
Activity of the Lateral Hypothalamus during Genetically Determined Absence Seizures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
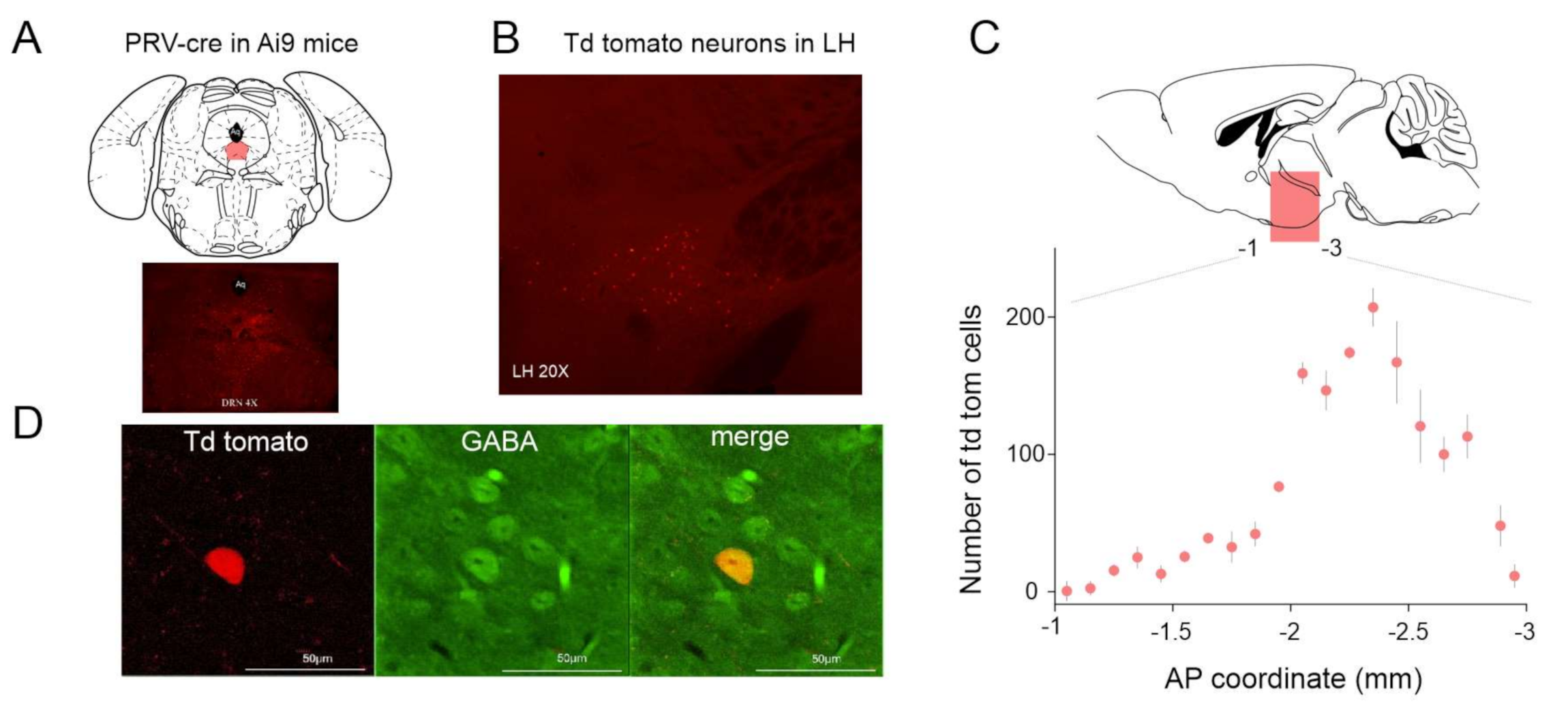
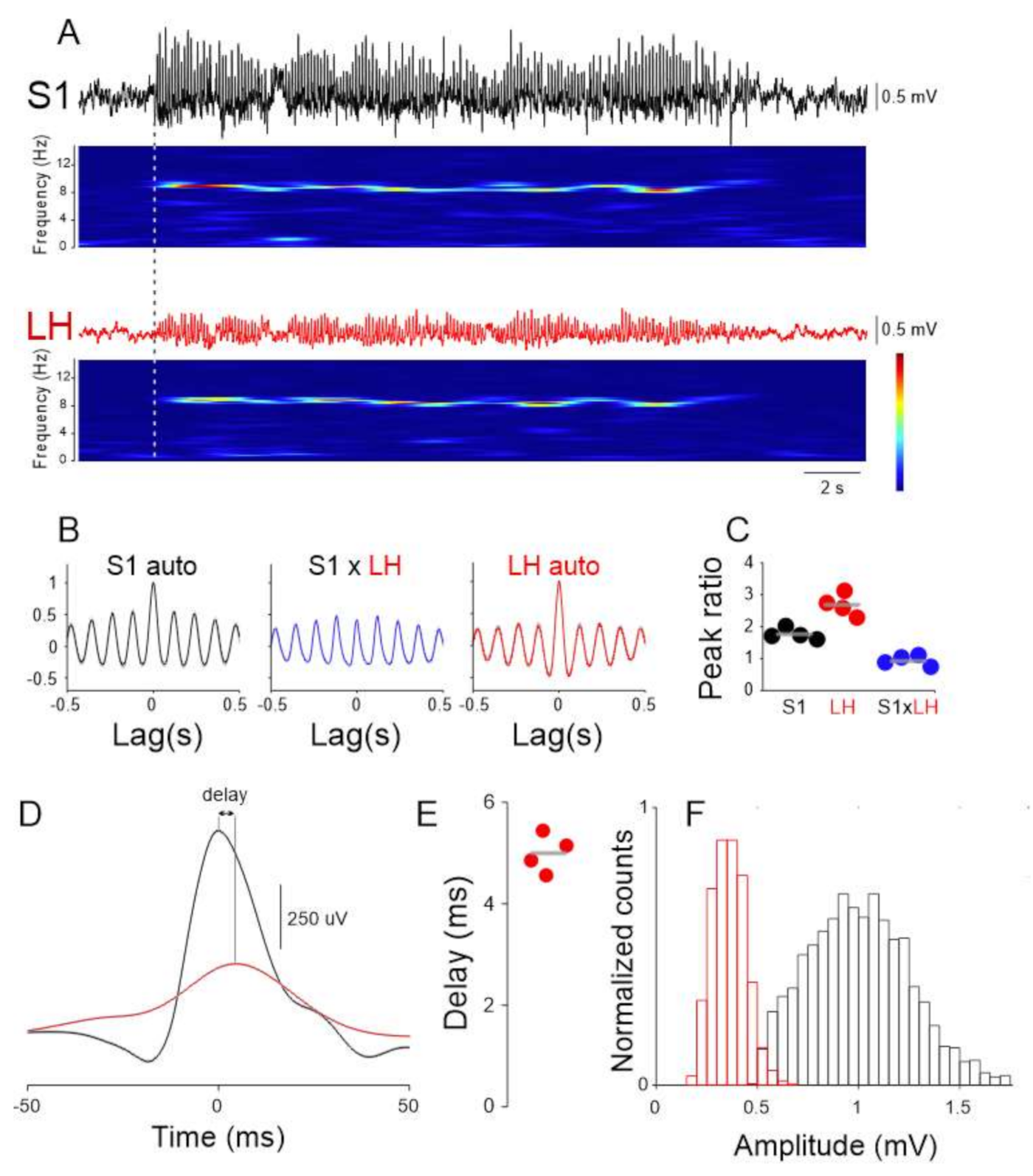
2.1. Functial Connectivity between LH and DRN
2.2. Local SWDs in the LH
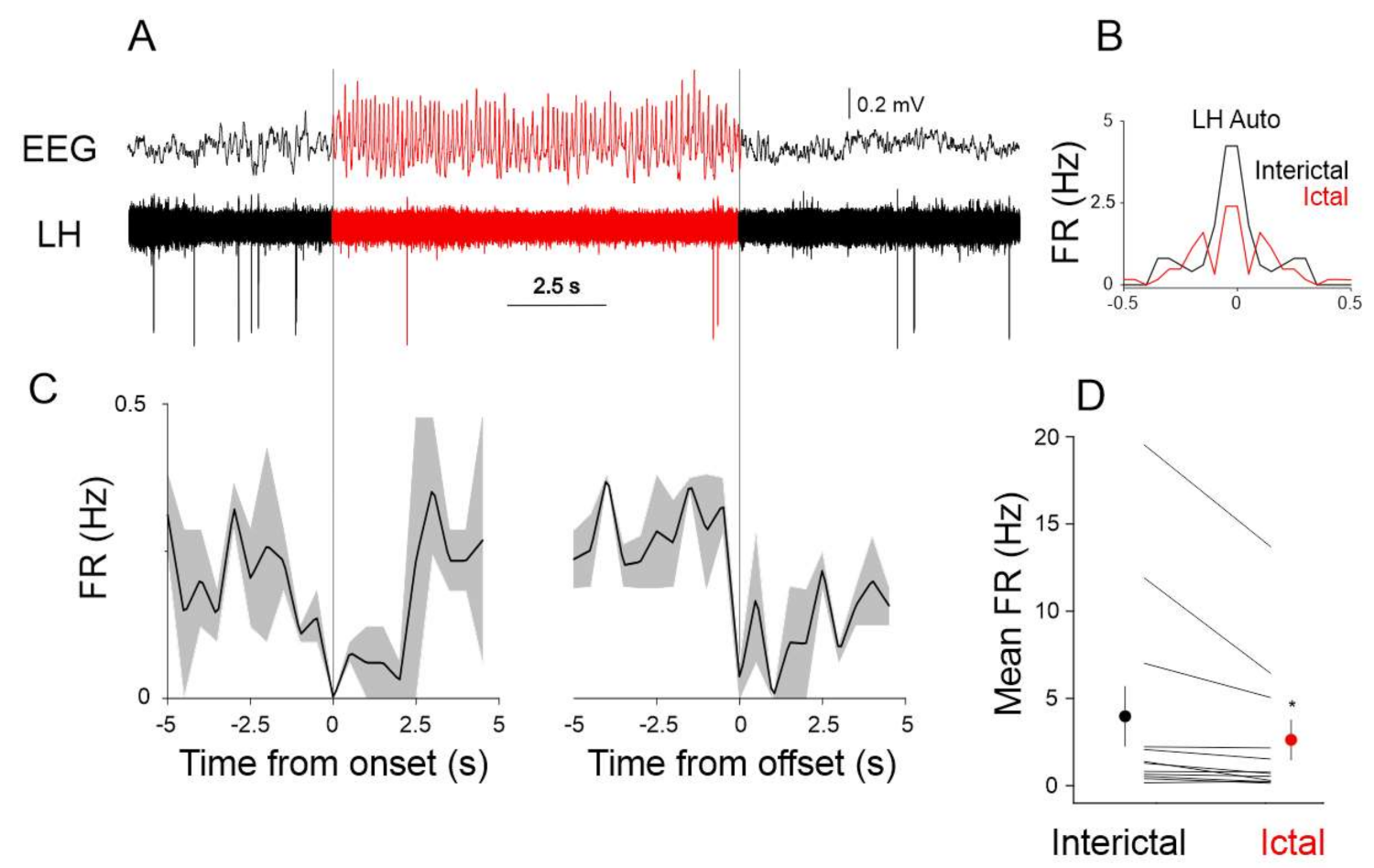
2.3. LH Neuronal Activity Correlates with SWDs
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crunelli, V.; Lőrincz, M.L.; McCafferty, C.; Lambert, R.C.; Leresche, N.; Di Giovanni, G.; David, F. Clinical and experimental insight into pathophysiology, comorbidity and therapy of absence seizures. Brain 2020, 143, 2341–2368. [Google Scholar] [CrossRef]
- Cnaan, A.; Shinnar, S.; Arya, R.; Adamson, P.C.; Clark, P.O.; Dlugos, D.; Hirtz, D.G.; Masur, D.; Glauser, T.A.; For the Childhood Absence Epilepsy Study Group. Second monotherapy in childhood absence epilepsy. Neurology 2017, 88, 182–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glauser, T.A.; Cnaan, A.; Shinnar, S.; Hirtz, D.G.; Dlugos, D.; Masur, D.; Clark, P.O.; Adamson, P.C.; for the Childhood Absence Epilepsy Study Team. Ethosuximide, valproic acid, and lamotrigine in childhood absence epilepsy: Initial monotherapy outcomes at 12 months. Epilepsia 2013, 54, 141–155. [Google Scholar] [CrossRef] [Green Version]
- Masur, D.; Shinnar, S.; Cnaan, A.; Shinnar, R.C.; Clark, P.; Wang, J.; Weiss, E.F.; Hirtz, D.G.; Glauser, T.A.; For the Childhood Absence Epilepsy Study Group. Pretreatment cognitive deficits and treatment effects on attention in childhood absence epilepsy. Neurology 2013, 81, 1572–1580. [Google Scholar] [CrossRef] [Green Version]
- Bagdy, G.; Kecskemeti, V.; Riba, P.; Jakus, R. Serotonin and epilepsy. J. Neurochem. 2007, 100, 857–873. [Google Scholar] [CrossRef]
