
Fine Particulate Matter-Induced Oxidative Stress Mediated by UVA-Visible Light Leads to Keratinocyte Damage

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
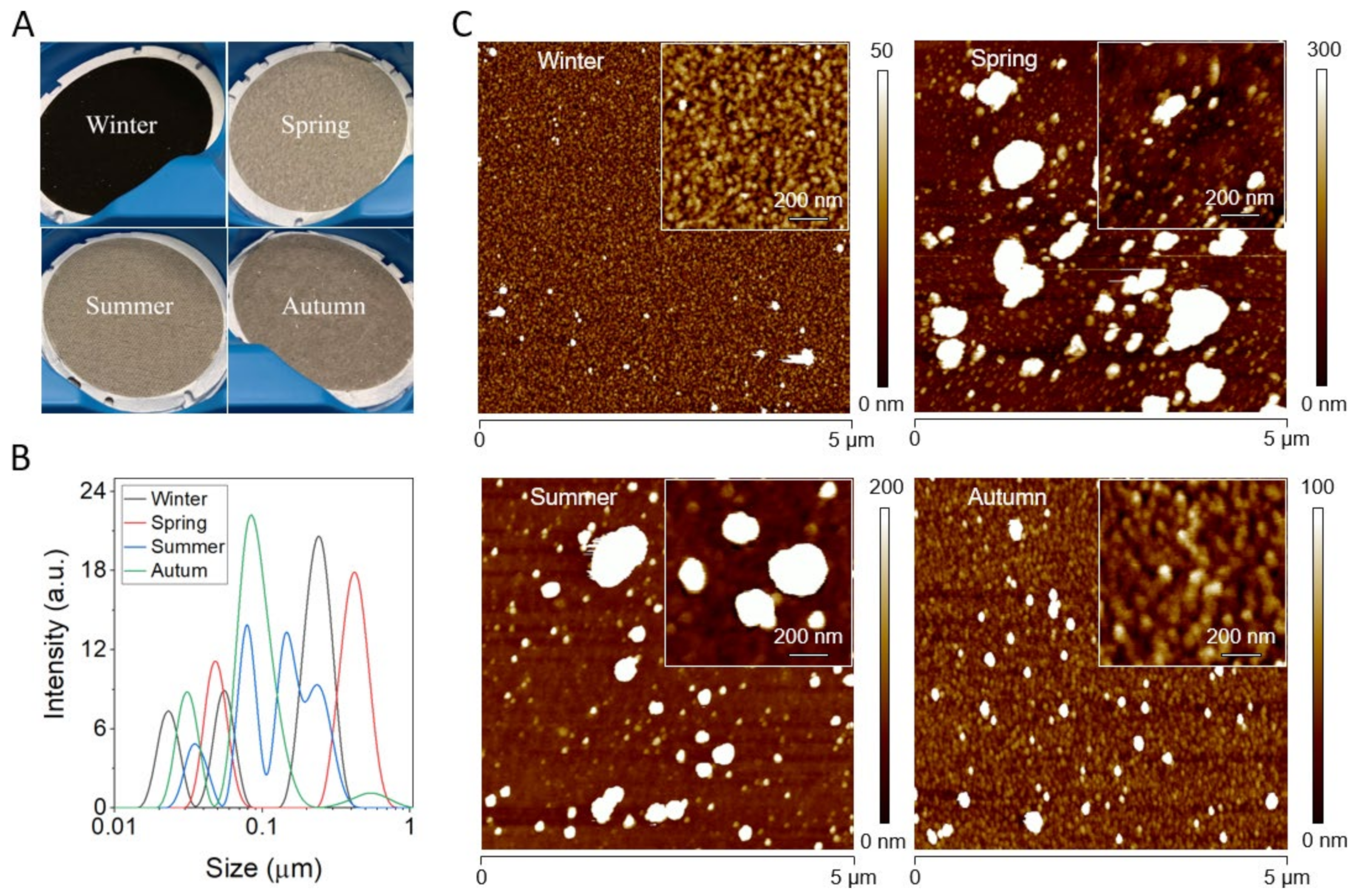
2.1. Size Analysis of PM Particles
2.2. Phototoxic Effect of Particulate Matter
2.3. Photogeneration of Free Radicals by PM
2.4. Photogeneration of Singlet Oxygen (1O2) by PM
2.5. Light-Induced Lipid Peroxidation by PM
2.6. The Relationship between Photoactivated PM and Apoptosis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Particulate Matter Extraction
4.3. Dynamic Light Scattering
4.4. Atomic Force Microscopy
4.5. Cell Treatment and Light Irradiation
4.6. Propidium Iodide Staining
4.7. MTT Assay
4.8. Detection of Free Radicals by EPR Spin Trapping
4.9. Time-Resolved Detection of Singlet Oxygen Phosphorescence
4.10. Liposome Preparation and Iodometric Assay for Lipid Hydroperoxide Measurements
4.11. Flow Cytometry
4.12. Caspase 3/7 Fluorometric Analysis
4.13. Real-Time PCR
4.14. Mitochondrial Membrane Potential Detection
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sotiropoulou, P.A.; Blanpain, C. Development and Homeostasis of the Skin Epidermis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008383. [Google Scholar] [CrossRef] [Green Version]
- Drakaki, E.; Dessinioti, C.; Antoniou, C.V. Air Pollution and the Skin. Front. Environ. Sci. 2014, 2, 1–6. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Ambient Air Pollution: A Global Assessment of Exposure and Burden of Disease. Available online: http://www.who.int/phe/publications/air-pollution-global-assessment/en/ (accessed on 22 April 2021).
- Aust, A.E.; Ball, J.C.; Hu, A.A.; Lighty, J.A.S.; Smith, K.R.; Straccia, A.M.; Veranth, J.M.; Young, W.C. Particle Characteristics Responsible for Effects on Human Lung Epithelial Cells. Res. Rep. Health Eff. Inst. 2002, 110, 1–76. [Google Scholar]
- Vejerano, E.P.; Rao, G.; Khachatryan, L.; Cormier, S.A.; Lomnicki, S. Environmentally Persistent Free Radicals: Insights on a New Class of Pollutants. Environ. Sci. Technol. 2018, 52, 2468–2481. [Google Scholar] [CrossRef] [PubMed]
- Szaciłowski, K.; Macyk, W.; Drzewiecka-Matuszek, A.; Brindell, M.; Stochel, G. Bioinorganic Photochemistry: Frontiers and Mechanisms. Chem. Rev. 2005, 105, 2647–2694. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Imai, N.; Mashima, R.; Konaka, R.; Inoue, M.; Dunlap, W.C. Singlet Oxygen from Irradiated Titanium Dioxide and Zinc Oxide. Methods Enzymol. 2000, 319, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Kozinska, A.; Zadlo, A.; Labuz, P.; Broniec, A.; Pabisz, P.; Sarna, T. The Ability of Functionalized Fullerenes and Surface-Modified TiO2 Nanoparticles to Photosensitize Peroxidation of Lipids in Selected Model Systems. Photochem. Photobiol. 2019, 95, 227–236. [Google Scholar] [CrossRef]
