Robbins, M.P.; Passeri, V.; Hauck, B.; Morris, P.; Rubini, A.; Damiani, F. The strawberry transcription factor FaMYB1 inhibits the biosynthesis of proanthocyanidins in Lotus corniculatus leaves. J. Exp. Bot. 2011, 62, 1189–1200. [Google Scholar] [CrossRef] [Green Version]
- Kadomura-Ishikawa, Y.; Miyawaki, K.; Takahashi, A.; Noji, S. RNAi-mediated silencing and overexpression of the FaMYB1 gene and its effect on anthocyanin accumulation in strawberry fruit. Biol. Plant. 2015, 59, 677–685. [Google Scholar] [CrossRef]
- Xiang, L.; Liu, X.; Li, H.; Yin, X.; Grierson, D.; Li, F.; Chen, K. CmMYB#7, an R3 MYB transcription factor, acts as a negative regulator of anthocyanin biosynthesis in chrysanthemum. J. Exp. Bot. 2019, 70, 3111–3123. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.H.; Song, J.H.; Kim, D.H.; Kim, J.K.; Lee, J.Y.; Kim, Y.M.; Ha, S.H. Activation of anthocyanin biosynthesis by expression of the radish R2R3-MYB transcription factor gene RsMYB1. Plant. Cell Rep. 2016, 35, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.H.; Kim, D.H.; Kim, J.K.; Lee, J.Y.; Ha, S.H. A radish basic helix-loop-helix transcription factor, RsTT8 acts a positive regulator for anthocyanin biosynthesis. Front. Plant. Sci. 2017, 8, 1917. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xu, Z.; Yu, X.; Zhao, L.; Zhao, M.; Han, X.; Qi, S. Identification of two novel R2R3-MYB transcription factors, PsMYB114L and PsMYB12L, related to anthocyanin biosynthesis in Paeonia suffruticosa. Int. J. Mol. Sci. 2019, 20, 1055. [Google Scholar] [CrossRef] [Green Version]
- Hatlestad, G.; Akhavan, N.; Sunnadeniya, R. The beet Y locus encodes an anthocyanin MYB-like protein that activates the betalain red pigment pathway. Nat. Genet. 2015, 47, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Roosa, A.E.; Laitinen, M.A.; Suvi, K.; Broholm, T.H.; Teeri, P.E. Identification of target genes for a MYB-type anthocyanin regulator in Gerbera hybrida. J. Exp. Bot. 2008, 59, 3691–3703. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.; Wang, Y.; Yang, S. Anthocyanin biosynthesis in pears is regulated by a R2R3-MYB transcription factor PyMYB10. Planta 2010, 232, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Castillejo, C.; Waurich, V.; Wagner, H.; Ramos, R.; Oiza, N.; Muñoz, P.; Amaya, I. Allelic variation of MYB10 is the major force controlling natural variation in skin and flesh color in strawberry (Fragaria spp.) fruit. Plant. Cell 2020, 32, 3723–3749. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, W.; Wang, X.; Yang, R.; Wu, Z.; Wang, H.; Fu, C. MiR156 regulates anthocyanin biosynthesis through SPL targets and other microRNAs in poplar. Hort. Res. 2020, 7, 1–12. [Google Scholar] [CrossRef]
- Hawkins, C.; Caruana, J.; Schiksnis, E. Genome-scale DNA variant analysis and functional validation of a SNP underlying yellow fruit color in wild strawberry. Sci. Rep. 2016, 6, 29017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; Wu, J.; Xue, Y. The novel gene BrMYB2, located on chromosome A07, with a short intron 1 controls the purple-head trait of Chinese cabbage (Brassica rapa L.). Hortic. Res. 2020, 7, 97. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.W.; Li, L. Characterization of the regulatory network of BoMYB2 in controlling anthocyanin biosynthesis in purple cauliflower. Planta 2012, 236, 1153–1164. [Google Scholar] [CrossRef]
- He, Q.; Lu, Q.; He, Y.; Wang, Y.; Zhang, N.; Zhao, W.; Zhang, L. Dynamic changes of the anthocyanin biosynthesis mechanism during the development of heading Chinese cabbage (Brassica rapa L.) and Arabidopsis under the control of BrMYB2. Front. Plant. Sci. 2020, 11, 593766. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Liu, J.X.; Duan, A.Q. AgMYB2 transcription factor is involved in the regulation of anthocyanin biosynthesis in purple celery (Apium graveolens L.). Planta 2018, 248, 1249–1261. [Google Scholar] [CrossRef]