- Graf, M.; Jakus, R.; Kantor, S.; Levay, G.; Bagdy, G. Selective 5-HT1A and 5-HT7 antagonists decrease epileptic activity in the WAG/Rij rat model of absence epilepsy. Neurosci. Lett. 2004, 359, 45–48. [Google Scholar] [CrossRef]
- Ohno, Y.; Sofue, N.; Imaoku, T.; Morishita, E.; Kumafuji, K.; Sasa, M.; Serikawa, T. Serotonergic modulation of absence-like seizures in groggy rats: A novel rat model of absence epilepsy. J. Pharmacol. Sci. 2010, 114, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Deransart, C.; Depaulis, A. The control of seizures by the basal ganglia? A review of experimental data. Epileptic Disord. 2002, 4 (Suppl. S3), S61–S72. [Google Scholar]
- Danober, L.; Depaulis, A.; Marescaux, C.; Vergnes, M. Effects of cholinergic drugs on genetic absence seizures in rats. Eur. J. Pharmacol. 1993, 234, 263–268. [Google Scholar] [CrossRef]
- Micheletti, G.; Warter, J.M.; Marescaux, C.; Depaulis, A.; Tranchant, C.; Rumbach, L.; Vergnes, M. Effects of drugs affecting noradrenergic neurotransmission in rats with spontaneous petit mal-like seizures. Eur. J. Pharmacol. 1987, 135, 397–402. [Google Scholar] [CrossRef]
- Sitnikova, E.; van Luijtelaar, G. Reduction of adrenergic neurotransmission with clonidine aggravates spike-wave seizures and alters activity in the cortex and the thalamus in WAG/Rij rats. Brain Res. Bull. 2005, 64, 533–540. [Google Scholar] [CrossRef]
- Franklin, M.E.; Foa, E.B. Treatment of obsessive compulsive disorder. Annu. Rev. Clin. Psychol. 2011, 7, 229–243. [Google Scholar] [CrossRef]
- Wong, D.T.; Perry, K.W.; Bymaster, F.P. Case history: The discovery of fluoxetine hydrochloride (Prozac). Nat. Rev. Drug Discov. 2005, 4, 764–774. [Google Scholar] [CrossRef]
