
MicroRNA-126b-5p Exacerbates Development of Adipose Tissue and Diet-Induced Obesity

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. miR-126b-5p Was Associated with Adipogenesis
2.2. miR-126b-5p Promotes 3T3-L1 Preadipocyte Proliferation and Differentiation
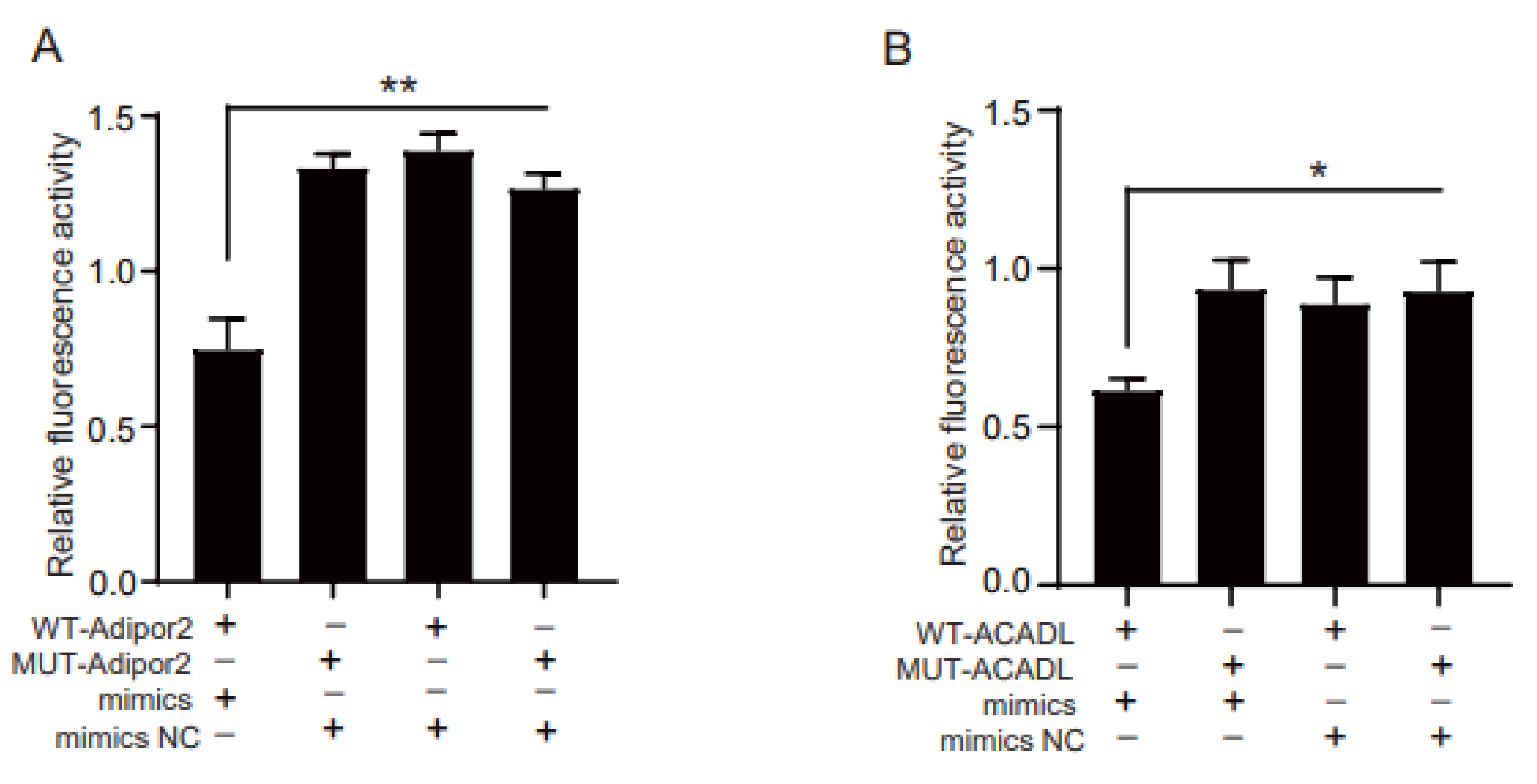
2.3. Adipor2 and ACADL Are Direct Target Genes of miR-126b-5p
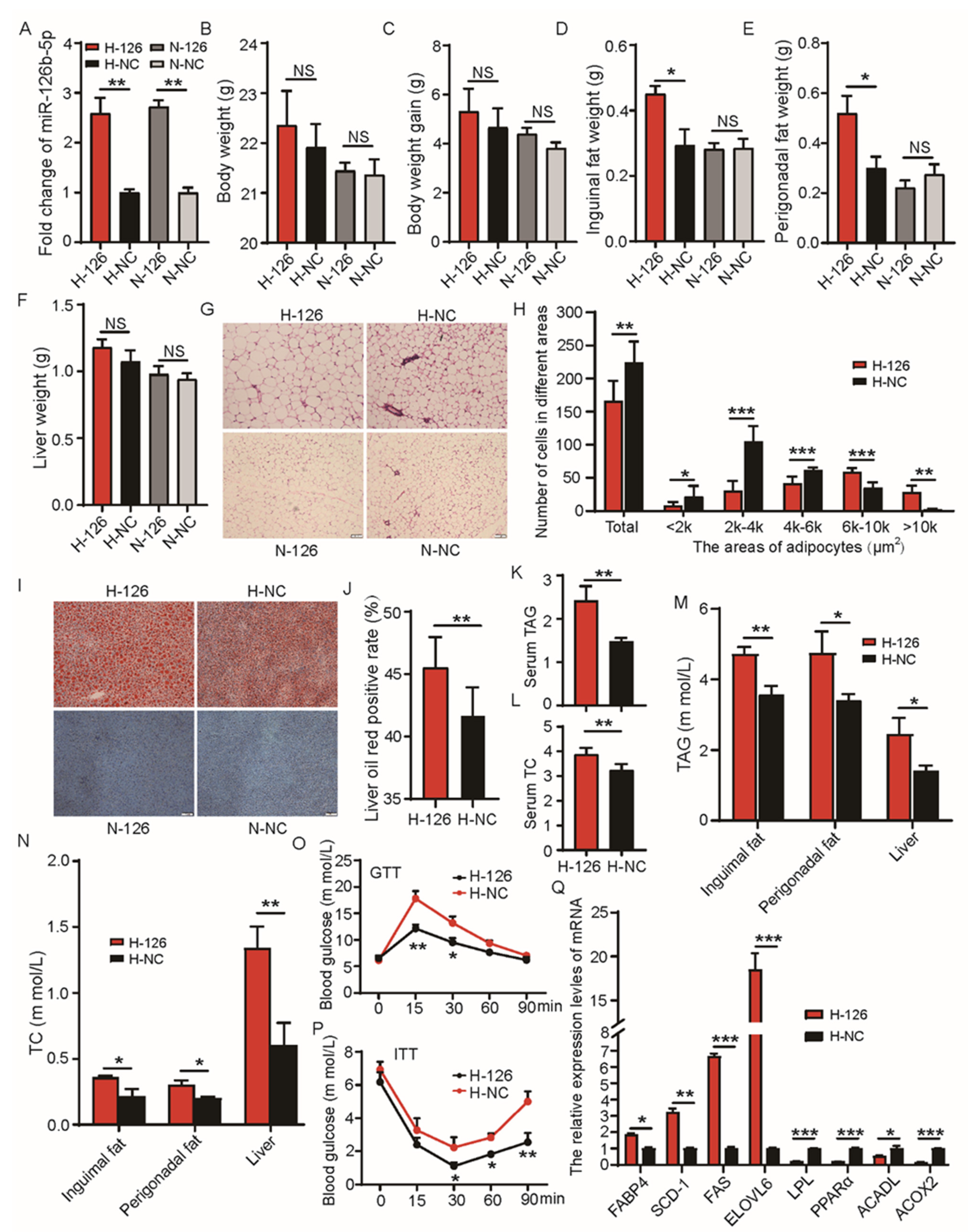
2.4. miR-126b-5p Exacerbated Obesity Induced by a High-Fat Diet
3. Discussion
4. Materials and Methods
4.1. Animal Treatment and Sample Collection
4.2. Cell Culture and Transfection
4.3. Proliferation Analysis
4.4. Triglyceride, Total Cholesterol Assay, and Cell Oil Red O Staining
4.5. Tissue H&E and Oil Red O Staining
4.6. Glucose Tolerance Test (GTT) and Insulin Tolerance Test (ITT)
4.7. Analysis of Fatty Acids
4.8. Quantitative Real-Time PCR
4.9. Luciferase Reporter Assay
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, G.M.; Danaei, G.; Farzadfar, F.; Stevens, G.A.; Woodward, M.; Wormser, D.; Kaptoge, S.; Whitlock, G.; Qiao, Q.; Lewington, S. The Age-Specific Quantitative Effects of Metabolic Risk Factors on Cardiovascular Diseases and Diabetes: A Pooled Analysis. PLoS ONE 2013, 8, e65174. [Google Scholar] [CrossRef]
- Gregor, M.F.; Hotamisligil, G.S. Inflammatory mechanisms in obesity. Annu. Rev. Immunol. 2011, 29, 415–445. [Google Scholar] [CrossRef] [Green Version]
- Malik, V.S.; Popkin, B.M.; Bray, G.A.; Després, J.-P.; Hu, F.B. Sugar-sweetened beverages, obesity, type 2 diabetes mellitus, and cardiovascular disease risk. Circulation 2010, 121, 1356–1364. [Google Scholar] [CrossRef]
- Pereira, S.S.; Alvarez-Leite, J.I. Low-grade inflammation, obesity, and diabetes. Curr. Obes. Rep. 2014, 3, 422–431. [Google Scholar] [PubMed]
- He, Y.; Pan, A.; Wang, Y.; Yang, Y.; Xu, J.; Zhang, Y.; Liu, D.; Wang, Q.; Shen, H.; Zhang, Y. Prevalence of overweight and obesity in 15.8 million men aged 15–49 years in rural China from 2010 to 2014. Sci. Rep. 2017, 7, 5012. [Google Scholar] [CrossRef] [Green Version]
- Meisinger, C.; Ezzati, M.; Di Cesare, M. Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar]