- Bonnett, R.; Berenbaum, M. Porphyrins as Photosensitizers. In Photosensitizing Compounds: Their Chemistry, Biology and Clinical Use; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 1989; pp. 40–53. [Google Scholar]
- Ibuki, Y.; Goto, R. Phototoxicity of Benzo[a]Pyrene by Ultraviolet A Irradiation: Induction of Apoptosis in Jurkat Cells. Environ. Toxicol. Pharmacol. 2001, 11, 101–109. [Google Scholar] [CrossRef]
- Mauthe, R.J.; Cook, V.M.; Coffing, S.L.; Baird, W.M. Exposure of Mammalian Cell Cultures to Benzo[a] and Light Results in Oxidative Dna Damage as Measured by 8-Hydroxydeoxyguanosine Formation. Carcinogenesis 1995, 16, 133–137. [Google Scholar] [CrossRef]
- Teranishi, M.; Toyooka, T.; Ohura, T.; Masuda, S.; Ibuki, Y. Benzo [a] Pyrene Exposed to Solar-Simulated Light Inhibits Apoptosis and Augments Carcinogenicity. Chem. Interact. 2010, 185, 4–11. [Google Scholar] [CrossRef]
- Von Koschembahr, A.; Youssef, A.; Béal, D.; Gudimard, L.; Giot, J.P.; Douki, T. Toxicity and DNA Repair in Normal Human Keratinocytes Co-Exposed to Benzo[a]Pyrene and Sunlight. Toxicol. Vitr. 2020, 63, 104744. [Google Scholar] [CrossRef]
- Ash, C.; Dubec, M.; Donne, K.; Bashford, T. Effect of Wavelength and Beam Width on Penetration in Light-Tissue Interaction Using Computational Methods. Lasers Med Sci. 2017, 32, 1909–1918. [Google Scholar] [CrossRef]
- Jin, S.P.; Li, Z.; Choi, E.K.; Lee, S.; Kim, Y.K.; Seo, E.Y.; Chung, J.H.; Cho, S. Urban Particulate Matter in Air Pollution Penetrates into the Barrier-Disrupted Skin and Produces ROS-Dependent Cutaneous Inflammatory Response in Vivo. J. Dermatol. Sci. 2018, 91, 175–183. [Google Scholar] [CrossRef]
- Liao, Z.; Nie, J.; Sun, P. The Impact of Particulate Matter (PM2.5) on Skin Barrier Revealed by Transcriptome Analysis: Focusing on Cholesterol Metabolism. Toxicol. Rep. 2020, 7, 1–9. [Google Scholar] [CrossRef]
- Magnani, N.D.; Muresan, X.M.; Belmonte, G.; Cervellati, F.; Sticozzi, C.; Pecorelli, A.; Miracco, C.; Marchini, T.; Evelson, P.; Valacchi, G. Skin Damage Mechanisms Related to Airborne Particulate Matter Exposure. Toxicol. Sci. 2016, 149, 227–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.-W.; Lin, Z.-C.; Hu, S.C.-S.; Chiang, Y.-C.; Hsu, L.-F.; Lin, Y.-C.; Lee, I.-T.; Tsai, M.-H.; Fang, J.-Y. Urban particulate matter down-regulates filaggrin via COX2 expression/PGE2 production leading to skin barrier dysfunction. Sci. Rep. 2016, 6, 27995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lao, J.-Y.; Xie, S.-Y.; Wu, C.-C.; Bao, L.-J.; Tao, S.; Zeng, E.Y. Importance of Dermal Absorption of Polycyclic Aromatic Hydrocarbons Derived from Barbecue Fumes. Environ. Sci. Technol. 2018, 52, 8330–8338. [Google Scholar] [CrossRef] [PubMed]
