LysM Receptor-Like Kinase LYK9 of Pisum Sativum L. May Regulate Plant Responses to Chitooligosaccharides Differing in Structure

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Identification of Pea Lyk9 Mutant Lines in Targeting-Induced Local Lesions in Genomes (TILLING) Collection
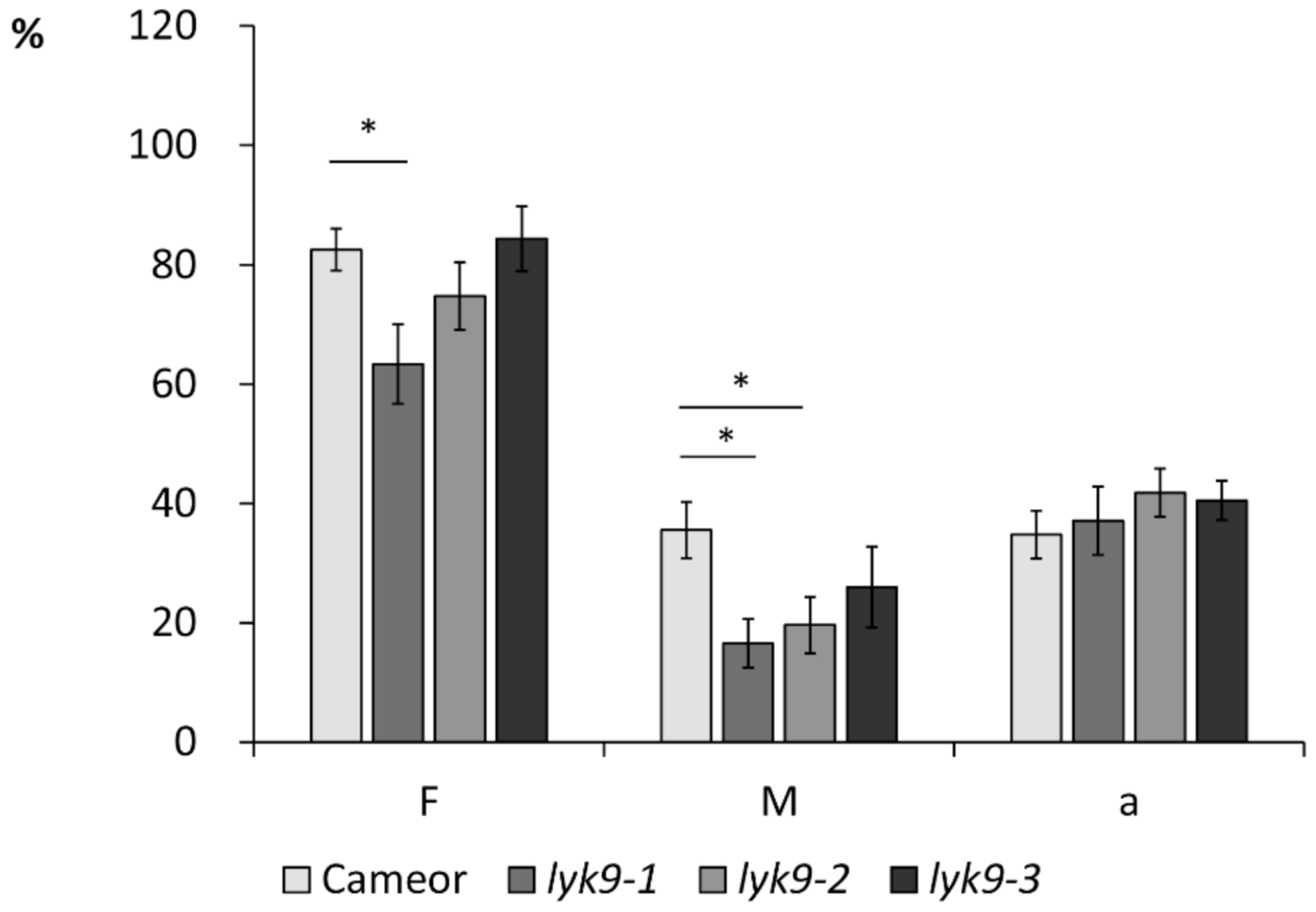
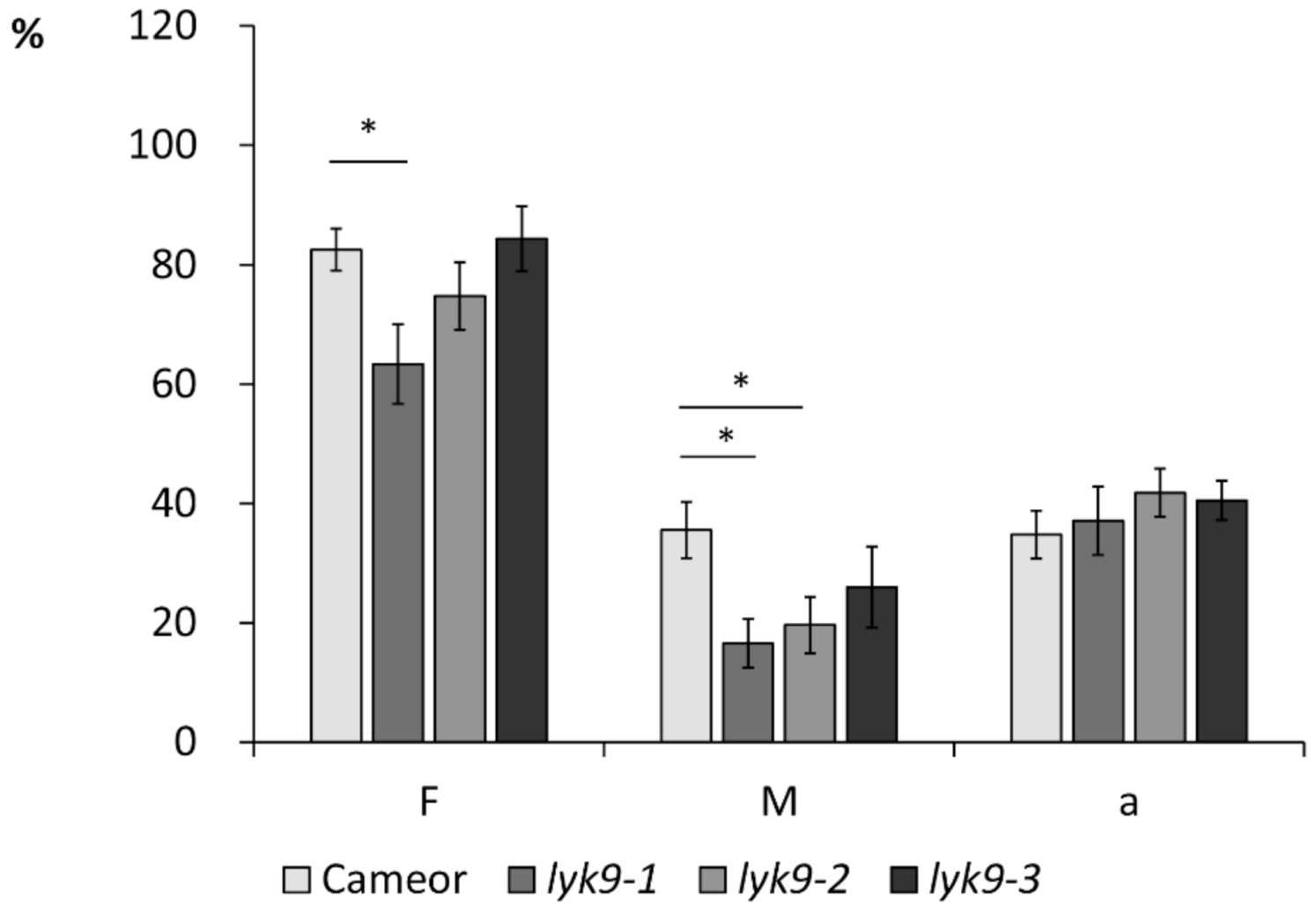
2.2. Analysis of Symbiosis Development with AM Fungi R. irregularis in Lyk9 Mutant Lines
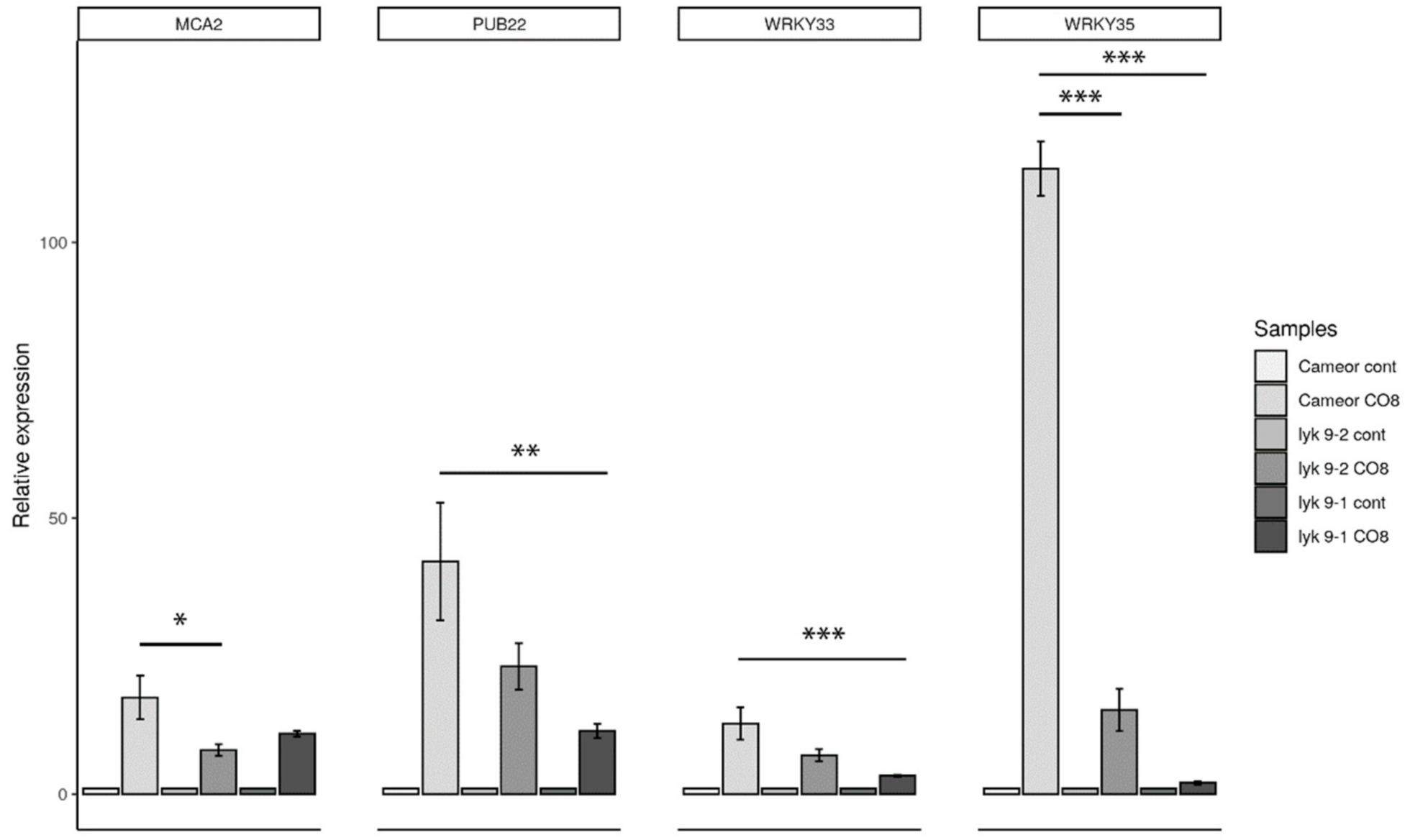
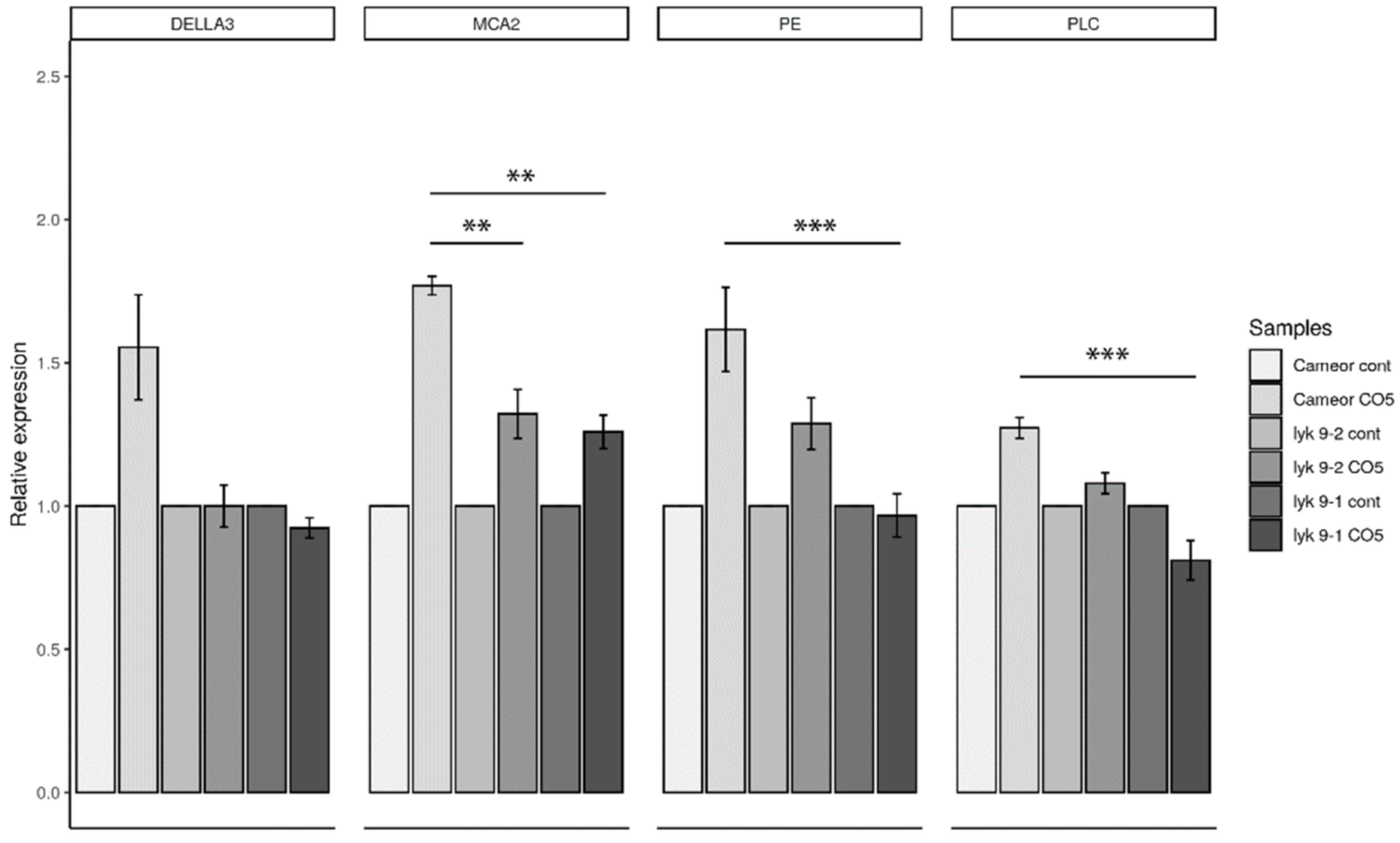
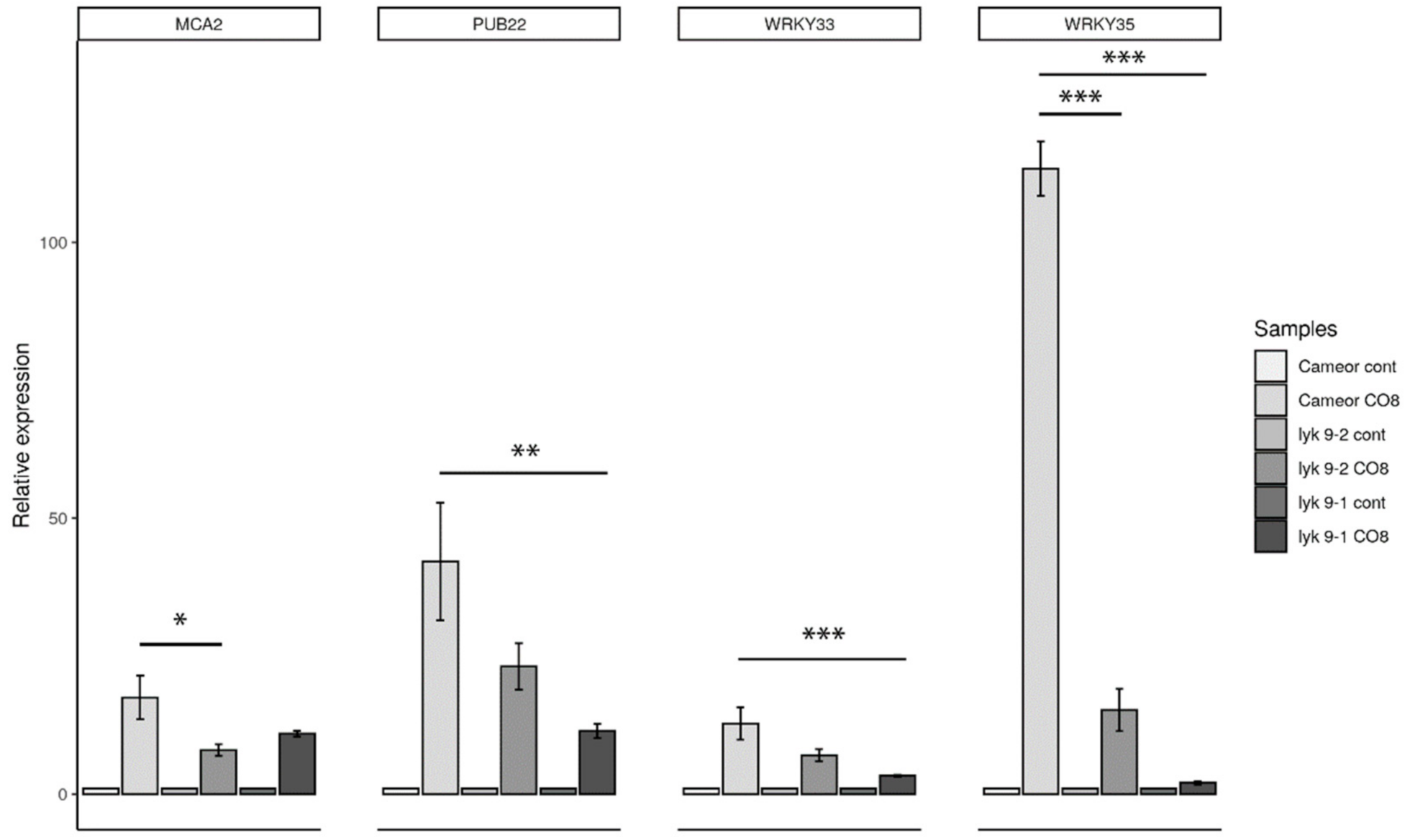
2.3. Analysis of the Symbiotic and Defense Marker Gene Expression in Wild-Type Pea Plants and Mutants Impaired in the Lyk9 Gene after CO5 and CO8-DA Treatment
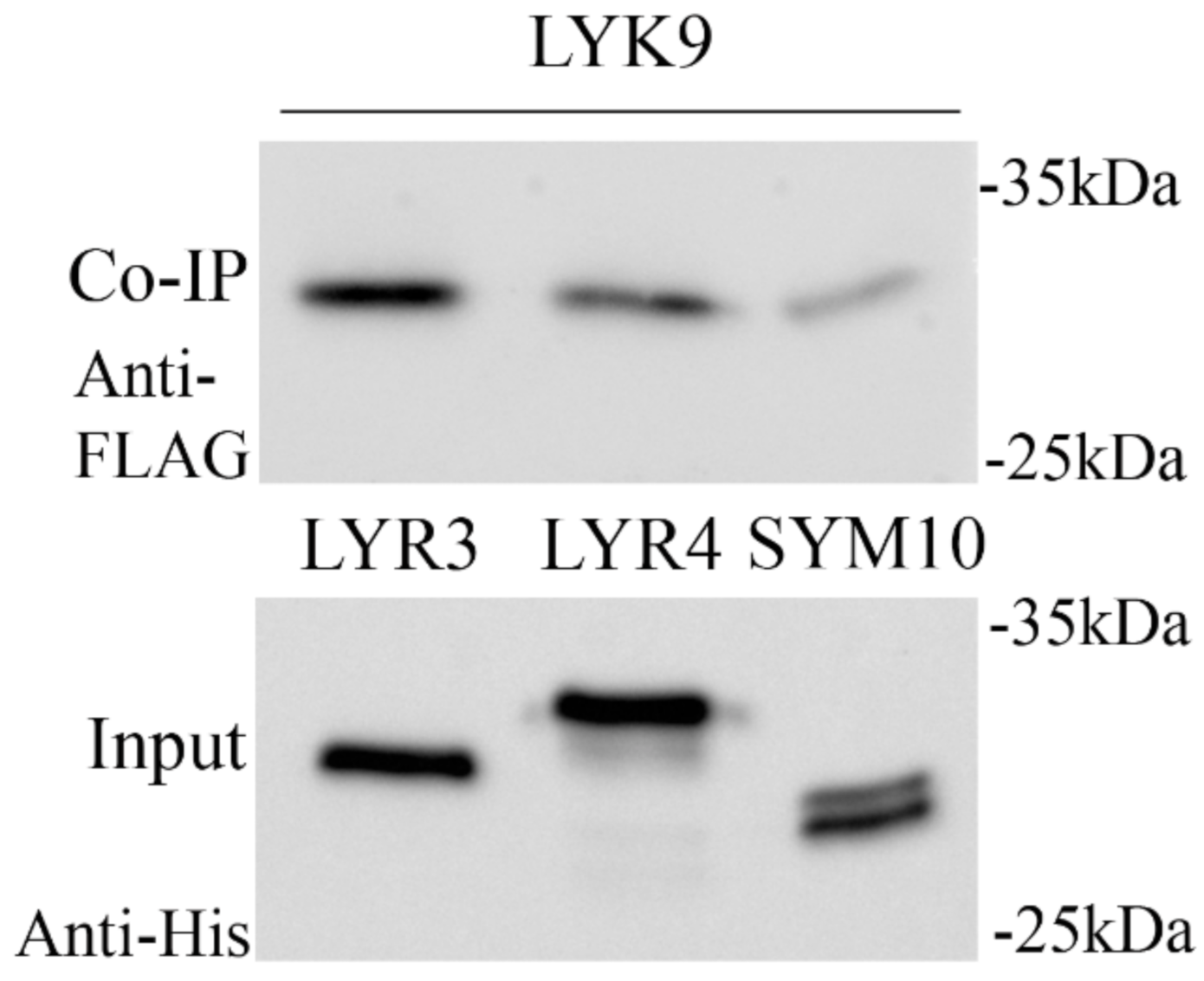
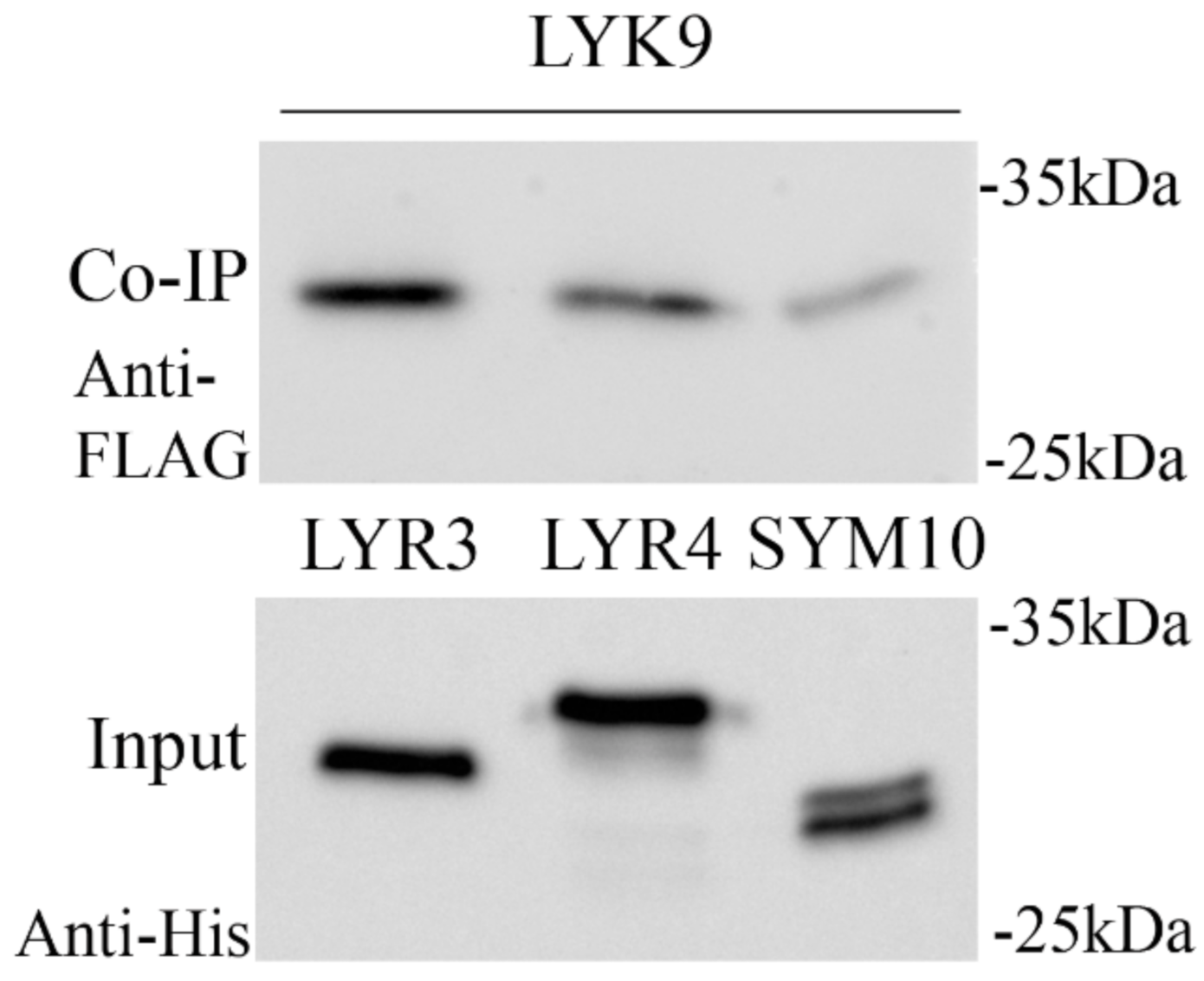
2.4. Synthesis of the PsLYK9-ECD, PsLYR4-ECD and PsLYR3-ECD Extracellular Domains (ECDs) in E. coli and Co-Immunoprecipitation Assay
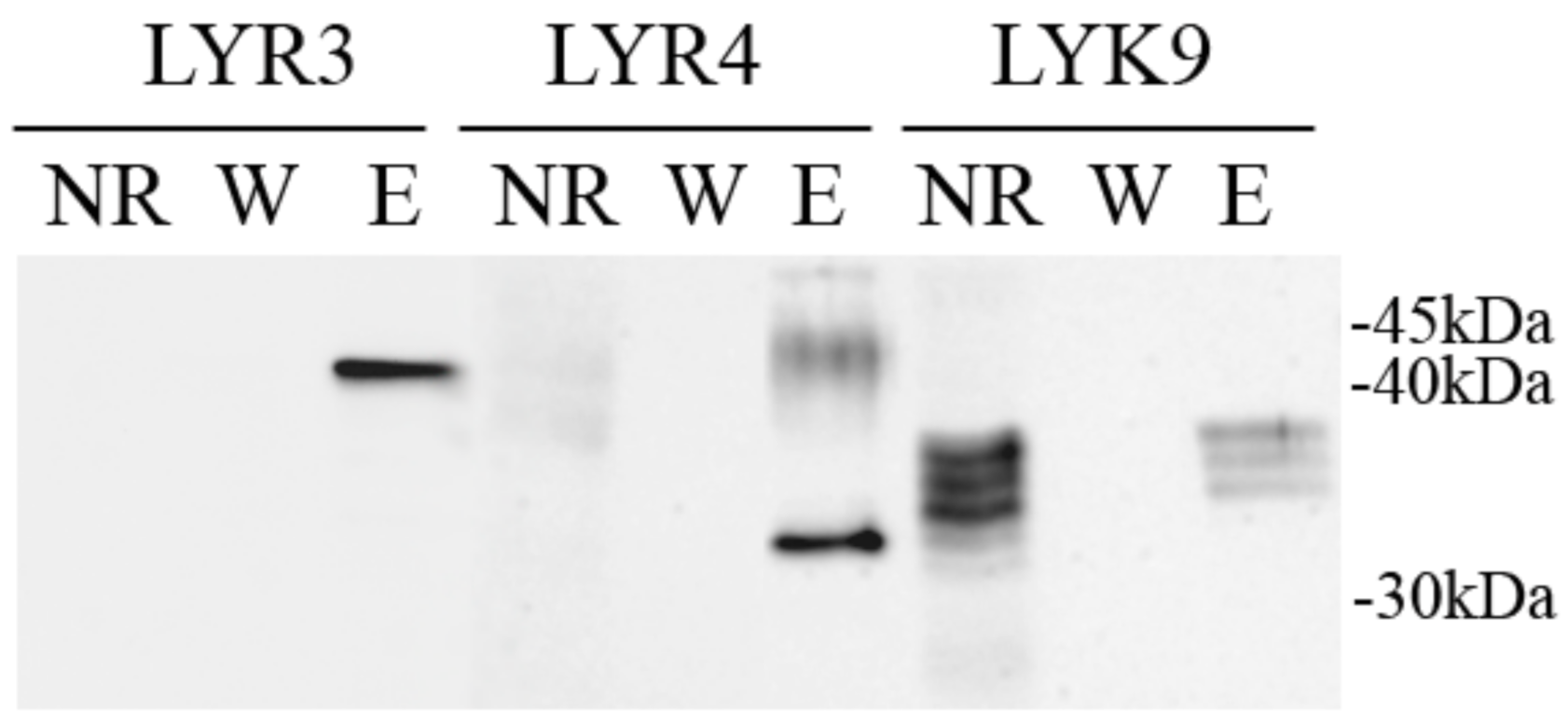
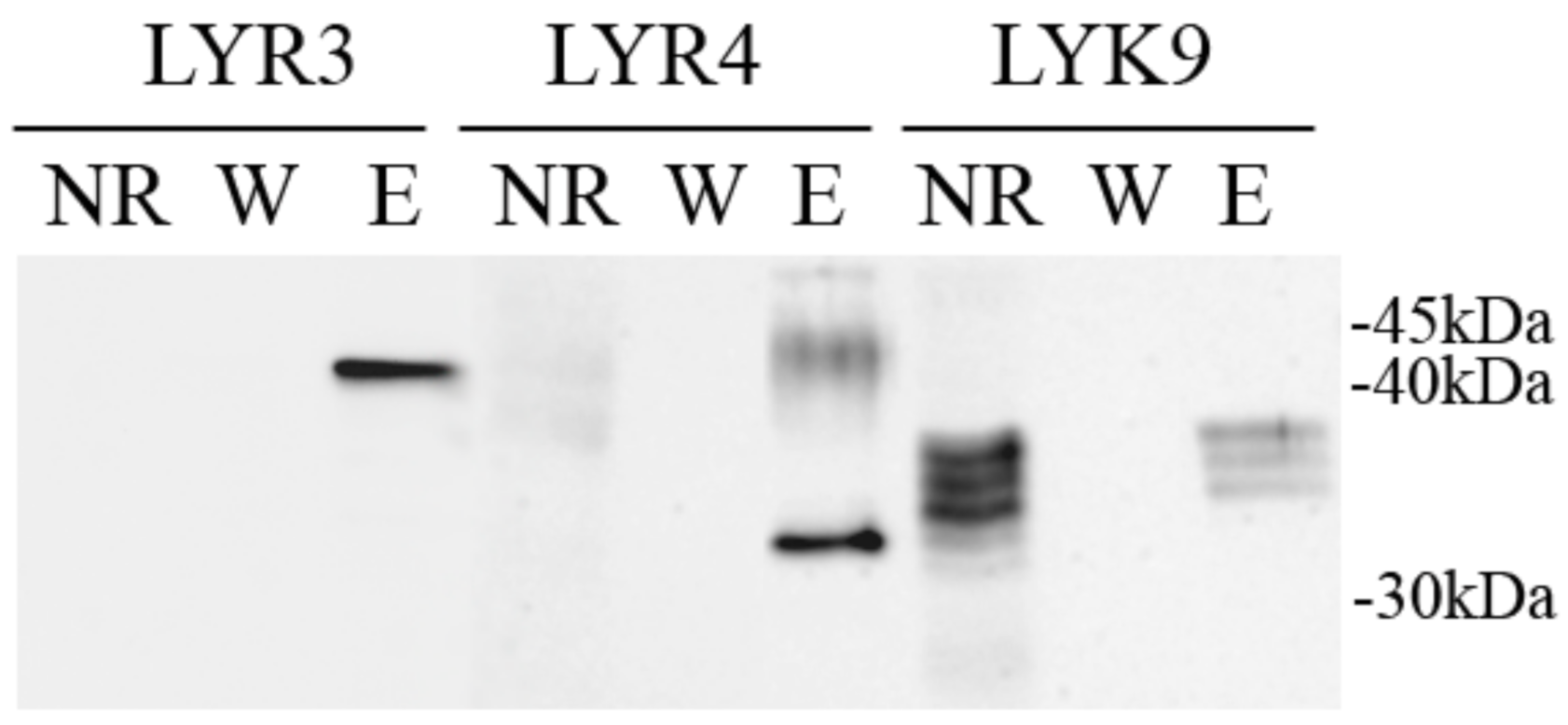
2.5. Synthesis of the PsLYK9-ECD, PsLYR4-ECD and PsLYR3-ECD ECDs in Insect Cells
2.6. Analysis of Receptor Binding with Ligands Using Thermophoresis
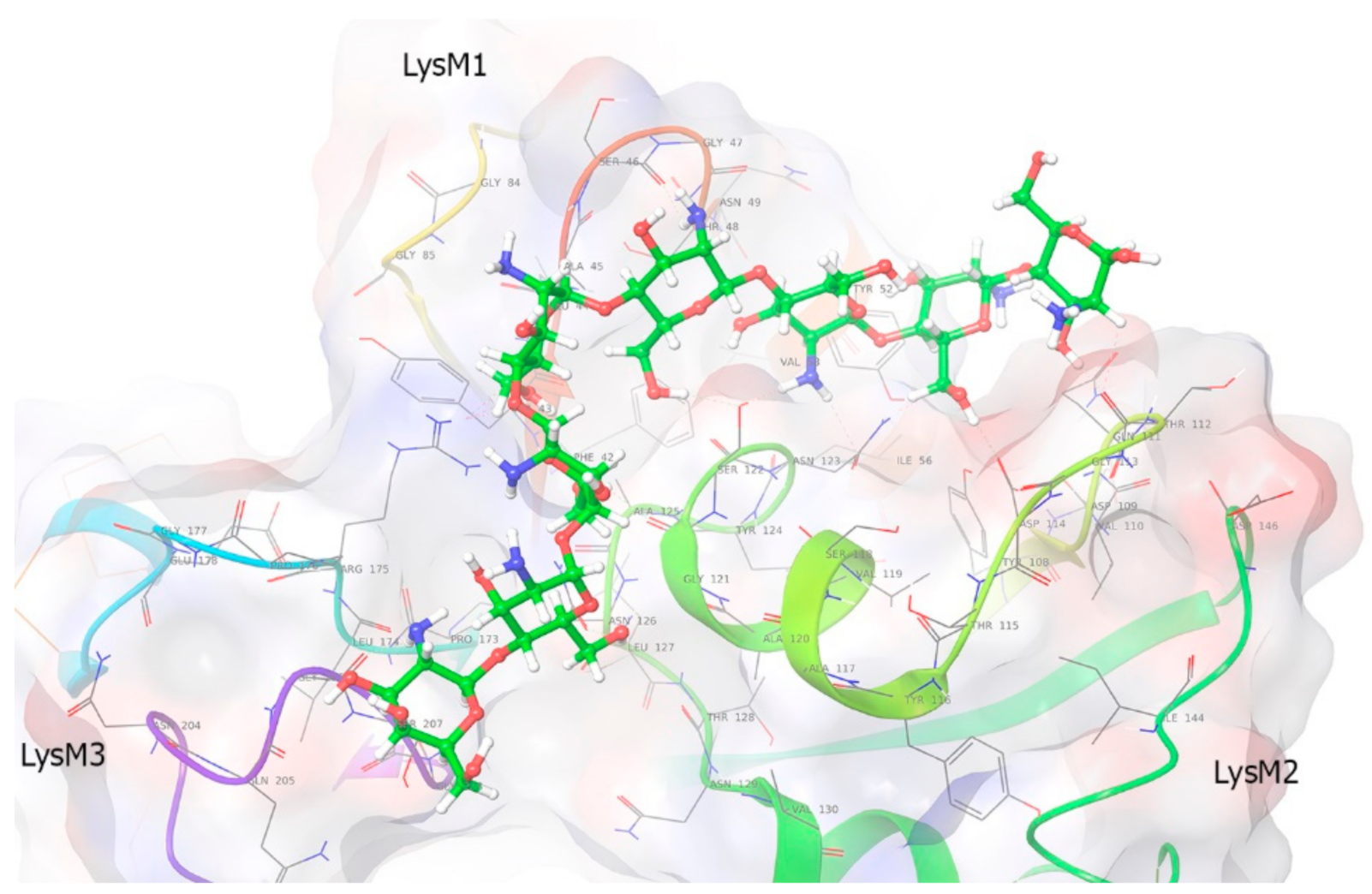
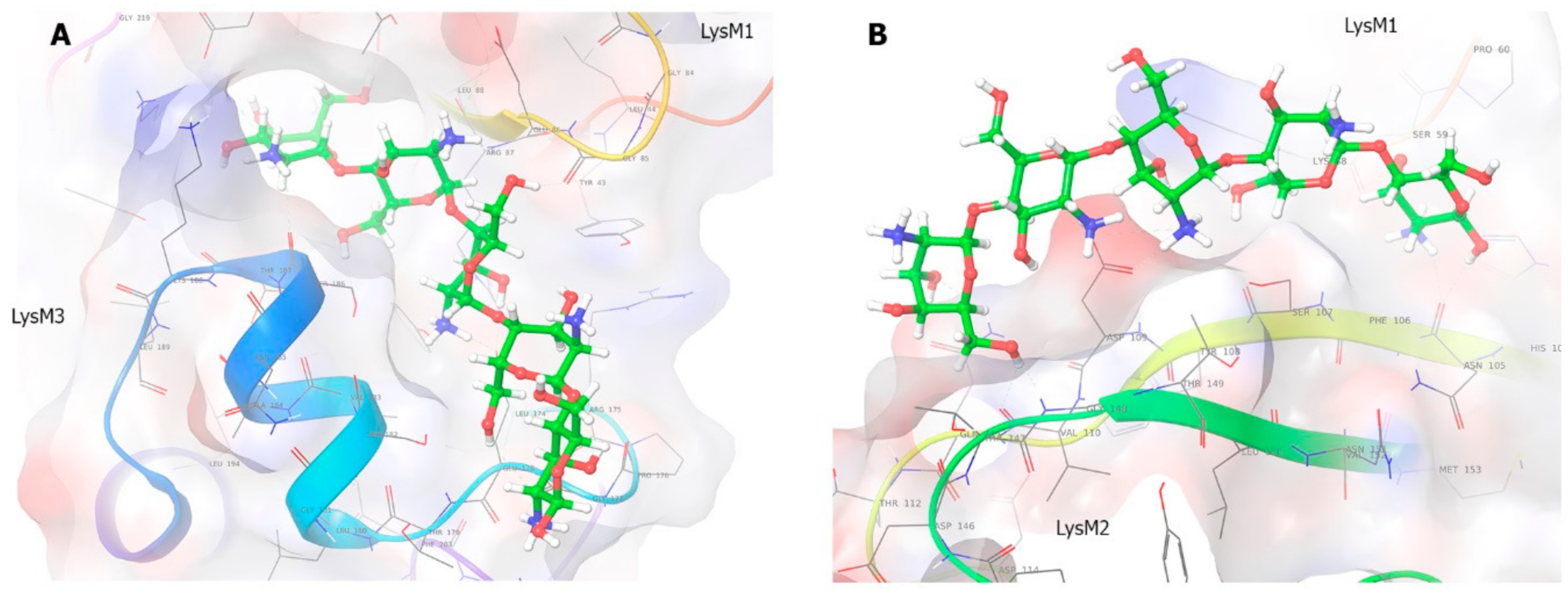
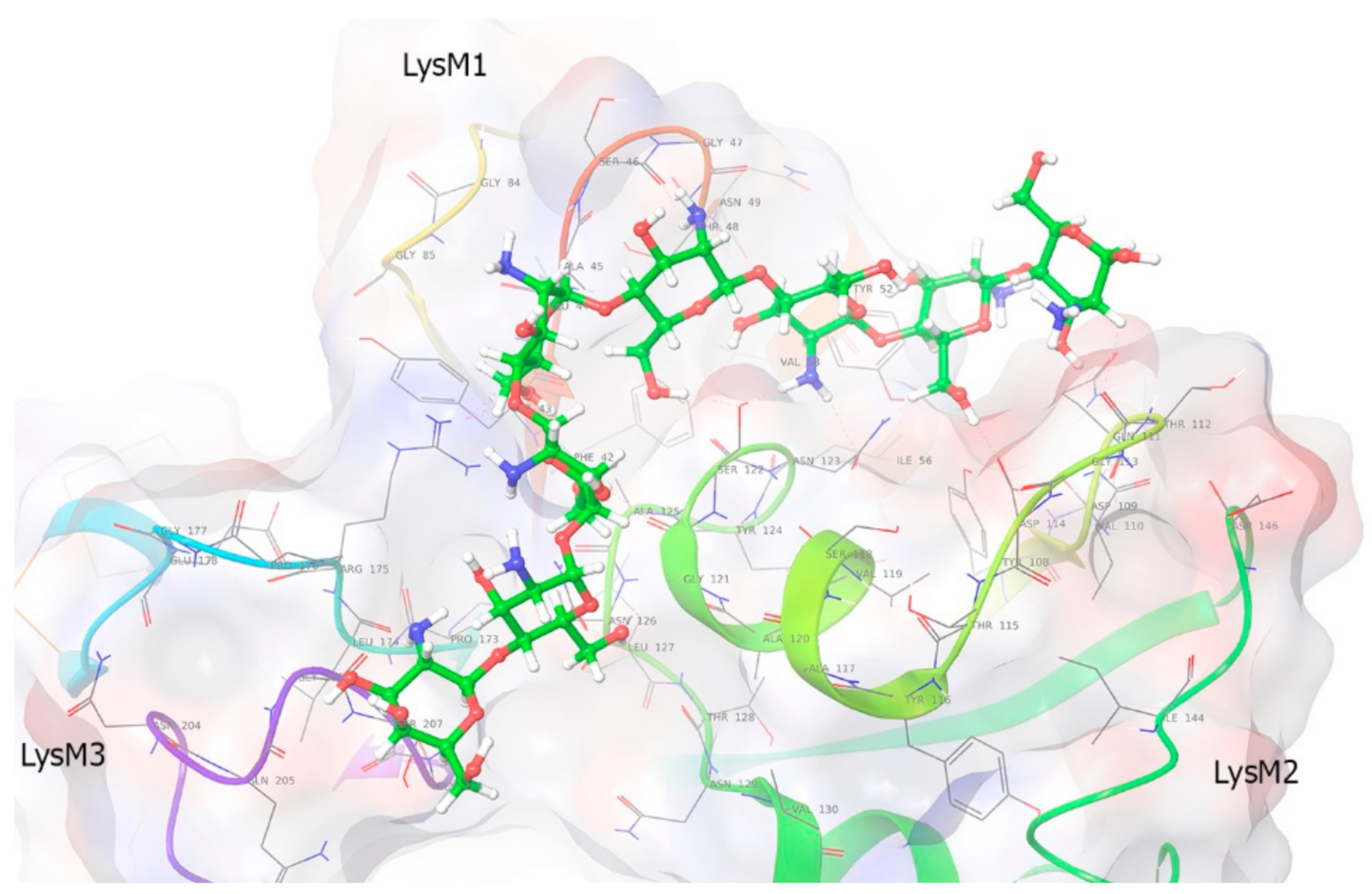
2.7. Molecular Modeling
3. Discussion
4. Materials and Methods
4.1. Plants and Microorganisms
4.2. Plant Growth Conditions
4.3. Targeting-Induced Local Lesions in Genomes (TILLING) Screens
4.4. Fungal Inoculum Preparation
4.5. Plant Inoculation with Rhizophagus Irregularis
4.6. Analysis of Mycorrhization
4.7. Treatment of Pea Seedlings with COs
4.8. Molecular Cloning
4.9. Bacmid Isolation and Purification
4.10. Heterologous Synthesis of PsLYK9-ECD, PsLYR4-ECD and PsLYR3-ECD Extracellular Domains in Sf9 Insect Cell
4.11. Co-Immunoprecipitation Assay
4.12. Microscale Thermophoresis
4.13. RNA Isolation and cDNA Synthesis
