In Vitro Macrophage Immunomodulation by Poly(ε-caprolactone) Based-Coated AZ31 Mg Alloy


 ,
, 

 and
and 
Abstract
:1. Introduction
2. Results
2.1. Materials Surface Characterization
2.2. In Vitro Release of Coumarin
2.3. Electrochemical Behaviour
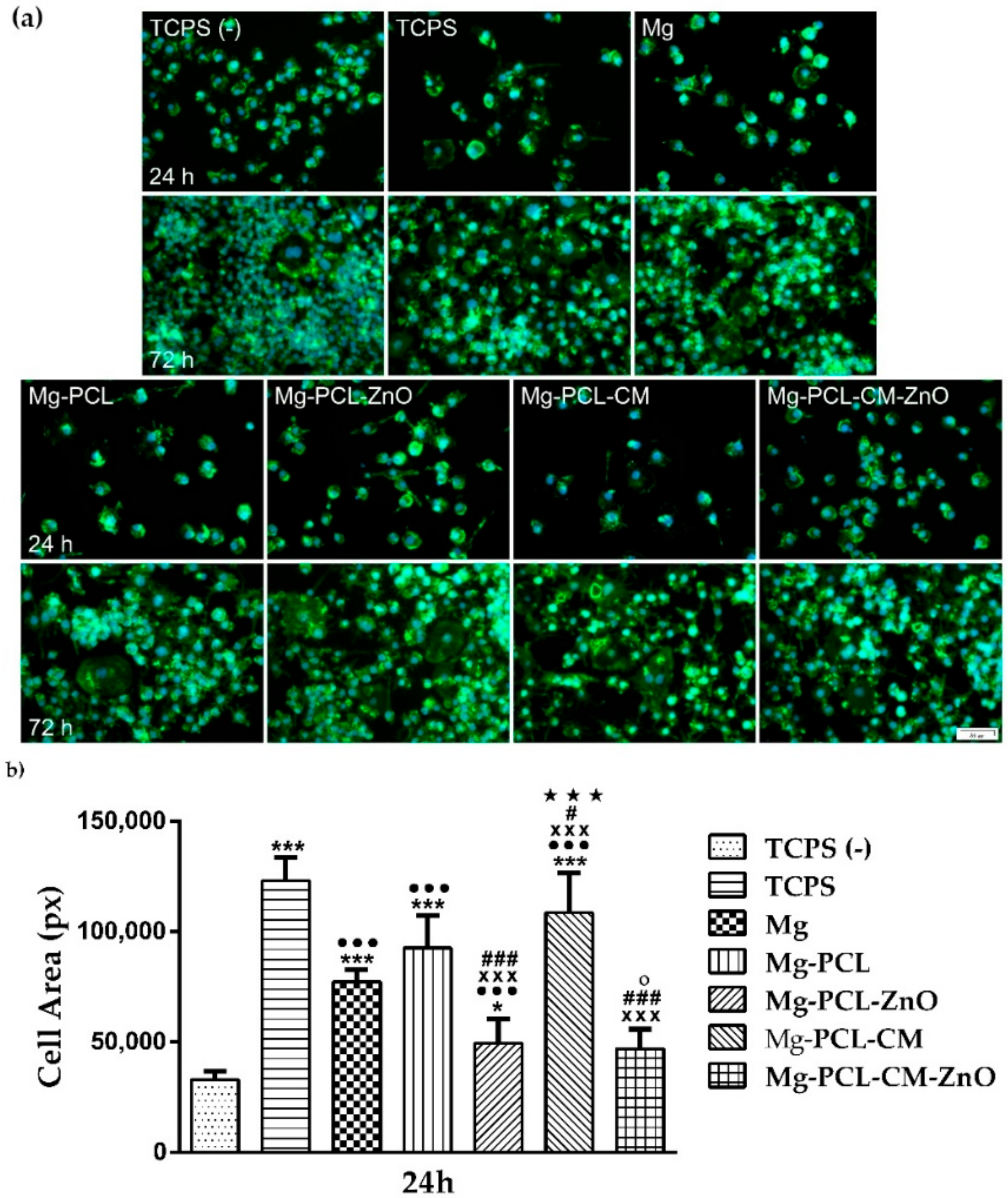
2.4. Macrophage Viability and Proliferation
2.5. Cell Morphology
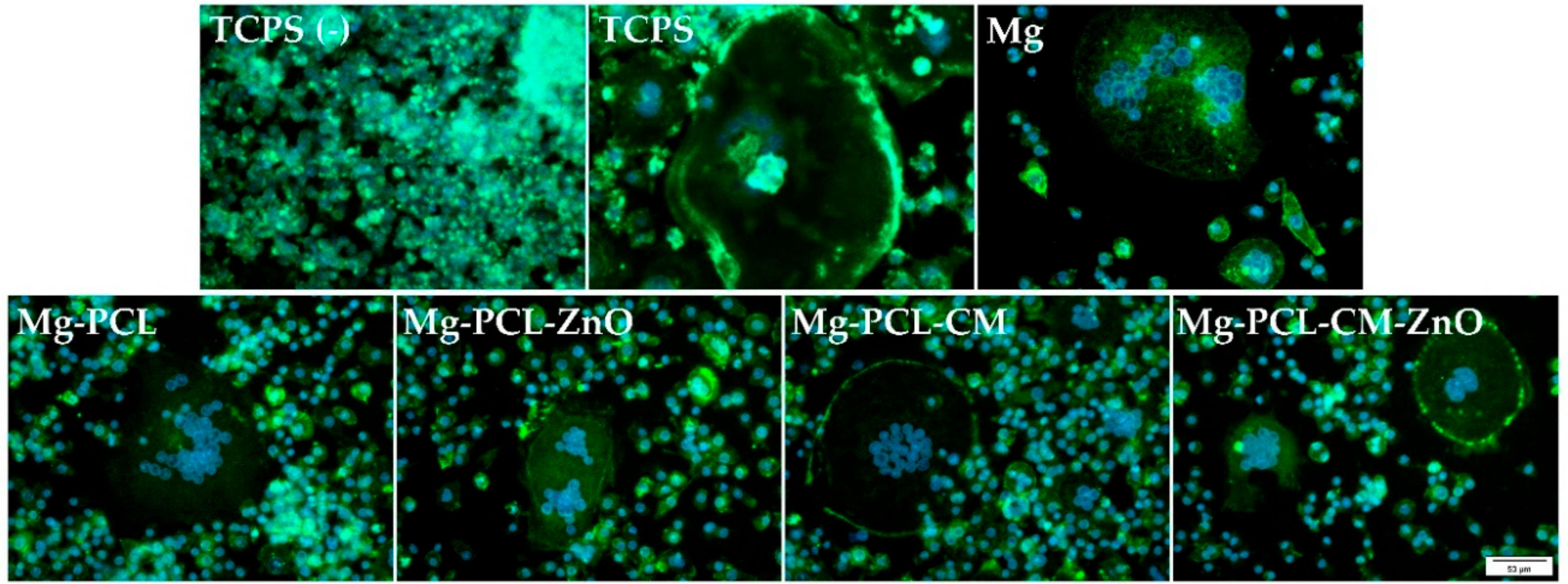
2.6. Multinuclear Foreign Body Giant Cell Formation
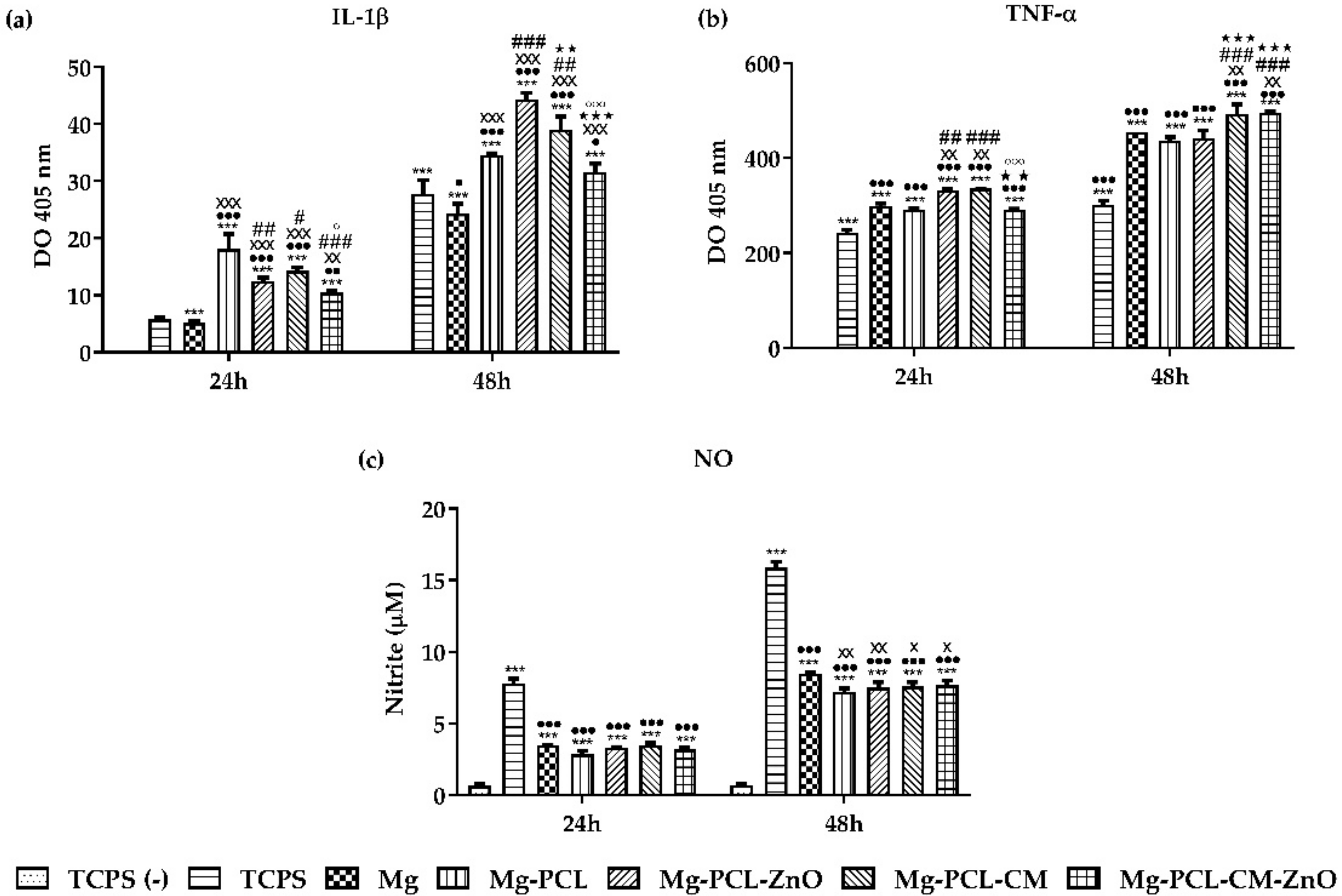
2.7. The Extracellular Release of the Pro-Inflammatory Mediators
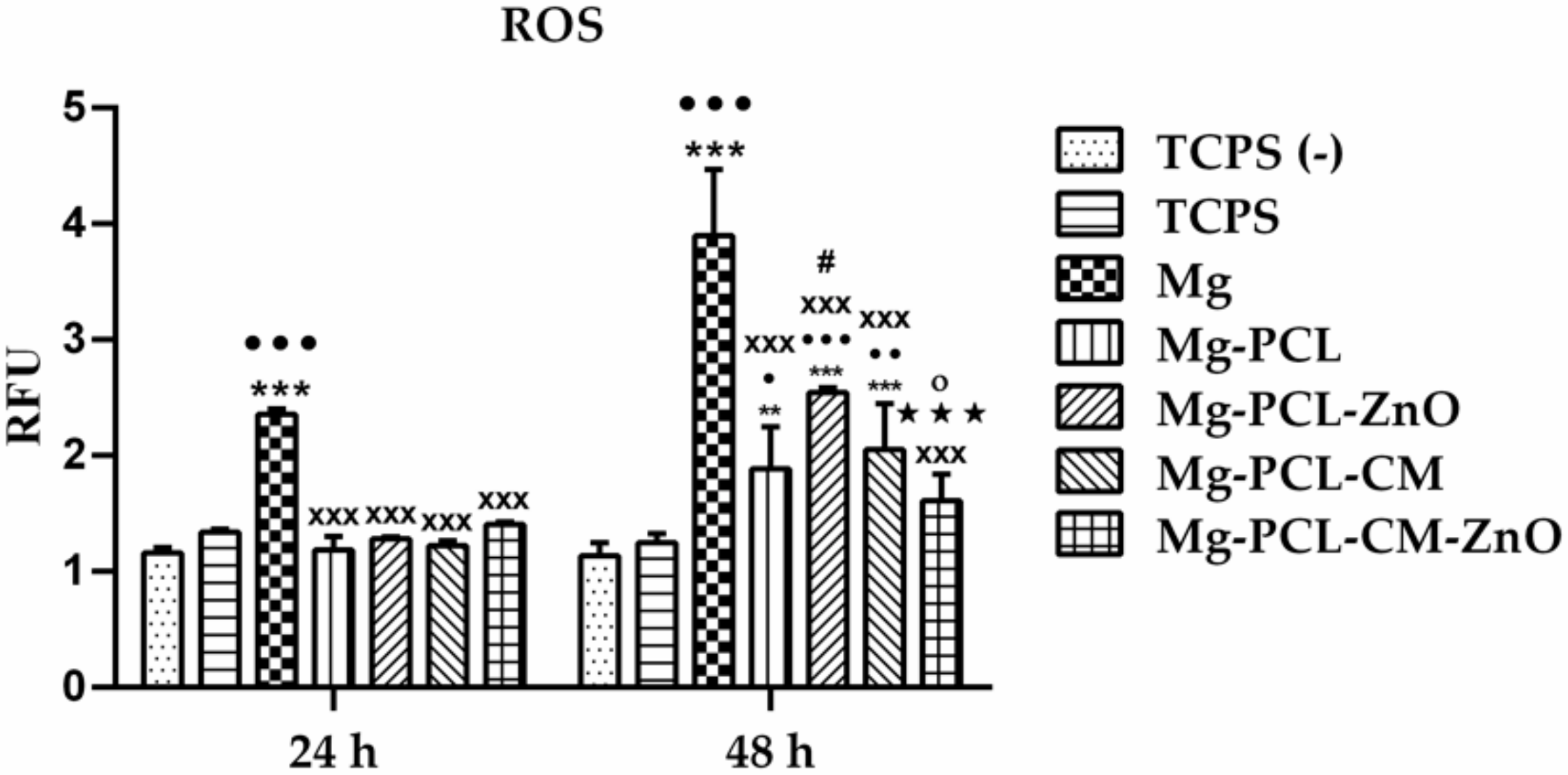
2.8. Intracellular Reactive Oxygen Species Generation
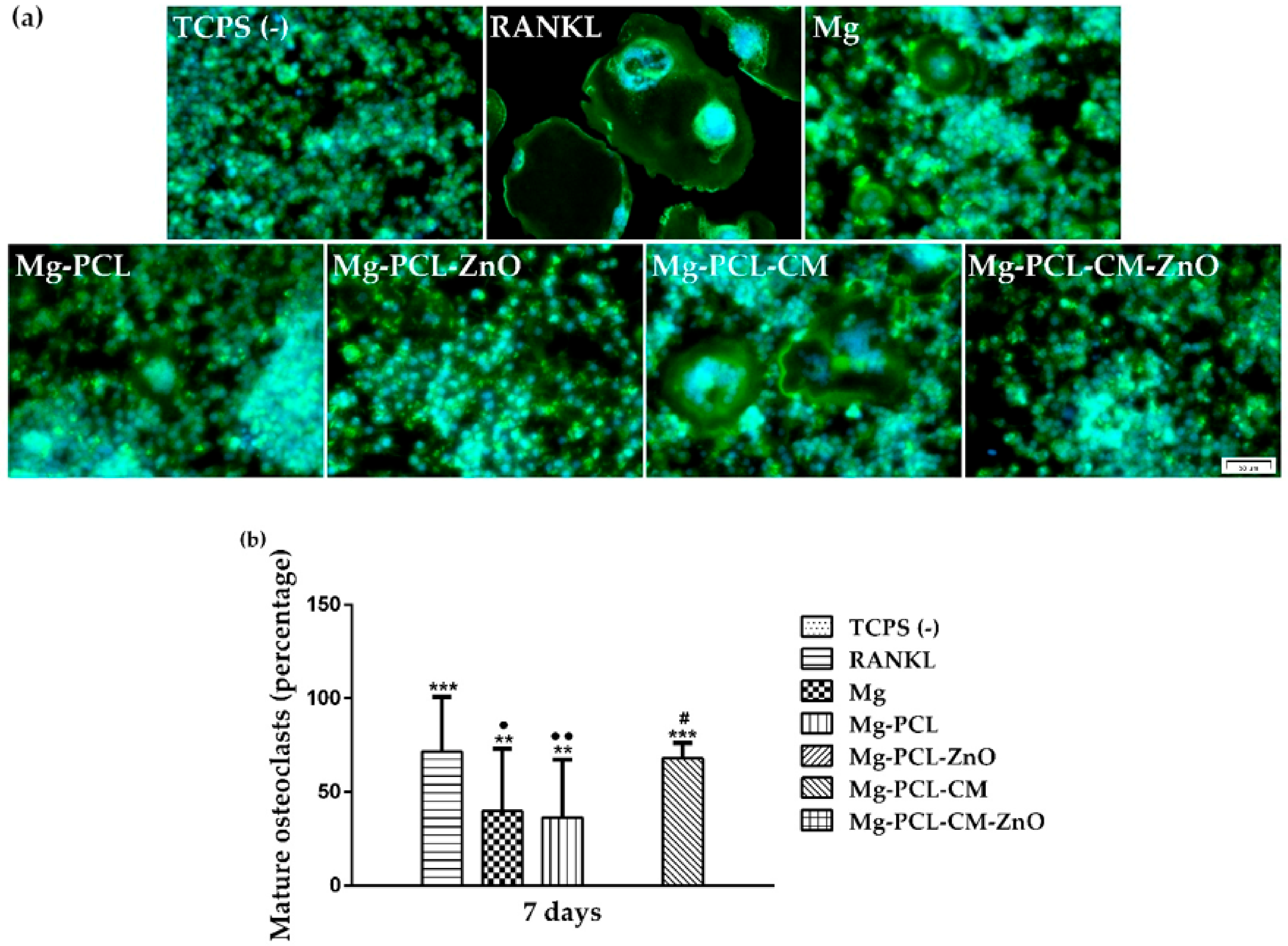
2.9. Macrophage-Osteoclast Differentiation
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Coating Precursor’s Preparation
4.3. Electrospinning Fabrication of the PCL-Coatings
4.4. Spectrometric Characterization of the Coated Alloys
4.5. In Vitro Release of Coumarin
4.6. Electrochemical Experiments
4.7. In Vitro Biological Experiments
4.7.1. Extract Preparation
4.7.2. Cell Culture
4.7.3. Cell Viability and Proliferation Assays
4.7.4. Cellular Morphology and Cytoskeleton Organization Assessment
4.7.5. In Vitro Macrophage Fusion Assay