- Zhang, P.; Du, J.; Wang, L.; Niu, L.; Zhao, Y.; Tang, G.; Jiang, Y.; Shuai, S.; Bai, L.; Li, X. MicroRNA-143a-3p modulates preadipocyte proliferation and differentiation by targeting MAPK7. Biomed. Pharmacother. 2018, 108, 531–539. [Google Scholar]
- Fan, Y.; Gan, M.; Tan, Y.; Chen, L.; Shen, L.; Niu, L.; Liu, Y.; Tang, G.; Jiang, Y.; Li, X. Mir-152 regulates 3T3-L1 preadipocyte proliferation and differentiation. Molecules 2019, 24, 3379. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Chen, F.-F.; Ge, J.; Zhu, J.-Y.; Shi, X.-E.; Li, X.; Yu, T.-Y.; Chu, G.-Y.; Yang, G.-S. miR-429 inhibits differentiation and promotes proliferation in porcine preadipocytes. Int. J. Mol. Sci. 2016, 17, 2047. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wu, H.; Luo, Z.; Xia, Y.; Guan, J.; Wang, T.; Gu, Y.; Chen, L.; Zhang, K.; Ma, J. An atlas of DNA methylomes in porcine adipose and muscle tissues. Nat. Commun. 2012, 3, 1–11. [Google Scholar] [CrossRef]
- Sun, Z.; Ou, C.; Liu, J.; Chen, C.; Zhou, Q.; Yang, S.; Li, G.; Wang, G.; Song, J.; Li, Z. YAP1-induced MALAT1 promotes epithelial–mesenchymal transition and angiogenesis by sponging miR-126-5p in colorectal cancer. Oncogene 2019, 38, 2627–2644. [Google Scholar] [CrossRef] [Green Version]
- Schober, A.; Nazari-Jahantigh, M.; Wei, Y.; Bidzhekov, K.; Gremse, F.; Grommes, J.; Megens, R.T.; Heyll, K.; Noels, H.; Hristov, M. MicroRNA-126-5p promotes endothelial proliferation and limits atherosclerosis by suppressing Dlk1. Nat. Med. 2014, 20, 368–376. [Google Scholar] [CrossRef] [Green Version]
- Xue, S.; Liu, D.; Zhu, W.; Su, Z.; Zhang, L.; Zhou, C.; Li, P. Circulating MiR-17-5p, MiR-126-5p and MiR-145-3p are novel biomarkers for diagnosis of acute myocardial infarction. Front. Physiol. 2019, 10, 123. [Google Scholar] [CrossRef] [Green Version]
- Dehghani, M.; Zarch, S.M.A.; Mehrjardi, M.Y.V.; Nazari, M.; Babakhanzadeh, E.; Ghadimi, H.; Zeinali, F.; Talebi, M. Evaluation of miR-181b and miR-126-5p expression levels in T2DM patients compared to healthy individuals: Relationship with NF-κB gene expression. Endocrinol. Diabetes Nutr. 2020, 67, 454–460. [Google Scholar] [CrossRef]