4.14. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
4.15. Molecular Modeling
4.16. Statistical Methods and Computer Software
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boller, T.; He, S.Y. Innate immunity in plants: An arms race between pattern recognition receptors in plants and effectors in microbial pathogens. Science 2009, 324, 742–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zipfel, C. Pattern-recognition receptors in plant innate immunity. Curr. Opin. Immunol. 2008, 20, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Felix, G.; Duran, J.D.; Volko, S.; Boller, T. Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. Plant J. 1999, 18, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gómez, L.; Boller, T. Fls2: An LRR Receptor-like Kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Mol. Cell 2000, 5, 1003–1011. [Google Scholar] [CrossRef]
- Chinchilla, D.; Zipfel, C.; Robatzek, S.; Kemmerling, B.; Nürnberger, T.; Jones, J.D.G.; Felix, G.; Boller, T. A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature 2007, 448, 497–500. [Google Scholar] [CrossRef]
- Wang, S.; Sun, Z.; Wang, H.; Liu, L.; Lu, F.; Yang, J.; Zhang, M.; Zhang, S.; Guo, Z.; Bent, A.F.; et al. Rice OsFLS2-Mediated Perception of Bacterial Flagellins Is Evaded by Xanthomonas oryzae pvs. oryzae and oryzicola. Mol. Plant 2015, 8, 1024–1037. [Google Scholar] [CrossRef] [Green Version]
- Kunze, G.; Zipfel, C.; Robatzek, S.; Niehaus, K.; Boller, T.; Felix, G. The N terminus of bacterial elongation factor Tu elicits innate immunity in Arabidopsis plants. Plant Cell 2004, 16, 3496–3507. [Google Scholar] [CrossRef] [Green Version]
