
In Silico Analysis of Fatty Acid Desaturases Structures in Camelina sativa, and Functional Evaluation of Csafad7 and Csafad8 on Seed Oil Formation and Seed Morphology



Abstract
:1. Introduction
2. Results
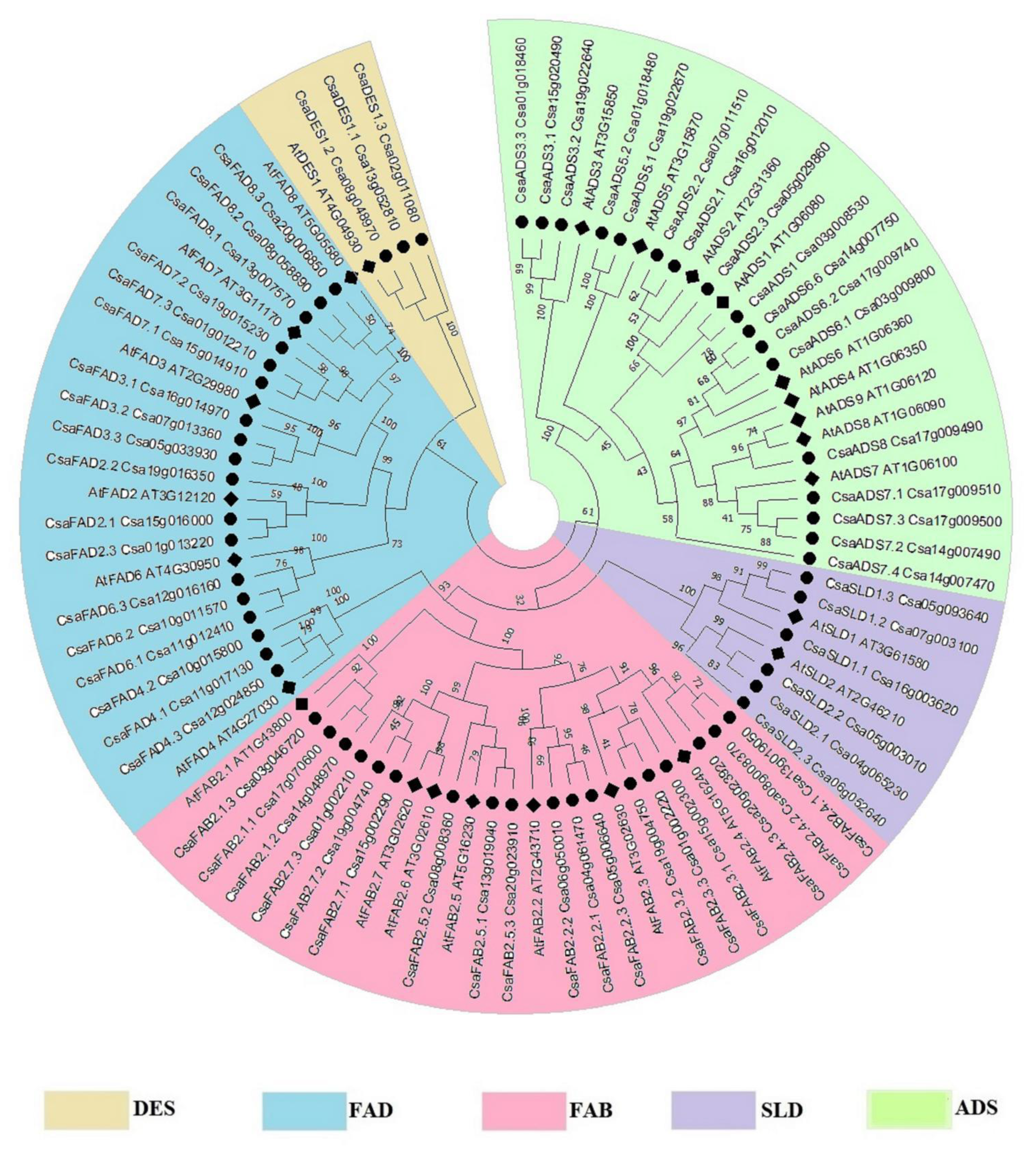
2.1. Synteny and Evolutionary Relationship of Fatty Acid Desaturase Genes in C. Sativa and A. thaliana
2.2. Diversity in Gene and Protein Structure in C. sativa Fatty Acid Desaturases
2.3. Expression Profile Analysis of C. sativa Desaturases Revealed an Increase in Expression in Young Leaves, Seeds, and Flowers
2.4. C. sativa Fatty Acid Desaturases Were Detected in Three Intracellular Compartments
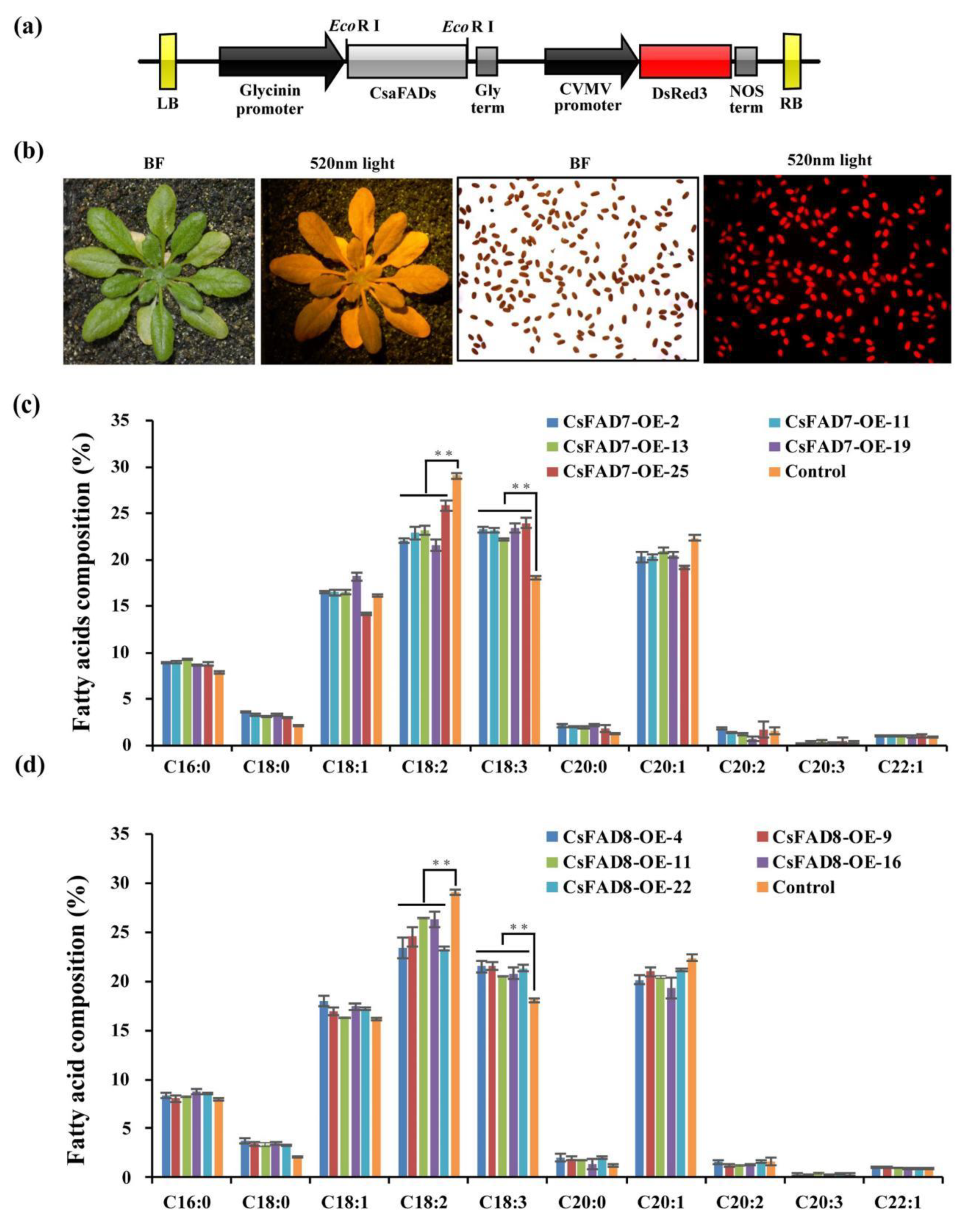
2.5. Overexpression of CsaFAD7 Caused Seed Size and Oil Content Reduction in A. thaliana Seed but Not CsaFAD8
3. Discussion
3.1. Evolution of A. thaliana and C. sativa and their Respective Fatty Acid Desaturases
3.2. Conservation and Diversification in Structure of the Fatty Acid Desaturases
3.3. C. sativa Desaturases Accumulation Was Associated with Different Cellular Compartmentations and Tissues
3.4. Seed-Specific Expression of the cDNA Encoding CsaFAD7 and CsaFAD8 Affected the Fatty Acid Composition of Seed Oil and the Seed Size in A. thaliana
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Identification of Desaturase Genes in C. sativa
4.3. Phylogenetic Analysis of Desaturase Genes
4.4. Gene Structure Analysis
4.5. Protein Domain Structure and Conserved Motif Analysis
4.6. Expression Pattern of Desaturase Genes
4.7. Subcellular Localization Analysis
4.8. Construction of CsaFAD7 and Csafad8 Seed-Specific Expression Vectors and Expression in A. thaliana
4.9. Transgenic Seed Size and Lipid Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harwood, J.L. Plant acyl lipids: Structure, distribution, and analysis. In Biochemistry of Plants; Stumpf, P.K., Conn, E.E., Eds.; Academic Press: New York, NY, USA, 1980; Volume 4, pp. 1–55. [Google Scholar]
- Stumpf, P.K. Biosynthesis of saturated and unsaturated fatty acids. In Biochemistry of Plants; Stumpf, P.K., Conn, E.E., Eds.; Academic Press: New York, NY, USA, 1980; Volume 4, pp. 177–204. [Google Scholar]
- Los, D.A.; Murata, N. Structure and expression of fatty acid desaturases. Biochim. Biophys. Acta 1998, 1394, 3–15. [Google Scholar] [CrossRef]
- Nakamura, M.T.; Nara, T.Y. Structure, function, and dietary regulation of delta6, delta5, and delta9 desaturases. Annu. Rev. Nutr. 2004, 24, 345–376. [Google Scholar] [CrossRef]
- Ohlrogge, J.; Browse, J. Lipid biosynthesis. Plant Cell 1995, 7, 957–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, X.Y.; Yang, Q.L.; Lu, Y.D.; Wang, J.Y.; Zhang, Q.F.; Pan, L.J.; Yu, S. Genome-wide analysis of fatty acid desaturases in soybean (Glycine max). Plant Mol. Biol. Rep. 2011, 29, 769–783. [Google Scholar] [CrossRef] [Green Version]
- Murphy, D.J. Production of novel oils in plants. Curr. Opin. Biotechnol. 1999, 10, 175–180. [Google Scholar] [CrossRef]
- Kargiotidou, A.; Deli, D.; Galanopoulou, D.; Tsaftaris, A.; Farmaki, T. Low temperature and light regulate delta 12 fatty acid desaturases (FAD2) at a transcriptional level in cotton (Gossypium hirsutum). J. Exp. Bot. 2008, 59, 2043–2056. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.A.; Dauk, M.; Ramadan, H.; Yang, H.; Seamons, L.E.; Haslam, R.P.; Forseille, L. Involvement of Arabidopsis ACYL-COENZYME A DESATURASE-LIKE2 (At2g31360) in the biosynthesis of the very-long-chain monounsaturated fatty acid components of membrane lipids. Plant Physiol. 2013, 161, 81–96. [Google Scholar] [CrossRef] [Green Version]
- Kachroo, A.; Shanklin, J.; Whittle, E.; Lapchyk, L.; Hildebrand, D.; Kachroo, P. The Arabidopsis stearoyl-acyl carrier protein-desaturase family and the contribution of leaf isoforms to oleic acid synthesis. Plant Mol. Biol. 2007, 63, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Okuley, J.; Lightner, J.; Feldmann, K.; Yadav, N.; Lark, E. Arabidopsis FAD2 gene encodes the enzyme that is essential for polyunsaturated lipid synthesis. Plant Cell 1994, 6, 147–158. [Google Scholar] [CrossRef]
- Falcone, D.L.; Gibson, S.; Lemieux, B.; Somerville, C. Identification of a gene that complements an Arabidopsis mutant deficient in chloroplast ω6 desaturase activity. Plant Physiol. 1994, 106, 1453–1459. [Google Scholar] [CrossRef] [Green Version]
- Arondel, V.; Lemieux, I.; Hwang, S.; Gibson, H.M. Goodman and C.R. Somerville, Map-based cloning of a gene controlling omega-3 fatty acid desaturation in Arabidopsis. Science 1992, 258, 1353–1355. [Google Scholar] [CrossRef] [PubMed]