- Chen, D.; Liu, Y.; Yin, S.; Qiu, J.; Jin, Q.; King, G.J.; Wang, J.; Ge, X.; Li, Z. Alternatively spliced bnapap2.a7 isoforms play opposing roles in anthocyanin biosynthesis of Brassica napus L. Front. Plant. Sci. 2020, 11, 983. [Google Scholar] [CrossRef]
- Xu, Z.S.; Feng, K.; Que, F. A MYB transcription factor, DcMYB6, is involved in regulating anthocyanin biosynthesis in purple carrot taproots. Sci. Rep. 2017, 7, 45324. [Google Scholar] [CrossRef] [Green Version]
- Naing, A.H.; Park, K.I.; Ai, T.N. Overexpression of snapdragon Delila (Del) gene in tobacco enhances anthocyanin accumulation and abiotic stress tolerance. BMC Plant. Biol. 2017, 17, 65. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Holme, I.B.; Dionisio, G. Cyanidin based anthocyanin biosynthesis in orange carrot is restored by expression of AmRosea1 and AmDelila, MYB and bHLH transcription factors. Plant. Mol. Biol. 2020, 103, 443–456. [Google Scholar] [CrossRef]
- Nie, X.; Zhao, Z.P.; Chen, G.P. Brassica napus possesses enhanced antioxidant capacity via heterologous expression of anthocyanin pathway gene transcription factors. Russ. J. Plant. Physiol. 2013, 60, 108–115. [Google Scholar] [CrossRef]
- Song, H.; Yi, H.; Lee, M.; Han, C.T.; Lee, J.; Kim, H.; Hur, Y. Purple Brassica oleracea var. capitata F. rubra is due to the loss of BoMYBL2–1 expression. BMC Plant. Biol. 2018, 18, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sawa, S. Overexpression of the AtmybL2 gene represses trichome development in Arabidopsis. DNA Res. 2002, 9, 31–34. [Google Scholar] [CrossRef] [Green Version]
- Gates, D.J.; Olson, B.J.; Clemente, T.E.; Smith, S.D. A novel R3 MYB transcriptional repressor associated with the loss of floral pigmentation in Iochroma. New Phytol. 2018, 217, 1346–1356. [Google Scholar] [CrossRef] [Green Version]
- Nemie-Feyissa, D.; Olafsdottir, S.M.; Heidari, B.; Lillo, C. Nitrogen depletion and small R3-MYB transcription factors affecting anthocyanin accumulation in Arabidopsis leaves. Phytochemistry 2014, 98, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Kirik, V.; Simon, M.; Huelskamp, M.; Schiefelbein, J. The ENHANCER OF TRY AND CPC1 gene acts redundantly with TRIPTYCHON and CAPRICE in trichome and root hair cell patterning in Arabidopsis. Dev. Biol. 2004, 268, 506–513. [Google Scholar] [CrossRef] [Green Version]
- Li, H.H.; Liu, X.; An, J.P.; Hao, Y.J.; Wang, X.F.; You, C.X. Cloning and elucidation of the functional role of apple MdLBD13 in anthocyanin biosynthesis and nitrate assimilation. Plant. Cell Tissue Organ. Cult. 2017, 130, 1–13. [Google Scholar] [CrossRef]
- Ren, J.; Liu, Z.; Chen, W.; Xu, H.; Feng, H. Anthocyanin degrading and chlorophyll accumulation lead to the formation of bicolor leaf in ornamental kale. Int. J. Mol. Sci. 2019, 20, 603. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Cao, M.; Chen, X.; Ye, M.; Zhao, P.; Nan, Y.; Chen, Q. Genome-wide analysis of the Lateral Organ Boundaries Domain (LBD) gene family in Solanum tuberosum. Int. J. Mol. Sci. 2019, 20, 5360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, G.; Li, Z.; Song, Y.; Zhu, H.; Lin, S.; Huang, R.; Duan, X. LcNAC13 Physically Interacts with LcR1MYB1 to Coregulate Anthocyanin Biosynthesis-Related Genes during Litchi Fruit Ripening. Biomolecules 2019, 9, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, J.Y.; Jung, Y.H.; Song, H. Alleles disrupting LBD37-like gene by an 136 bp insertion show different distributions between green and purple cabbages (Brassica oleracea var. capitata). Genes Genom. 2021, 43, 679–688. [Google Scholar] [CrossRef]