- Dashty, M. A quick look at biochemistry: Carbohydrate metabolism. Clin. Biochem. 2013, 46, 1339–1352. [Google Scholar] [CrossRef]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose tissue dysfunction as determinant of obesity-associated metabolic complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [Green Version]
- Caruso, J.A.; Duong, M.T.; Carey, J.P.; Hunt, K.K.; Keyomarsi, K. Low-molecular-weight cyclin E in human cancer: Cellular consequences and opportunities for targeted therapies. Cancer Res. 2018, 78, 5481–5491. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Hannon, G.J.; Zhang, H.; Casso, D.; Kobayashi, R.; Beach, D. p21 is a universal inhibitor of cyclin kinases. Nature 1993, 366, 701–704. [Google Scholar] [CrossRef]
- El-Deiry, W.S.; Tokino, T.; Velculescu, V.E.; Levy, D.B.; Parsons, R.; Trent, J.M.; Lin, D.; Mercer, W.E.; Kinzler, K.W.; Vogelstein, B. WAF1, a potential mediator of p53 tumor suppression. Cell 1993, 75, 817–825. [Google Scholar] [CrossRef]
- Dulić, V.; Kaufmann, W.K.; Wilson, S.J.; Tisty, T.D.; Lees, E.; Harper, J.W.; Elledge, S.J.; Reed, S.I. p53-dependent inhibition of cyclin-dependent kinase activities in human fibroblasts during radiation-induced G1 arrest. Cell 1994, 76, 1013–1023. [Google Scholar] [CrossRef]
- Christy, R.; Yang, V.; Ntambi, J.; Geiman, D.; Landschulz, W.; Friedman, A.; Nakabeppu, Y.; Kelly, T.; Lane, M. Differentiation-induced gene expression in 3T3-L1 preadipocytes: CCAAT/enhancer binding protein interacts with and activates the promoters of two adipocyte-specific genes. Genes Dev. 1989, 3, 1323–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, J.; Moller, D.E. The mechanisms of action of PPARs. Annu. Rev. Med. 2002, 53, 409–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Liu, D. Hydrodynamic delivery of adiponectin and adiponectin receptor 2 gene blocks high-fat diet-induced obesity and insulin resistance. Gene Ther. 2013, 20, 846–852. [Google Scholar] [CrossRef] [Green Version]