- Iba, K.; Gibson, S.; Nishiuchi, T.; Fuse, T.; Nishimura, M.; Arondel, V.; Somerville, C. A gene encoding a chloroplast omega-3 fatty acid desaturase complements alterations in fatty acid desaturation and chloroplast copy number of the fad7 mutant of Arabidopsis thaliana. J. Biol. Chem. 1993, 268, 24099–24105. [Google Scholar] [CrossRef]
- Gibson, S.; Arondel, V.; Iba, K.; Somerville, C. Cloning of a temperature-regulated gene encoding a chloroplast ω-3 desaturase from Arabidopsis thaliana. Plant Physiol. 1994, 106, 1615–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Ajjawi, I.; Manoli, A.; Sawin, A.; Xu, C.; Froehlich, J.E.; Benning, C. Fatty acid desaturase4 of Arabidopsis encodes a protein distinct from characterized fatty acid desaturases. Plant J. 2009, 60, 832–839. [Google Scholar] [CrossRef]
- Ryan, P.R.; Liu, Q.; Sperling, P.; Dong, B.; Franke, S.; Delhaize, E. A higher plant Δ8 sphingolipid desaturase with a preference for (Z)-isomer formation confers aluminum tolerance to yeast and plants. Plant Physiol. 2007, 144, 1968–1977. [Google Scholar] [CrossRef] [Green Version]
- Shanklin, J.; Cahoon, E. Desaturation and related modifications of fatty acids. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 611–641. [Google Scholar] [CrossRef] [Green Version]
- López Alonso, D.; García-Maroto, F.; Rodríguez-Ruiz, J.; Garrido, J.; Vilches, M. Evolution of the membrane-bound fatty acid desaturases. Biochem. Syst. Ecol. 2003, 31, 1111–1124. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Ohlrogge, J. Acyl-lipid metabolism. Arabidopsis Book 2013, 11, e161. [Google Scholar] [CrossRef] [Green Version]
- Gehringer, A.; Friedt, W.; Luhs, W.; Snowdon, R.J. Genetic mapping of agronomic traits in false flax (Camelina sativa subsp. sativa). Genome 2006, 49, 1555–1563. [Google Scholar] [CrossRef]
- Hutcheon, C.; Ditt, R.F.; Beilstein, M.; Comai, L.; Schroeder, J.; Goldstein, E.; Kiser, J. Polyploid genome of Camelina sativa revealed by isolation of fatty acid synthesis genes. BMC Plant Biol. 2010, 10, 233. [Google Scholar] [CrossRef] [Green Version]
- Kagale, S.; Koh, C.; Nixon, J.; Bollina, V.; Clarke, W.E.; Tuteja, R.; Parkin, I.A. The emerging biofuel crop Camelina sativa retains a highly undifferentiated hexaploid genome structure. Nat. Commun. 2014, 5, 3706. [Google Scholar] [CrossRef] [Green Version]
- Mandáková, T.; Pouch, M.; Brock, J.R.; Al-Shehbaz, I.A.; Lysak, M.A. Origin and evolution of diploid and allopolyploid Camelina genomes were accompanied by chromosome shattering. Plant Cell 2019, 31, 2596–2612. [Google Scholar] [CrossRef]
- Lysak, M.A.; Berr, A.; Pecinka, A.; Schmidt, R.; McBreen, K.; Schubert, I. Mechanisms of chromosome number reduction in Arabidopsis thaliana and related Brassicaceae species. Proc. Natl. Acad. Sci. USA 2006, 103, 5224–5229. [Google Scholar] [CrossRef] [Green Version]
- Schranz, M.E.; Lysak, M.A.; Mitchell-Olds, T. The ABC’s of comparative genomics in the Brassicaceae: Building blocks of crucifer genomes. Trends Plant Sci. 2006, 11, 535–542. [Google Scholar] [CrossRef]
- Lysak, M.A.; Mandáková, T.; Schranz, M.E. Comparative paleogenomics of crucifers: Ancestral genomic blocks revisited. Curr. Opin. Plant Biol. 2016, 30, 108–115. [Google Scholar] [CrossRef]
- Moser, B.R. Biodiesel from alternative oilseed feedstocks: Camelina and field pennycress. Biofuels 2012, 3, 193–209. [Google Scholar] [CrossRef] [Green Version]
- Shonnard, D.R.; Williams, L.; Kalnes, T.M. Camelina derived jet fuel and diesel: Sustainable advanced biofuels. Environ. Prog. Sustain. Energy 2010, 29, 382–392. [Google Scholar] [CrossRef]
- Iskandarov, U.; Kim, H.J.; Cahoon, E.B. Camelina: An emerging oilseed platform for advanced biofuels and bio-based materials. In Plants and Bioenergy; McCann, M.C., Buckeridge, M.S., Carpita, N.C., Eds.; Springer: Berlin, Germany, 2014; pp. 131–140. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Park, J.H.; Kim, D.J.; Kim, A.Y.; Su, M.C. Functional analysis of diacylglycerol acyltransferase1 genes from Camelina sativa and effects of CsDGAT1B overexpression on seed mass and storage oil content in C. sativa. Plant Biotechnol. Rep. 2016, 10, 141–153. [Google Scholar] [CrossRef]
- Chhikara, S.; Abdullah, H.M.; Akbari, P.; Schnell, D.; Dhankher, O.P. Engineering Camelina sativa (L.) Crantz for enhanced oil and seed yields by combining diacylglycerol acyltransferase1 and glycerol-3-phosphate dehydrogenase expression. Plant Biotechnol. J. 2017, 16, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kim, R.J.; Kim, H.U.; Suh, M.C. Development of Camelina enhanced with drought stress resistance and seed oil production by co-overexpression of MYB96A and DGAT1C. Ind. Crops Prod. 2019, 138, 111475. [Google Scholar] [CrossRef]