4.7.6. Quantification of the Secreted Pro-Inflammatory Mediators
4.7.7. Intracellular ROS Level Determination
4.7.8. Osteoclast Differentiation Assay
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Persaud, S.D.; McGoron, A. Biodegradable Magnesium Alloys: A Review of Material Development and Applications. J. Biomim. Biomater. Tissue Eng. 2012, 12, 25–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Ren, Z.; Xu, Y.; Pang, S.; Zhao, X.; Zhao, Y. Biodegradable Magnesium Alloys Developed as Bone Repair Materials: A Review. Scanning 2018, 2018, 9216314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamrani, S.; Fleck, C. Biodegradable Magnesium Alloys as Temporary Orthopaedic Implants: A Review. BioMetals 2019, 32, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Sezer, N.; Evis, Z.; Kayhan, S.M.; Tahmasebifar, A.; Koç, M. Review of Magnesium-Based Biomaterials and Their Applications. J. Magnes. Alloys 2018, 6, 23–43. [Google Scholar] [CrossRef]
- Farraro, K.F.; Kim, K.E.; Woo, S.L.-Y.; Flowers, J.R.; McCullough, M.B. Revolutionizing Orthopaedic Biomaterials: The Potential of Biodegradable and Bioresorbable Magnesium-Based Materials for Functional Tissue Engineering. J. Biomech. 2014, 47, 1979–1986. [Google Scholar] [CrossRef] [Green Version]