- Tomita, K.; Oike, Y.; Teratani, T.; Taguchi, T.; Noguchi, M.; Suzuki, T.; Mizutani, A.; Yokoyama, H.; Irie, R.; Sumimoto, H. Hepatic AdipoR2 signaling plays a protective role against progression of nonalcoholic steatohepatitis in mice. Hepatology 2008, 48, 458–473. [Google Scholar] [CrossRef]
- Liu, Y.; Michael, M.D.; Kash, S.; Bensch, W.R.; Monia, B.P.; Murray, S.F.; Otto, K.A.; Syed, S.K.; Bhanot, S.; Sloop, K.W. Deficiency of adiponectin receptor 2 reduces diet-induced insulin resistance but promotes type 2 diabetes. Endocrinology 2007, 148, 683–692. [Google Scholar] [CrossRef]
- Kraus, M.; Greither, T.; Wenzel, C.; Bräuer-Hartmann, D.; Wabitsch, M.; Behre, H.M. Inhibition of adipogenic differentiation of human SGBS preadipocytes by androgen-regulated microRNA miR-375. Mol. Cell. Endocrinol. 2015, 414, 177–185. [Google Scholar] [CrossRef]
- Chegary, M.; te Brinke, H.; Ruiter, J.P.; Wijburg, F.A.; Stoll, M.S.; Minkler, P.E.; van Weeghel, M.; Schulz, H.; Hoppel, C.L.; Wanders, R.J. Mitochondrial long chain fatty acid β-oxidation in man and mouse. Biochim. Biophys. Acta 2009, 1791, 806–815. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Liu, Z.-X.; Choi, C.S.; Tian, L.; Kibbey, R.; Dong, J.; Cline, G.W.; Wood, P.A.; Shulman, G.I. Mitochondrial dysfunction due to long-chain Acyl-CoA dehydrogenase deficiency causes hepatic steatosis and hepatic insulin resistance. Proc. Natl. Acad. Sci. USA 2007, 104, 17075–17080. [Google Scholar] [CrossRef] [Green Version]
- Bakermans, A.J.; Geraedts, T.R.; van Weeghel, M.; Denis, S.; João Ferraz, M.; Aerts, J.M.; Aten, J.; Nicolay, K.; Houten, S.M.; Prompers, J.J. Fasting-induced myocardial lipid accumulation in long-chain acyl-CoA dehydrogenase knockout mice is accompanied by impaired left ventricular function. Circ. Cardiovasc. Imaging 2011, 4, 558–565. [Google Scholar] [CrossRef] [Green Version]
- Cox, K.B.; Liu, J.; Tian, L.; Barnes, S.; Yang, Q.; Wood, P.A. Cardiac hypertrophy in mice with long-chain acyl-CoA dehydrogenase or very long-chain acyl-CoA dehydrogenase deficiency. Lab. Investig. 2009, 89, 1348–1354. [Google Scholar] [CrossRef] [Green Version]
