
Expression and Regulation of CD73 during the Estrous Cycle in Mouse Uterus



Abstract
:1. Introduction
2. Results
2.1. Expression of CD73 in the Mouse Uterus during Estrous Cycle
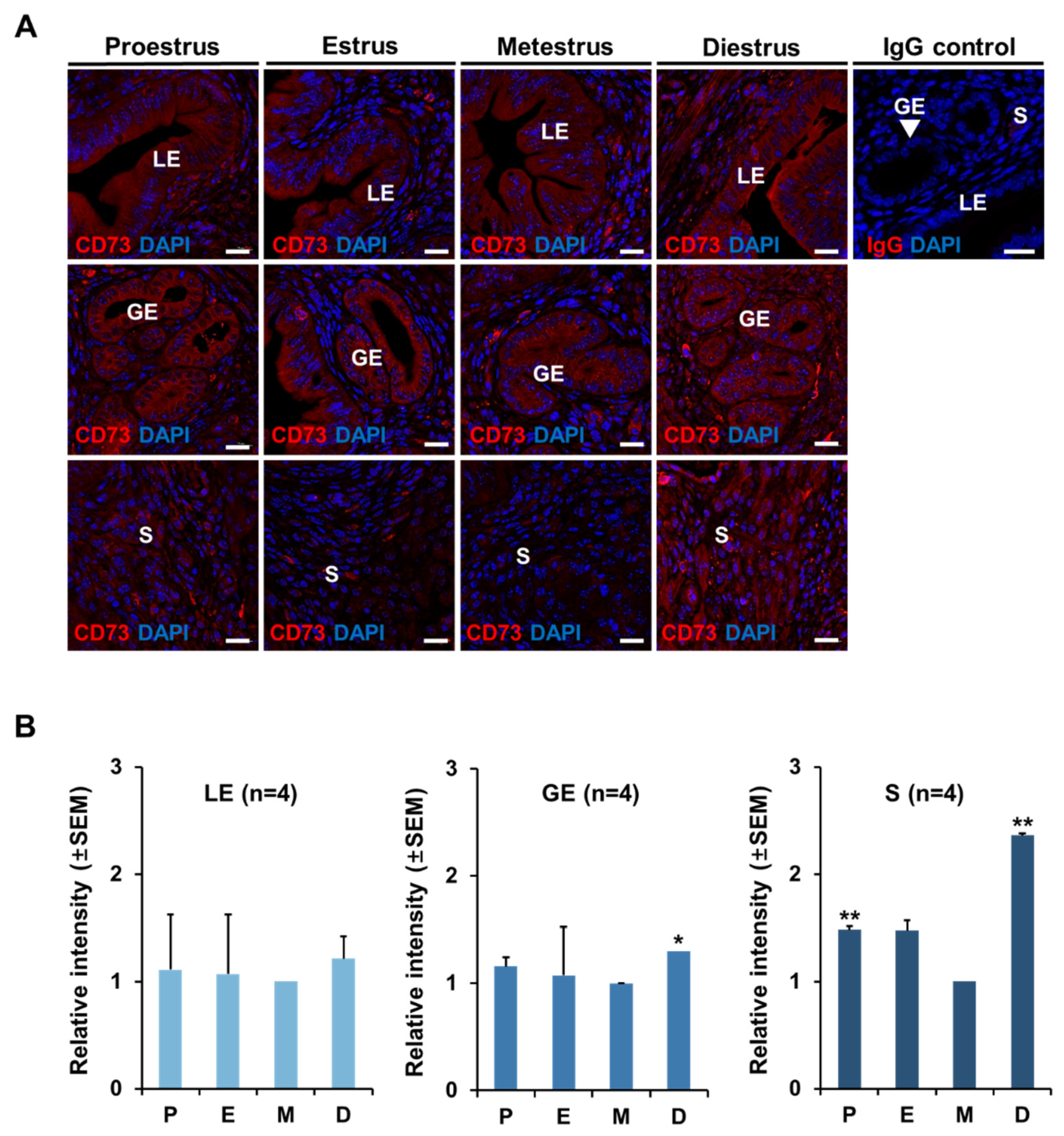
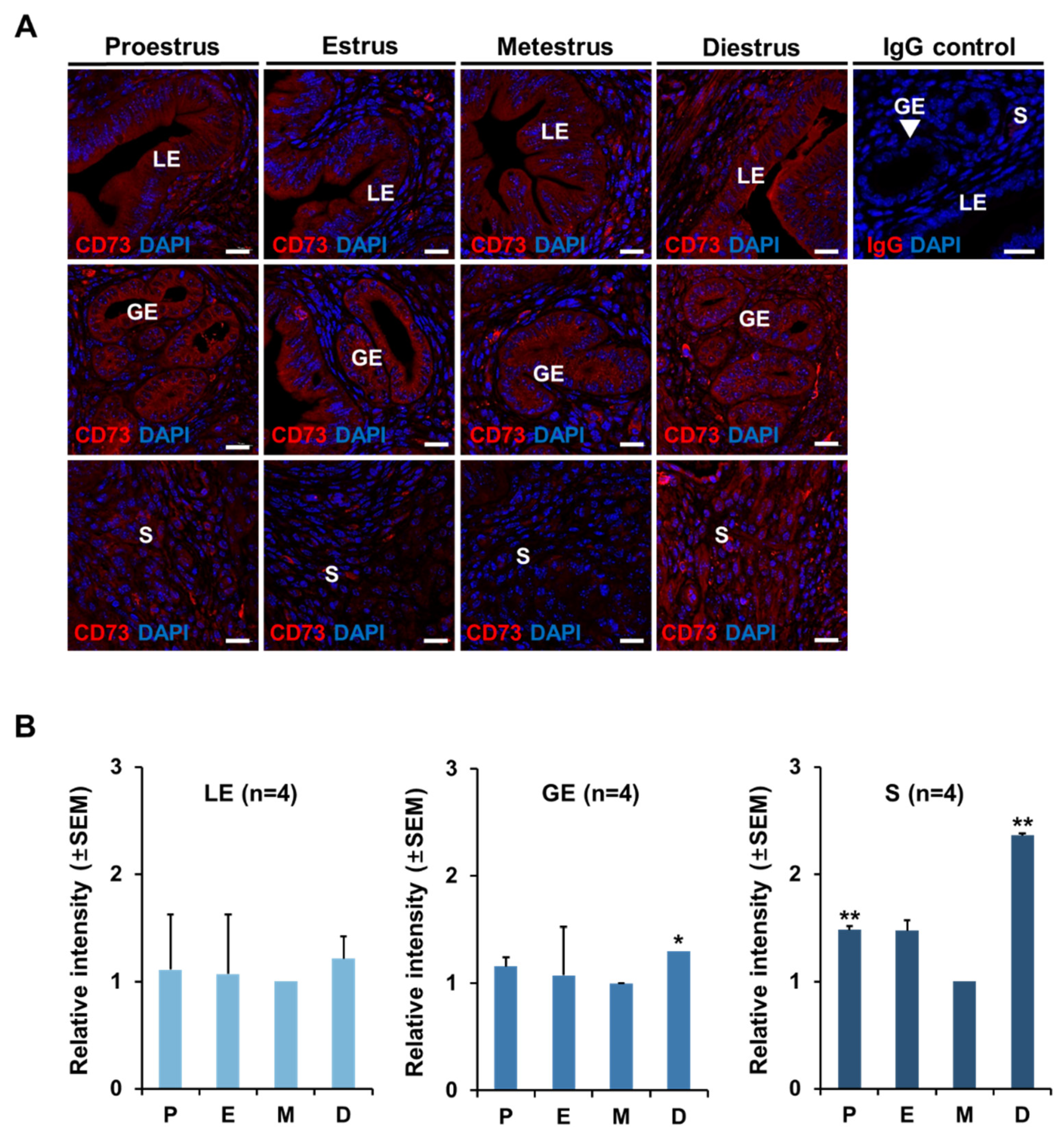
2.2. The Localization of CD73 in Mouse Uterus at Each Stage of Estrous Cycle
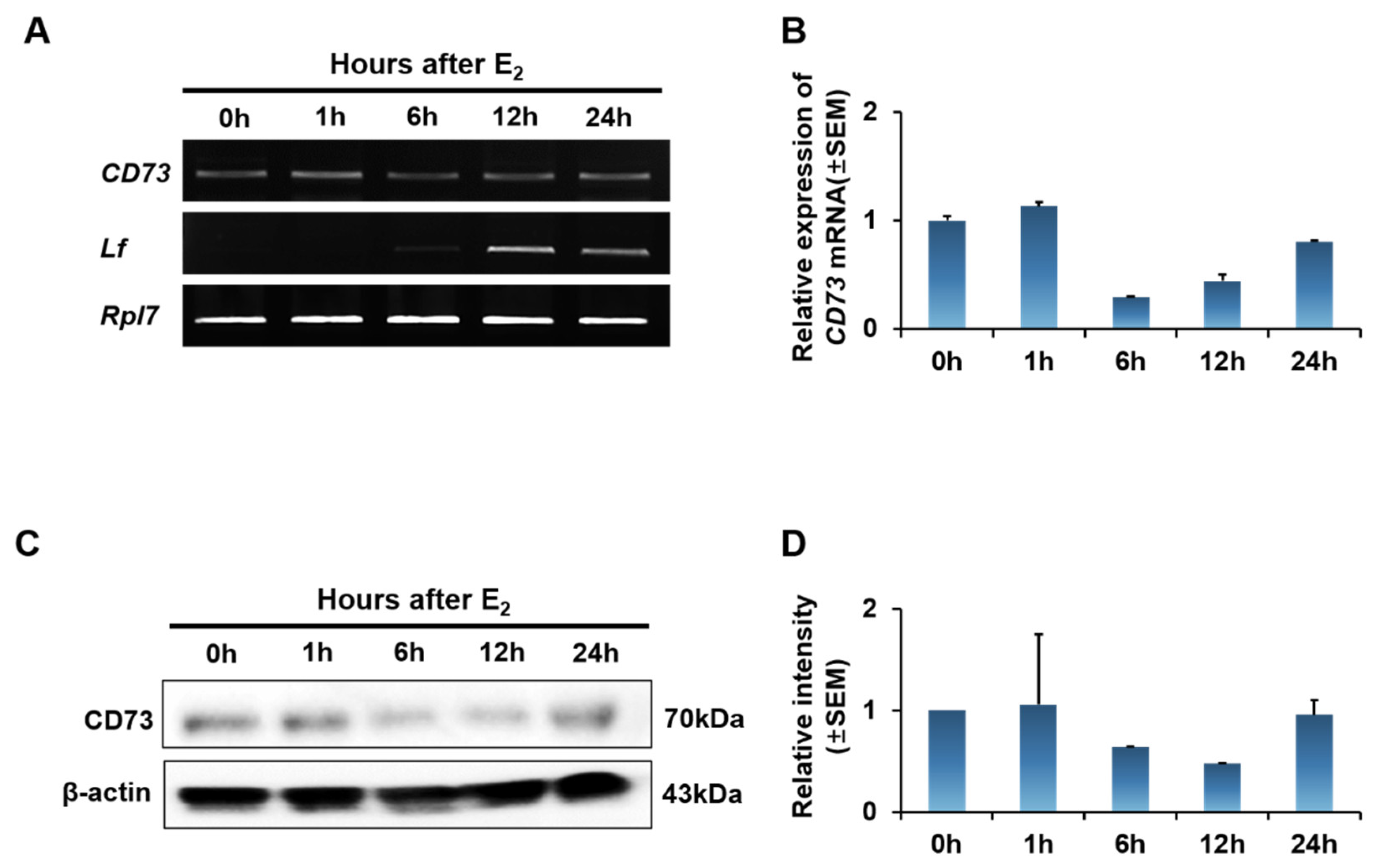
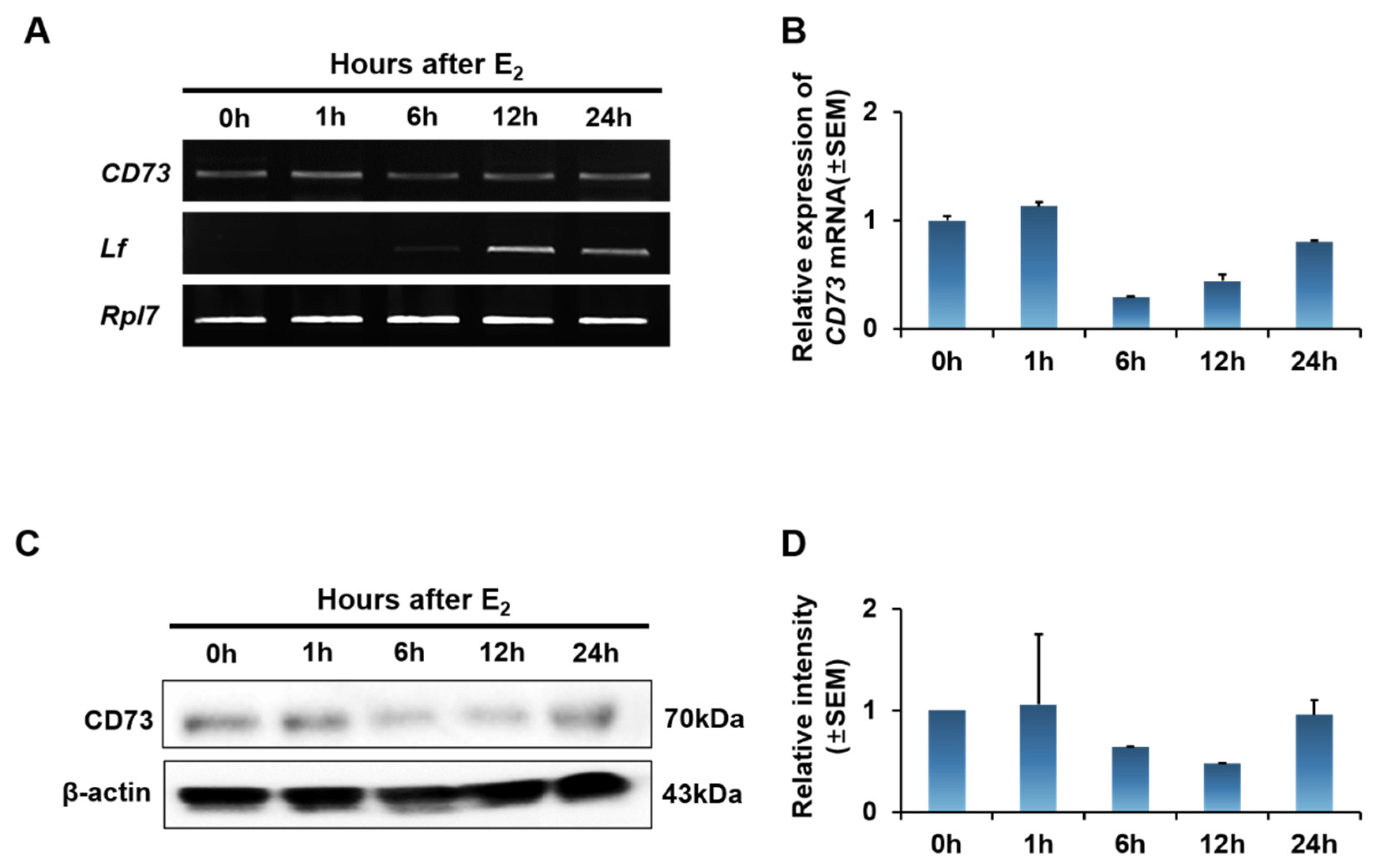
2.3. Estrogen Effect on CD73 Expression in Ovariectomized (OVX) Mouse Uterus
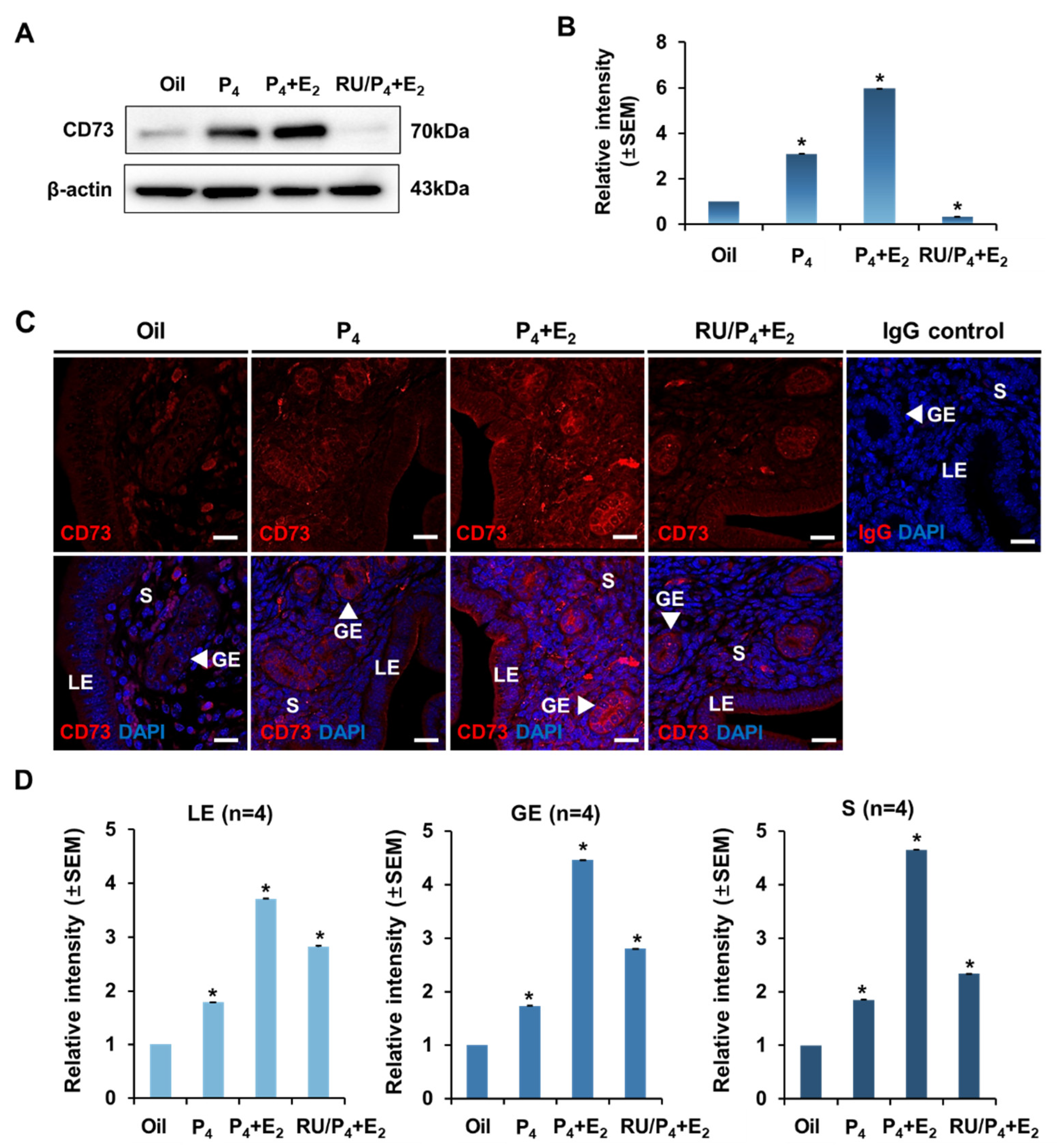
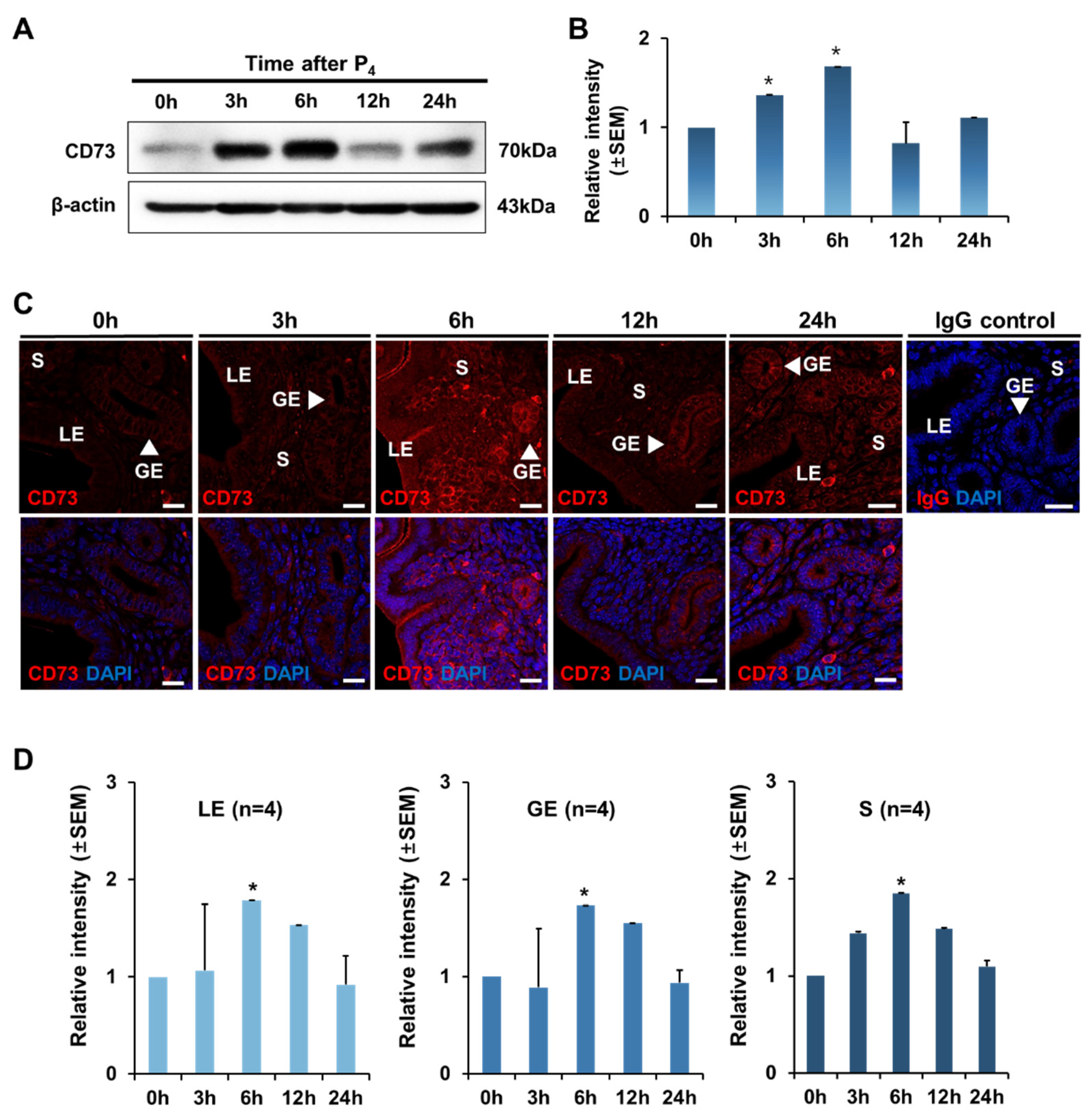
2.4. Regulation and Localization of CD73 Expression by Progesterone
2.5. Effect of Estrogen and Progesterone on the Expression of CD73
3. Discussion
4. Materials and Methods
4.1. Animal Care and Experimentation
4.2. Confirmation of Estrous Cycle and Uterus Sampling
4.3. Ovariectomy and Hormone Injection
4.4. RNA Preparation, Reverse Transcription PCR (RT-PCR), and Quantitative Real-Time PCR (qRT-PCR)
4.5. Protein Preparation and Western Blot Analysis
4.6. Immunofluorescence
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kobayashi, A.; Behringer, R.R. Developmental genetics of the female reproductive tract in mammals. Nat. Rev. Genet. 2003, 4, 969–980. [Google Scholar] [CrossRef]
- Mihm, M.; Gangooly, S.; Muttukrishna, S. The normal menstrual cycle in women. Anim. Reprod. Sci. 2011, 124, 229–236. [Google Scholar] [CrossRef]
- Bertolin, K.; Murphy, B.D. Reproductive tract changes during the mouse estrous cycle. In The Guide to Investigation of Mouse Pregnancy; Elsevier: Amsterdam, The Netherlands, 2014; pp. 85–94. [Google Scholar]