- Weschler, C.J.; Bekö, G.; Koch, H.M.; Salthammer, T.; Schripp, T.; Toftum, J.; Clausen, G. Transdermal Uptake of Diethyl Phthalate and Di(n-Butyl) Phthalate Directly from Air: Experimental Verification. Environ. Health Perspect. 2015, 123, 928–934. [Google Scholar] [CrossRef] [Green Version]
- Kelly, F.J.; Fussell, J.C. Size, source and chemical composition as determinants of toxicity attributable to ambient particulate matter. Atmos. Environ. 2012, 60, 504–526. [Google Scholar] [CrossRef]
- Reiss, R.; Anderson, E.L.; Cross, C.E.; Hidy, G.; Hoel, D.; McClellan, R.; Moolgavkar, S. Evidence of Health Impacts of Sulfate-and Nitrate-Containing Particles in Ambient Air. Inhal. Toxicol. 2007, 19, 419–449. [Google Scholar] [CrossRef]
- Costa, M.; Mollenhauer, H.H. Phagocytosis of Nickel Subsulfide Particles during the Early Stages of Neoplastic Transformation in Tissue Culture. Cancer Res. 1980, 40, 2688–2694. [Google Scholar] [PubMed]
- Cervellati, F.; Benedusi, M.; Manarini, F.; Woodby, B.; Russo, M.; Valacchi, G.; Pietrogrande, M.C. Proinflammatory Properties and Oxidative Effects of Atmospheric Particle Components in Human Keratinocytes. Chemosphere 2020, 240, 124746. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, J.; Bae, J.S. ROS Homeostasis and Metabolism: A Critical Liaison for Cancer Therapy. Exp. Mol. Med. 2016, 48, e269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romani, A.; Cervellati, C.; Muresan, X.M.; Belmonte, G.; Pecorelli, A.; Cervellati, F.; Benedusi, M.; Evelson, P.; Valacchi, G. Keratinocytes Oxidative Damage Mechanisms Related to Airbone Particle Matter Exposure. Mech. Ageing Dev. 2018, 172, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Bickers, D.R.; Athar, M. Oxidative Stress in the Pathogenesis of Skin Disease. J. Investig. Dermatol. 2006, 126, 2565–2575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bito, T.; Nishigori, C. Impact of reactive oxygen species on keratinocyte signaling pathways. J. Dermatol. Sci. 2012, 68, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.A.; Sinha, A.A. Oxidative stress and autoimmune skin disease. Eur. J. Dermatol. 2013, 23, 5–13. [Google Scholar] [CrossRef]
- Vierkötter, A.; Schikowski, T.; Ranft, U.; Sugiri, D.; Matsui, M.; Krämer, U.; Krutmann, J. Airborne Particle Exposure and Extrinsic Skin Aging. J. Investig. Dermatol. 2010, 130, 2719–2726. [Google Scholar] [CrossRef] [Green Version]
- Buettner, G. Spin Trapping: ESR parameters of spin adducts 1474 1528V. Free. Radic. Biol. Med. 1987, 3, 259–303. [Google Scholar] [CrossRef]
- Mokrzynski, K.; Ito, S.; Wakamatsu, K.; Camenish, T.G.; Sarna, T.; Sarna, M. Photoreactivity of Hair Melanin from Different Skin Phototypes—Contribution of Melanin Subunits to the Pigments Photoreactive Properties. Int. J. Mol. Sci. 2021, 22, 4465. [Google Scholar] [CrossRef]
- Furso, J.; Zadlo, A.; Szewczyk, G.; Sarna, T.J. Photoreactivity of Bis-Retinoid A2E Complexed with a Model Protein in Selected Model Systems. Cell Biophys. 2020, 78, 415–427. [Google Scholar] [CrossRef]
