Abstract
Currently, infertility affects 8–12% of reproductive age couples worldwide, a problem that also affects women suffering from recurrent implantation failure (RIF). RIF is a complex condition resulting from many physiological and molecular mechanisms involving dynamic endometrium–blastocyst interaction. The most important are the endometrial receptivity process, decidualization, trophoblast invasion, and blastocyst nesting. Although the exact multifactorial pathogenesis of RIF remains unclear, many studies have suggested the association between hormone level imbalance, disturbances of angiogenic and immunomodulatory factors, certain genetic polymorphisms, and occurrence of RIF. These studies were performed in quite small groups. Additionally, the results are inconsistent between ethnicities. The present review briefly summarizes the importance of factors involved in RIF development that could also serve as diagnostic determinants. Moreover, our review could constitute part of a new platform for discovery of novel diagnostic and therapeutic solutions for RIF.
1. Introduction
Recurrent implantation failure (RIF) is defined as three or more consecutive failed in vitro attempts with at least four high-quality embryos in a minimum of three fresh or frozen cycles [1]. This serious complication refers to women younger than 40 [2,3]. It is estimated that 5% of women suffer from recurrent pregnancy loss (RPL), in which 75% of cases are diagnosed as RIF. Among patients under infertility treatment, 15% experience RIF [4].
A successful pregnancy is a result of proper embryo implantation, followed by adequate decidualization, as well as placentation [2]. Each step requires many molecular and physiological processes for making it possible to achieve the balance necessary for pregnancy development. Implantation is a combination of proper endometrial function and dynamic interaction between the endometrium and the blastocyst. The most important is the implantation phase, which is a multipart process and requires stability in adhesion of the blastocyst, invasion of trophoblast cells, and immune modulation. Achieving implantation strongly depends on proper structure and adequate receptivity of the endometrium (Figure 1). The specific time of the higher rate of successful implantation, called the window of implantation (WOI) (days 20–24 of the cycle, during the secretory phase), depends on high receptivity of the endometrium. Endometrial receptivity is noted upon the appearance of pinopodes, which are microvilli on the apical surface of the epithelial cells (days 20 and 21 of the cycle, up to 5 days of variation). However, without a doubt, endometrial receptivity is a result of many physiological and molecular mechanisms. During the in vitro process and embryo transfer procedure, it is essential to recognize WOI and to maintain endometrial receptivity. Growing evidence suggests that performing elective frozen or thawed embryo transfer may enhance proper implantation (“freeze-all policy”) [5].
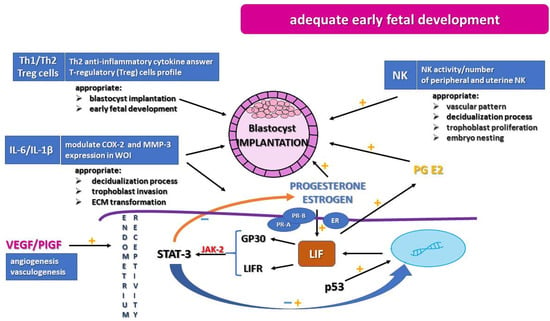
Figure 1.
Creation of utero environment for appropriate endometrium receptivity, trophoblast proliferation, and embryo nesting. COX-2, cyclooxygenase-2; ECM, extracellular matrix; ER, estrogen receptors; GP130, glycoprotein 130; IL-1β, interleukin-1β; IL-6, interleukin-6; JAK-2, Janus kinase 2; LIF, leukemia inhibitor factor; LIFR, membrane LIF receptor; MMP-3, metalloproteinase-3; NK, natural killer; p53, tumor protein p53; PlGF, placental growth factor; PR-A/PR-B, progesterone receptors; STAT-3, signal transducer and activator of transcription 3; Th1/Th2, T cells involved in inflammatory response; Treg, T-regulatory cell profile; WOI, window of implantation; VEGF, vascular endothelial growth factor.
Implantation failure is diagnosed as a lack of ultrasound signs of pregnancy in the uterine cavity. In several studies, a biochemical pregnancy was included (an increase in β-hCG without any ultrasound sign of pregnancy) to the definition of RIF. Because the implantation process is complex, the assessment of causes of RIF should be performed on several levels. The most common analyses are chromosomal testing of both parents, the estimation of ovarian function (FSH, LH, AMH measurement) in women, and sperm DNA fragmentation in men, as well as assessment of uterine pathologies and fallopian tube permeability (hysterosalpingogram, laparoscopy) [6,7].
The known risk factors of RIF include childbearing age, BMI, tobacco and alcohol intake, and history of endometriosis. The reasons for RIF could be also divided into embryo and uterine factors. On the other hand, the influence of the male and female factors on the occurrence of RIF has also been shown (Table 1). Although the exact multifactorial pathogenesis of RIF remains unclear, the current review tries to briefly summarize the factors involved in RIF development.

Table 1.
The frequent risk factors of RIF.
2. Immunological Profile
The proper immune response plays a crucial role in receptive endometrium during the WOI [34]. A precise immunologic balance at the maternal–fetal interface in the endometrium is needed to obtain a positive pregnancy outcome. In the early stage of pregnancy, the stability at the maternal–fetal surface is complex and involves immunological processes, including activity of innate lymphocytes, macrophages, decidual dendritic cells, and T cells. All of these cells play a pivotal role in immune acceptance of the embryo, and hence they are also involved in adverse pregnancy outcomes such as the RIF pathomechanism [35]. Innate lymphoid cells (ILCs) were identified in human decidua as playing an important role in maternal–fetal interface immunity and were classified into two subsets: natural killer (NK) cells and non-cytotoxic helper ILCs (ILC1s, ILC2s, ILC3s). The uterine dendritic cells play a vital role in the subtle balance in the decidual space. They are involved in angiogenesis and endometrial stromal cell proliferation and differentiation, as well as in controlling the immune response via T cell-mediated effects [36]. Macrophages are important cells regulating the microenvironment of the developing embryo and are involved in implantation, placentation, and embryo development. These cells are also linked with vascular remodeling at the maternal–fetal interface via promoting the invasion of trophoblasts into spiral arteries. Additionally, they remove apoptotic cells and cellular debris [37]. In the decidua, macrophages are divided into M1 (classically activated) and M2 (alternatively activated) cells. In the luteal phase and physiological healthy pregnancy, M2 macrophages are present in high numbers in the endometrium [37]. It is a crucial to maintain the correct proportion of M1/M2 cells for the appropriate pregnancy outcome; thus, a disproportion in the M1/M2 macrophage ratio leads to several complications during pregnancy, such as preeclampsia, intrauterine growth restriction, spontaneous and recurrent abortion, and RIF [37,38,39].
Many studies have indicated the significant role of dysregulation in the cellular immune response, including imbalance in natural killer (NK) and T cells in receptive endometrium development (Figure 1). This imbalance manifests as intensification of NK activity and augmentation of peripheral and uterine NK cell numbers, indicating the role in RIF etiology [10]. Apart from a uterine presence, NK cells exist in three subpopulations with different immunological features, and separation of the peripheral and uterine NK cells is difficult, so the activity of NK cells should be considered carefully. The abnormal activity of uterine NK cells could be a cause of disturbances in vascular pattern, ischemic symptoms, and augmentation of oxidative stress, which is very unfavorable in the early stage of trophoblast invasion [9,40]. On the other hand, it is recognized that the Th2 anti-inflammatory cytokine response, such as the T-regulatory (Treg) cell profile, supports appropriate implantation and early fetal development. Disturbances in Th1/Th2 profile are associated with recurrent miscarriages, and increased Th1 cytokine pro-inflammatory profile and Th17 phenotype are implicated in recurrent implantation failure [41].
All these pathological processes could lead to the creation of a cytotoxic environment in utero, with inappropriate endometrium receptivity, difficult trophoblast proliferation, and impossibility of embryo nesting. Considering the above-mentioned facts, many studies focus on the inflammatory immune response, which probably plays a vital role in RIF etiology. It was revealed that several cytokines could be factors predicting the occurrence of RIF [7].
Interleukins, such as interleukin-6 (IL-6), leukemia inhibitor factor (LIF), and IL-1 cytokines, play an important role in embryo development and the occurrence of the receptivity window. IL-6, which is synthesized by macrophages, fibroblasts, epithelial cells, and placental trophoblasts, is a multifunctional cytokine greatly contributing to fertility process as well as to the acute immune response. IL-6 level is observed to be high in the luteal phase, particularly in the receptivity window. It is involved in placenta and pregnancy development [42,43]. IL-1a, IL-1b, and the receptor antagonist IL-Ira belong to the same family, have similar biological effects, and are involved in the immune response. They act by the same receptor: IL-1 receptor (IL-1R). These cytokines are shown to be very important in modulation of the process of maternal endometrium decidual change and embryo development. IL-1b, expressed in a high level in the first trimester of pregnancy, is secreted by cytotrophoblast cells. Importantly, IL-1b modulates the expression of cyclooxygenase-2 (COX-2) and metalloproteinase-3 (MMP-3), both recognized to be crucial in homeostasis of decidualization, trophoblast invasion, and extracellular matrix (ECM) transformation [44].