- Byers, S.L.; Wiles, M.V.; Dunn, S.L.; Taft, R.A. Mouse estrous cycle identification tool and images. PLoS ONE 2012, 7, e35538. [Google Scholar] [CrossRef] [Green Version]
- Caligioni, C.S. Assessing reproductive status/stages in mice. Curr. Protoc. Neurosci. 2009, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gargett, C.E.; Nguyen, H.P.; Ye, L. Endometrial regeneration and endometrial stem/progenitor cells. Rev. Endocr. Metab. Disord. 2012, 13, 235–251. [Google Scholar] [CrossRef]
- Sträter, N. Ecto-5′-nucleotidase: Structure function relationships. Purinergic Signal. 2006, 2, 343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adzic, M.; Nedeljkovic, N. Unveiling the role of ecto-5′-nucleotidase/CD73 in astrocyte migration by using pharmacological tools. Front. Pharmacol. 2018, 9, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, J. La nucleotidase et sa reaction avec les desaminations des nucleotides dans le coeur et dans le muscle. Bull. Soc. Chim. Biol. 1934, 16, 385–399. [Google Scholar]
- Zimmermann, H. 5′-Nucleotidase: Molecular structure and functional aspects. Biochem. J. 1992, 285, 345–365. [Google Scholar] [CrossRef] [Green Version]
- Trapero, C.; Vidal, A.; Fernández-Montolí, M.E.; Coroleu, B.; Tresserra, F.; Barri, P.; Gómez de Aranda, I.; Sévigny, J.; Ponce, J.; Matias-Guiu, X. Impaired Expression of Ectonucleotidases in Ectopic and Eutopic Endometrial Tissue Is in Favor of ATP Accumulation in the Tissue Microenvironment in Endometriosis. Int. J. Mol. Sci. 2019, 20, 5532. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.R.; Oh, H.K.; Park, S.H.; Jeong, Y.J. Expression of CD73 is associated with tumor progression and intratumoral inflammation in breast cancer. Asia-Pac. J. Clin. Oncol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Buisseret, L.; Pommey, S.; Allard, B.; Garaud, S.; Bergeron, M.; Cousineau, I.; Ameye, L.; Bareche, Y.; Paesmans, M.; Crown, J. Clinical significance of CD73 in triple-negative breast cancer: Multiplex analysis of a phase III clinical trial. Ann. Oncol. 2018, 29, 1056–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B. CD73: A novel target for cancer immunotherapy. Cancer Res. 2010, 70, 6407–6411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minor, M.; Alcedo, K.P.; Battaglia, R.A.; Snider, N.T. Cell type-and tissue-specific functions of ecto-5′-nucleotidase (CD73). Am. J. Physiol.-Cell Physiol. 2019, 317, C1079–C1092. [Google Scholar] [CrossRef]
- Augusto, E.; Matos, M.; Sévigny, J.; El-Tayeb, A.; Bynoe, M.S.; Müller, C.E.; Cunha, R.A.; Chen, J.-F. Ecto-5′-nucleotidase (CD73)-mediated formation of adenosine is critical for the striatal adenosine A2A receptor functions. J. Neurosci. 2013, 33, 11390–11399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulesskaya, N.; Võikar, V.; Peltola, M.; Yegutkin, G.G.; Salmi, M.; Jalkanen, S.; Rauvala, H. CD73 is a major regulator of adenosinergic signalling in mouse brain. PLoS ONE 2013, 8, e66896. [Google Scholar] [CrossRef] [PubMed]
- Shirley, D.G.; Vekaria, R.M.; Sévigny, J. Ectonucleotidases in the kidney. Purinergic Signal. 2009, 5, 501. [Google Scholar] [CrossRef] [Green Version]