- Choudhury, S.R.; Riesselman, A.J.; Sona Pandey, S. Constitutive or seed-specific overexpression of Arabidopsis G-protein c subunit 3 (AGG3) results in increased seed and oil production and improved stress tolerance in Camelina sativa. Plant Biotechnol. J. 2014, 12, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Park, W.; Feng, Y.; Kim, H.; Suh, M.C.; Ahn, S.J. Changes in fatty acid content and composition between wild type and CsHMA3 overexpressing Camelina sativa under heavy-metal stress. Plant Cell Rep. 2015, 34, 1489–1498. [Google Scholar] [CrossRef]
- An, D.; Suh, M.C. Overexpression of Arabidopsis WRI1 enhanced seed mass and storage oil content in Camelina sativa. Plant Biotechnol. Rep. 2015, 9, 137–148. [Google Scholar] [CrossRef]
- Li, M.; Wei, F.; Tawfall, A.; Tang, M.; Saettele, A.; Wang, X. Overexpression of patatin-related phospholipase AIIIδ altered plant growth and increased seed oil content in Camelina. Plant Biotechnol. J. 2015, 13, 766–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, G.; Wang, G.; Kim, S.C.; Li, J.; Zhou, Y.; Wang, X. Increased expression of fatty acid and ABC transporters enhances seed oil production in Camelina. Biotechnol. Biofuels 2021, 14, 49. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Xie, L.; Chen, G.Q.; Lee, M.Y.; Loque, D.; Scheller, H.V. A transgene design for enhancing oil content in Arabidopsis and Camelina seeds. Biotechnol. Biofuels 2018, 11, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, M.F.; Sánchez-García, A.; Salas, J.J.; Garcés, R.; Martínez-Force, E. Characterization of soluble acyl-ACP desaturases from Camelina sativa, Macadamia tetraphylla and Dolichandra unguis-cati. J. Plant Physiol. 2015, 178, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.; Snapp, A.R.; Lu, C. Identification of three genes encoding microsomal oleate desaturases (FAD2) from the oilseed crop Camelina sativa. Plant Physiol. Biochem. 2011, 49, 223–229. [Google Scholar] [CrossRef]
- Kim, H.; Go, Y.S.; Kim, A.Y.; Lee, S.; Kim, K.N.; Lee, G.J.; Suh, M.C. Isolation and functional analysis of three microsomal delta-12 fatty acid desaturase genes from Camelina sativa (L.) cv. CAME. J. Plant Biotechnol. 2014, 41, 146–158. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.Z.; Henry, I.M.; Lynagh, P.G.; Comai, L.; Cahoon, E.B.; Weeks, D.P. Significant enhancement of fatty acid composition in seeds of the allohexaploid, Camelina sativa, using CRISPR/Cas9 gene editing. Plant Biotechnol. J. 2017, 15, 648–657. [Google Scholar] [CrossRef] [Green Version]
- Morineau, C.; Bellec, Y.; Tellier, F.; Gissot, L.; Kelemen, Z.; Nogué, F.; Faure, J.D. Selective gene dosage by CRISPR-Cas9 genome editing in hexaploid Camelina sativa. Plant Biotechnol. J. 2017, 15, 729–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Rodríguez, M.F.; Salas, J.J.; Venegas-Calerón, M.; Garcés, R.; Martínez-Force, E. Molecular cloning and characterization of the genes encoding a microsomal oleate Δ12 desaturase (CsFAD2) and linoleate Δ15 desaturase (CsFAD3) from Camelina sativa. Ind. Crops Prod. 2016, 89, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Guo, Z.; Chen, K.; Tian, T.; Tan, J.; Chen, X.; Li, M. Ultra-high α-linolenic acid accumulating developmental defective embryo was rescued by lysophosphatidic acid acyltransferase 2. Plant J. 2020, 103, 2151–2167. [Google Scholar] [CrossRef] [PubMed]
- Ahmadizadeh, M.; Rezaee, S.; Heidari, P. Genome-wide characterization and expression analysis of fatty acid desaturase gene family in Camelina sativa. Gene Rep. 2020, 21, 100894. [Google Scholar] [CrossRef]
- Gomez-Cano, F.; Carey, L.; Lucas, K.; García Navarrete, T.; Mukundi, E.; Lundback, S.; Grotewold, E. CamRegBase: A gene regulation database for the biofuel crop, Camelina sativa. Database 2020. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Choudhuri, S. Chapter 1-Fundamentals of Genes and Genomes. Bioinformatics for Beginners; Academic Press: Cambridge, MA, USA, 2014; pp. 1–25. [Google Scholar] [CrossRef]
- Itoh, M.; Nacher, J.C.; Kuma, K.; Goto, S.; Kanehisa, M. Evolutionary history and functional implications of protein domains and their combinations in eukaryotes. Genome Biol. 2007, 8, R121. [Google Scholar] [CrossRef] [Green Version]
- The Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar] [CrossRef] [Green Version]
- Berti, M.; Gesch, R.; Eynck, C.; Anderson, J.; Cermak, S. Camelina uses, genetics, genomics, production, and management. Ind. Crops Prod. 2016, 94, 690–710. [Google Scholar] [CrossRef]
- Kumar, A.; Bennetzen, J.L. Plant retrotransposons. Annu. Rev. Genet. 1999, 33, 479–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennetzen, J.L. Mechanisms and rates of genome expansion and contraction in flowering plants. Genetica 2002, 115, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Lagercrantz, U. Comparative mapping between Arabidopsis thaliana and Brassica nigra indicates that Brassica genomes have evolved through extensive genome replication accompanied by chromosome fusions and frequent rearrangements. Genetics 1998, 150, 1217–1228. [Google Scholar] [CrossRef]