- Zipfel, C.; Felix, G. Plants and animals: A different taste for microbes? Curr. Opin. Plant Biol. 2005, 8, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C.; Kunze, G.; Chinchilla, D.; Caniard, A.; Jones, J.D.G.; Boller, T.; Felix, G. Perception of the Bacterial PAMP EF-Tu by the Receptor EFR Restricts Agrobacterium-Mediated Transformation. Cell 2006, 125, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Roux, M.; Schwessinger, B.; Albrecht, C.; Chinchilla, D.; Jones, A.; Holton, N.; Malinovsky, F.G.; Tör, M.; de Vries, S.; Zipfel, C. The Arabidopsis leucine-rich repeat receptor-like kinases BAK1/SERK3 and BKK1/SERK4 are required for innate immunity to hemibiotrophic and biotrophic pathogens. Plant Cell 2011, 23, 2440–2455. [Google Scholar] [CrossRef] [Green Version]
- Feng, F.; Sun, J.; Radhakrishnan, G.V.; Lee, T.; Bozsóki, Z.; Fort, S.; Gavrin, A.; Gysel, K.; Thygesen, M.B.; Andersen, K.R.; et al. A combination of chitooligosaccharide and lipochitooligosaccharide recognition promotes arbuscular mycorrhizal associations in Medicago truncatula. Nat. Commun. 2019, 10, 5047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maillet, F.; Poinsot, V.; André, O.; Puech-Pagés, V.; Haouy, A.; Gueunier, M.; Cromer, L.; Giraudet, D.; Formey, D.; Niebel, A.; et al. Fungal lipochitooligosaccharide symbiotic signals in arbuscular mycorrhiza. Nature 2011, 469, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Genre, A.; Chabaud, M.; Balzergue, C.; Puech-Pagès, V.; Novero, M.; Rey, T.; Fournier, J.; Rochange, S.; Bécard, G.; Bonfante, P.; et al. Short-chain chitin oligomers from arbuscular mycorrhizal fungi trigger nuclear Ca2+ spiking in Medicago truncatula roots and their production is enhanced by strigolactone. New Phytol. 2013, 198, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Bozsoki, Z.; Cheng, J.; Feng, F.; Gysel, K.; Vinther, M.; Andersen, K.R.; Oldroyd, G.; Blaise, M.; Radutoiu, S.; Stougaard, J. Receptor-mediated chitin perception in legume roots is functionally separable from Nod factor perception. Proc. Natl. Acad. Sci. USA 2017, 114, E8118–E8127. [Google Scholar] [CrossRef] [Green Version]