- Witte, F.; Hort, N.; Feyerabend, F.; Vogt, C. Magnesium (Mg) Corrosion: A Challenging Concept for Degradable Implants. In Corrosion of Magnesium Alloys; Elsevier BV: Amsterdam, The Netherlands, 2011; pp. 403–425. [Google Scholar]
- Negrescu, A.M.; Necula, M.-G.; Costache, M.; Cimpean, A. In Vitro and in Vivo Biological Performance of Mg-Based Bone Implants. Rev. Biol. Biomed. Sci. 2020, 3, 11–41. [Google Scholar] [CrossRef]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign Body Reaction to Biomaterials. Semin. Immunol. 2008, 20, 86–100. [Google Scholar] [CrossRef] [Green Version]
- Franz, S.; Rammelt, S.; Scharnweber, D.; Simon, J.C. Immune Responses to Implants—A Review of the Implications for the Design of Immunomodulatory Biomaterials. Biomaterials 2011, 32, 6692–6709. [Google Scholar] [CrossRef]
- Takayanagi, H. Inflammatory Bone Destruction and Osteoimmunology. J. Periodontal Res. 2005, 40, 287–293. [Google Scholar] [CrossRef]
- Chen, Z.; Wu, C.; Gu, W.; Klein, T.J.; Crawford, R.W.; Xiao, Y. Osteogenic Differentiation of Bone Marrow MSCs by β-Tricalcium Phosphate Stimulating Macrophages via BMP2 Signalling Pathway. Biomaterials 2014, 35, 1507–1518. [Google Scholar] [CrossRef]