- Wong, M.L.; Licinio, J. Research and treatment approaches to depression. Nat. Rev. Neurosci. 2001, 2, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Q.; Buchanan, G.F.; Motelow, J.E.; Andrews, J.; Vitkovskiy, P.; Chen, W.C.; Serout, F.; Gummadavelli, A.; Kundishora, A.; Furman, M.; et al. Impaired serotonergic brainstem function during and after seizures. J. Neurosci. 2016, 36, 2711–2722. [Google Scholar] [CrossRef] [Green Version]
- Lőrincz, M.; Olah, M.; Baracskay, P.; Szilagyi, N.; Juhasz, G. Propagation of spike and wave activity to the medial prefrontal cortex and dorsal raphe nucleus of WAG/Rij rats. Physiol. Behav. 2007, 90, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Venzi, M.; David, F.; Bellet, J.; Cavaccini, A.; Bombardi, C.; Crunelli, V.; Di Giovanni, G. Role for serotonin2A (5-HT2A) and 2C (5-HT2C) receptors in experimental absence seizures. Neuropharmacology 2016, 108, 292–304. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, S.K.; Cohen, J.Y.; Hwang, D.; Uchida, N.; Watabe-Uchida, M. Organization of monosynaptic inputs to the serotonin and dopamine neuromodulatory systems. Cell Rep. 2014, 8, 1105–1118. [Google Scholar] [CrossRef] [Green Version]
- Weissbourd, B.; Ren, J.; DeLoach, K.E.; Guenthner, C.J.; Miyamichi, K.; Luo, L. Presynaptic partners of dorsal raphe serotonergic and GABAergic neurons. Neuron 2014, 83, 645–662. [Google Scholar] [CrossRef] [Green Version]
- Saper, C.B.; Fuller, P.M.; Pedersen, N.P.; Lu, J.; Scammell, T.E. Sleep state switching. Neuron 2010, 68, 1023–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardis, L.L.; Bellinger, L.L. The lateral hypothalamic area revisited: Ingestive behavior. Neurosci. Biobehav. Rev. 1996, 20, 189–287. [Google Scholar] [CrossRef]