- St. Denis, T.G.; Vecchio, D.; Zadlo, A.; Rineh, A.; Sadasivam, M.; Avci, P.; Huang, L.; Kozinska, A.; Chandran, R.; Sarna, T.; et al. Thiocyanate Potentiates Antimicrobial Photodynamic Therapy: In Situ Generation of the Sulfur Trioxide Radical Anion by Singlet Oxygen. Free. Radic. Biol. Med. 2013, 65, 800–810. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.S.; Olejnik, D. Photolysis of Polycyclic Aromatic Hydrocarbons in Water. Water Res. 2001, 35, 233–243. [Google Scholar] [CrossRef]
- Gilmore, T.D. Introduction to NF-ΚB: Players, Pathways, Perspectives. Oncogene 2006, 25, 6680–6684. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.O.; Thundiyil, J.G.; Stolbach, A. Clearing the Air: A Review of the Effects of Particulate Matter Air Pollution on Human Health. J. Med. Toxicol. 2012, 8, 166–175. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-H.; Kabir, E.; Kabir, S. A Review on the Human Health Impact of Airborne Particulate Matter. Environ. Int. 2015, 74, 136–143. [Google Scholar] [CrossRef]
- Chevion, M. A site-specific mechanism for free radical induced biological damage: The essential role of redox-active transition metals. Free. Radic. Biol. Med. 1988, 5, 27–37. [Google Scholar] [CrossRef]
- Stohs, S.J.; Bagchi, D. Oxidative Mechanisms in the Toxicity of Metal Ions. Free. Radic. Biol. Med. 1995, 18, 321–336. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.J.; Hayes, J.D.; Higgins, L.G.; Wolf, C.R.; Dinkova-Kostova, A.T. Activation of the NRF2 Signaling Pathway by Copper-Mediated Redox Cycling of Para- and Ortho-Hydroquinones. Chem. Biol. 2010, 17, 75–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milnerowicz, H.; Ściskalska, M.; Dul, M. Pro-inflammatory effects of metals in persons and animals exposed to tobacco smoke. J. Trace Elements Med. Biol. 2015, 29, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Jomova, K.; Rhodes, C.J.; Kuča, K.; Musílek, K. Redox- and non-redox-metal-induced formation of free radicals and their role in human disease. Arch. Toxicol. 2016, 90, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Styszko, K.; Samek, L.; Szramowiat, K.; Korzeniewska, A.; Kubisty, K.; Rakoczy-Lelek, R.; Kistler, M.; Giebl, A.K. Oxidative Potential of PM10 and PM2.5 Collected at High Air Pollution Site Related to Chemical Composition: Krakow Case Study. Air Qual. Atmos. Health 2017, 10, 1123–1137. [Google Scholar] [CrossRef]
- Krutmann, J.; Liu, W.; Li, L.; Pan, X.; Crawford, M.; Sore, G.; Seite, S. Pollution and skin: From epidemiological and mechanistic studies to clinical implications. J. Dermatol. Sci. 2014, 76, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, E.; Schäfer, C.; Calles, C.; Bernsmann, T.; Bernshausen, T.; Wurm, M.; Hübenthal, U.; Cline, J.E.; Hajimiragha, H.; Schroeder, P.; et al. Lightening up the UV Response by Identification of the Arylhydrocarbon Receptor as a Cytoplasmatic Target for Ultraviolet B Radiation. Proc. Natl. Acad. Sci. USA 2007, 104, 8851–8856. [Google Scholar] [CrossRef] [Green Version]