- He, L.; Tang, R.; Shi, X.; Wang, W.; Cao, Q.; Liu, X.; Jia, X. Biosynthesis related microRNAs and their target genes by small RNA and degradome sequencing in tuberous roots of sweetpotato. BMC Plant. Biol. 2019, 19, 1–19. [Google Scholar] [CrossRef]
- Li, X.; Hou, Y.; Xie, X.; Li, H.; Li, X.; Zhu, Y.; Bian, S. A blueberry MIR156a–SPL12 module coordinates the accumulation of chlorophylls and anthocyanins during fruit ripening. J. Exp. Bot. 2020, 71, 5976–5989. [Google Scholar] [CrossRef] [PubMed]
- Feyissa, B.A.; Arshad, M.; Gruber, M.Y.; Kohalmi, S.E.; Hannoufa, A. The interplay between miR156/SPL13 and DFR/WD40–1 regulate drought tolerance in alfalfa. BMC Plant. Biol. 2019, 19, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Zheng, T.; Tan, W.; Yang, H.; Zhang, L.; Li, T.; Liu, B.; Zhang, D.; Lin, H. Regulation of anthocyanin accumulation via MYB75/HAT1/TPL-mediated transcriptional repression. PLoS Genetics 2019, 15, e1007993. [Google Scholar] [CrossRef]
- Li, C.; Qiu, J.; Yang, G. Isolation and characterization of a R2R3-MYB transcription factor gene related to anthocyanin biosynthesis in the spathes of Anthurium andraeanum (Hort.). Plant. Cell Rep. 2016, 35, 2151–2165. [Google Scholar] [CrossRef]
- Li, C.; Qiu, J.; Ding, L.; Huang, M.; Huang, S.; Yang, G.; Yin, J. Anthocyanin biosynthesis regulation of DhMYB2 and DhbHLH1 in Dendrobium hybrids petals. Plant. Physiol. Biochem. 2017, 112, 335–345. [Google Scholar] [CrossRef]
- Anwar, M.; Wang, G.; Wu, J.; Waheed, S.; Allan, A.C.; Zeng, L. Ectopic overexpression of a novel R2R3-MYB, NtMYB2 from Chinese narcissus represses anthocyanin biosynthesis in tobacco. Molecules 2018, 23, 781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, J.H.; Liu, C.; Xiao, X.; Dixon, R.A. The transcriptional repressor MYB2 regulates both spatial and temporal patterns of proanthocyandin and anthocyanin pigmentation in Medicago truncatula. Plant. Cell 2015, 27, 2860–2879. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.J.; Shen, X.F.; Zhang, Z. The myb transcription factor MdMYB6 suppresses anthocyanin biosynthesis in transgenic Arabidopsis. Plant. Cell Tiss Organ. Cult. 2011, 106, 235–242. [Google Scholar] [CrossRef]
- Xi, W.; Feng, J.; Liu, Y.; Zhang, S.; Zhao, G. The R2R3-MYB transcription factor PaMYB10 is involved in anthocyanin biosynthesis in apricots and determines red blushed skin. BMC Plant. Biol. 2019, 19, 287. [Google Scholar] [CrossRef]
- Ahmed, N.U.; Park, J.-I.; Jung, H.-J.; Hur, Y.; Nou, I.-S. Anthocyanin biosynthesis for cold and freezing stress tolerance and desirable color in Brassica rapa. Funct. Integr. Genomics 2015, 15, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hu, Z.; Zhu, M.; Zhu, Z.; Wang, Z.; Tian, S.; Chen, G. Anthocyanin accumulation and molecular analysis of correlated genes in purple Kohlrabi (Brassica oleracea var. gongylodes L.). J. Agric. Food Chem. 2015, 63, 4160–4169. [Google Scholar] [CrossRef]
- Mushtaq, M.A.; Pan, Q.; Chen, D.; Zhang, Q.; Ge, X.; Li, Z. Comparative leaves transcriptome analysis emphasizing on accumulation of anthocyanins in Brassica: Molecular regulation and potential interaction with photosynthesis. Front. Plant. Sci. 2016, 7, 311. [Google Scholar] [CrossRef] [Green Version]
- Padmaja, L.K.; Agarwal, P.; Gupta, V.; Mukhopadhyay, A.; Sodhi, Y.S.; Pental, D.; Pradhan, A.K. Natural mutations in two homoeologous TT8 genes control yellow seed coat trait in allotetraploid Brassica juncea (AABB). Theor. Appl. Genet. 2014, 127, 339–347. [Google Scholar] [CrossRef]
- Li, X.; Chen, L.; Hong, M.; Zhang, Y.; Zu, F.; Wen, J.; Yi, B.; Ma, C.; Shen, J.; Tu, J. A large insertion in bHLH transcription factor BrTT8 resulting in yellow seed coat in Brassica rapa. PLoS ONE 2012, 7, e44145. [Google Scholar] [CrossRef] [PubMed]