The significant element of the endometrium during the WOI period is the pinopodes, considered to have an important role in endometrial receptivity. Qiong et al. evaluated the clinical value of pinopodes in humans. They concluded that pinopode scoring is a reliable marker of endometrial receptivity and could predict successful embryo transfer [45].
3. Leukemia Inhibitor Factor
One of the most important factors modulating endometrium receptivity is leukemia inhibitor factor (LIF), a pleiotropic cytokine belonging to the IL-6 superfamily and known to play a major role in a wide array of physiological processes. During the receptivity window, LIF promoting the development of pinopodes supports blastocyst attachment and invasion and the decidualization process [46].
Several studies have shown the decreasing level of LIF in women with recurrent implantation failure. In the study of Stewart et al., knockout mice deficient in the LIF gene were fertile and were able to conceive and develop a blastocyst, but implantation did not occur [47]. Song et al. suggest that reduction in prostaglandin synthesis may influence the implantation failure connected with LIF alteration [11]. LIF provides upregulation of prostaglandin E2 and stimulates trophoblast development. Prostaglandins are mediators of decidualization [48]. Implantation may be induced after administration of exogenous LIF; hence, subcutaneous injection of LIF may increase the success rate of implantation [12,49]. Another association between LIF and implantation was found in the research on emergency contraception. One of the properties of levonorgestrel is to inhibit LIF, which is a suggested contraceptive mechanism [50]. LIF regulates cellular function via binding to the membrane receptor LIF receptor (LIFR) and glycoprotein 130 (GP130) and activates the downstream signal transduction pathway as a signal transducer and activator of transcription (STAT) [13]. After binding, the receptors dimerize and recruit the kinase Janus kinase 2 (JAK-2), which activates signal transducer and activator of transcription 3 (STAT-3). STAT-3 acts as a gene regulator in the nucleus, and activation of STAT-3 by LIF coincides with the beginning of the WOI [51]. A study of Hiraoka et al. addressed the role of STAT-3 and showed that uterine and stromal STAT-3 may have a crucial role in the implantation process and may suppress the estrogenic response and endometrial proliferation. Thus, it indicates embryo attachment [52].
LIF expression could be altered by estrogen and progesterone. In a mouse model, estrogen can significantly increase expression of LIF in the endometrium during the WOI [49]. Choi et al. observed an alteration in progesterone/estradiol and the LIF pathway. Although changes in expression of hormonal receptors were not observed, decreases in expression of STAT-3 and a gene set associated with the JAK–STAT pathway were demonstrated in patients with RIF. Downregulation of 22 genes metabolizing estrogen and progesterone in the endometrium was detected [53]. Sun et al. observed an elevated estrogenic response in the endometrium connected with STAT-3 gene deficiency [54]. STAT-3 knockout mice demonstrated upregulation of an estrogen-responsive gene and downregulation of some progesterone-responsive genes, which highlighted the reduction in P4-signaling pathways in the endometrium [54,55]. Additionally, LIF is a gene target for p53, which increases its expression. The molecule p53 binds to the p53-binding element in the first intron and alters the expression of LIF in different tissue, including endometrial tissue. The absence of p53 leads to the reduction in LIF and impairment of the implantation process [49]. Considering pleiotropic LIF function, it remains clear that this factor is recognized as a clinical marker of endometrial receptivity occurrence.
4. Glycodelin-A
Glycodelin-A (GdA), an abundant secretory glycoprotein in the first-trimester decidua, is one of the important molecules taking part in feto-maternal tolerance, as well as in placental development in early pregnancy. GdA is a progesterone-induced glycoprotein secreted by endometrial glands into the uterus cavity. GdA, dominant in mid-secretory phase, occurs in six different glycosylated glycoforms, which differ in expression in each phase of the menstrual cycle. The presence of GdA in endometrial epithelial and stromal cells has been observed [15].
During proper implantation, GdA stimulates an endometrial proliferation and regulates an attachment of trophoblast and affects cell proliferation, differentiation, adhesion, and motility. Therefore, GdA is a vital molecule during trophoblast and placenta development [16,56]. There is also evidence that GdA could play a major role during implantation, modulating numerous processes, especially the immunosuppression mechanism. In this field, GdA reduces the cytotoxic effect of NK cells and modulates NK cells to cytokine production, with a balance shift to Th2 cytokines [57,58,59]. Moreover, GdA inhibits T cells proliferation, as well as stimulates T cells apoptosis and modulates the immune answer from B cells [17,60]. The important role of GdA in modulating the suitable phenotype of dendritic cells has also been shown [61].
GdA level could indicated to appropriate endometrial receptivity and the chance for successful implantation. It was revealed that GdA, as well as mucin-1 (MUC-1) in the human endometrium and in serum blood during WOI period, were significantly lower in RIF women compared with fertile women. Additionally, the correlation of blood and tissue levels of MUC-1 and GdA has been observed. [62]. These remarks indicate that the changed expression profile of GdA could be associated with impaired endometrial receptivity and could play an important role in recurrent implantation failure, early miscarriage, and the recurrent miscarriage pathomechanism [15,16]. Considering the above-mentioned findings, the GdA could be a good potential biomarker of endometrial activity.
5. Progesterone, Estrogen, and Their Hormonal Receptors
Extensive studies have demonstrated the important role of progesterone, estrogen, and hormonal receptors during implantation. Physiologically, after ovulation the estrogen level decreases and the progesterone level increases. Increased levels of estrogen and LIF in endometrium coincide with implantation time [49].
The correct balance between estrogen and progesterone enhances proper endometrial proliferation during the WOI and prepares the endometrium for blastocyst invasion. In women with RIF, the implantation could be hampered by an elevated estradiol level during ovarian stimulation that could lead to premature rising progesterone levels and maturation of the endometrium [49]. Several studies focus on this significant problem. Prapas et al. suggest that the time of endometrial exposure to a high progesterone level may distinguish the WOI during the menstrual cycle [63]. Klonos et al. assessed expression of estrogen (ER) and progesterone receptors (PR-A, PR-B), comparing receptor expression in the endometria of healthy women undergoing an oocyte donation program on days 0 and 5 of oocyte retrieval. A statistically significant increase in expression of PR-B and decrease in ER were observed, confirming the physiological changes in the endometrium during preparation for implantation. The expression of PR-A remained constant [64].
PR-B gene knockout mice had a reduced implantation rate in comparison with PR-A knockout mice, suggesting the relevance of PR-B expression for implantation [65]. In patients suffering from RIF, a decrease in ER was observed [66]. Similar results were obtained in another study, which showed a statistically significant correlation between ESR1 polymorphism and implantation failure [67].
Several authors suggest crucial interaction between p53, LIF, and hormonal receptors in the reproduction process. The multi-functional protein that is the p53 molecule plays a crucial role in several physiological processes, such as genomic stability, tumor suppression, and gene expression regulation. In numerous studies, its role in the reproductive system was demonstrated. Through inducing apoptosis and angiogenesis, it influences endometrial receptivity, as well as oocyte maturation and quality [68]. The p53 impact on implantation is meditated by LIF [49]. Furthermore, VEGF is one of the gene targets of p53, and alteration in p53 expression leads to an increase in VEGF tissue expression through hypoxia-induced factor [69].
6. Angiogenic Factors
Vascular endothelial growth factor (VEGF) is an angiogenic factor with a pleiotropic role in numerous pathophysiological processes. The VEGF family includes several proteins (VEGF-A to VEGF-F), as well as placental growth factor (PlGF), among which VEGF-A is recognized as the most important factor of angiogenesis involved in the regulation of endothelial cell function. It also seems that VEGF-A plays a crucial role in embryo implantation and placenta development during the early stage of pregnancy. In addition, placental expression of VEGF-A increases endometrial receptivity and positively improves the interactions between the embryo and endometrial tissue. VEGF also dynamically acts in angiogenesis and vascularization of the embryo. It was demonstrated that proper VEGF and PlGF gene expression is vital for embryo implantation, trophoblast invasion, adequate angiogenesis, and placenta development in successful pregnancy [70] (Figure 1). Both progesterone and estrogen are regulators of VEGF synthesis [71]. That means that embryo implantation is conditioned by estrogen, progesterone, and angiogenetic factors and that all of these should be taken into account as potential factors in this mechanism.
Moreover, both VEGF and PlGF could also modulate the immuno-tolerance processes in the maternal immune system throughout the embryo implantation, activating the monocytes and macrophages [72]. In addition, PlGF promotes the proliferation and differentiation of uterine NK cells, as well as dendritic cells, and could be able to balance towards the Th2 phenotype. The above-mentioned immunoregulatory roles of VEGF and PlGF are crucial for the initiation and development of pregnancy [73].