- Vogel, C.F.A.; William Chang, W.L.; Kado, S.; McCulloh, K.; Vogel, H.; Wu, D.; Haarmann-Stemmann, T.; Yang, G.X.; Leung, P.S.C.; Matsumura, F.; et al. Transgenic Overexpression of Aryl Hydrocarbon Receptor Repressor (AhRR) and AhR-Mediated Induction of CYP1A1, Cytokines, and Acute Toxicity. Environ. Health Perspect. 2016, 124, 1071–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valacchi, G.; Virgili, F.; Cervellati, C.; Pecorelli, A. OxInflammation: From Subclinical Condition to Pathological Biomarker. Front. Physiol. 2018, 9, 858. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, F.; Woodby, B.; Pecorelli, A.; Schiavone, M.L.; Pambianchi, E.; Messano, N.; Therrien, J.P.; Choudhary, H.; Valacchi, G. Additive Effect of Combined Pollutants to UV Induced Skin OxInflammation Damage. Evaluating the Protective Topical Application of a Cosmeceutical Mixture Formulation. Redox Biol. 2020, 34, 101481. [Google Scholar] [CrossRef]
- Samek, L.; Furman, L.; Mikrut, M.; Regiel-Futyra, A.; Macyk, W.; Stochel, G.; van Eldik, R. Chemical Composition of Submicron and Fine Particulate Matter Collected in Krakow, Poland. Consequences for the APARIC Project. Chemosphere 2017, 187, 430–439. [Google Scholar] [CrossRef]
- Samek, L.; Stegowski, Z.; Styszko, K.; Furman, L.; Fiedor, J. Seasonal Contribution of Assessed Sources to Submicron and Fine Particulate Matter in a Central European Urban Area. Environ. Pollut. 2018, 241, 406–411. [Google Scholar] [CrossRef]
- Samek, L.; Stegowski, Z.; Furman, L.; Styszko, K.; Szramowiat, K.; Fiedor, J. Quantitative Assessment of PM2.5 Sources and Their Seasonal Variation in Krakow. Water Air Soil Pollut. 2017, 228, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tong, H.; Zhang, Y.; Filippi, A.; Wang, T.; Li, C.; Liu, F.; Leppla, D.; Kourtchev, I.; Wang, K.; Keskinen, H.M.; et al. Radical Formation by Fine Particulate Matter Associated with Highly Oxygenated Molecules. Environ. Sci. Technol. 2019, 53, 12506–12518. [Google Scholar] [CrossRef]
- Mikrut, M.; Regiel-Futyra, A.; Samek, L.; Macyk, W.; Stochel, G.; van Eldik, R. Generation of Hydroxyl Radicals and Singlet Oxygen by Particulate Matter and Its Inorganic Components. Environ. Pollut. 2018, 238, 638–646. [Google Scholar] [CrossRef]
- Gilbert, B.C.; Norman, R.O.C.; Sealy, R.C. Electron Spin Resonance Studies. Part XLIII. Reaction of Dimethyl Sulphoxide with the Hydroxyl Radical. J. Chem. Soc. Perkin Trans. 2 1975, 303, 303–308. [Google Scholar] [CrossRef]
- Veltwisch, D.; Janata, E.; Asmus, K.-D. Primary Processes in the Reaction of OH·-Radicals with Sulphoxides. J. Chem. Soc. Perkin Trans. 2 1980, 146–153. [Google Scholar] [CrossRef]