- Brown, B.N.; Ratner, B.D.; Goodman, S.B.; Amar, S.; Badylak, S.F. Macrophage Polarization: An Opportunity for Improved Outcomes in Biomaterials and Regenerative Medicine. Biomaterials 2012, 33, 3792–3802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, F.O.; Sica, A.; Mantovani, A.; Locati, M. Macrophage Activation and Polarization. Front. Biosci. 2008, 13, 453–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadowska, J.M.; Wei, F.; Guo, J.; Guillem-Marti, J.; Lin, Z.; Ginebra, M.-P.; Xiao, Y. The Effect of Biomimetic Calcium Deficient Hydroxyapatite and Sintered β-Tricalcium Phosphate on Osteoim-Mune Reaction and Osteogenesis. Acta Biomater. 2019, 96, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Schlundt, C.; Schell, H.; Goodman, S.B.; Vunjak-Novakovic, G.; Duda, G.N.; Schmidt-Bleek, K. Immune Modulation as a Therapeutic Strategy in Bone Regeneration. J. Exp. Orthop. 2015, 2, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, M.; Keshav, S.; Harris, N.; Gordon, S. Interleukin 4 Potently Enhances Murine Macrophage Mannose Receptor Activity: A Marker of Alternative Immunologic Macrophage Activation. J. Exp. Med. 1992, 176, 287–292. [Google Scholar] [CrossRef]