Endometrial VEGF expression is reduced in the peri-implantation period in infertile women [74], as well as in women with recurrent miscarriages [75]. In patients with RIF, an elevated level of VEGF in blood and a decreased level of endometrial expression are observed. The results show the significantly lower level of MMP-7 and VEGF in peripheral blood in the RIF women compared with the controls, reflecting the role of these circulating molecules in the implantation process. Both of these molecules could be proposed as biomarkers of RIF development [76]. In several studies, altered expression of VEGF in recurrent miscarriages shows the importance of the angiogenic factor in placenta development [77,78].
The main functions of PlGF, another important member of the VEGF family, are chemotactic, mitogenic, and angiogenic activity of endothelial cells. On the other hand, PlGF also has chemotactic function for monocytes and several other hemopoietic cells. This molecule is expressed during embryo implantation by maternal uterine NK (uNK) and fetal trophoblast cells. PlGF is reported to be associated with pathological angiogenesis, which occurs in inflammatory processes and tumor angiogenesis [79]. The decreasing concentration of PlGF in preeclamptic women suggests that this molecule could predict occurrence of the disease [80,81]. It was suggested that PlGF could play an important role in uterine NK cell proliferation, as well as in differentiation.
Because PlGF and VEGF act by the same VEGF receptor (VEGFR-1, fms-like tyrosine kinase-1), they each potentiate the biological effects of the other. PlGF and VEGF are expressed by maternal endometrial cells, and in addition, PlGF is abundantly expressed in the trophoblast and placenta (villous cytotrophoblasts, syncytiotrophoblasts, extravillous trophoblasts, villous mesenchyme). Both PlGF and VEGF are expressed by uterine NK, endometrial glands, and macrophages [74,82].
The significant role of uterine NK in pregnancy is to modulate the angiogenesis and vasculogenesis in the early stage of trophoblast invasion [73].
7. Genetic Factors
Various genetic factors are involved in the critical process of embryo implantation, and several single-nucleotide polymorphisms (SNPs) have been reported to be associated with RIF.
MicroRNAs, recognized function modulators, could control the expression of many genes involved in the peri-implantation period and in this way could play a vital role in fetal/placental development. On the other hand, these factors might be closely involved in recurrent implantation failure (IVF) and recurrent pregnancy loss (RPL) pathogenesis [28].
In the population of Korean women, the significant role of miR-449bA>G polymorphism (AG + GG genotype was associated with RIF prevalence, OR = 1.584, p = 0.046) [3] as well as miR-27a rs895819 and miR-449b rs10061133 [83] in RIF predisposition and RPL development was observed. Despite these positive observations, the exact mechanism underlying the function of microRNAs in RIF etiopathology remains unclear [3].
In several analyses it was shown that inherited thrombophilia could be a risk factor of recurrent RIF failure. Safdarian et al. observed that the presence of factor V Leiden mutation (OR = 3.06, p = 0.01) and the homozygote form of methylene tetrahydrofolate reductase (MTHFR) mutation (OR = 12.33, p = 0.05) were significant risk factors for recurrent IVF failure [27]. In addition, the occurrence of at least one thrombophilia was a risk factor of recurrent IVF failure (OR = 3.15, p = 0.00). In the study performed by Qublan et al., at least one inherited or acquired thrombophilia was determined in 68.9% of the RIF group vs. 25.6% in the group with successful pregnancy after the first IVF–embryo transfer cycle and 25% in the control group, which included women with uneventful pregnancy and without miscarriages in their history (p < 0.01). Combined thrombophilia was more frequent in the repeated IVF failure group vs. the two above-mentioned control groups (35.6% vs. 4.4% and 3%) (p < 0.0001). The authors demonstrated the crucial role of acquired and inherited thrombophilia in IVF–embryo transfer implantation failure and concluded that women with repeated IVF–embryo transfer failure should be screened for thrombophilia [84]. In contrast, several analyses did not reveal a significant role of thrombophilic factors in IVF–embryo transfer implantation failure [85,86,87]. In a meta-analysis performed by Zeng et al. (nine studies including 1812 women: 754 RIF vs. 1058 controls), no association between either MTHFR polymorphism (MTHFR C677T or MTHFR A1298C) and RIF occurrence was confirmed [88].
Several studies have revealed the possible role of p53 genetic polymorphisms in RIF occurrence. An SNP in codon 72, replacing G with C, which results in proline (P72) instead of arginine (R72) in the protein chain, was identified as modulating the biological activity of protein p53, consequently influencing the reproductive capacity [30,87,88]. Interestingly, the women carriers of the C allele (proline) of p53 codon 72 polymorphism showed decreased leukemia inhibitory factor expression, thus leading to a lower rate of implantation success [14].
An earlier study by Kang et al. showed that p53 P72 allele presence was higher in women undergoing in vitro treatment in comparison with the general population (33.1% vs. 22.7%, respectively, p < 0.005). Additionally, this polymorphic variant identified in women younger than 35 appears to be a risk factor for implantation failure (implantation rate 19% vs. 42% in homozygous and heterozygous for P72 allele, respectively, p = 0.0028) [89]. These findings were confirmed in an analysis conducted by Lledo et al., who indicated that R72P polymorphism of the p53 gene is more frequent in the group of RIF and RPL patients compared with the fertile population [90]. In addition, it was also suggested that rs1042522 (R72P, G/C) and rs17878362 (Ins16bp, N/D) variants of the p53 gene could be a genetic factor predisposing to RIF [91,92].
Human leukocyte antigen (HLA)-G is recognized as an important factor of the immunomodulatory system, modulating the fertilization process as well as influencing the early stage of pregnancy; thus, its expression has been the subject of numerous analyses in women with RIF. Several studies have shown that polymorphisms of the HLA-G gene could be involved in effective implantation after the IVF–ETs technique. The significant role of several HLA-G polymorphism (rs1632947, rs1233334, rs371194629 HLA-G) in infertile patients, such as soluble HLA-G in the early stages of pregnancy, was indicated in the study of Nowak et al. in a population of Polish women [31]. On the other hand, in a large meta-analysis, Fan et al. found that an HLA-G 14-bp insertion allele probably increases the risk of RIF in Caucasians. In a meta-analysis, the authors found that HLA-G 14 bp polymorphism was significantly correlated with RIF occurrence (OR = 1.74) [93].
The significant role of angiogenetic factors, especially VEGF, in the implantation process is not in doubt. In a metanalysis, in a group of 683 patients (305 RIF women, 378 controls), the authors observed the strongest association between the −1154A allele of the VEGF gene and RIF occurrence (OR 1.39, p = 0.01). These results indicated that the –1154A>G variant may be perceived as a risk factor of RIF [54]. Additionally, other studies showed that women carrying several genetic variants of VEGF genes would be at significant risk of RIF occurrence [92].
Additional factors associated with the implantation process are several variants of the estrogen receptor 1 (ESR1) gene. Vagnini et al. investigated the role of the ESR1/AA (rs12199722) genotype, which was common in the RIF group compared with women who became pregnant on their first cycle of IVF/intracytoplasmic sperm injection (OR = 7.9), as well as compared with women who became pregnant without treatment (OR = 2.8) [67]. This observation reveals the important role of this genetic polymorphism in RIF etiology.
8. Vaginal and Endometrial Microbiome (Microbiota) Disturbances
Many studies have shown that Lactobacillus species (mainly L. crispatus, L. gasseri, L. iners, L. jensenii), as the dominant component of the vaginal microbiota, can play a protective role and may possess a eubiotic effect on the vaginal microenvironment [77,94,95]. It is known that Lactobacillus species inhibit invasion and colonization of pathogenic bacteria by production of a high concentration of lactic acid and short-chain fatty acids, which can maintain the acidic and anaerobic environment in the vaginal area [18,96] and which possess antibacterial, antiviral and immunomodulatory properties [97]. Vaginal lactic acid may disrupt the outer membrane of Gram-negative bacteria such as Pseudomonas aeruginosa and Escherichia coli [98,99,100]; it can inhibit opportunistic infections caused by Gardnerella vaginalis, Trichomonas vaginalis, Neisseria gonorrhoeae, Chlamydia trachomatis, herpes simplex virus (HSV), and human Papillomavirus (HPV) [101]. Moreover, Lactobacillus species can produce antimicrobial peptides (bacteriocins, bacteriocin-like substances) and biosurfactants [68]. According to the latest data, it is estimated that the vaginal microbiota contains approximately 1010–1011 bacteria, whereas the endometrium harbors 4 orders of magnitude fewer bacteria than the vagina [19,77].