- El-Nouby Adam, M. Effect of The Atmosphere on UVB Radiation Reaching the Earth’s Surface: Dependence on Solar Zenith Angle. Atmos. Ocean. Sci. Lett. 2011, 4, 139–145. [Google Scholar] [CrossRef]
- Lim, H.W.; Honingsmann, H.; Hawk, J.L.M. Photodermatology; CRC Press: Boca Raton, FL, USA, 2007; ISBN 978036745319. [Google Scholar]
- Kozinska, A.; Oles, T.; Sarna, T. Photoactivation and Detection of Photoexcited Molecules and Photochemical Products. Isr. J. Chem. 2012, 52, 745–756. [Google Scholar] [CrossRef]
- Krasnovsky, A.A. Primary Mechanisms of Photoactivation of Molecular Oxygen. History of Development and the Modern Status of Research. Biochemistry 2007, 72, 1065–1080. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Kang, Z.; Jiang, S.; Zhao, J.; Yan, S.; Xu, F.; Xu, J. Effects of Ambient Fine Particles PM 2.5 on Human HaCaT Cells. Int. J. Environ. Res. Public Health 2017, 14, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.J.; Kim, J.H.; Jeong, G.J.; Park, K.Y.; Lee, M.K.; Seo, S.J. Particulate Matter Induces Pro-Inflammatory Cytokines via Phosphorylation of P38 MAPK Possibly Leading to Dermal Inflammaging. Exp. Dermatol. 2019, 28, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Kwon, K.; Park, S.-H.; Han, B.S.; Oh, S.W.; Lee, S.E.; Yoo, J.A.; Park, S.J.; Kim, J.; Kim, J.W.; Cho, J.Y.; et al. Negative Cellular Effects of Urban Particulate Matter on Human Keratinocytes Are Mediated by P38 MAPK and NF-κB-dependent Expression of TRPV 1. Int. J. Mol. Sci. 2018, 19, 2660. [Google Scholar] [CrossRef] [Green Version]
- Winikoff, S.E.; Zeh, H.J.; DeMarco, R.; Lotze, M.T. Cytolytic Assays. In Measuring Immunity. Basic Biology and Clinical Assessment; Lotze, M.T., Thomson, A.W., Eds.; Academic Press: London, UK, 2005; pp. 343–349. ISBN 978-0-12-455900-4. [Google Scholar]
- Hu, R.; Xie, X.-Y.; Xu, S.-K.; Wang, Y.-N.; Jiang, M.; Wen, L.-R.; Lai, W.; Guan, L. PM2.5 Exposure Elicits Oxidative Stress Responses and Mitochondrial Apoptosis Pathway Activation in HaCaT Keratinocytes. Chin. Med. J. 2017, 130, 2205–2214. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, L.; Tuo, J.; Liu, Q.; Zhang, X.; Xu, Z.; Liu, S.; Sui, G. Analysis of PM2.5-Induced Cytotoxicity in Human HaCaT Cells Based on a Microfluidic System. Toxicol. Vitr. 2017, 43, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Desagher, S.; Martinou, J.-C. Mitochondria as the central control point of apoptosis. Trends Cell Biol. 2000, 10, 369–377. [Google Scholar] [CrossRef]
- Ryu, Y.S.; Kang, K.A.; Piao, M.J.; Ahn, M.J.; Yi, J.M.; Hyun, Y.-M.; Kim, S.H.; Ko, M.K.; Park, C.O.; Hyun, J.W. Particulate matter induces inflammatory cytokine production via activation of NFκB by TLR5-NOX4-ROS signaling in human skin keratinocyte and mouse skin. Redox Biol. 2019, 21, 101080. [Google Scholar] [CrossRef]