- Lysak, M.A.; Koch, M.A.; Pecinka, A.; Schubert, I. Chromosome triplication found across the tribe Brassiceae. Genome Res. 2005, 15, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Parkin, I.A.; Gulden, S.M.; Sharpe, A.G.; Lukens, L.; Trick, M.; Osborn, T.C.; Lydiate, D.J. Segmental structure of the Brassica napus genome based on comparative analysis with Arabidopsis thaliana. Genetics 2005, 71, 765–781. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Wu, J.; Wang, X. Genome triplication drove the diversification of Brassica plants. Hortic. Res. 2014, 1, 14024. [Google Scholar] [CrossRef] [Green Version]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [Green Version]
- Celik Altunoglu, Y.; Unel, N.M.; Baloglu, M.C.; Ulu, F.; Can, T.H.; Cetinkaya, R. Comparative identification and evolutionary relationship of fatty acid desaturase (FAD) genes in some oil crops: The sunflower model for evaluation of gene expression pattern under drought stress. Biotechnol. Biotechnol. Equip. 2018, 32, 846–857. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Li, W.; He, Q.; Daud, M.K.; Chen, J.; Zhu, S. Characterization of 19 genes encoding membrane-bound fatty acid desaturases and their expression profiles in Gossypium raimondii under low temperature. PLoS ONE 2015, 10, e0123281. [Google Scholar] [CrossRef] [Green Version]
- Hajiahmadi, Z.; Abedi, A.; Wei, H.; Sun, W.; Ruan, H.; Zhuge, Q.; Movahedi, A. Identification, evolution, expression, and docking studies of fatty acid desaturase genes in wheat (Triticum aestivum L.). BMC genomics 2020, 21, 778. [Google Scholar] [CrossRef]
- Zhang, J.Z. Evolution by gene duplication: An update. Trends Ecol. Evol. 2003, 18, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, S.; Liberles, D.A. Subfunctionalization of duplicated genes as a transition state to neofunctionalization. BMC Evol. Biol. 2005, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Freeling, M.; Scanlon, M.J.; Fowler, J.E. Fractionation and subfunctionalization following genome duplications: Mechanisms that drive gene content and their consequences. Curr. Opin. Genet. Dev. 2015, 5, 110–118. [Google Scholar] [CrossRef]
- Hu, T.T.; Pattyn, P.; Bakker, E.G.; Cao, J.; Cheng, J.F.; Clark, R.M.; Guo, Y.L. The Arabidopsis lyrata genome sequence and the basis of rapid genome size change. Nat. Genet. 2011, 43, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Shi, F. Evolution of the RALF gene family in plants: Gene duplication and selection patterns. Evol. Bioinf. 2012, 8, S9652. [Google Scholar] [CrossRef]
- Chen, C.; Yang, J.; Tong, H.; Li, T.; Wang, L.; Chen, H. Genome-wide analysis of fatty acid desaturase genes in rice (Oryza sativa L.). Sci. Rep. 2019, 9, 19445. [Google Scholar] [CrossRef] [Green Version]
- Díaz, M.L.; Cuppari, S.; Soresi, D.; Carrera, A. In silico analysis of fatty acid desaturase genes and proteins in grasses. Appl. Biochem. Biotechnol. 2018, 184, 484–499. [Google Scholar] [CrossRef]
- Sperling, P.; Zahringer, U.; Heinz, E. A sphingolipid desaturase from higher plants. Identification of a new cytochrome b5 fusion protein. J. Biol. Chem. 1998, 273, 28590–28596. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Xian, Z.; Kang, X.; Tang, N.; Li, Z. Genome-wide identification, phylogeny and expression analysis of GRAS gene family in tomato. BMC Plant Biol. 2015, 15, 209. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, H.; Sun, J.; Li, B.; Zhu, Q.; Chen, S.; Zhang, H. Arabidopsis fatty acid desaturase fad2 is required for salt tolerance during seed germination and early seedling growth. PLoS ONE 2012, 7, e30355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Barg, R.; Yin, M.; Gueta-Dahan, Y.; Leikin-Frenkel, A.; Salts, Y.; Ben-Hayyim, G. Modulated fatty acid desaturation via overexpression of two distinct ω-3 desaturases differentially alters tolerance to various abiotic stresses in transgenic tobacco cells and plants. Plant J. 2005, 44, 361–371. [Google Scholar] [CrossRef]
- Lan, T.; Gao, J.; Zeng, Q.Y. Genome-wide analysis of the LEA (late embryogenesis abundant) protein gene family in Populus trichocarpa. Tree Genet. Genom. 2013, 9, 253–264. [Google Scholar] [CrossRef]
- Xie, D.W.; Wang, X.N.; Fu, L.S.; Sun, J.; Zheng, W.; Li, Z.F. Identification of the trehalose-6-phosphate synthase gene family in winter wheat and expression analysis under conditions of freezing stress. J Genet. 2015, 94, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Janin, J.; Wodak, S.J. Structural domains in proteins and their role in the dynamics of protein function. Prog. Biophys. Mol. Biol. 1983, 42, 21–78. [Google Scholar] [CrossRef]