It should be emphasized that the vagino–uterine microbiota and its dysbiosis may influence the pathogenesis of several gynecological infectious and non-infectious illnesses [18,20,102], including gynecologic cancers [103]. Recently, it was observed that dysbiosis of the vaginal microbiome may be involved in recurrent implantation failure (RIF) at various stages, such as formation of gametes, implantation, and delivery [18,104]. Moreover, Moreno et al. [105] observed that the microbiota in the endometrial fluid is mainly dominated by Lactobacillus spp. (>90%), in comparison with Gardnerella, Streptococcus, and Bifidobacterium. However, in another group of women the endometrial microbiota was dominated by bacteria other than Lactobacillus spp., such as Atopobium, Bifidobacterium, Gardnerella, Megasphaera, Prevotella, Sneathia, and Streptococcus [19]. This kind of endometrial microbiota, along with a low level of Lactobacillus spp., has been associated with negative reproductive outcomes in patients—women had significantly lower implantation, pregnancy, ongoing pregnancy, and live birth rates, as well as higher miscarriage rates [105]. Another study [19] showed that endometrial dysbiosis may cause infertility. Although the endometrial and vaginal microbial populations are different, the key to recurrent implantation failure is Lactobacillus spp. It is well known that uterine infections caused by Streptococcus spp., Staphylococcus spp., Enterococcus spp., Escherichia coli, and Klebsiella pneumoniae are risk factors of infertility because inflammation in the reproductive tract may lead to impairment of embryo implantation and the onset of a successful pregnancy [19]. According to Schoenmakers and Laven [96], the relative abundance of Lactobacillus crispatus (>60%) and Lactobacillus iners (>60%) in the vagina may be used for the classification of women with higher chances to achieve a pregnancy during assisted reproductive technology. A recent review of studies [106] confirmed that Lactobacillus abundance in the female genital tract is associated with better reproductive outcomes and results of assisted reproductive technology. All risks factors are summarized in Table 1.
9. Treatment of RIF
It is worth underlining that the identification of RIF risk factors could also be useful in clinical practice. There is growing evidence that immunological therapeutic methods have an effective role in RIF patients. Ahmadi et al. revealed the benefits of using intravenous immunoglobulin (IVIG) therapy in RIF women. The authors evaluated 72 women with RIF who were divided into two subgroups: women who received IVIG, aspirin, and heparin (anoxaparin) and women who received aspirin and heparin (anoxaparin) and no IVIG. In all women, the function of Th17 and Treg cells, as two subgroups of CD4+ T cells, in implantation and pregnancy rates were assessed. It was found that the use of IVIG affects the regulation of immune mechanisms, especially Treg cytokines, and in this way could be an optimal choice in the treatment of implantation failure [107]. Furthermore, for the improvement of pregnancy rate, lymphocyte immunotherapy (LIT) and IVIG have been applied. In combination, they might augment the efficacy of embryo implantation through several processes, such as modulating Th1/Th2 balance, increasing T regulatory cells (Tregs), and inhibiting NK activity [107,108]. The benefits of using immunomodulatory treatment before embryo transfer were demonstrated in an interesting study by Kolanska et al. In RIF women, steroids and intralipid treatment were used. In this group, clinical pregnancies occurred significantly more frequently in treated versus untreated embryo transfers (p < 0.001). Both steroids and intralipids resulted in higher clinical pregnancy rates [109]. In the other study in women after IVF with a history of recurrent pregnancy loss or recurrent implantation failure, it was also indicated that immunomodulatory (prednisone, intravenous immunoglobulin G (IVIG)) and anticoagulation treatment (low molecular weight heparin and low dose aspirin) suggestively improved the pregnancy [108].
It has also been demonstrated that daily administration of hydroxychloroquine (HCQ) in RIF women downregulated Th17 cytokines and additionally upregulated Treg cytokines. Both of these processes, through influencing the Th17/Treg ratio, compensate for the immune response [110].
10. Materials and Methods
In this review, publications available in PubMed and Scopus databases as well as in Google Scholar were taken into account. The following keywords and their combinations were used: “recurrent implantation failure”, “endometrial receptivity”, “vaginal microbiota”, “risk factors”. Additional searches included references from identified publications. In the screening process, articles published in predatory journals and studies published earlier than 1980 were excluded.
11. Conclusions
Summarizing the pathogenesis of RIF, the most important roles are played by embryo and uterine factors and disturbances of the hormonal and cytokines network, along with angiogenic and immunomodulatory factors, as well as several multi-functional proteins, such as the p53 molecule. Other factors include the vaginal microbiome, as well as a genetic polymorphism of the gene involved in embryo implantation, immune response, and endometrial receptivity (Table 2).
Table 2.
Summary of pathophysiological processes in relation to RIF development according to environmental and pathophysiological risk factors.
Recurrent implantation failure is an important problem and a great challenge for human reproductive medicine. Implantation disorders affect the physical and mental health of women and are a common cause of disappointment in both partners, as well as constituting a great difficulty for clinicians. Undoubtedly, identifying the factors predisposing the occurrence of RIF will make it possible to individualize treatment plans and consider the “freeze-all policy” during in vitro treatment, which altogether could increase the rate of implantation success. Studies of RIF biomarkers could improve the chance of pregnancy. Many studies have revealed the association between hormones, angiogenic and immunomodulatory factors, as well as genetic polymorphisms and RIF occurrence, but these observations require further research on large samples of different ethnic populations.
Currently, it is a fact that infertility affects 8–12% of couples in reproductive age worldwide, and it is a growing problem. There is clearly a great need for progress in the development of diagnostic tests that would allow the assessment of RIF and RPL risk.
Author Contributions
Conceptualization, A.E.M. and P.J.; methodology, A.E.M.; software, A.E.M.; formal analysis, P.J.; writing—original draft preparation, A.E.M.; writing—review and editing, A.E.M., P.J., M.O.; visualization, A.E.M.; supervision, P.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The study did not report any data.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sun, Y.; Zhang, Y.; Ma, X.; Jia, W.; Su, Y. Determining diagnostic criteria of unexplained recurrent implantation failure: A ret-rospective study of two vs. three or more implantation failure. Front. Endocrinol. 2021, 12, 619437. [Google Scholar] [CrossRef]
- Coughlan, C. What to do when good-quality embryos repeatedly fail to implant. Best Pr. Res. Clin. Obstet. Gynaecol. 2018, 53, 48–59. [Google Scholar] [CrossRef]
- Kim, J.O.; Ahn, E.H.; Sakong, J.H.; An, H.J.; Park, H.S.; Kim, Y.R.; Lee, J.R.; Lee, W.S.; Kim, N.K. Association of miR-27aA>G, miR-423C>a, miR-449bA>G, and miR-604A>G Polymorphisms with Risk of Recurrent Implantation Failure. Reprod. Sci. 2020, 27, 29–38. [Google Scholar] [CrossRef]
- Busnelli, A.; Reschini, M.; Cardellicchio, L.; Vegetti, W.; Somigliana, E.; Vercelllini, P. How common is real repeated implantation failure? An indirect estimate of the prevalence. Reprod. Biomed. Online 2020, 40, 91–97. [Google Scholar] [CrossRef]