- Carotenuto, G.; Chabaud, M.; Miyata, K.; Capozzi, M.; Takeda, N.; Kaku, H.; Shibuya, N.; Nakagawa, T.; Barker, D.G.; Genre, A. The rice LysM receptor-like kinase OsCERK1 is required for the perception of short-chain chitin oligomers in arbuscular mycorrhizal signaling. New Phytol. 2017, 214, 1440–1446. [Google Scholar] [CrossRef] [Green Version]
- Leppyanen, I.V.; Kirienko, A.N.; Lobov, A.A.; Dolgikh, E.A. Differential proteome analysis of pea roots at the early stages of symbiosis with nodule bacteria. Vavilovskii Zhurnal Genet. Selektsii 2018, 22, 196–204. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Liu, Z.; Song, C.; Hu, Y.; Han, Z.; She, J.; Fan, G.; Wang, J.; Jin, C.; Chang, J.; et al. Chitin-induced dimerization activates a plant immune receptor. Science 2012, 336, 1160–1164. [Google Scholar] [CrossRef] [Green Version]
- Willmann, R.; Lajunen, H.M.; Erbs, G.; Newman, M.-A.; Kolb, D.; Tsuda, K.; Katagiri, F.; Fliegmann, J.; Bono, J.-J.; Cullimore, J.V.; et al. Arabidopsis lysin-motif proteins LYM1 LYM3 CERK1 mediate bacterial peptidoglycan sensing and immunity to bacterial infection. Proc. Natl. Acad. Sci. USA 2011, 108, 19824–19829. [Google Scholar] [CrossRef] [Green Version]
- Kaku, H.; Nishizawa, Y.; Ishii-Minami, N.; Akimoto-Tomiyama, C.; Dohmae, N.; Takio, K.; Minami, E.; Shibuya, N. Plant cells recognize chitin fragments for defense signaling through a plasma membrane receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 11086–11091. [Google Scholar] [CrossRef] [Green Version]
- Miya, A.; Albert, P.; Shinya, T.; Desaki, Y.; Ichimura, K.; Shirasu, K.; Narusaka, Y.; Kawakami, N.; Kaku, H.; Shibuya, N. CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 19613–19618. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Liang, Y.; Tanaka, K.; Nguyen, C.T.; Jedrzejczak, R.P.; Joachimiak, A.; Stacey, G. The kinase LYK5 is a major chitin receptor in Arabidopsis and forms a chitin-induced complex with related kinase CERK1. eLife 2014, 3, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Nakano, T.; Takamizawa, D.; Desaki, Y.; Ishii-Minami, N.; Nishizawa, Y.; Minami, E.; Okada, K.; Yamane, H.; Kaku, H.; et al. Two LysM receptor molecules, CEBiP and OsCERK1, cooperatively regulate chitin elicitor signaling in rice. Plant J. 2010, 64, 204–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gubaeva, E.; Gubaev, A.; Melcher, R.L.J.; Cord-Landwehr, S.; Singh, R.; El Gueddari, N.E.; Moerschbacher, B.M. Slipped sandwich’ model for chitin and chitosan perception in arabidopsis. Mol. Plant-Microbe Interact. 2018, 31, 1145–1153. [Google Scholar] [CrossRef] [Green Version]
- Shinya, T.; Motoyama, N.; Ikeda, A.; Wada, M.; Kamiya, K.; Hayafune, M.; Kaku, H.; Shibuya, N. Functional characterization of CEBiP and CERK1 homologs in arabidopsis and rice reveals the presence of different chitin receptor systems in plants. Plant Cell Physiol. 2012, 53, 1696–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouzai, Y.; Mochizuki, S.; Nakajima, K.; Desaki, Y.; Hayafune, M.; Miyazaki, H.; Yokotani, N.; Ozawa, K.; Minami, E.; Kaku, H.; et al. Targeted gene disruption of OsCERK1 reveals its indispensable role in chitin perception and involvement in the peptidoglycan response and immunity in rice. Mol. Plant Microbe Interact. 2014, 27, 975–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Dong, W.; Sun, J.; Feng, F.; Deng, Y.; He, Z.; Oldroyd, G.E.D.; Wang, E. The receptor kinase CERK1 has dual functions in symbiosis and immunity signalling. Plant J. 2015, 81, 258–267. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Gao, C.; Guo, G.; Liu, J.; Gao, Y.; Pan, R.; Guan, Y.; Hu, J. Maize annexin genes ZmANN33 and ZmANN35 encode proteins that function in cell membrane recovery during seed germination. J. Exp. Bot. 2019, 70, 1183–1195. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yuan, L.; Staehelin, C.; Li, Y.; Ruan, J.; Liang, Z.; Xie, Z.; Wang, W.; Xie, J.; Huang, S. The LYSIN MOTIF-CONTAINING RECEPTOR-LIKE KINASE 1 protein of banana is required for perception of pathogenic and symbiotic signals. New Phytol. 2019, 223, 1530–1546. [Google Scholar] [CrossRef]