- Dong, L.; Hu, R.; Yang, D.; Zhao, J.; Kan, H.; Tan, J.; Guan, M.; Kang, Z.; Xu, F. Fine Particulate Matter (PM2.5) Upregulates Expression of Inflammasome NLRP1 via ROS/NF-ΚB Signaling in Hacat Cells. Int. J. Med. Sci. 2020, 17, 2200–2206. [Google Scholar] [CrossRef]
- Piao, M.J.; Ahn, M.J.; Kang, K.A.; Ryu, Y.S.; Hyun, Y.J.; Shilnikova, K.; Zhen, A.X.; Jeong, J.W.; Choi, Y.H.; Kang, H.K.; et al. Particulate Matter 2.5 Damages Skin Cells by Inducing Oxidative Stress, Subcellular Organelle Dysfunction, and Apoptosis. Arch. Toxicol. 2018, 92, 2077–2091. [Google Scholar] [CrossRef] [Green Version]
- Fernando, I.P.S.; Jayawardena, T.U.; Kim, H.-S.; Vaas, A.P.J.P.; De Silva, H.I.C.; Nanayakkara, C.M.; Abeytunga, D.T.U.; Lee, W.; Ahn, G.; Lee, D.-S.; et al. A keratinocyte and integrated fibroblast culture model for studying particulate matter-induced skin lesions and therapeutic intervention of fucosterol. Life Sci. 2019, 233, 116714. [Google Scholar] [CrossRef] [PubMed]
- Piao, M.J.; Kang, K.A.; Zhen, A.X.; Fernando, P.D.S.M.; Ahn, M.J.; Koh, Y.S.; Kang, H.K.; Yi, J.M.; Choi, Y.H.; Hyun, J.W. Particulate Matter 2.5 Mediates Cutaneous Cellular Injury by Inducing Mitochondria-Associated Endoplasmic Reticulum Stress: Protective Effects of Ginsenoside Rb1. Antioxidants 2019, 8, 383. [Google Scholar] [CrossRef] [Green Version]
- Sreedhar, A.; Aguilera-Aguirre, L.; Singh, K.K. Mitochondria in Skin Health, Aging, and Disease. Cell Death Dis. 2020, 11, 444. [Google Scholar] [CrossRef]
- Muro, E.; Ekin Atilla-Gokcumen, G.; Eggert, U.S. Lipids in Cell Biology: How Can We Understand Them Better? Mol. Biol. Cell 2014, 25, 1819–1823. [Google Scholar] [CrossRef] [PubMed]
- Pappas, A. Epidermal Surface Lipids. Derm.-Endocrinol. 2009, 1, 72–76. [Google Scholar] [CrossRef] [Green Version]
- Zhen, A.X.; Piao, M.J.; Kang, K.A.; Fernando, P.D.S.M.; Kang, H.K.; Koh, Y.S.; Yi, J.M.; Hyun, J.W. Niacinamide Protects Skin Cells from Oxidative Stress Induced by Particulate Matter. Biomol. Ther. 2019, 27, 562–569. [Google Scholar] [CrossRef]
- Bein, K.J.; Wexler, A.S. A High-Efficiency, Low-Bias Method for Extracting Particulate Matter from Filter and Impactor Substrates. Atmos. Environ. 2014, 90, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Wytrwal, M.; Leduc, C.; Sarna, M.; Goncalves, C.; Kepczynski, M.; Midoux, P.; Nowakowska, M.; Pichon, C. Gene Delivery Efficiency and Intracellular Trafficking of Novel Poly(Allylamine) Derivatives. Int. J. Pharm. 2015, 478, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Zadlo, A.; Mokrzyński, K.; Ito, S.; Wakamatsu, K.; Sarna, T. The Influence of Iron on Selected Properties of Synthetic Pheomelanin. Cell Biochem. Biophys. 2020, 78, 181–189. [Google Scholar] [CrossRef]