- Kimura, I.; Ichimura, A.; Ohue-Kitano, R.; Igarashi, M. Free fatty acid receptors in health and disease. Physiol. Rev. 2020, 100, 171–210. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Ntambi, J.M.; Friedman, J.M. Stearoyl-CoA desaturase-1 and the metabolic syndrome. Curr. Drug Targets Immune Endocr. Metabol. Disord. 2003, 3, 271–280. [Google Scholar] [CrossRef]
- Matsuzaka, T.; Shimano, H. Elovl6: A new player in fatty acid metabolism and insulin sensitivity. J. Mol. Med. 2009, 87, 379–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Shen, L.; Tan, Z.; Zhang, P.; Zhao, X.; Xu, Y.; Yang, Q.; Ma, J.; Jiang, A.A.; Tang, G. Betaine supplementation enhances lipid metabolism and improves insulin resistance in mice fed a high-fat diet. Nutrients 2018, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Sang, L.-J.; Ju, H.-Q.; Liu, G.-P.; Tian, T.; Ma, G.-L.; Lu, Y.-X.; Liu, Z.-X.; Pan, R.-L.; Li, R.-H.; Piao, H.-l. LncRNA CamK-A regulates Ca2+-signaling-mediated tumor microenvironment remodeling. Mol. Cell. 2018, 72, 71–83.e77. [Google Scholar] [CrossRef] [Green Version]
- Hengge, U.R.; Walker, P.S.; Vogel, J.C. Expression of naked DNA in human, pig, and mouse skin. J. Clin. Investig. 1996, 97, 2911–2916. [Google Scholar] [CrossRef]
- Bai, H.; Lester, G.M.S.; Petishnok, L.C.; Dean, D.A. Cytoplasmic transport and nuclear import of plasmid DNA. Biosci. Rep. 2017, 37, BSR20160616. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | Concentration (μg/mL) | Up/Down | |
|---|---|---|---|
| Mimics NC | miR-126b-5p Mimics | ||
| C6:0 | 0 | 0 | ---- |
| C8:0 | 0 | 0 | ---- |
| C10:0 | 0 | 0.01 ± 0.0004 | Up |
| C11:0 | 0 | 0 | ---- |
| C12:0 | 0 | 0.02 ± 0.002 | Up |
| C13:0 | 0.01 ± 0.00 | 0.02 ± 0.001 | Up *** |
| C14:0 | 0.54 ± 0.05 | 1.23 ± 0.04 | Up *** |
| C14:1 | 0 | 0 | ---- |
| C15:0 | 0.46 ± 0.05 | 1.01 ± 0.02 | Up *** |
| C15:1 | 0 | 0 | ---- |
| C16:0 | 8.29 ± 0.38 | 18.16 ± 0.13 | Up *** |
| C16:1 | 4.57 ± 0.44 | 11.01 ± 0.55 | Up *** |
| C17:0 | 0.42 ± 0.04 | 0.77 ± 0.02 | Up *** |
| C17:1 | 0.54 ± 0.05 | 1.10 ± 0.09 | Up *** |
| C18:0 | 3.60 ± 0.76 | 5.02 ± 0.03 | Up * |
| C18:1n9t | 0 | 0 | ---- |