- Jackson, E.K.; Cheng, D.; Verrier, J.D.; Janesko-Feldman, K.; Kochanek, P.M. Interactive roles of CD73 and tissue nonspecific alkaline phosphatase in the renal vascular metabolism of 5′-AMP. Am. J. Physiol.-Ren. Physiol. 2014, 307, F680–F685. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, S.; Eto, S.; Kato, K.; Tashiro, Y. Ferritin immunoelectron microscopic localization of 5’-nucleotidase on rat liver cell surface. J. Cell Biol. 1984, 99, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Peng, Z.; Fernandez, P.; Wilder, T.; Yee, H.; Chiriboga, L.; Chan, E.S.; Cronstein, B.N. Ecto-5′-nucleotidase (CD73)-mediated extracellular adenosine production plays a critical role in hepatic fibrosis. FASEB J. 2008, 22, 2263–2272. [Google Scholar] [CrossRef]
- Eckle, T.; Füllbier, L.; Wehrmann, M.; Khoury, J.; Mittelbronn, M.; Ibla, J.; Rosenberger, P.; Eltzschig, H.K. Identification of ectonucleotidases CD39 and CD73 in innate protection during acute lung injury. J. Immunol. 2007, 178, 8127–8137. [Google Scholar] [CrossRef]
- Li, H.; Karmouty-Quintana, H.; Chen, N.-Y.; Mills, T.; Molina, J.; Blackburn, M.R.; Davies, J. Loss of CD73-mediated extracellular adenosine production exacerbates inflammation and abnormal alveolar development in newborn mice exposed to prolonged hyperoxia. Pediatr. Res. 2017, 82, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Eckle, T.; Krahn, T.; Grenz, A.; Kohler, D.; Mittelbronn, M.; Ledent, C.; Jacobson, M.A.; Osswald, H.; Thompson, L.F.; Unertl, K. Cardioprotection by ecto-5’-nucleotidase (CD73) and A2B adenosine receptors. Circulation 2007, 