- Gibelin-Viala, C.; Amblard, E.; Puech-Pages, V.; Bonhomme, M.; Garcia, M.; Bascaules-Bedin, A.; Fliegmann, J.; Wen, J.; Mysore, K.S.; le Signor, C.; et al. The Medicago truncatula LysM receptor-like kinase LYK9 plays a dual role in immunity and the arbuscular mycorrhizal symbiosis. New Phytol. 2019, 223, 1516–1529. [Google Scholar] [CrossRef]
- Dalmais, M.; Schmidt, J.; Le Signor, C.; Moussy, F.; Burstin, J.; Savois, V.; Aubert, G.; Brunaud, V.; de Oliveira, Y.; Guichard, C.; et al. UTILLdb, a Pisum sativum in silico forward and reverse genetics tool. Genome Biol. 2008, 9, R43. [Google Scholar] [CrossRef] [Green Version]
- Kendra, D.F.; Hadwiger, L.A. Characterization of the smallest chitosan oligomer that is maximally antifungal to Fusarium solani and elicits pisatin formation in Pisum sativum. Exp. Mycol. 1984, 8, 276–281. [Google Scholar] [CrossRef]
- Kendra, D.F.; Christian, D.; Hadwiger, L.A. Chitosan oligomers from Fusarium solani/pea interactions, chitinase/β-glucanase digestion of sporelings and from fungal wall chitin actively inhibit fungal growth and enhance disease resistance. Physiol. Mol. Plant Pathol. 1989, 35, 215–230. [Google Scholar] [CrossRef]
- Hadwiger, L.A.; Ogawa, T.; Kuyama, H. Chitosan polymer sizes effective in inducing phytoalexin accumulation and fungal suppression are verified with synthesized oligomers. Mol. Plant-Microbe Interact. 1994, 7, 531–533. [Google Scholar] [CrossRef] [PubMed]