- An, X.H.; Tian, Y.; Chen, K.Q.; Wang, X.F.; Hao, Y.J. The apple WD40 protein MdTTG1 interacts with bHLH but not MYB proteins to regulate anthocyanin accumulation. J. Plant. Physiol. 2012, 169, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Gao, L.; Wang, H. The R2R3-MYB, bHLH, WD40, and related transcription factors in flavonoid biosynthesis. Funct. Integr. Genomics 2013, 13, 75–98. [Google Scholar] [CrossRef]
- Liu, Y.; Hou, H.; Jiang, X.; Wang, P.; Dai, X.; Chen, W.; Xia, T. A WD40 repeat protein from Camellia sinensis regulates anthocyanin and Proanthocyanidin accumulation through the formation of MYB–bHLH–WD40 ternary complexes. Int. J. Mol. Sci. 2018, 19, 1686. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.R.; Davison, P.A.; Bolognesi-Winfield, A.C.; James, C.M.; Srinivasan, N.; Blundell, T.L.; Esch, J.J.; Marks, M.D.; Gray, J.C. The TRANSPARENT TESTA GLABRA1 Locus, which regulates trichome differentiation and anthocyanin biosynthesis in Arabidopsis, encodes a WD40 repeat protein. Plant. Cell 1999, 11, 1337–1349. [Google Scholar] [CrossRef] [Green Version]
- Pang, Y.; Wenger, J.P.; Saathoff, K.; Peel, G.J.; Wen, J.; Huhman, D.; Dixon, R.A. A WD40 repeat protein from Medicago truncatula is necessary for tissue-specific anthocyanin and proanthocyanidin biosynthesis but not for trichome development. Plant. Physiol. 2009, 151, 1114–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaFountain, A.M.; Yuan, Y.W. Repressors of anthocyanin biosynthesis. New Phytol. 2021. [Google Scholar] [CrossRef]
- Yoshida, K.; Ma, D.; Constabel, C.P. The MYB182 protein down-regulates proanthocyanidin and anthocyanin biosynthesis in poplar by repressing both structural and regulatory flavonoid genes. Plant. Physiol. 2015, 167, 693–710. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Wang, N.; Liu, J.; Qu, C.; Wang, Y.; Jiang, S.; Chen, X. The molecular mechanism underlying anthocyanin metabolism in apple using the MdMYB16 and MdbHLH33 genes. Plant. Mol. Biol. 2017, 94, 149–165. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.M.; Zhang, S.; Yang, Q.S.; Gao, H.J.; Sheng, O.; Bi, F.C.; Li, C.Y.; Dong, T.; Yi, G.J.; He, W.D.; et al. MaMYB4, an R2R3-MYB Repressor Transcription Factor, Negatively Regulates the Biosynthesis of Anthocyanin in Banana. Front. Plant. Sci. 2021, 11, 600704. [Google Scholar] [CrossRef]
- Jin, H.; Cominelli, E.; Bailey, P.; Parr, A.; Mehrtens, F.; Jones, J.; Martin, C. Transcriptional repression by AtMYB4 controls production of UV-protecting sunscreens in Arabidopsis. EMBO J. 2000, 19, 6150–6161. [Google Scholar] [CrossRef] [Green Version]
- Albert, N.W.; Davies, K.M.; Lewis, D.H.; Zhang, H.; Montefiori, M.; Brendolise, C.; Boase, M.R.; Ngo, H.; Jameson, P.E.; Schwinn, K.E. A conserved network of transcriptional activators and repressors regulates anthocyanin pigmentation in eudicots. Plant. Cell 2014, 26, 962–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cominelli, E.; Galbiati, M.; Vavasseur, A.; Conti, L.; Sala, T.; Vuylsteke, M.; Tonelli, C. A guard-cell-specific MYB transcription factor regulates stomatal movements and plant drought tolerance. Curr. Biol. 2005, 15, 1196–1200. [Google Scholar] [CrossRef] [PubMed]
- De la Guardia, A.R.H.; Ugalde, M.B.; Lobos-Diaz, V.; Romero-Romero, J.L.; Meyer-Regueiro, C.; Inostroza-Blancheteau, C.; Arce-Johnson, P. Isolation and molecular characterization of MYB60 in Solanum lycopersicum. Mol. Biol. Rep. 2021, 48, 1579–1587. [Google Scholar] [CrossRef] [PubMed]
- Galbiati, M.; Matus, J.T.; Francia, P.; Rusconi, F.; Cañón, P.; Medina, C.; Arce-Johnson, P. The grapevine guard cell-related VvMYB60 transcription factor is involved in the regulation of stomatal activity and is differentially expressed in response to ABA and osmotic stress. BMC Plant. Biol. 2011, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Hu, B.; Qin, Y.; Hu, G.; Zhao, J. Advance of the negative regulation of anthocyanin biosynthesis by MYB transcription factors. Plant. Physiol. Biochem. 2019, 136, 178–187. [Google Scholar] [CrossRef]