- Herrera, C.G.; Cadavieco, M.C.; Jego, S.; Ponomarenko, A.; Korotkova, T.; Adamantidis, A. Hypothalamic feedforward inhibition of thalamocortical network controls arousal and consciousness. Nat. Neurosci. 2016, 19, 290–298. [Google Scholar] [CrossRef] [Green Version]
- Gazea, M.; Furdan, S.; Sere, P.; Oesch, L.; Molnar, B.; Di Giovanni, G.; Fenno, L.E.; Ramakrishnan, C.; Mattis, J.; Deisseroth, K.; et al. Reciprocal Lateral Hypothalamic and Raphe GABAergic Projections Promote Wakefulness. J. Neurosci. 2021, 41, 4840–4849. [Google Scholar] [CrossRef]
- Floyd, N.S.; Price, J.L.; Ferry, A.T.; Keay, K.A.; Bandler, R. Orbitomedial prefrontal cortical projections to hypothalamus in the rat. J. Comp. Neurol. 2001, 432, 307–328. [Google Scholar] [CrossRef]
- Gabbott, P.L.; Warner, T.A.; Jays, P.R.; Salway, P.; Busby, S.J. Prefrontal cortex in the rat: Projections to subcortical autonomic, motor, and limbic centers. J. Comp. Neurol. 2005, 492, 145–177. [Google Scholar] [CrossRef]
- Sesack, S.R.; Deutch, A.Y.; Roth, R.H.; Bunney, B.S. Topographical organization of the efferent projections of the medial prefrontal cortex in the rat: An anterograde tract-tracing study with Phaseolus vulgaris leucoagglutinin. J. Comp. Neurol. 1989, 290, 213–242. [Google Scholar] [CrossRef] [PubMed]
- McCafferty, C.; David, F.; Venzi, M.; Lőrincz, M.L.; Delicata, F.; Atherton, Z.; Recchia, G.; Orban, G.; Lambert, R.C.; di Giovanni, G.; et al. Cortical drive and thalamic feed-forward inhibition control thalamic output synchrony during absence seizures. Nat. Neurosci. 2018, 21, 744–756. [Google Scholar] [CrossRef] [PubMed]
- Vergnes, M.; Marescaux, C.; Depaulis, A. Mapping of spontaneous spike and wave discharges in Wistar rats with genetic generalized non-convulsive epilepsy. Brain Res. 1990, 523, 87–91. [Google Scholar] [CrossRef]