- Han, J.H.; Batey, S.; Nickson, A.A.; Teichmann, S.A.; Clarke, J. The folding and evolution of multidomain proteins. Nat. Rev. Mol. Cell Biol. 2007, 8, 319–330. [Google Scholar] [CrossRef]
- Lupas, A.N.; Ponting, C.P.; Russell, R.B. On the evolution of protein folds: Are similar motifs in different protein folds the result of convergence, insertion, or relics of an ancient peptide world? J. Struct. Biol. 2001, 134, 191–203. [Google Scholar] [CrossRef]
- Ponting, C.P.; Russell, R.R. The natural history of protein domains. Annu. Rev. Biophys. Biomol. Struct. 2002, 31, 45–71. [Google Scholar] [CrossRef]
- Vogel, C.; Bashton, M.; Kerrison, N.D.; Chothia, C.; Teichmann, S.A. Structure, function and evolution of multidomain proteins. Curr. Opin. Struct. Biol. 2004, 14, 208–216. [Google Scholar] [CrossRef]
- Yang, S.; Bourne, P.E. The evolutionary history of protein domains viewed by species phylogeny. PLoS ONE 2009, 4, e8378. [Google Scholar] [CrossRef]
- Wang, M.; Caetano-Anollés, G. The evolutionary mechanics of domain organization in proteomes and the rise of modularity in the protein world. Structure 2009, 17, 66–78. [Google Scholar] [CrossRef] [Green Version]
- Illergård, K.; Ardell, D.H.; Elofsson, A. Structure is three to ten times more conserved than sequence—a study of structural response in protein cores. Proteins Struct. Funct. Bioinforma. 2009, 77, 499–508. [Google Scholar] [CrossRef]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [Green Version]
- Vogel, C.; Teichmann, S.A.; Pereira-Leal, J. The relationship between domain duplication and recombination. J. Mol. Biol. 2005, 346, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Grishin, N.V. Fold change in evolution of protein structures. J. Struct. Biol. 2001, 134, 167–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bateman, A.; Coggill, P.; Finn, R.D. DUFs: Families in search of function. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 1148–1152. [Google Scholar] [CrossRef] [Green Version]
- Lodish, H.; Berk, A.; Zipursky, S. Insertion of Membrane Proteins into the ER Membrane, 4th ed.; Freeman, W.H., Ed.; W.H. Freeman & Company: New York, NY, USA, 2000. [Google Scholar]
- Dailey, H.A.; Strittmatter, P. Characterization of the interaction of amphipathic cytochrome b5 with stearoyl coenzyme A desaturase and NADPH:cytochrome P-450 reductase. J. Biol. Chem. 1980, 255, 5184–5189. [Google Scholar] [CrossRef]
- Mitchell, A.G.; Martin, C.E. A novel cytochrome b5-like domain is linked to the carboxyl terminus of the Saccharomyces cerevisiae v-9 fatty acid desaturase. J. Biol. Chem. 1995, 270, 29766–29772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ternes, P.; Franke, S.; Zähringer, U.; Sperling, P.; Heinz, E. Identification and characterization of a sphingolipid delta 4-desaturase family. J. Biol. Chem. 2002, 277, 25512–25518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheeff, E.D.; Bourne, P.E. Structural evolution of the protein kinase-like superfamily. PLoS Comput. Biol. 2005, 1, e49. [Google Scholar] [CrossRef]
- Gough, J. Convergent evolution of domain architectures (is rare). Bioinformatics 2005, 21, 1464–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kummerfeld, S.K.; Teichmann, S.A. Relative rates of gene fusion and fission in multi-domain proteins. Trends Genet. 2005, 21, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Björklund, A.K.; Ekman, D.; Elofsson, A. Expansion of protein domain repeats. PLoS Comput. Biol. 2006, 2, e114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, C.; Morea, V. Duplication, divergence and formation of novel protein topologies. Bioessays 2006, 28, 973–978. [Google Scholar] [CrossRef]
- Somerville, C.; Browse, J. Dissecting desaturation: Plants prove advantageous. Trends Cell Biol. 1996, 6, 148–153. [Google Scholar] [CrossRef]
- Xue, Y.; Chen, B.; Win, A.N.; Fu, C.; Lian, J.; Liu, X.; Wang, R.; Zhang, X.; Chai, Y. Omega-3 fatty acid desaturase gene family from two ω-3 sources, Salvia hispanica and Perilla frutescens: Cloning, characterization and expression. PLoS ONE 2018, 13, e0191432. [Google Scholar] [CrossRef]
- Engelman, D.M.; Steitz, T.A.; Goldman, A. Identifying nonpolar transbilayer helices in amino acid sequences of membrane proteins. Annu. Rev. Biophys. Biophys. Chem. 1986, 15, 321–353. [Google Scholar] [CrossRef]