- Cimadomo, D.; Craciunas, L.; Vermeulen, N.; Vomstein, K.; Toth, B. Definition, diagnostic and therapeutic options in recurrent implantation failure: An international survey of clinicians and embryologists. Hum. Reprod. 2021, 36, 305–317. [Google Scholar] [CrossRef]
- Coughlan, C.; Ledger, W.; Wang, Q.; Liu, F.; Demirol, A.; Gurgan, T.; Cutting, R.; Ong, K.; Sallam, H.; Li, T. Recurrent implantation failure: Definition and management. Reprod. Biomed. Online 2014, 28, 14–38. [Google Scholar] [CrossRef] [PubMed]
- Bashiri, A.; Halper, K.I.; Orvieto, R. Recurrent Implantation Failure-update overview on etiology, diagnosis, treatment and future directions. Reprod. Biol. Endocrinol. 2018, 16, 121. [Google Scholar] [CrossRef]
- Sanders, B. Uterine factors and infertility. J. Reprod. Med. 2006, 51, 169–176. [Google Scholar]
- Kwak-Kim, J.; Bao, S.; Lee, S.K.; Kim, J.W.; Gilman-Sachs, A. Immunological Modes of Pregnancy Loss: Inflammation, Immune Effectors, and Stress. Am. J. Reprod. Immunol. 2014, 72, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Santillán, I.; Lozano, I.; Illán, J.; Verdú, V.; Coca, S.; Bajo-Arenas, J.M.; Martínez-Pastor, F. Where and when should natural killer cells be tested in women with repeated implantation failure? J. Reprod. Immunol. 2015, 108, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Lim, H.; Das, S.K.; Paria, B.C.; Dey, S.K. Dysregulation of EGF Family of Growth Factors and COX-2 in the Uterus during the Preattachment and Attachment Reactions of the Blastocyst with the Luminal Epithelium Correlates with Implantation Failure in LIF- Deficient Mice. Mol. Endocrinol. 2000, 14, 1147–1161. [Google Scholar] [CrossRef]
- Chen, J.R.; Cheng, J.-G.; Shatzer, T.; Sewell, L.; Hernandez, L.; Stewart, C.L. Leukemia Inhibitory Factor Can Substitute for Nidatory Estrogen and Is Essential to Inducing a Receptive Uterus for Implantation But Is Not Essential for Subsequent Embryogenesis1. Endocrinology 2000, 141, 4365–4372. [Google Scholar] [CrossRef] [PubMed]
- Salleh, N. Diverse Roles of Prostaglandins in Blastocyst Implantation. Sci. World J. 2014, 2014, 968141. [Google Scholar] [CrossRef]
- Kang, H.-J.; Rosenwaks, Z. p53 and reproduction. Fertil. Steril. 2018, 109, 39–43. [Google Scholar] [CrossRef]
- Focarelli, R.; Luddi, A.; De Leo, V.; Capaldo, A.; Stendardi, A.; Pavone, V.; Benincasa, L.; Belmonte, G.; Petraglia, F.; Piomboni, P. Dysregulation of GdA Expression in Endometrium of Women With Endometriosis: Implication for Endometrial Receptivity. Reprod. Sci. 2017, 25, 579–586. [Google Scholar] [CrossRef]
- Uchida, H.; Maruyama, T.; Nishikawa-Uchida, S.; Miyazaki, K.; Masuda, H.; Yoshimura, Y. Glycodelin in reproduction. Reprod. Med. Biol. 2013, 12, 79–84. [Google Scholar] [CrossRef]
- Lee, C.-L.; Lam, E.Y.F.; Lam, K.K.W.; Koistinen, H.; Seppälä, M.; Ng, E.H.Y.; Yeung, W.S.B.; Chiu, P.C.N. Glycodelin-A Stimulates Interleukin-6 Secretion by Human Monocytes and Macrophages through L-selectin and the Extracellular Signal-regulated Kinase Pathway. J. Biol. Chem. 2012, 287, 36999–37009. [Google Scholar] [CrossRef]
- Fu, M.; Zhang, X.; Liang, Y.; Lin, S.; Qian, W.; Fan, S. Alterations in Vaginal Microbiota and Associated Metabolome in Women with Recurrent Implantation Failure. MBio 2020, 11, e03242-19. [Google Scholar] [CrossRef]
- Moreno, I.; Simon, C. Relevance of assessing the uterine microbiota in infertility. Fertil. Steril. 2018, 110, 337–343. [Google Scholar] [CrossRef]
- Han, Y.; Liu, Z.; Chen, T. Role of Vaginal Microbiota Dysbiosis in Gynecological Diseases and the Potential Interventions. Front. Microbiol. 2021, 12, 643422. [Google Scholar] [CrossRef]
- Raziel, A.; Friedler, S.; Schachter, M.; Kasterstein, E.; Strassburger, D.; Ron-El, R. Increased frequency of female partner chromosomal abnormalities in patients with high-order implantation failure after in vitro fertilization. Fertil. Steril. 2002, 78, 515–519. [Google Scholar] [CrossRef]
- Voullaire, L.; Collins, V.; Callaghan, T.; McBain, J.; Williamson, R.; Wilton, L. High incidence of complex chromosome abnormality in cleavage embryos from patients with repeated implantation failure. Fertil. Steril. 2007, 87, 1053–1058. [Google Scholar] [CrossRef] [PubMed]
- Saleh, R.; Agarwal, A.; Nelson, D.R.; Nada, E.; El-Tonsy, M.H.; Alvarez, J.G.; Thomas, A.J.; Sharma, R. Increased sperm nuclear DNA damage in normozoospermic infertile men: A prospective study. Fertil. Steril. 2002, 78, 313–318. [Google Scholar] [CrossRef]
- Nikolaeva, M.; Arefieva, A.; Babayan, A.; Chagovets, V.; Kitsilovskaya, N.; Starodubtseva, N.; Frankevich, V.; Kalinina, E.; Krechetova, L.; Sukhikh, G. Immunoendocrine Markers of Stress in Seminal Plasma at IVF/ICSI Failure: A Preliminary Study. Reprod. Sci. 2021, 28, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Azem, F.; Many, A.; Yovel, I.; Amit, A.; Lessing, J.B.; Kupferminc, M.J. Increased rates of thrombophilia in women with repeated IVF failures. Hum. Reprod. 2004, 19, 368–370. [Google Scholar] [CrossRef]
- Grandone, E.; Colaizzo, D.; Bue, A.L.; Checola, M.G.; Cittadini, E.; Margaglione, M. Inherited thrombophilia and in vitro fertilization implantation failure. Fertil. Steril. 2001, 76, 201–202. [Google Scholar] [CrossRef]
- Safdarian, L.; Najmi, Z.; Aleyasin, A.; Aghahosseini, M.; Rashidi, M.; Asadollah, S. Recurrent IVF failure and hereditary thrombophilia. Iran. J. Reprod. Med. 2014, 12, 467–470. [Google Scholar]
- Chen, C.H.; Lu, F.; Yang, W.J.; Yang, P.E.; Chen, W.M.; Kang, S.T.; Huang, Y.S.; Kao, Y.C.; Feng, C.T.; Chang, P.C.; et al. A novel platform for discovery of differentially expressed microRNAs in patients with repeated implantation failure. Fertil. Steril. 2021, 116, 181–188. [Google Scholar] [CrossRef]
- Zeng, H.; Hu, L.; Xie, H.; Ma, W.; Quan, S. Polymorphisms of vascular endothelial growth factor and recurrent implantation failure: A systematic review and meta-analysis. Arch. Gynecol. Obstet. 2021, 304, 297–307. [Google Scholar] [CrossRef]
- Dumont, P.; Leu, J.I.-J.; Iii, A.C.D.P.; George, D.L.; Murphy, M. The codon 72 polymorphic variants of p53 have markedly different apoptotic potential. Nat. Genet. 2003, 33, 357–365. [Google Scholar] [CrossRef]
- Nowak, I.; Wilczyńska, K.; Radwan, P.; Wiśniewski, A.; Krasiński, R.; Radwan, M.; Wilczyński, J.R.; Malinowski, A.; Kuśnierczyk, P. Association of Soluble HLA-G Plasma Level and HLA-G Genetic Polymorphism With Pregnancy Outcome of Patients Undergoing in vitro Fertilization Embryo Transfer. Front. Immunol. 2020, 10, 2982. [Google Scholar] [CrossRef]
- Chen, X.; Diao, L.; Lian, R.; Qi, L.; Yu, S.; Liu, S.; Lin, S.; Xue, Z.; Zeng, Y. Potential impact of maternal vitamin D status on peripheral blood and endometrium cellular immunity in women with recurrent implantation failure. Am. J. Reprod. Immunol. 2020, 84, e13243. [Google Scholar] [CrossRef]
- Schröder-Heurich, B.; Springer, C.J.P.; Von Versen-Höynck, F. Vitamin D Effects on the Immune System from Periconception through Pregnancy. Nutrients 2020, 12, 1432. [Google Scholar] [CrossRef]
- Makrigiannakis, A.; Makrygiannakis, F.; Vrekoussis, T. Approaches to Improve Endometrial Receptivity in Case of Repeated Implantation Failures. Front. Cell Dev. Biol. 2021, 9, 613277. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zheng, L.; Zhao, D.; Xu, Y.; Wang, Y. The Role of Immune Cells in Recurrent Spontaneous Abortion. Reprod. Sci. 2021, 28, 1–13. [Google Scholar] [CrossRef]
- Ticconi, C.; Pietropolli, A.; Di Simone, N.; Piccione, E.; Fazleabas, A. Endometrial Immune Dysfunction in Recurrent Pregnancy Loss. Int. J. Mol. Sci. 2019, 20, 5332. [Google Scholar] [CrossRef]
- Tsao, F.-Y.; Wu, M.-Y.; Chang, Y.-L.; Wu, C.-T.; Ho, H.-N. M1 macrophages decrease in the deciduae from normal pregnancies but not from spontaneous abortions or unexplained recurrent spontaneous abortions. J. Formos. Med. Assoc. 2018, 117, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Nagamatsu, T.; Schust, D.J. The Contribution of Macrophages to Normal and Pathological Pregnancies. Am. J. Reprod. Immunol. 2010, 63, 460–471. [Google Scholar] [CrossRef]