- Koh, T.J.; DiPietro, L.A. Inflammation and Wound Healing: The Role of the Macrophage. Expert Rev. Mol. Med. 2011, 13, e23. [Google Scholar] [CrossRef] [Green Version]
- Recalcati, S.; Locati, M.; Marini, A.; Santambrogio, P.; Zaninotto, F.; De Pizzol, M.; Zammataro, L.; Girelli, D.; Cairo, G. Differential Regulation of Iron Homeostasis during Human Macrophage Polarized Activation. Eur. J. Immunol. 2010, 40, 824–835. [Google Scholar] [CrossRef]
- Mantovani, A.; Biswas, S.K.; Galdiero, M.R.; Sica, A.; Locati, M. Macrophage Plasticity and Polarization in Tissue Repair and Remodelling. J. Pathol. 2013, 229, 176–185. [Google Scholar] [CrossRef]
- Ho, V.W.; Sly, L.M. Derivation and Characterization of Murine Alternatively Activated (M2) Macrophages. Comput. Biol. 2009, 531, 173–185. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The Chemokine System in Diverse Forms of Macrophage Activation and Polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Corliss, B.A.; Azimi, M.S.; Munson, J.M.; Poirce, S.M.; Murfee, W.L. Macrophages: An Inflammatory Link Between Angiogenesis and Lymphogiogenesis. Microcirculation 2016, 23, 95–121. [Google Scholar] [CrossRef]