- Petreanu, L.; Mao, T.; Sternson, S.M.; Svoboda, K. The subcellular organization of neocortical excitatory connections. Nature 2009, 457, 1142–1145. [Google Scholar] [CrossRef]
- Oyibo, H.K.; Znamenskiy, P.; Oviedo, H.V.; Enquist, L.W.; Zador, A.M. Long-term Cre-mediated retrograde tagging of neurons using a novel recombinant pseudorabies virus. Front. Neuroanat. 2014, 8, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marescaux, C.; Vergnes, M.; Depaulis, A. Genetic absence epilepsy in rats from Strasbourg—A review. J. Neural Transm. Suppl. 1992, 35, 37–69. [Google Scholar] [PubMed]
- Polack, P.O.; Mahon, S.; Chavez, M.; Charpier, S. Inactivation of the somatosensory cortex prevents paroxysmal oscillations in cortical and related thalamic neurons in a genetic model of absence epilepsy. Cereb. Cortex 2009, 19, 2078–2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mickelsen, L.E.; Kolling, F.W., IV; Chimileski, B.R.; Fujita, A.; Norris, C.; Chen, K.; Nelson, C.E.; Jackson, A.C. Neurochemical heterogeneity among lateral hypothalamic hypocretin/orexin and melanin-concentrating hormone neurons identified through single-cell gene expression analysis. eNeuro 2017, 4. [Google Scholar] [CrossRef] [Green Version]
- Allers, K.A.; Sharp, T. Neurochemical and anatomical identification of fast- and slow-firing neurones in the rat dorsal raphe nucleus using juxtacellular labelling methods in vivo. Neuroscience 2003, 122, 193–204. [Google Scholar] [CrossRef]
- Lottem, E.; Lőrincz, M.L.; Mainen, Z.F. Optogenetic activation of dorsal raphe serotonin neurons rapidly inhibits spontaneous but not odor-evoked activity in olfactory cortex. J. Neurosci. 2016, 36, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Gurgu, R.S.; Ciobanu, A.M.; Danasel, R.I.; Panea, C.A. Psychiatric comorbidities in adult patients with epilepsy (A systematic review). Exp. Ther. Med. 2021, 22, 909. [Google Scholar] [CrossRef] [PubMed]