- Li, S. Transcriptional control of flavonoid biosynthesis: Fine-tuning of the MYB-bHLH-WD40 (MBW) complex. Plant. Signal. Behav. 2014, 9, e27522. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.P.; Xing, H.L.; Dong, L. Egg cell-specific promoter-controlled CRISPR/Cas9 efficiently generates homozygous mutants for multiple target genes in Arabidopsis in a single generation. Genome Biol. 2015, 16, 144. [Google Scholar] [CrossRef] [Green Version]
- Song, B.; Tang, Z.; Li, X.; Li, J.; Zhang, M.; Zhao, K.; Wu, J. Mining and evolution analysis of lateral organ boundaries domain (LBD) genes in Chinese white pear (Pyrus bretschneideri). BMC Genomics 2020, 21, 1–17. [Google Scholar] [CrossRef]
- Guan, X.Y.; Li, Q.J.; Shan, C.M.; Wang, S.; Mao, Y.B.; Wang, L.J.; Chen, X.Y. The HD-Zip IV gene GaHOX1 from cotton is a functional homologue of the Arabidopsis GLABRA2. Physiol. Plant. 2008, 134, 174–182. [Google Scholar] [CrossRef]
- Zhang, D.; Ye, H.; Guo, H.; Johnson, A.; Zhang, M.; Lin, H. Transcription factor HAT1 is phosphorylated by BIN2 kinase and mediates brassinosteroid repressed gene expression in Arabidopsis. Plant. J. 2014, 77, 59–70. [Google Scholar] [CrossRef]
- Tan, W.; Zhang, D.; Zhou, H.; ZhengTing, Y.Y.; Lin, H. Transcription factor HAT1 is a substrate of SnRK2.3 Kinase and negatively regulates ABA synthesis and signaling in Arabidopsis responding to drought. PLoS Genetics 2018, 14, e1007336. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lian, W.; Cao, Y.; Wang, X.; Wang, G.; Qi, C.; Liu, L.; Qin, S.; Yuan, X.; Li, X.; et al. Overexpression of BoNAC019, a NAC transcription factor from Brassica oleracea, negatively regulates the dehydration response and anthocyanin biosynthesis in Arabidopsis. Sci. Rep. 2018, 8, 13349. [Google Scholar] [CrossRef] [Green Version]
- Morishita, T.; Kojima, Y.; Maruta, T.; Nishizawa-Yokoi, A.; Yabuta, Y.; Shigeoka, S. Arabidopsis NAC Transcription Factor, ANAC078, Regulates Flavonoid Biosynthesis under High-light. Plant. Cell Physiol. 2009, 50, 2210–2222. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, K.; Xu, Z.; El-Kereamy, A.; Casaretto, J.A.; Rothstein, S.J. The Arabidopsis Transcription Factor ANAC032 Represses Anthocyanin Biosynthesis in Response to High Sucrose and Oxidative and Abiotic Stresses. Front. Plant. Sci 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Jiang, S.; Zhang, T.; Xu, H.; Fang, H.; Zhang, J.; Chen, X. Apple NAC transcription factor MdNAC52 regulates biosynthesis of anthocyanin and proanthocyanidin through MdMYB9 and MdMYB11. Plant. Sci. 2019, 289, 110286. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Allu, A.D.; Garapati, P.; Siddiqui, H.; Dortay, H.; Zanor, M.I.; Asensi-Fabado, M.A.; Munné-Bosch, S.; Antonio, C.; Tohge, T.; et al. JUNGBRUNNEN1, a reactive oxygen species–responsive NAC transcription factor, regulates longevity in Arabidopsis. Plant. Cell 2012, 24, 482–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alshareef, N.O.; Wang, J.Y.; Ali, S.; Al-Babili, S.; Tester, M.; Schmöckel, S.M. Overexpression of the NAC transcription factor JUNGBRUNNEN1 (JUB1) increases salinity tolerance in tomato. Plant. Physiol. Biochem. 