| C18:1n9c | 5.20 ± 0.71 | 10.02 ± 0.56 | Up *** |
| C18:2n6t | 0 | 0 | ---- |
| C18:2n6c | 0.52 ± 0.04 | 0.55 ± 0.02 | Up NS |
| C18:3n6 | 0.25 ± 0.001 | 0.26 ± 0.01 | Up * |
| C18:3n3 | 0 | 0 | ---- |
| C20:0 | 0.20 ± 0.01 | 0.21 ± 0.01 | Up NS |
| C20:1 | 0 | 0 | ---- |
| C20:2 | 0.68 ± 0.0005 | 0.66 ± 0.05 | Down NS |
| C21:0 | 0 | 0 | ---- |
| C20:3n6 | 0.484 ± 0.03 | 0.485 ± 0.02 | Up NS |
| C20:4n6 | 1.61 ± 0.23 | 1.65 ± 0.11 | Up NS |
| C20:3n3 | 0 | 0 | ---- |
| C20:5n3 | 0.53 ± 0.05 | 0.55 ± 0.04 | Up NS |
| C22:0 | 0.168 ± 0.005 | 0.174 ± 0.003 | Up NS |
| C22:1n9 | 0 | 0 | ---- |
| C22:2n6 | 0 | 0 | ---- |
| C23:0 | 0 | 0 | ---- |
| C24:0 | 0.25 ± 0.01 | 0.26 ± 0.005 | Up NS |
| C24:1 | 0 | 0 | ---- |
| C22:6 | 0.846 ± 0.05 | 0.853 ± 0.03 | Up NS |
| total | 29.17 ± 2.60 | 54.02 ± 0.97 | Up *** |
| SFA | 13.94 ± 1.09 | 26.88 ± 0.02 | Up *** |
| MUFA | 10.3 ± 1.15 | 22.13 ± 1.20 | Up *** |
| PUFA | 4.92 ± 0.40 | 5.01 ± 0.25 | Up NS |
| C16:0/C16:1 | 1.82 ± 0.090 | 1.65 ± 0.094 | Down NS |
| C16:0/C18:0 | 2.37 ± 0.49 | 3.62 ± 0.044 | Up * |
| C18:0/C18:1 | 0.69 ± 0.057 | 0.50 ± 0.026 | Down ** |
| C16:1/C18:1 | 0.89 ± 0.074 | 1.10 ± 0.0070 | Up ** |
| SFA/total | 0.48 ± 0.0065 | 0.50 ± 0.0091 | Up * |
| MUFA/total | 0.35 ± 0.0081 | 0.41 ± 0.015 | Up ** |
| PUFA/total | 0.17 ± 0.0068 | 0.093 ± 0.0061 | Down *** |
| Fatty Acid | Concentration (μg/g) | Up/Down | |
|---|---|---|---|
| H-NC | H-126 | ||
| C6:0 | 27.84 ± 2.75 | 28.85 ± 0.66 | NS |
| C8:0 | 9.84 ± 0.19 | 11.82 ± 0.80 | Up * |
| C10:0 | 49.24 ± 2.76 | 68.67 ± 3.51 | Up ** |
| C11:0 | 1.04 ± 0.14 | 1.19 ± 0.02 | NS |
| C12:0 | 299.48 ± 25.58 | 335.03 ± 35.38 | NS |
| C13:0 | 6.11 ± 0.74 | 6.86 ± 0.02 | NS |
| C14:0 | 6586.37 ± 365.78 | 7352.71 ± 133.51 | Up * |
| C14:1 | 320.40 ± 29.42 | 342.04 ± 5.08 | NS |
| C15:0 | 341.88 ± 11.10 | 333.69 ± 5.41 | NS |
| C15:1 | 0.75 ± 0.27 | 2.01 ± 0.13 | Up ** |
| C16:0 | 67,993.57 ± 1538.06 | 64,103.13 ± 3242.12 | NS |
| C16:1 | 21,977.17 ± 844.14 | 25,118.60 ± 333.46 | Up ** |
| C17:0 | 622.01 ± 26.90 | 636.70 ± 2.65 | NS |
| C17:1 | 836.88 ± 6.36 | 835.63 ± 17.52 | NS |