- Krzyszczyk, P.; Schloss, R.; Palmer, A.; Berthiaume, F. The Role of Macrophages in Acute and Chronic Wound Healing and Interventions to Promote Pro-wound Healing Phenotypes. Front. Physiol. 2018, 9, 419. [Google Scholar] [CrossRef]
- Gordon, S. Alternative Activation of Macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Ogle, M.E.; Segar, C.E.; Sridhar, S.; Botchwey, E.A. Monocytes and Macrophages in Tissue Repair: Implications for Immunoregenerative Biomaterial Design. Exp. Biol. Med. 2016, 241, 1084–1097. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Helming, L.; Gordon, S. Alternative Activation of Macrophages: An Immunologic Functional Perspective. Annu. Rev. Immunol. 2009, 27, 451–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, S.; Martinez, F.O. Alternative Activation of Macrophages: Mechanism and Functions. Immunology 2010, 32, 593–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosser, D.M.; Edwards, J.P. Exploring the Full Spectrum of Macrophage Activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K.; Mantovani, A. Macrophage Plasticity and Interaction with Lymphocyte Subsets: Cancer as a Paradigm. Nat. Immunol. 2010, 11, 889–896. [Google Scholar] [CrossRef]
- Mills, C.D.; Thomas, A.C.; Lenz, L.L.; Munder, M. Macrophage: SHIP of Immunity. Front. Immunol. 2014, 5, 620. [Google Scholar] [CrossRef] [Green Version]