115, 1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, X.T.; Taniai, S.; Zhao, G.; Huang, C.X.; Velenosi, T.J.; Xue, J.; Urquhart, B.L.; Karmazyn, M. CD73-TNAP crosstalk regulates the hypertrophic response and cardiomyocyte calcification due to α 1 adrenoceptor activation. Mol. Cell. Biochem. 2014, 394, 237–246. [Google Scholar] [CrossRef]
- Antonioli, L.; Blandizzi, C.; Pacher, P.; Haskó, G. Immunity, inflammation and cancer: A leading role for adenosine. Nat. Rev. Cancer 2013, 13, 842–857. [Google Scholar] [CrossRef] [PubMed]
- Young, A.; Mittal, D.; Stagg, J.; Smyth, M.J. Targeting cancer-derived adenosine: New therapeutic approaches. Cancer Discov. 2014, 4, 879–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalekar, L.A.; Mueller, D.L. Relationship between CD4 regulatory T cells and anergy in vivo. J. Immunol. 2017, 198, 2527–2533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kordaß, T.; Osen, W.; Eichmüller, S.B. Controlling the immune suppressor: Transcription factors and microRNAs regulating CD73/NT5E. Front. Immunol. 2018, 9, 813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neo, S.Y.; Yang, Y.; Record, J.; Ma, R.; Chen, X.; Chen, Z.; Tobin, N.P.; Blake, E.; Seitz, C.; Thomas, R.; et al. CD73 immune checkpoint defines regulatory NK cells within the tumor microenvironment. J. Clin. Investig. 2020, 130, 1185–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Ham, A.; Kim, J.Y.; Brown, K.M.; D’Agati, V.D.; Lee, H.T. The volatile anesthetic isoflurane induces ecto-5’-nucleotidase (CD73) to protect against renal ischemia and reperfusion injury. Kidney Int. 2013, 84, 90–103. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Alter, C.; Ding, Z.; Borg, N.; Schrader, J. Adenosine Formed by CD73 on T Cells Inhibits Cardiac Inflammation and Fibrosis and Preserves Contractile Function in Transverse Aortic Constriction-Induced Heart Failure. Circ. Heart Fail. 2017, 10. [Google Scholar] [CrossRef]
- Jia, W.Q.; Zhou, T.C.; Dai, J.W.; Liu, Z.N.; Zhang, Y.F.; Zang, D.D.; Lv, X.W. CD73 regulates hepatic stellate cells activation and proliferation through Wnt/beta-catenin signaling pathway. Eur. J. Pharmacol. 2021, 890, 173667. [Google Scholar] [CrossRef]
- Tepekoy, F.; Akkoyunlu, G.; Demir, R. The role of Wnt signaling members in the uterus and embryo during pre-implantation and implantation. J. Assist. Reprod. Genet. 2015, 32, 337–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnstock, G. Purinergic signalling in the reproductive system in health and disease. Purinergic Signal. 2014, 10, 157–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackburn, M.R.; Gao, X.; Airhart, M.J.; Skalko, R.G.; Thompson, L.F.; Knudsen, T.B. Adenosine levels in the postimplantation mouse uterus: Quantitation by HPLC-fluorometric detection and spatiotemporal regulation by 5′-nucleotidase and adenosine deaminase. Dev. Dyn. 1992, 194, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Gillman, T.A.; Pennefather, J.N. Evidence for the presence of both P1 and P2 purinoceptors in the rat myometrium. Clin. Exp. Pharmacol. Physiol. 