- Aygun, H.; Ayyildiz, M. Vortioxetine increases absence-like seizures in WAG/Rij rats but decreases penicillin- and pentylenetetrazole-induced seizures in Wistar rats. Epilepsy Behav. 2021, 116, 107797. [Google Scholar] [CrossRef] [PubMed]
- Madisen, L.; Zwingman, T.A.; Sunkin, S.M.; Oh, S.W.; Zariwala, H.A.; Gu, H.; Ng, L.L.; Palmiter, R.D.; Hawrylycz, M.J.; Jones, A.R.; et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat. Neurosci. 2010, 13, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sere, P.; Zsigri, N.; Raffai, T.; Furdan, S.; Győri, F.; Crunelli, V.; Lőrincz, M.L. Activity of the Lateral Hypothalamus during Genetically Determined Absence Seizures. Int. J. Mol. Sci. 2021, 22, 9466. https://doi.org/10.3390/ijms22179466
Sere P, Zsigri N, Raffai T, Furdan S, Győri F, Crunelli V, Lőrincz ML. Activity of the Lateral Hypothalamus during Genetically Determined Absence Seizures. International Journal of Molecular Sciences. 2021; 22(17):9466. https://doi.org/10.3390/ijms22179466
Chicago/Turabian StyleSere, Péter, Nikolett Zsigri, Timea Raffai, Szabina Furdan, Fanni Győri, Vincenzo Crunelli, and Magor L. Lőrincz. 2021. "Activity of the Lateral Hypothalamus during Genetically Determined Absence Seizures" International Journal of Molecular Sciences 22, no. 17: 9466. https://doi.org/10.3390/ijms22179466
APA StyleSere, P., Zsigri, N., Raffai, T., Furdan, S., Győri, F., Crunelli, V., & Lőrincz, M. L. (2021). Activity of the Lateral Hypothalamus during Genetically Determined Absence Seizures. International Journal of Molecular Sciences, 22(17), 9466. https://doi.org/10.3390/ijms22179466