- Sharpe, H.J.; Stevens, T.J.; Munro, S. A comprehensive comparison of transmembrane domains reveals organelle-specific properties. Cell 2010, 142, 158–169. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, K.; Yoshizawa, A.C.; Okuda, S.; Kuma, K.; Goto, S.; Kanehisa, M. The repertoire of desaturases and elongases reveals fatty acid variations in 56 eukaryotic genomes. J. Lipid Res. 2008, 49, 183–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Markham, J.E.; Cahoon, E.B. Sphingolipid Δ8 unsaturation is important for glucosylceramide biosynthesis and low-temperature performance in Arabidopsis. Plant J. 2012, 69, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Soria-Garcïa, Ï.N.; Rubio, M.A.C.; Lagunas, B.; Lï Pez-Gomollï, N.S.; Lujï, N.M.A.L.Ï.N.; Dï Az-Guerra, R.L. Tissue Distribution and Specific Contribution of Arabidopsis FAD7 and FAD8 plastid desaturases to the JA- and ABA-mediated cold stress or defense responses. Plant Cell Physiol. 2019, 60, 1025–1040. [Google Scholar] [CrossRef] [Green Version]
- Damude, H.G.; Zhang, H.; Farrall, L.; Ripp, K.G.; Tomb, J.F.; Hollerbach, D.; Yadav, N.S. Identification of bifunctional Δ12/ω-3 fatty acid desaturases for improving the ratio of ω-3 to ω-6 fatty acid in microbes and plants. Proc. Nat. Acad. Sci. USA 2006, 103, 9446–9451. [Google Scholar] [CrossRef] [Green Version]
- Eckert, H.; LaVallee, B.; Schweiger, B.J.; Kinney, A.J.; Cahoon, E.B.; Clemente, T. Co-expression of the borage Δ6 desaturase and the Arabidopsis Δ15 desaturase results in high accumulation of stearidonic acid in the seeds of transgenic soybean. Planta 2006, 224, 1050–1057. [Google Scholar] [CrossRef]
- Liu, H.L.; Yin, Z.J.; Xiao, L.; Xu, Y.N.; Qu, L.Q. Identification and evaluation of ω-3 fatty acid desaturase genes for hyperfortifying α-linolenic acid in transgenic rice seed. J. Exp. Bot. 2012, 63, 3279–3287. [Google Scholar] [CrossRef] [Green Version]
- D’Angeli, S.; Matteucci, M.; Fattorini, L.; Gismondi, A.; Ludovici, M.; Canini, A.; Altamura, M.M. OeFAD8, OeLIP and OeOSM expression and activity in cold-acclimation of Olea europaea, a perennial dicot without winter-dormancy. Planta 2016, 243, 1279–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, M.L.; Sicardo, M.D.; Martinez-Rivas, J.M. Differential contribution of endoplasmic reticulum and chloroplast ω-3 fatty acid desaturase genes to the linolenic acid content of olive (Olea europaea) fruit. Plant Cell Physiol. 2016, 57, 138–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, M.L.; Lima-Cabello, E.; Alché, J.D.; Martínez-Rivas, J.M.; Castro, A.J. Lipid Composition and Associated Gene Expression Patterns during Pollen Germination and Pollen Tube Growth in Olive (Olea europaea L.). Plant Cell Physiol. 2020, 6, 1348–1364. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.Y.; Ruan, J.; Tian, H.Y.; Shan, L.; Meng, J.J.; Guo, F.; Li, X. The family of peanut fatty acid desaturase genes and a functional analysis of four ω-3 AhFAD3 members. Plant Mol. Biol. Rep. 2020, 38, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Dmitriev, A.A.; Kezimana, P.; Rozhmina, T.A.; Zhuchenko, A.A.; Povkhova, L.V.; Pushkova, E.N.; Melnikova, N.V. Genetic diversity of SAD and FAD genes responsible for the fatty acid composition in flax cultivars and lines. BMC Plant Biol. 2020, 20, 301. [Google Scholar] [CrossRef]
- Liu, K.; Zhao, S.; Wang, S.; Wang, H.; Zhang, Z. Identification and analysis of the FAD gene family in walnuts (Juglans regia L.) based on transcriptome data. BMC Genom. 2020, 21, 299. [Google Scholar] [CrossRef] [Green Version]
- Torres-Franklin, M.L.; Repellin, A.; Huynh, V.B.; d’Arcy-Lameta, A.; Zuily-Fodil, Y.; Pham-Thi, A.T. Omega-3 fatty acid desaturase (FAD3, FAD7, FAD8) gene expression and linolenic acid content in cowpea leaves submitted to drought and after rehydration. Environ. Exp. Bot. 2009, 65, 162–169. [Google Scholar] [CrossRef]
- Zhang, Z.; Jin, X.; Liu, Z.; Zhang, J.; Liu., W. Genome-wide identification of FAD gene family and functional analysis of MsFAD3.1 involved in the accumulation of α-linolenic acid in alfalfa. Crop Sci. 2020, 1–14. [Google Scholar] [CrossRef]
- Li, J.; Galla, A.L.; Avila, C.A.; Flattmann, K.; Vaughn, K.L.; Goggin, F. Fatty acid desaturases in the chloroplast and endoplasmic reticulum promote susceptibility to the green peach aphid, myzus persicae, in Arabidopsis thaliana. Mol. Plant Microbe Interact 2021. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, N.D.; Bates, P.D.; Browse, J. Wrinkled1 rescues feedback inhibition of fatty acid synthesis in hydroxylase-expressing seeds. Plant Physiol. 2016, 171, 179–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, P.D.; Johnson, S.R.; Cao, X.; Li, J.; Nam, J.W.; Jaworski, J.G.; Ohlrogge, J.B.; Browse, J. Fatty acid synthesis is inhibited by inefficient utilization of unusual fatty acids for glycerolipid assembly. Proc. Natl. Acad. Sci. USA 2014, 111, 1204–1209. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, S.; Sinha, S.; Das, N.; Maiti, M.K. Increasing the stearate content in seed oil of Brassica juncea by heterologous expression of MlFatB affects lipid content and germination frequency of transgenic seeds. Plant Physiol. Biochem. 2015, 96, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Cahoon, E.B.; Dietrich, C.R.; Meyer, K.; Damude, H.G.; Dyer, J.M.; Kinney, A.J. Conjugated fatty acids accumulate to high levels in phospholipids of metabolically engineered soybean and Arabidopsis seeds. Phytochemistry 2006, 67, 1166–1176. [Google Scholar] [CrossRef]