- Mori, A.; Nishi, H.; Sasaki, T.; Nagamitsu, Y.; Kawaguchi, R.; Okamoto, A.; Kuroda, M.; Isaka, K. HLA-G expression is regulated by miR-365 in trophoblasts under hypoxic conditions. Placenta 2016, 45, 37–41. [Google Scholar] [CrossRef]
- Donoghue, J.; Paiva, P.; Teh, W.T.; Cann, L.M.; Nowell, C.; Rees, H.; Bittinger, S.; Obers, V.; Bulmer, J.N.; Stern, C.; et al. Endometrial uNK cell counts do not predict successful implantation in an IVF population. Hum. Reprod. 2019, 34, 2456–2466. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.B.; Jeelall, Y.; Pennell, C.; Hart, R.; McLean-Tooke, A.; Lucas, M. The role of immunological testing and intervention in reproductive medicine: A fertile collaboration? Am. J. Reprod. Immunol. 2018, 79, e12784. [Google Scholar] [CrossRef]
- Perrier d’Hauterive, S.; Charlet-Renard, C.; Berndt, S.; Dubois, M.; Munaut, C.; Gon, F.; Hagelstein, M.T.; Noel, A.; Hazout, A.; Foidart, J.M.; et al. Human chorionic gonadotropin and growth factors at the embryonic-endometrial interface control leukemia inhibitory factor (LIF) and interleukin 6 (IL-6) secretion by human endometrial epithelium. Hum. Reprod. 2004, 19, 2633–2643. [Google Scholar] [CrossRef]
- Achache, H.; Revel, A. Endometrial receptivity markers, the journey to successful embryo implantation. Hum. Reprod. Updat. 2006, 12, 731–746. [Google Scholar] [CrossRef]
- Strakova, Z.; Mavrogianis, P.; Meng, X.; Hastings, J.M.; Jackson, K.S.; Cameo, P.; Brudney, A.; Knight, O.; Fazleabas, A.T. In vivo infusion of interleukin-1beta and chorionic gonadotropin induces endometrial changes that mimic early pregnancy events in the baboon. Endocrinology 2005, 146, 4097–4104. [Google Scholar] [CrossRef][Green Version]
- Qiong, Z.; Jie, H.; Yonggang, W.; Bin, X.; Jing, Z.; Yanping, L. Clinical validation of pinopode as a marker of endometrial receptivity: A randomized controlled trial. Fertil. Steril. 2017, 108, 513–517.e2. [Google Scholar] [CrossRef]
- Yue, X.; Wu, L.; Hu, W. The Regulation of Leukemia Inhibitory Factor. Cancer Cell Microenviron. 2015, 2, 877. [Google Scholar] [CrossRef]
- Stewart, C.L.; Kaspar, P.; Brunet, L.J.; Bhatt, H.; Gadi, I.; Köntgen, F.; Abbondanzo, S.J. Blastocyst implantation depends on maternal expression of leukaemia inhibitory factor. Nature 1992, 359, 76–79. [Google Scholar] [CrossRef]
- Salleh, N.; Giribabu, N. Leukemia Inhibitory Factor: Roles in Embryo Implantation and in Nonhormonal Contraception. Sci. World J. 2014, 2014, 201514. [Google Scholar] [CrossRef]
- Hu, W.; Feng, Z.; Teresky, A.K.; Levine, A.J. p53 regulates maternal reproduction through LIF. Nat. Cell Biol. 2007, 450, 721–724. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, M.; Hirota, Y.; Fukui, Y.; Fujita, H.; Saito-Fujita, T.; Kaku, T.; Gebril, M.; Hirata, T.; Akaeda, S.; Hiraoka, T.; et al. Levonorgestrel Inhibits Embryo Attachment by Eliminating Uterine Induction of Leukemia Inhibitory Factor. Endocrinology 2019, 161, bqz005. [Google Scholar] [CrossRef]
- Cheng, J.-G.; Chen, J.R.; Hernandez, L.; Alvord, W.G.; Stewart, C.L. Dual control of LIF expression and LIF receptor function regulate Stat3 activation at the onset of uterine receptivity and embryo implantation. Proc. Natl. Acad. Sci. USA 2001, 98, 8680–8685. [Google Scholar] [CrossRef]
- Hiraoka, T.; Hirota, Y.; Fukui, Y.; Gebril, M.; Kaku, T.; Aikawa, S.; Hirata, T.; Akaeda, S.; Matsuo, M.; Haraguchi, H.; et al. Differential roles of uterine epithelial and stromal STAT3 coordinate uterine receptivity and embryo attachment. Sci. Rep. 2020, 10, 15523. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Kim, H.-R.; Lim, E.J.; Park, M.; Yoon, J.A.; Kim, Y.S.; Kim, E.-K.; Shin, J.-E.; Kim, J.H.; Kwon, H.; et al. Integrative Analyses of Uterine Transcriptome and MicroRNAome Reveal Compromised LIF-STAT3 Signaling and Progesterone Response in the Endometrium of Patients with Recurrent/Repeated Implantation Failure (RIF). PLoS ONE 2016, 11, e0157696. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Bartos, A.; Whitsett, J.A.; Dey, S.K. Uterine Deletion ofGp130orStat3Shows Implantation Failure with Increased Estrogenic Responses. Mol. Endocrinol. 2013, 27, 1492–1501. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Ahn, E.; Park, H.; Kim, J.; Kim, Y.; Lee, W.; Kim, N. Association between HOX Transcript Antisense RNA Single-Nucleotide Variants and Recurrent Implantation Failure. Int. J. Mol. Sci. 2021, 22, 3021. [Google Scholar] [CrossRef]
- So, K.; Lee, C.-L.; Yeung, W.S.; Lee, K.-F. Glycodelin suppresses endometrial cell migration and invasion but stimulates spheroid attachment. Reprod. Biomed. Online 2012, 24, 639–645. [Google Scholar] [CrossRef]
- Lee, C.L.; Chiu, P.C.; Lam, K.K.; Chan, R.W.; Chu, I.K.; Koistinen, R.; Koistinen, H.; Seppälä, M.; Lee, K.F.; Yeung, W.S. Glycodelin-A modulates cytokine production of peripheral blood natural killer cells. Fertil. Steril. 2010, 94, 769–771. [Google Scholar] [CrossRef]
- Lee, C.-L.; Lam, K.K.; Koistinen, H.; Seppala, M.; Kurpisz, M.; Fernandez, N.; Pang, R.T.; Yeung, W.S.; Chiu, C.N. Glycodelin-A as a paracrine regulator in early pregnancy. J. Reprod. Immunol. 2011, 90, 29–34. [Google Scholar] [CrossRef]
- Dixit, A.; Karande, A.A. Glycodelin regulates the numbers and function of peripheral natural killer cells. J. Reprod. Immunol. 2020, 137, 102625. [Google Scholar] [CrossRef]
- Lee, C.-L.; Lam, K.K.W.; Vijayan, M.; Koistinen, H.; Seppala, M.; Ng, E.; Yeung, S.B.W.; Chiu, C.N. The Pleiotropic Effect of Glycodelin-A in Early Pregnancy. Am. J. Reprod. Immunol. 2016, 75, 290–297. [Google Scholar] [CrossRef]
- Scholz, C.; Toth, B.; Brunnhuber, R.; Rampf, E.; Weissenbacher, T.; Santoso, L.; Friese, K.; Jeschke, U. Glycodelin-A induces a tolerogenic phenotype in monocyte-derived dendritic cells in vitro. Am. J. Reprod. Immunol. 2008, 60, 501–512. [Google Scholar] [CrossRef]
- Bastu, E.; Mutlu, M.F.; Yasa, C.; Dural, O.; Aytan, A.N.; Celik, C.; Buyru, F.; Yeh, J. Role of Mucin 1 and Glycodelin A in recurrent implantation failure. Fertil. Steril. 2015, 103, 1059–1064.e2. [Google Scholar] [CrossRef]
- Prapas, Y.; Prapas, N.; Jones, E.E.; Duleba, A.J.; Olive, D.L.; Chatziparasidou, A.; Vlassis, G. The window for embryo transfer in oocyte donation cycles depends on the duration of progesterone therapy. Hum. Reprod. 1998, 13, 720–723. [Google Scholar] [CrossRef]
- Klonos, E.; Katopodis, P.; Karteris, E.; Papanikolaou, E.; Tarlatzis, B.; Pados, G. Endometrial changes in estrogen and progesterone receptor expression during implantation in an oocyte donation program. Exp. Ther. Med. 2020, 20, 178. [Google Scholar] [CrossRef] [PubMed]
- Mulac-Jericevic, B.; Lydon, J.P.; DeMayo, F.; Conneely, O.M. Defective mammary gland morphogenesis in mice lacking the progesterone receptor B isoform. Proc. Natl. Acad. Sci. USA 2003, 100, 9744–9749. [Google Scholar] [CrossRef]
- Long, N.; Liu, N.; Liu, X.; Li, J.; Cai, B.; Cai, X. Endometrial expression of telomerase, progesterone, and estrogen receptors during the implantation window in patients with recurrent implantation failure. Genet. Mol. Res. 2016, 15, 49. [Google Scholar] [CrossRef] [PubMed]
- Vagnini, L.D.; Renzi, A.; Petersen, B.; Canas, M.D.C.T.; Petersen, C.G.; Mauri, A.L.; Mattila, M.C.; Ricci, J.; Dieamant, F.; Oliveira, J.B.A.; et al. Association between estrogen receptor 1 (ESR1) and leukemia inhibitory factor (LIF) polymorphisms can help in the prediction of recurrent implantation failure. Fertil. Steril. 2019, 111, 527–534. [Google Scholar] [CrossRef]
- Suh, E.-K.; Yang, A.; Kettenbach, A.; Bamberger, T.C.; Michaelis, A.H.; Zhu, Z.; Elvin, J.A.; Bronson, R.T.; Crum, C.P.; McKeon, F. p63 protects the female germ line during meiotic arrest. Nat. Cell Biol. 2006, 444, 624–628. [Google Scholar] [CrossRef]
- Ravi, R.; Mookerjee, B.; Bhujwalla, Z.M.; Sutter, C.H.; Artemov, D.; Zeng, Q.; Dillehay, L.E.; Madan, A.; Semenza, G.L.; Bedi, A. Regulation of tumor angiogenesis by p53-induced degradation of hypoxia-inducible factor 1α. Genome Res. 2000, 14, 34–44. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC316350/ (accessed on 15 August 2021).