2019, 140, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Peragine, A.; Yoshikawa, M.; Wu, G.; Albrecht, H.L.; Poethig, R.S. SGS3 and SGS2/SDE1/RDR6 are required for juvenile development and the production of trans-acting siRNAs in Arabidopsis. Genes Dev. 2004, 18, 2368–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez, F.; Vaucheret, H.; Rajagopalan, R.; Lepers, C.; Gasciolli, V.; Mallory, A.C.; Hilbert, J.L.; Bartel, D.P.; Crete, P. Endogenous trans-acting siRNAs regulate the accumulation of Arabidopsis mRNAs. Mol. Cell 2004, 16, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.; Xie, Z.; Gustafson, A.M.; Carrington, J.C. MicroRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 2005, 121, 207–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Yang, H.J.; Yang, Y.Z.; Zhu, Z.Z.; Li, Y.N.; Qu, D.; Zhao, Z.Y. mdm-miR828 participates in the feedback loop to regulate anthocyanin accumulation in apple peel. Front. Plant. Sci. 2020, 11, 608109. [Google Scholar] [CrossRef]
- Yamagishi, M.; Sakai, M. The MicroRNA828/MYB12 Module Mediates Bicolor Pattern Development in Asiatic Hybrid Lily (Lilium spp.) Flowers. Front. Plant. Sci. 2020, 11, 590791. [Google Scholar] [CrossRef]
- Bonar, N.; Liney, M.; Zhang, R.; Austin, C.; Dessoly, J.; Davidson, D.; Hornyik, C. Potato miR828 is associated with purple tuber skin and flesh color. Front. Plant. Sci. 2018, 9, 1742. [Google Scholar] [CrossRef] [Green Version]
- Tirumalai, V.; Swetha, C.; Nair, A.; Pandit, A.; Shivaprasad, P.V. miR828 and miR858 regulate VvMYB114 to promote anthocyanin and flavonol accumulation in grapes. J. Exp. Bot. 2019, 70, 4775–4792. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Singh, R.K.; Navarre, D.A. R2R3-MYB transcription factors, StmiR858 and sucrose mediate potato flavonol biosynthesis. Hort. Res. 2021, 8, 1–13. [Google Scholar] [CrossRef]
- Piya, S.; Kihm, C.; Rice, J.H.; Baum, T.J.; Hewezi, T. Cooperative regulatory functions of miR858 and MYB83 during Cyst nematode parasitism. Plant. Physiol. 2017, 174, 1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, D.; Tiwari, M.; Pandey, A.; Bhatia, C.; Sharma, A.; Trivedi, P.K. MicroRNA858 is a potential regulator of phenylpropanoid pathway and plant development. Plant. Physiol. 2016, 171, 944–959. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, Y.; Song, Z.; Zhang, H. Repression of MYBL2 by both microRNA858a and HY5 leads to the activation of anthocyanin biosynthetic pathway in Arabidopsis. Mol. Plant. 2016, 9, 1395–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Cui, W.; Qi, X.; Lin, M.; Qiao, C.; Zhong, Y.; Fang, J. MicroRNA858 negatively regulates anthocyanin biosynthesis by repressing AaMYBC1 expression in kiwifruit (Actinidia arguta). Plant. Sci. 2020, 296, 110476. [Google Scholar] [CrossRef]
- Zhou, G.K.; Kubo, M.; Zhong, R.; Demura, T.; Ye, Z.H. Overexpression of miR165 affects apical meristem formation, organ polarity establishment and vascular development in Arabidopsis. Plant. Cell Physiol. 2007, 48, 391–404. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Qiu, Y.; Duan, M.; Wang, J.; Zhang, X.; Wang, H.; Li, X. Identification of anthocyanin biosynthesis related microRNAs in a distinctive Chinese radish (Raphanus sativus L.) by high-throughput sequencing. Mol. Genetics Genom. 2017, 292, 215–229. [Google Scholar] [CrossRef]
- Gou, J.Y.; Felippes, F.F.; Liu, C.J.; Weigel, D.; Wang, J.W. Negative regulation of anthocyanin biosynthesis in Arabidopsis by a miR156-targeted SPL transcription factor. Plant. Cell 2011, 23, 1512–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukusaki, E.; Kawasaki, K.; Kajiyama, S.; An, C.-I.; Suzuki, K.; Tanaka, Y.; Kobayashi, A. Flower color modulations of Torenia hybrida by downregulation of chalcone synthase genes with RNA interference. J. Biotechnol. 2004, 111, 229–240. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Gene | Organism | Function | Method of Genome Editing | Editing Events |
|---|---|---|---|---|
| PAP1 | A. thaliana | MYB transcriptional activator of anthocyanin biosynthesis | activation | no changes in leaf color [29]; purple pigmentation of the leaves [12] |