- Hadwiger, L.A.; Chang, M.-M. Low level DNA damage occurs as PAMPs, chitin and flg22, activates PR genes, and increases pisatin and disease resistance in pea endocarp tissue. New Negat. Plant Sci. 2015, 1–2, 6–15. [Google Scholar] [CrossRef] [Green Version]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Ng, P.C.; Henikoff, S. SIFT: Predicting amino acid changes that affect protein function. Nucleic Acids Res. 2003, 31, 3812–3814. [Google Scholar] [CrossRef] [Green Version]
- Shtark, O.Y.; Sulima, A.S.; Zhernakov, A.I.; Kliukova, M.S.; Fedorina, J.V.; Pinaev, A.G.; Kryukov, A.A.; Akhtemova, G.A.; Tikhonovich, I.A.; Zhukov, V.A. Arbuscular mycorrhiza development in pea (Pisum sativum L.) mutants impaired in five early nodulation genes including putative orthologs of NSP1 and NSP2. Symbiosis 2016, 68, 129–144. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. Agricultural experiment station the water-culture method for growing plants without soil. Calif. Exp. Stn. 1938, 347, 32. [Google Scholar]
- Vierheilig, H.; Coughlan, A.P.; Wyss, U.; Piché, Y. Ink and vinegar, a simple staining technique for arbuscular-mycorrhizal fungi. Appl. Environ. Microbiol. 1998, 64, 5004–5007. [Google Scholar] [CrossRef] [Green Version]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Mesure du taux de mycorhization VA d’un système radiculaire. Recherche de méthodes d’estimation ayant une signification fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae, INRA ed.; Gianinazzi-Pearson, V., Gianinazzi, S., Eds.; INRA: Paris, France, 1986. [Google Scholar]
- Little, T.M.; Hills, F.J. Agricultural Experimentation: Design and Analysis; John Wiley & Sons, Ltd.: New York, NY, USA, 1978; ISBN 978-0-471-02352-4. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutant Line | Mutation | DNA Position | Protein Position | Localization |
|---|---|---|---|---|
| 3631 (lyk9-1) | G→A | 4533 | G485R | kinase domain, the activation loop |
| 3079 (lyk9-2) | C→T | 4340 | L470F | kinase domain, the activation loop |
| 4249 (lyk9-3) | C→T | 512 | P145L | LysM2 motif of extracellular domain |
| Protein | Ligand | Kd, µM (±SD) |