- Guo, X.; Yi, H.; Li, T.; Wang, Y.; Wang, H.; Chen, X. Role of Vascular Endothelial Growth Factor (VEGF) in Human Embryo Implantation: Clinical Implications. Biomolecules 2021, 11, 253. [Google Scholar] [CrossRef]
- Okada, H.; Okamoto, R.; Tsuzuki, T.; Tsuji, S.; Yasuda, K.; Kanzaki, H. Progestins inhibit estradiol-induced vascular endothelial growth factor and stromal cell–derived factor 1 in human endometrial stromal cells. Fertil. Steril. 2011, 96, 786–791. [Google Scholar] [CrossRef]
- Selvaraj, S.K.; Giri, R.K.; Perelman, N.; Johnson, C.; Malik, P.; Kalra, V.K. Mechanism of monocyte activation and expression of proinflammatory cytochemokines by placenta growth factor. Blood 2003, 102, 1515–1524. [Google Scholar] [CrossRef]
- Tayade, C.; Hilchie, D.; He, H.; Fang, Y.; Moons, L.; Carmeliet, P.; Foster, R.A.; Croy, B.A. Genetic Deletion of Placenta Growth Factor in Mice Alters Uterine NK Cells. J. Immunol. 2007, 178, 4267–4275. [Google Scholar] [CrossRef]
- Lash, G.E.; Innes, B.A.; Drury, J.A.; Robson, S.C.; Quenby, S.; Bulmer, J.N. Localization of angiogenic growth factors and their receptors in the human endometrium throughout the menstrual cycle and in recurrent miscarriage. Hum. Reprod. 2012, 27, 183–195. [Google Scholar] [CrossRef]
- Sadekova, O.N.; Nikitina, L.A.; Rashidov, T.N.; Voloschuk, I.N.; Samokhodskaya, L.M.; Demidova, E.M.; Tkachuk, V.A. Luteal phase defect is associated with impaired VEGF mRNA expression in the secretory phase endometrium. Reprod. Biol. 2015, 15, 65–68. [Google Scholar] [CrossRef]
- Benkhalifa, M.; Zidi, W.; Bahri, H.; Mahjoub, S.; Boudhraa, K.; Sanhaji, H.; Khorsi-Cauet, H.; Feki, M.; Benkhalifa, M.; Allal-Elasmi, M. Circulating MMP-7 and VEGF as potential predictive biomarkers for recurrent implantation failures. Zygote 2021, 9, 1–7. [Google Scholar] [CrossRef]
- Chen, X.; Man, G.C.W.; Liu, Y.; Wu, F.; Huang, J.; Li, T.C.; Wang, C.C. Physiological and pathological angiogenesis in endometrium at the time of embryo implantation. Am. J. Reprod. Immunol. 2017, 78, e12693. [Google Scholar] [CrossRef]
- Bansal, R.; Ford, B.; Bhaskaran, S.; Thum, M.; Bansal, A. Elevated Levels of Serum Vascular Endothelial Growth Factor-A Are Not Related to NK Cell Parameters in Recurrent IVF Failure. J. Reprod. Infertil. 2017, 18, 280–287. [Google Scholar]
- Van Bergen, T.; Etienne, I.; Cunningham, F.; Moons, L.; Schlingemann, R.O.; Feyen, J.H.; Stitt, A.W. The role of placental growth factor (PlGF) and its receptor system in retinal vascular diseases. Prog. Retin. Eye Res. 2019, 69, 116–136. [Google Scholar] [CrossRef]
- Albonici, L.; Benvenuto, M.; Focaccetti, C.; Cifaldi, L.; Miele, M.T.; Limana, F.; Manzari, V.; Bei, R. PlGF Immunological Impact during Pregnancy. Int. J. Mol. Sci. 2020, 21, 8714. [Google Scholar] [CrossRef]
- Ukah, U.V.; Payne, B.A.; Hutcheon, J.A.; Chappell, L.; Seed, P.T.; Conti-Ramsden, F.I.; Ansermino, J.M.; Magee, L.A.; Von Dadelszen, P. Placental growth factor for the prognosis of women with preeclampsia (fullPIERS model extension): Context matters. BMC Pregnancy Childbirth 2020, 20, 668. [Google Scholar] [CrossRef] [PubMed]
- Li, X.F.; Charnock-Jones, D.S.; Zhang, E.; Hiby, S.; Malik, S.; Day, K.; Licence, D.; Bowen, J.M.; Gardner, L.; King, A.; et al. Angiogenic Growth Factor Messenger Ribonucleic Acids in Uterine Natural Killer Cells1. J. Clin. Endocrinol. Metab. 2001, 86, 1823–1834. [Google Scholar] [CrossRef] [PubMed]
- Rah, H.; Chung, K.W.; Ko, K.H.; Kim, E.S.; Kim, J.O.; Sakong, J.H.; Kim, J.H.; Lee, W.S.; Kim, N.K. miR-27a and miR-449b polymorphisms associated with a risk of idiopathic recurrent pregnancy loss. PLoS ONE 2017, 12, e0177160. [Google Scholar] [CrossRef] [PubMed]
- Qublan, H.S.; Eid, S.S.; Ababneh, H.A.; Amarin, Z.O.; Smadi, A.Z.; Al-Khafaji, F.F.; Khader, Y. Acquired and inherited thrombophilia: Implication in recurrent IVF and embryo transfer failure. Hum. Reprod. 2006, 21, 2694–2698. [Google Scholar] [CrossRef]
- Simur, A.; Ozdemir, S.; Acar, H.; Colakoğlu, M.C.; Görkemli, H.; Balci, O.; Nergis, S. Repeated in vitro Fertilization Failure and Its Relation with Thrombophilia. Gynecol. Obstet. Investig. 2009, 67, 109–112. [Google Scholar] [CrossRef]
- Zeng, H.; He, D.; Zhao, Y.; Liu, N.G.; Xie, H. Association between MTHFR polymorphisms (MTHFR C677T, MTHFR A1298C) and recurrent implantation failure: A systematic review and meta-analysis. Arch. Gynecol. Obstet. 2021, 303, 1089–1098. [Google Scholar] [CrossRef]
- Bautch, V.L. VEGF-Directed Blood Vessel Patterning: From Cells to Organism. Cold Spring Harb. Perspect. Med. 2012, 2, a006452. [Google Scholar] [CrossRef]
- Bond, G.L.; Hu, W.; Bond, E.E.; Robins, H.; Lutzker, S.G.; Arva, N.C.; Bargonetti, J.; Bartel, F.; Taubert, H.; Wuerl, P.; et al. A Single Nucleotide Polymorphism in the MDM2 Promoter Attenuates the p53 Tumor Suppressor Pathway and Accelerates Tumor Formation in Humans. Cell 2004, 119, 591–602. [Google Scholar] [CrossRef]
- Kang, H.-J.; Feng, Z.; Sun, Y.; Atwal, G.; Murphy, M.; Rebbeck, T.R.; Rosenwaks, Z.; Levine, A.J.; Hu, W. Single-nucleotide polymorphisms in the p53 pathway regulate fertility in humans. Proc. Natl. Acad. Sci. USA 2009, 106, 9761–9766. [Google Scholar] [CrossRef]
- Lledó, B.; Turienzo, A.; Ortiz, J.A.; Morales, R.; Ten, J.; Llacer, J.; Bernabeu, R. Negative effect of P72 polymorphism on p53 gene in IVF outcome in patients with repeated implantation failure and pregnancy loss. J. Assist. Reprod. Genet. 2013, 31, 169–172. [Google Scholar] [CrossRef][Green Version]
- Mohammadzadeh, M.; Ghorbian, S.; Nouri, M. Evaluation of clinical utility of P53 gene variations in repeated implantation failure. Mol. Biol. Rep. 2019, 46, 2885–2891. [Google Scholar] [CrossRef]
- Turienzo, A.; Lledó, B.; Ortiz, J.A.; Morales, R.; Sanz, J.; Llácer, J.; Bernabeu, R. Prevalence of candidate single nucleotide polymorphisms on p53, IL-11, IL-10, VEGF and APOE in patients with repeated implantation failure (RIF) and pregnancy loss (RPL). Hum. Fertil. 2018, 23, 117–122. [Google Scholar] [CrossRef]
- Fan, W.; Huang, Z.; Li, S.; Xiao, Z. The HLA-G 14-bp polymorphism and recurrent implantation failure: A meta-analysis. J. Assist. Reprod. Genet. 2017, 34, 1559–1565. [Google Scholar] [CrossRef]
- Amabebe, E.; Anumba, D.O.C. The Vaginal Microenvironment: The Physiologic Role of Lactobacilli. Front. Med. 2018, 5, 181. [Google Scholar] [CrossRef]