| ANT1 | Solanum lycopersicum | MYB transcriptional activator of anthocyanin biosynthesis | knock-in | dark purple plants [16,33] |
| AN2 | S. lycopersicum | MYB transcriptional activator of anthocyanin biosynthesis | knockout | reduced anthocyanin content in tomato fruits [65]; decreased anthocyanin content in vegetative tissues [66] |
| MYB113-like | Daucus carota | MYB transcriptional activator of anthocyanin biosynthesis | knockout | depigmentation [67] |
| MYBA7 (PAP2) | Vitis vinifera | MYB transcriptional activator of anthocyanin biosynthesis | knockout | change in color were not evaluated [59] |
| TT8 | B. napus, N. tabacum | bHLH transcriptional activatior of anthocyanin biosynthesis | knockout | depigmentation of seeds, elevated seed oil and protein content and altered fatty acid composition [68]; elevated protein and lipid content, depigmentation of seeds and flowers [69] |
| TTG1 | A. thaliana, Oryza sativa | WD40 transcriptional activatior of anthocyanin biosynthesis | knockout | absence of trichomes and pale seeds [70], decreased anthocyanin accumulation in various organs, reduction of trichomes in grains [71] |
| MYB57 | P. nyctaginiflora | MYB transcriptional repressor of anthocyanin biosynthesis | knockout | high anthocyanin and proanthocyanidin content [72] |
| GLABRA2 | A. thaliana | homeodomain-leucine zipper repressor of anthocyanin biosynthesis | knockout | trichome-less phenotype [8,73]; anthocyanin accumulation and increased seed oil content in natural mutants [74,75] |
| DFR | Ipomoea nil, O. sativa, S. lycopersicum, A. thaliana, N. benthamiana, Zea mays | structural gene | knockout, activation, knock-in | reduced anthocaynin pigmentation [76,77,78]; knock-in of the deleted part of DFR gene restored gene function [78]; activation of DFR gene increased its expression level 400-10000 fold [79]; knockout was successful, but phenotypic effect was not evaluated [80] |
| UGT79B2 and UGT79B3 | A. thaliana | structural gene | knockout | mutants were more susceptible to stress and had reduced anthocyanin content [81] |
| 5GT, 3′GT, 3′AT | Gentiana triflora × Gentiana scabra | structural gene | knockout | change in flower color shade due to predominance of different delphinidin derivatives [47] |
| F3′H | Euphorbia pulcherrima, O. sativa | structural gene | knockout | bract color changed from red to yellow [82], depigmented seeds and reduced anthocyanin content [77]. |
| F3H | Torenia fournieri, D. carota, S. lycopersicum | structural gene | knockout | depigmentation of flowers [83], cells [84] and hypocotyls [85]. |
| Gene | Organism | Function | Method used to Verify Gene Function | Phenotypic Effect |
|---|---|---|---|---|
| MYB10 | A. thaliana, Gerbera hybrida, Fragaria vesca F.×ananassa | MYB transcriptional activator of anthocyanin biosynthesis | generation of transgenic plants, SNP analysis | increased anthocyanin pigmentation in undifferentiated callus, petioles, leaf veins and flower scapes [113] and immature seeds [114]. Transposon insertions in the promoter region changed the fruit color [115]. An insertion in the coding region resulted in white fruit color [116]. An amino acid change caused the loss of fruit pigmentation [117]. |
| MYB1 | Allium cepa | MYB transcriptional activator of anthocyanin biosynthesis | RNA interference | anthocyanin pigmentation decreased [100]. |
| MYB1 | F.×ananassa | MYB transcriptional repressor of anthocyanin biosynthesis | RNA interference, generation of transgenic plants | down-regulation of this gene promoted an increase in anthocyanin content [107]; accumulation of anthocyanins and flavonols in transgenic tobacco was suppressed [103]. |