- Cahoon, E.B.; Carlson, T.J.; Ripp, K.G.; Schweiger, B.J.; Cook, G.A.; Hall, S.E.; Kinney, A.J. Biosynthetic origin of conjugated double bonds: Production of fatty acid components of high-value drying oils in transgenic soybean embryos. Proc. Natl. Acad. Sci. USA 1999, 96, 12935–12940. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Yu, K.; Wu, Y.; Tateno, M.; Hatanaka, T.; Hildebrand, D.F. Vernonia DGATs can complement the disrupted oil and protein metabolism in epoxygenase-expressing soybean seeds. Metab. Eng. 2012, 14, 29–38. [Google Scholar] [CrossRef]
- Eccleston, V.S.; Ohlrogge, J.B. Expression of lauroyl-acyl carrier protein thioesterase in Brassica napus seeds induces pathways for both fatty acid oxidation and biosynthesis and implies a set point for triacylglycerol accumulation. Plant Cell 1998, 10, 613–622. [Google Scholar] [CrossRef] [Green Version]
- Yeom, W.W.; Kim, H.J.; Lee, K.; Cho, H.S.; Kim, J.; Jung, H.W.; Oh, S.; Jun, S.E.; Kim, H.U.; Chung, Y. Increased Production of α-Linolenic Acid in Soybean Seeds by Overexpression of Lesquerella FAD3-1. Front. Plant Sci. 2020, 10. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucl. Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Zuckerkandl, E.; Pauling, L. Evolutionary divergence and convergence in proteins. In Evolving Genes and Proteins; Bryson, V., Vogel, H.J., Eds.; Academic Press: New York, NY, USA, 1965; pp. 97–166. [Google Scholar]
- Higgins, D.G.; Sharp, P.M. CLUSTAL: A package for performing multiple sequence alignment on a microcomputer. Gene 1988, 73, 237–244. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchler-Bauer, A.; Bryant, S.H. CD-search: Protein domain annotations on the fly. Nucl. Acids Res. 2004, 32, W327–W331. [Google Scholar] [CrossRef]
- Krogh, A.; von Heijne, B.L.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirokawa, T.; Boon-Chieng, S.; Mitaku, S. SOSUI: Classification and secondary structure prediction system for membrane proteins. Bioinformatics 1998, 14, 378–379. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins Struct. Funct. Bioinf. 2006, 64, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.; Cho, Y.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Prot. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [Green Version]
- Nelson, B.K.; Cai, X.; Nebenführ, A. A multicolored set of in vivo organelle markers for colocalization studies in Arabidopsis and other plants. Plant J. 2007, 51, 1126–1136. [Google Scholar] [CrossRef]
- Shao, G.; Wei, X.; Chen, M.; Tang, S.; Luo, J.; Jiao, G.; Hu, P. Allelic variation for a candidate gene for GS7, responsible for grain shape in rice. Theor. Appl. Genet. 2012, 125, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.Q.; Shao, G.N.; Wei, X.J.; Chen, M.L.; Sheng, Z.H.; Luo, J.; Hu, P.S. QTL mapping of grain weight in rice and the validation of the QTL qTGW3.2. Gene 2013, 527, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Li, H.; Li, S.; Xu, L.; Zhao, Z.; Wang, J. Genetic dissection on rice grain shape by the two-dimensional image analysis in one japonica × indica population consisting of recombinant inbred lines. Theor. Appl. Genet. 2015, 128, 1969–1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | A. thaliana (n = 5) | C. sativa (n = 20) | ||||||
|---|---|---|---|---|---|---|---|---|
| Gene Name | Gene ID | Chr | Size (AA) | Gene ID | Chr | Size (AA) | Identity (%) | |
| Soluble | FAB2.1 | AT1G43800 | 1 | 391 | Csa17g070600 | 17 | 391 | 92 |
| Csa14g048970 | 14 | 391 | 92 | |||||
| Csa03g046720 | 3 | 394 | 87 | |||||
| FAB2.2 | AT2G43710 | 2 | 401 | Csa04g061470 | 4 | 401 | 98 | |
| Csa06g050010 | 6 | 401 | 98 | |||||
| Csa05g006640 | 5 | 466 | 97 | |||||
| FAB2.3 | AT3G02630 | 3 | 396 | Csa15g002300 | 15 | 396 | 96 | |
| Csa19g004750 | 19 | 396 | 97 | |||||
| Csa01g002220 | 1 | 544 | 96 | |||||
| FAB2.4 | AT5G16240 | 5 | 394 | Csa13g019050 | 13 | 333 | 94 | |
| Csa08g008370 | 8 | 406 | 88 | |||||
| Csa20g023920 | 20 | 427 | 89 | |||||
| FAB2.5 | AT5G16230 | 5 | 401 | Csa13g019040 | 13 | 402 | 95 | |
| Csa08g008360 | 8 | 425 | 95 | |||||
| Csa20g023910 | 20 | 403 | 96 | |||||
| FAB2.6 | AT3G02610 | 3 | 411 | * | ||||
| FAB2.7 | AT3G02620 | 3 | 403 | Csa15g002290 | 15 | 412 | 90 | |
| Csa19g004740 | 19 | 409 | 89 | |||||
| Csa01g002210 | 1 | 411 | 89 | |||||