|---|---|---|
| LYR4-ECD | CO4 | - |
| CO5 | - | |
| CO5-DA | 45 ± 20 | |
| CO6-DA | 22 ± 11 | |
| CO7-DA | 19 ± 13 | |
| CO8-DA | 8 ± 5 | |
| LYK9-ECD | CO4 | 2.1 ± 0.6 |
| CO5 | 5 ± 1.9 | |
| CO5-DA | 76 ± 25 | |
| CO6-DA | 42 ± 23 | |
| CO7-DA | 31 ± 11 | |
| CO8-DA | 26 ± 15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leppyanen, I.V.; Pavlova, O.A.; Vashurina, M.A.; Bovin, A.D.; Dolgikh, A.V.; Shtark, O.Y.; Sendersky, I.V.; Dolgikh, V.V.; Tikhonovich, I.A.; Dolgikh, E.A. LysM Receptor-Like Kinase LYK9 of Pisum Sativum L. May Regulate Plant Responses to Chitooligosaccharides Differing in Structure. Int. J. Mol. Sci. 2021, 22, 711. https://doi.org/10.3390/ijms22020711
Leppyanen IV, Pavlova OA, Vashurina MA, Bovin AD, Dolgikh AV, Shtark OY, Sendersky IV, Dolgikh VV, Tikhonovich IA, Dolgikh EA. LysM Receptor-Like Kinase LYK9 of Pisum Sativum L. May Regulate Plant Responses to Chitooligosaccharides Differing in Structure. International Journal of Molecular Sciences. 2021; 22(2):711. https://doi.org/10.3390/ijms22020711
Chicago/Turabian StyleLeppyanen, Irina V., Olga A. Pavlova, Maria A. Vashurina, Andrey D. Bovin, Alexandra V. Dolgikh, Oksana Y. Shtark, Igor V. Sendersky, Vyacheslav V. Dolgikh, Igor A. Tikhonovich, and Elena A. Dolgikh. 2021. "LysM Receptor-Like Kinase LYK9 of Pisum Sativum L. May Regulate Plant Responses to Chitooligosaccharides Differing in Structure" International Journal of Molecular Sciences 22, no. 2: 711. https://doi.org/10.3390/ijms22020711
APA StyleLeppyanen, I. V., Pavlova, O. A., Vashurina, M. A., Bovin, A. D., Dolgikh, A. V., Shtark, O. Y., Sendersky, I. V., Dolgikh, V. V., Tikhonovich, I. A., & Dolgikh, E. A. (2021). LysM Receptor-Like Kinase LYK9 of Pisum Sativum L. May Regulate Plant Responses to Chitooligosaccharides Differing in Structure. International Journal of Molecular Sciences, 22(2), 711. https://doi.org/10.3390/ijms22020711