| MYB2 | B. rapa, B. oleracea, B. napus A. graveolens | MYB transcriptional activator of anthocyanin biosynthesis | generation of transgenic plants, sequencing | increased anthocyanin pigmentation [118,119,120,121]; mutation in promoter region of BnaPAP2.A7 resulted in purple pigmentation in B. napus [122]. |
| MYB6 | D. carota | MYB transcriptional activator of anthocyanin biosynthesis | generation of transgenic plants | enhanced anthocyanin accumulation in vegetative and reproductive tissues [123]. |
| Delila | Antirrhinum majus | bHLH transcriptional activatior of anthocyanin biosynthesis | generation of transgenic plants | enhanced anthocyanin production in leaves and flowers, salt and drought tolerance [124]; anthocyanin accumulation in the taproots [125] leaves [126]. |
| MYBL2 | B. oleracea, A. thaliana, I. loxense | MYB transcriptional repressor of anthocyanin biosynthesis | Sequence analysis of naturally purple plants, silencing, generation of transgenic plants | deletion of coding sequences or substitution in the promoter region were responsible for purple coloration [127]; loss of function promoted an increase in the anthocyanin content [50,51] and prevented trichome initiation [128]; expression of IlMYBL1 gene in tobacco resulted in depigmentation [129]. |
| CPC | A. thaliana | MYB transcriptional repressor of anthocyanin biosynthesis | generation of transgenic plants, loss of function mutants | positive regulator of root hair formation and negative regulator of trichome formation and anthocyanin accumulation [52,130,131]. |
| LBD37, LBD38, and LBD39 | A. thaliana, B. oleracea, Malus domestica, Pyrus pyrifolia, S. tuberosum | LATERAL ORGAN BOUNDARY DOMAIN repressors of anthocyanin biosynthesis | generation of transgenic plants, loss of function mutants, expression profiling | increased accumulation of anthocyanins in loss of function mutants [53,127]; repression of anthocyanin biosynthesis due to overexpression of LBD gene [132,133,134,135]; In purple cabbage pigmentation depended on an insertion in LBD37 gene [136]. |
| miR156 | A. thaliana, Ipomoea batatas, Populus alba ×P. tremula var. glandulosa, Vaccinium corymbosum, S. lycopersicum. Medicago sativa L. | MicroRNA activator of anthocyanin biosynthesis | generation of transgenic plants | Increased expression of miR156 promoted accumulation of anthocyanins [116,137,138,139]; reduction of miR156 activity promoted synthesis of flavonols [137]; miR156 positively regulated drought tolerance and photosynthetic efficiency [139]. |
| HAT1 | A. thaliana | homeodomain-leucine zipper repressor of anthocyanin biosynthesis | generation of transgenic plants | drought tolerance and anthocyanin accumulation increased in loss-of-function mutants and decreased in transgenic plants overexpressing HAT1 gene [140] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khusnutdinov, E.; Sukhareva, A.; Panfilova, M.; Mikhaylova, E. Anthocyanin Biosynthesis Genes as Model Genes for Genome Editing in Plants. Int. J. Mol. Sci. 2021, 22, 8752. https://doi.org/10.3390/ijms22168752
Khusnutdinov E, Sukhareva A, Panfilova M, Mikhaylova E. Anthocyanin Biosynthesis Genes as Model Genes for Genome Editing in Plants. International Journal of Molecular Sciences. 2021; 22(16):8752. https://doi.org/10.3390/ijms22168752
Chicago/Turabian StyleKhusnutdinov, Emil, Anna Sukhareva, Maria Panfilova, and Elena Mikhaylova. 2021. "Anthocyanin Biosynthesis Genes as Model Genes for Genome Editing in Plants" International Journal of Molecular Sciences 22, no. 16: 8752. https://doi.org/10.3390/ijms22168752
APA StyleKhusnutdinov, E., Sukhareva, A., Panfilova, M., & Mikhaylova, E. (2021). Anthocyanin Biosynthesis Genes as Model Genes for Genome Editing in Plants. International Journal of Molecular Sciences, 22(16), 8752. https://doi.org/10.3390/ijms22168752