| C18:0 | 20,838.49 ± 9.99 | 26,268.43 ± 227.55 | Up *** |
| C18:1n9t | 0 | 0 | ---- |
| C18:1n9c | 118,554.11 ± 1652.67 | 117,502.15 ± 3730.93 | NS |
| C18:2n6t | 0 | 0 | ---- |
| C18:2n6c | 60,435.67 ± 402.57 | 58,232.17 ± 2572.42 | NS |
| C18:3n6 | 0 | 0 | ---- |
| C18:3n3 | 4034.85 ± 94.16 | 4145.20 ± 39.24 | NS |
| C20:0 | 576.07 ± 29.08 | 646.11 ± 6.99 | Up * |
| C20:1 | 3601.34 ± 35.07 | 3583.60 ± 19.50 | NS |
| C20:2 | 1420.30 ± 12.40 | 1459.13 ± 4.35 | Down ** |
| C21:0 | 0 | 0 | ---- |
| C20:3n6 | 536.93 ± 22.31 | 556.34 ± 1.11 | NS |
| C20:4n6 | 685.24 ± 10.66 | 821.83 ± 46.59 | Up ** |
| C20:3n3 | 200.46 ± 5.48 | 201.28 ± 1.71 | NS |
| C20:5n3 | 182.35 ± 13.88 | 154.11 ± 1.73 | Down * |
| C22:0 | 95.49 ± 3.51 | 103.27 ± 1.26 | Up * |
| C22:1n9 | 170.41 ± 0.57 | 171.81 ± 3.16 | Up NS |
| C22:2n6 | 8.83 ± 0.33 | 10.20 ± 0.19 | Up ** |
| C23:0 | 15.42 ± 1.45 | 19.39 ± 1.35 | Up * |
| C24:0 | 34.29 ± 8.86 | 64.93 ± 4.91 | Up ** |
| C24:1 | 107.11 ± 4.00 | 106.56 ± 0.61 | NS |
| C22:6 | 905.71 ± 80.71 | 876.80 ± 49.26 | NS |
| total | 311,475.65 ± 4127.86 | 314,100.24 ± 2094.52 | NS |
| SFA | 97,497.15 ± 1739.71 | 99,980.78 ± 3565.31 | NS |
| MUFA | 145,568.17 ± 2210.44 | 147,662.40 ± 4110.32 | NS |
| PUFA | 68,410.34 ± 519.11 | 66,457.06 ± 2638.75 | NS |
| C16:0/C16:1 | 3.10 ± 0.10 | 2.55 ± 0.16 | Down ** |
| C16:0/C18:0 | 3.26 ± 0.073 | 2.44 ± 0.10 | Down *** |
| C18:0/C18:1 | 0.18 ± 0.0024 | 0.22 ± 0.0090 | Up *** |
| C16:1/C18:1 | 0.19 ± 0.0063 | 0.21 ± 0.0040 | Up ** |
| SFA/total | 0.31 ± 0.0016 | 0.32 ± 0.0092 | NS |
| MUFA/total | 0.47 ± 0.0023 | 0.47 ± 0.016 | NS |
| PUFA/total | 0.22 ± 0.0024 | 0.21 ± 0.0070 | NS |
| Gene | Primer Sequences (5′→3′) | TM (°C) |
|---|---|---|
| β-actin | F: GTGACGTTGACATCCGTAAAGA | 60 |
| R: GCCGGACTCATCGTACTCC | ||
| U6 | F: CTCGCTTCGGCAGCACA | 60 |
| R: AACGCTTCACGAATTTGCGT | ||
| C/EBPα | F: GCGGGAACGCAACAACATC | 62 |
| R: GTCACTGGTCAACTCCAGCAC | ||
| CDK4 | F: GTCAGTTTCTAAGCGGCCTG | 60 |
| R: CACGGGTGTTGCGTATGTAG | ||
| FABP4 | F: AAGGTGAAGAGCATCATAACCCT | 52 |
| R: TCACGCCTTTCATAACACATTCC | ||
| SCD-1 | F: TTCTTGCGATACACTCTGGTGC | 63.3 |