- Nuriel-Ohayon, M.; Neuman, H.; Koren, O. Microbial Changes during Pregnancy, Birth, and Infancy. Front. Microbiol. 2016, 7, 1031. [Google Scholar] [CrossRef] [PubMed]
- Schoenmakers, S.; Laven, J. The vaginal microbiome as a tool to predict IVF success. Curr. Opin. Obstet. Gynecol. 2020, 32, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Tachedjian, G.; Aldunate, M.; Bradshaw, C.S.; Cone, R.A. The role of lactic acid production by probiotic Lactobacillus species in vaginal health. Res. Microbiol. 2017, 168, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Luna, Y.; Yu, P.; Fan, H. Lactobacilli Inactivate Chlamydia trachomatis through Lactic Acid but Not H2O2. PLoS ONE 2014, 9, e107758. [Google Scholar] [CrossRef] [PubMed]
- Cadieux, P.A.; Burton, J.; Devillard, E.; Reid, G. Lactobacillus by-products inhibit the growth and virulence of uropathogenic Escherichia coli. J. Physiol. Pharmacol. 2009, 60, 13–18. [Google Scholar]
- Alakomi, H.-L.; Skyttä, E.; Saarela, M.; Mattila-Sandholm, T.; Latva-Kala, K.; Helander, I.M. Lactic Acid Permeabilizes Gram-Negative Bacteria by Disrupting the Outer Membrane. Appl. Environ. Microbiol. 2000, 66, 2001–2005. [Google Scholar] [CrossRef]
- Aldunate, M.; Tyssen, D.; Johnson, A.; Zakir, T.; Sonza, S.; Moench, T.; Cone, R.; Tachedjian, G. Vaginal concentrations of lactic acid potently inactivate HIV. J. Antimicrob. Chemother. 2013, 68, 2015–2025. [Google Scholar] [CrossRef]
- Brotman, R.M. Vaginal microbiome and sexually transmitted infections: An epidemiologic perspective. J. Clin. Investig. 2011, 121, 4610–4617. [Google Scholar] [CrossRef] [PubMed]
- Chase, D.; Goulder, A.; Zenhausern, F.; Monk, B.; Herbst-Kralovetz, M. The vaginal and gastrointestinal microbiomes in gynecologic cancers: A review of applications in etiology, symptoms and treatment. Gynecol. Oncol. 2015, 138, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Hyman, R.W.; Herndon, C.N.; Jiang, H.; Palm, C.; Fukushima, M.; Bernstein, D.; Vo, K.C.; Zelenko, Z.; Davis, R.W.; Giudice, L.C. The dynamics of the vaginal microbiome during infertility therapy with in vitro fertilization-embryo transfer. J. Assist. Reprod. Genet. 2012, 29, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Moreno, I.; Codoñer, F.M.; Vilella, F.; Valbuena, D.; Martinez-Blanch, J.F.; Jimenez-Almazán, J.; Alonso, R.; Alamá, P.; Remohí, J.; Pellicer, A.; et al. Evidence that the endometrial microbiota has an effect on implantation success or failure. Am. J. Obstet. Gynecol. 2016, 215, 684–703. [Google Scholar] [CrossRef]
- Punzón-Jiménez, P.; Labarta, E. The impact of the female genital tract microbiome in women health and reproduction: A review. J. Assist. Reprod. Genet. 2021, 38, 1–23. [Google Scholar] [CrossRef]
- Ahmadi, M.; Abdolmohammadi-Vahid, S.; Ghaebi, M.; Aghebati-Maleki, L.; Dolati, S.; Farzadi, L.; Ghasemzadeh, A.; Hamdi, K.; Younesi, V.; Nouri, M.; et al. Regulatory T cells improve pregnancy rate in RIF patients after additional IVIG treatment. Syst. Biol. Reprod. Med. 2017, 63, 350–359. [Google Scholar] [CrossRef]
- Sung, N.; Han, A.R.; Park, C.W.; Park, D.W.; Park, J.C.; Kim, N.Y.; Lim, K.S.; Shin, J.E.; Joo, C.W.; Lee, S.E.; et al. Intravenous immunoglobulin G in women with reproductive failure: The Korean Society for Reproductive Immunology practice guidelines. Clin. Exp. Reprod. Med. 2017, 44, 1–7. [Google Scholar] [CrossRef]
- Kolanska, K.; Bendifallah, S.; Cohen, J.; Placais, L.; Selleret, L.; Johanet, C.; Suner, L.; Delhommeau, F.; Chabbert-Buffet, N.; Darai, E.; et al. Unexplained recurrent implantation failures: Predictive factors of pregnancy and therapeutic management from a French multicentre study. J. Reprod. Immunol. 2021, 145, 103313. [Google Scholar] [CrossRef] [PubMed]
- Sadeghpour, S.; Berenji, M.G.; Nazarian, H.; Ghasemnejad, T.; Nematollahi, M.H.; Abroon, S.; Paktinat, S.; Khoei, H.H.; Berenji, H.G.; Novin, M.G. Effects of treatment with hydroxychloroquine on the modulation of Th17/Treg ratio and pregnancy outcomes in women with recurrent implantation failure: Clinical trial. Immunopharmacol. Immunotoxicol. 2020, 42, 632–642. [Google Scholar] [CrossRef]
- Shapiro, B.S.; Daneshmand, S.T.; Desai, J.; Garner, F.C.; Aguirre, M.; Hudson, C. The risk of embryo-endometrium asynchrony increases with maternal age after ovarian stimulation and IVF. Reprod. Biomed. Online 2016, 33, 50–55. [Google Scholar] [CrossRef]
- National Centers for Disease Control and Prevention and Health Promotion of the Centers for Disease Control and Prevention in Consultation with the American Society for Reproductive Medicine and the Society for Assisted Reproductive Technology. Assisted Reproductive Technology National Summary Report; US Departmentof Health and Human Services: Atlanta, GA, USA, 2017.
- Orvieto, R.; Meltcer, S.; Nahum, R.; Rabinson, J.; Anteby, E.Y.; Ashkenazi, J. The influence of body mass index on in vitro fertilization outcome. Int. J. Gynecol. Obstet. 2008, 104, 53–55. [Google Scholar] [CrossRef] [PubMed]
- Moragianni, V.A.; Jones, S.-M.L.; Ryley, D.A. The effect of body mass index on the outcomes of first assisted reproductive technology cycles. Fertil. Steril. 2012, 98, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Waylen, A.; Metwally, M.; Jones, G.; Wilkinson, A.; Ledger, W. Effects of cigarette smoking upon clinical outcomes of assisted reproduction: A meta-analysis. Hum. Reprod. Updat. 2008, 15, 31–44. [Google Scholar] [CrossRef]
- Anblagan, D.; Jones, N.W.; Costigan, C.; Parker, A.J.J.; Allcock, K.; Aleong, R.; Coyne, L.H.; Deshpande, R.; Raine-Fenning, N.; Bugg, G.; et al. Maternal Smoking during Pregnancy and Fetal Organ Growth: A Magnetic Resonance Imaging Study. PLoS ONE 2013, 8, e67223. [Google Scholar] [CrossRef]
- Künzle, R.; Mueller, M.; Hänggi, W.; Birkhäuser, M.H.; Drescher, H.; Bersinger, N.A. Semen quality of male smokers and nonsmokers in infertile couples. Fertil. Steril. 2003, 79, 287–291. [Google Scholar] [CrossRef]
- Nepomnaschy, P.A.; Welch, K.B.; McConnell, D.S.; Low, B.S.; Strassmann, B.I.; England, B.G. Cortisol levels and very early pregnancy loss in humans. Proc. Natl. Acad. Sci. USA 2006, 103, 3938–3942. [Google Scholar] [CrossRef] [PubMed]
- Paskulin, D.D.; Paixão-Côrtes, V.R.; Hainaut, P.; Bortolini, M.C.; Ashton-Prolla, P. The TP53 fertility network. Genet. Mol. Biol. 2012, 35, 939–946. [Google Scholar] [CrossRef] [PubMed][Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).