1998, 25, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Aliagas, E.; Vidal, A.; Torrejón-Escribano, B.; del Rosario Taco, M.; Ponce, J.; de Aranda, I.G.; Sévigny, J.; Condom, E.; Martín-Satué, M. Ecto-nucleotidases distribution in human cyclic and postmenopausic endometrium. Purinergic Signal. 2013, 9, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Texidó, L.; Romero, C.; Vidal, A.; García-Valero, J.; Fernandez Montoli, M.E.; Baixeras, N.; Condom, E.; Ponce, J.; García-Tejedor, A.; Martín-Satué, M. Ecto-nucleotidases activities in the contents of ovarian endometriomas: Potential biomarkers of endometriosis. Mediat. Inflamm. 2014, 2014. [Google Scholar] [CrossRef]
- Gupta, R.C. Veterinary Toxicology: Basic and Clinical Principles; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Jeong, J.-H.; Park, M.; Park, M.; Lim, E.J.; Kim, H.-R.; Song, H.; Park, S.G.; Choi, E.-J.; Hong, K.-H.; Lee, D.R. The expression of aminoacyl-tRNA-synthetase-interacting multifunctional protein-1 (Aimp1) is regulated by estrogen in the mouse uterus. Mol. Cell. Endocrinol. 2015, 399, 78–86. [Google Scholar] [CrossRef]
- Kim, H.-R.; Cho, K.-S.; Kim, E.; Lee, O.-H.; Yoon, H.; Lee, S.; Moon, S.; Park, M.; Hong, K.; Na, Y. Rapid expression of RASD1 is regulated by estrogen receptor-dependent intracellular signaling pathway in the mouse uterus. Mol. Cell. Endocrinol. 2017, 446, 32–39. [Google Scholar] [CrossRef]
- Paria, B.C.; Lim, H.; Wang, X.N.; Liehr, J.; Das, S.K.; Dey, S.K. Coordination of differential effects of primary estrogen and catecholestrogen on two distinct targets mediates embryo implantation in the mouse. Endocrinology 1998, 139, 5235–5246. [Google Scholar] [CrossRef] [PubMed]