| Membrane-bound | ADS1 | AT1G06080 | 1 | 305 | Csa03g008530 | 3 | 923 | 97 |
| ADS2 | AT2G31360 | 2 | 307 | Csa16g012010 | 16 | 307 | 96 | |
| Csa07g011510 | 7 | 307 | 96 | |||||
| Csa05g029860 | 5 | 305 | 96 | |||||
| ADS3/FAD5 | AT3G15850 | 3 | 371 | Csa15g020490 | 15 | 372 | 95 | |
| Csa19g022640 | 19 | 537 | 95 | |||||
| Csa01g018460 | 1 | 373 | 95 | |||||
| ADS4 | AT1G06350 | 1 | 300 | * | ||||
| ADS5 | AT3G15870 | 3 | 361 | Csa19g022670 | 19 | 584 | 80 | |
| Csa01g018480 | 1 | 361 | 81 | |||||
| ADS6 | AT1G06360 | 1 | 299 | Csa03g009800 | 3 | 299 | 92 | |
| Csa17g009740 | 17 | 422 | 89 | |||||
| Csa14g007750 | 14 | 299 | 92 | |||||
| ADS7 | AT1G06100 | 1 | 299 | Csa17g009510 | 17 | 347 | 85 | |
| Csa14g007490 | 14 | 259 | 81 | |||||
| Csa17g009500 | 17 | 288 | 80 | |||||
| Csa14g007470 | 14 | 898 | 80 | |||||
| ADS8 | AT1G06090 | 1 | 299 | Csa17g009490 | 17 | 300 | 91 | |
| ADS9 | AT1G06120 | 1 | 299 | * | ||||
| DES1 | AT4G04930 | 4 | 332 | Csa13g052810 | 13 | 266 | 87 | |
| Csa08g048970 | 8 | 337 | 94 | |||||
| Csa02g011080 | 2 | 307 | 87 | |||||
| FAD2 | AT3G12120 | 3 | 383 | Csa15g016000 | 15 | 385 | 96 | |
| Csa19g016350 | 19 | 502 | 96 | |||||
| Csa01g013220 | 1 | 384 | 97 | |||||
| FAD3 | AT2G29980 | 2 | 386 | Csa16g014970 | 16 | 439 | 96 | |
| Csa07g013360 | 7 | 387 | 97 | |||||
| Csa05g033930 | 5 | 387 | 96 | |||||
| FAD4 | AT4G27030 | 4 | 323 | Csa11g017130 | 11 | 321 | 90 | |
| Csa10g015800 | 10 | 555 | 91 | |||||
| Csa12g024850 | 12 | 323 | 91 | |||||
| FAD6 | AT4G30950 | 4 | 448 | Csa11g012410 | 11 | 558 | 95 | |
| Csa10g011570 | 10 | 445 | 95 | |||||
| Csa12g016160 | 12 | 496 | 95 | |||||
| FAD7 | AT3G11170 | 3 | 446 | Csa15g014910 | 15 | 448 | 90 | |
| Csa19g015230 | 19 | 448 | 90 | |||||
| Csa01g012210 | 1 | 448 | 90 | |||||
| FAD8 | AT5G05580 | 5 | 435 | Csa13g007570 | 13 | 433 | 94 | |
| Csa08g058890 | 8 | 433 | 94 | |||||
| Csa20g006850 | 20 | 433 | 94 | |||||
| SLD1 | AT3G61580 | 3 | 449 | Csa16g003620 | 16 | 766 | 95 | |
| Csa07g003100 | 7 | 449 | 91 | |||||
| Csa05g093640 | 5 | 450 | 93 | |||||
| SLD2 | AT2G46210 | 2 | 449 | Csa04g065230 | 4 | 449 | 95 | |
| Csa06g052640 | 6 | 449 | 96 | |||||
| Csa05g003010 | 5 | 508 | 95 | |||||
| Motifs | Location (AA) | |
|---|---|---|
| FAB2 | HxxxxH | 133–136 |
| ENRHG | 283–287 | |
| DEKRHE | 394–400 | |
| ADS1 | HRNLAH | 83–88 |
| HRYHH | 120–124 | |
| HNNHH | 252–256 | |
| ADS2 | HRNLAH | 85–90 |
| HRYHH | 122–126 | |
| HNNHH | 254–258 | |
| ADS3 | HRYHH | 354–358 |
| HNNHH | 486–490 | |
| ADS5 | HRNLSH | 361–366 |
| HRNLSH | 398–403 | |
| HNNHH | 530–534 | |
| ADS6 | HRFHH | 114–118 |
| HNNHH | 369–373 | |
| ADS7 | HRFHH | 714–718 |
| HNNHH | 951–955 | |
| DES1 | HELSH | 106-110 |
| HLEHH | 143–147 | |
| HPMAGHFISEH | 238–248 | |
| FAD2 | HECGHH | 107–112 |
| HRRHH | 143–147 | |
| HVAHH | 435–439 | |
| FAD3 | HDCGH | 154–158 |
| HHQNH | 193–197 | |
| HHHGH | 314–318 | |
| HVIHH | 357–361 | |
| FAD4 | HAWAH | 465–469 |
| HAEHH | 494–498 | |
| FAD6 | HDCAH | 319–323 |
| HDRHH | 355–359 | |
| HHTAPH | 472–477 | |
| HIPHH | 515–519 | |
| FAD7 | HDCGH | 170–174 |
| HHQNH | 209–213 | |
| HHHGH | 330–334 | |
| HVIHH | 373–377 | |
| FAD8 | HDCGH | 158–162 |
| HRTHH | 198–198 | |
| HHHGH | 318–322 | |
| HVIHH | 361–365 | |
| SLD | HPGTAWHH | 182–189 |
| HIKDFH | 398–403 | |
| HDSGH | 477–481 | |
| HNAHH | 573–577 | |
| HDPDLQH | 585–591 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raboanatahiry, N.; Yin, Y.; Chen, K.; He, J.; Yu, L.; Li, M. In Silico Analysis of Fatty Acid Desaturases Structures in Camelina sativa, and Functional Evaluation of Csafad7 and Csafad8 on Seed Oil Formation and Seed Morphology. Int. J. Mol. Sci. 2021, 22, 10857. https://doi.org/10.3390/ijms221910857
Raboanatahiry N, Yin Y, Chen K, He J, Yu L, Li M. In Silico Analysis of Fatty Acid Desaturases Structures in Camelina sativa, and Functional Evaluation of Csafad7 and Csafad8 on Seed Oil Formation and Seed Morphology. International Journal of Molecular Sciences. 2021; 22(19):10857. https://doi.org/10.3390/ijms221910857
Chicago/Turabian StyleRaboanatahiry, Nadia, Yongtai Yin, Kang Chen, Jianjie He, Longjiang Yu, and Maoteng Li. 2021. "In Silico Analysis of Fatty Acid Desaturases Structures in Camelina sativa, and Functional Evaluation of Csafad7 and Csafad8 on Seed Oil Formation and Seed Morphology" International Journal of Molecular Sciences 22, no. 19: 10857. https://doi.org/10.3390/ijms221910857
APA StyleRaboanatahiry, N., Yin, Y., Chen, K., He, J., Yu, L., & Li, M. (2021). In Silico Analysis of Fatty Acid Desaturases Structures in Camelina sativa, and Functional Evaluation of Csafad7 and Csafad8 on Seed Oil Formation and Seed Morphology. International Journal of Molecular Sciences, 22(19), 10857. https://doi.org/10.3390/ijms221910857