- Sarna, M.; Olchawa, M.; Zadlo, A.; Wnuk, D.; Sarna, T. The Nanomechanical Role of Melanin Granules in the Retinal Pigment Epithelium. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 801–807. [Google Scholar] [CrossRef]
- Krzysztynska-Kuleta, O.I.; Olchawa, M.M.; Sarna, T.J. Melanopsin Signaling Pathway in HEK293 Cell Line with Stable Expression of Human Melanopsin: Possible Participation of Phospholipase C Beta 4 and Diacylglycerol. Photochem. Photobiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Olchawa, M.M.; Krzysztynska-Kuleta, O.I.; Mokrzynski, K.T.; Sarna, P.M.; Sarna, T.J. Quercetin Protects ARPE-19 Cells against Photic Stress Mediated by the Products of Rhodopsin Photobleaching. Photochem. Photobiol. Sci. 2020, 19, 1022–1034. [Google Scholar] [CrossRef]
- Zareba, M.; Szewczyk, G.; Sarna, T.; Hong, L.; Simon, J.D.; Henry, M.M.; Burke, J.M. Effects of Photodegradation on the Physical and Antioxidant Properties of Melanosomes Isolated from Retinal Pigment Epithelium. Photochem. Photobiol. 2006, 82, 1024–1029. [Google Scholar] [CrossRef]
- Stoll, S.; Schweiger, A. EasySpin, a Comprehensive Software Package for Spectral Simulation and Analysis in EPR. J. Magn. Reson. 2006, 178, 42–55. [Google Scholar] [CrossRef]
- Szewczyk, G.; Zadlo, A.; Sarna, M.; Ito, S.; Wakamatsu, K.; Sarna, T. Aerobic Photoreactivity of Synthetic Eumelanins and Pheomelanins: Generation of Singlet Oxygen and Superoxide Anion. Pigment Cell Melanoma Res. 2016, 29, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Zawrotniak, M.; Wojtalik, K.; Rapala-Kozik, M. Farnesol, a Quorum-Sensing Molecule of Candida Albicans Triggers the Release of Neutrophil Extracellular Traps. Cells 2019, 8, 1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mokrzyński, K.; Krzysztyńska-Kuleta, O.; Zawrotniak, M.; Sarna, M.; Sarna, T. Fine Particulate Matter-Induced Oxidative Stress Mediated by UVA-Visible Light Leads to Keratinocyte Damage. Int. J. Mol. Sci. 2021, 22, 10645. https://doi.org/10.3390/ijms221910645
Mokrzyński K, Krzysztyńska-Kuleta O, Zawrotniak M, Sarna M, Sarna T. Fine Particulate Matter-Induced Oxidative Stress Mediated by UVA-Visible Light Leads to Keratinocyte Damage. International Journal of Molecular Sciences. 2021; 22(19):10645. https://doi.org/10.3390/ijms221910645
Chicago/Turabian StyleMokrzyński, Krystian, Olga Krzysztyńska-Kuleta, Marcin Zawrotniak, Michał Sarna, and Tadeusz Sarna. 2021. "Fine Particulate Matter-Induced Oxidative Stress Mediated by UVA-Visible Light Leads to Keratinocyte Damage" International Journal of Molecular Sciences 22, no. 19: 10645. https://doi.org/10.3390/ijms221910645
APA StyleMokrzyński, K., Krzysztyńska-Kuleta, O., Zawrotniak, M., Sarna, M., & Sarna, T. (2021). Fine Particulate Matter-Induced Oxidative Stress Mediated by UVA-Visible Light Leads to Keratinocyte Damage. International Journal of Molecular Sciences, 22(19), 10645. https://doi.org/10.3390/ijms221910645