- Paria, B.C.; Das, S.K.; Andrews, G.K.; Dey, S.K. Expression of the epidermal growth factor receptor gene is regulated in mouse blastocysts during delayed implantation. Proc. Natl. Acad. Sci. USA 1993, 90, 55–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Accession No. | Primer Sequence | Product Size (bp) |
|---|---|---|---|
| CD73 | NM_011851.4 | 1 F; AGGTTGTGGGGATTGTTGGA 2 R; CCCCAGGGCGATGATCTTAT | 152 |
| Rpl7 | NM_011291.5 | 1 F; TCAATGGAGTAAGCCCAAAG 2 R; GAAGAGACCGAGCAATCAAG | 246 |
| Lf | NM_008522.3 | 1 F; AGGAAAGCCCCCCTACAAAC 2 R; GGAACACAGCTCTTTGAGAA | 141 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.; Park, H.; Moon, S.; Do, J.-T.; Hong, K.; Choi, Y. Expression and Regulation of CD73 during the Estrous Cycle in Mouse Uterus. Int. J. Mol. Sci. 2021, 22, 9403. https://doi.org/10.3390/ijms22179403
Lee J, Park H, Moon S, Do J-T, Hong K, Choi Y. Expression and Regulation of CD73 during the Estrous Cycle in Mouse Uterus. International Journal of Molecular Sciences. 2021; 22(17):9403. https://doi.org/10.3390/ijms22179403
Chicago/Turabian StyleLee, Jihyun, Haeun Park, Sohyeon Moon, Jeong-Tae Do, Kwonho Hong, and Youngsok Choi. 2021. "Expression and Regulation of CD73 during the Estrous Cycle in Mouse Uterus" International Journal of Molecular Sciences 22, no. 17: 9403. https://doi.org/10.3390/ijms22179403
APA StyleLee, J., Park, H., Moon, S., Do, J. -T., Hong, K., & Choi, Y. (2021). Expression and Regulation of CD73 during the Estrous Cycle in Mouse Uterus. International Journal of Molecular Sciences, 22(17), 9403. https://doi.org/10.3390/ijms22179403