| R: CGGGATTGAATGTTCTTGTCGT | ||
| ACADL | F: TGCCCTATATTGCGAATTACGG | 63.3 |
| R: CTATGGCACCGATACACTTGC | ||
| ACOX2 | F: CATCCAACGTGACCCAGTGTT | 63.3 |
| R: AAATGCGTTCAGGACCGTCTT | ||
| CDK2 | F: GCGACCTCCTCCCAATATCG | 60 |
| R: GTCTGATCTCTTTCCCCAACTCT | ||
| LPL | F: GGTTGCGCGTAGAGAGGATG | 59.6 |
| R: CTCACGCTCTGACATGCCTTC | ||
| PPARα | F: TACTGCCGTTTTCACAAGTGC | 62 |
| R: AGGTCGTGTTCACAGGTAAGA | ||
| Adipor2 | F: TGTTCCTCTTAATCCTGCCCA | 52.8 |
| R: CCAACCTGCACAAGTTCCCTT | ||
| FAS | F: AGGTGGACTGGATACACAGAC | 62 |
| R: TCTCCTGCCCAAACTCTTTGC | ||
| Cyclin E1 | F: CTCCGACCTTTCAGTCCGC | 60 |
| R: CACAGTCTTGTCAATCTTGGCA | ||
| p21 | F: GATGGCTTCGACACCATTCC | 60 |
| R: AGACGACACAGGTGAGGAAG | ||
| Cyclin D1 | F: CTCCGTATCTTACTTCAAGTGCG | 60 |
| R: CTTCTCGGCAGTCAAGGGAA | ||
| p53 | F: CTCTCCCCCGCAAAAGAAAAA | 60 |
| R: CGGAACATCTCGAAGCGTTTA | ||
| mmu-miR-126b-5p | ATTATTACTCACGGTACGAGTT | 60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, L.; He, J.; Zhao, Y.; Niu, L.; Chen, L.; Tang, G.; Jiang, Y.; Hao, X.; Bai, L.; Li, X.; et al. MicroRNA-126b-5p Exacerbates Development of Adipose Tissue and Diet-Induced Obesity. Int. J. Mol. Sci. 2021, 22, 10261. https://doi.org/10.3390/ijms221910261
Shen L, He J, Zhao Y, Niu L, Chen L, Tang G, Jiang Y, Hao X, Bai L, Li X, et al. MicroRNA-126b-5p Exacerbates Development of Adipose Tissue and Diet-Induced Obesity. International Journal of Molecular Sciences. 2021; 22(19):10261. https://doi.org/10.3390/ijms221910261
Chicago/Turabian StyleShen, Linyuan, Jin He, Ye Zhao, Lili Niu, Lei Chen, Guoqing Tang, Yanzhi Jiang, Xiaoxia Hao, Lin Bai, Xuewei Li, and et al. 2021. "MicroRNA-126b-5p Exacerbates Development of Adipose Tissue and Diet-Induced Obesity" International Journal of Molecular Sciences 22, no. 19: 10261. https://doi.org/10.3390/ijms221910261
APA StyleShen, L., He, J., Zhao, Y., Niu, L., Chen, L., Tang, G., Jiang, Y., Hao, X., Bai, L., Li, X., Zhang, S., & Zhu, L. (2021). MicroRNA-126b-5p Exacerbates Development of Adipose Tissue and Diet-Induced Obesity. International Journal of Molecular Sciences, 22(19), 10261. https://doi.org/10.3390/ijms221910261