- Sridharan, R.; Cameron, A.R.; Kelly, D.J.; Kearney, C.J.; O’Brien, F.J. Biomaterial Based Modulation of Macrophage Polarization: A Review and Suggested Design Principles. Mater. Today 2015, 18, 313–325. [Google Scholar] [CrossRef]
- Chen, Z.; Klein, T.; Murray, R.Z.; Crawford, R.; Chang, J.; Wu, C.; Xiao, Y. Osteoimmunomodulation for the Development of Advanced Bone Biomaterials. Mater. Today 2016, 19, 304–321. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Mao, X.; Tan, L.; Friis, T.; Wu, C.; Crawford, R.; Xiao, Y. Osteoimmunomodulatory Properties of Magnesium Scaffolds Coated with β-Tricalcium Phosphate. Biomaterials 2014, 35, 8553–8565. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Zhang, D.; Li, M.; Liu, X.; Hang, Y.; Qian, S. Osteogenesis, Angiogenesis and Immune Response of Mg-Al Layered Double Hydroxide Coating on Pure Mg. Bioact. Mater. 2021, 6, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Gao, P.; Zhang, H.; Guo, Z.; Zheng, Y.; Han, Y. Osteoimmunomodulation, Osseointegration, and in Vivo Mechanical in-Tegrity of Pure MG Coated with Ha Na-Norod/Pore-Sealed MgO Bilayer. Biomater. Sci. 2018, 6, 3202–3218. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wang, B.; Jia, Z.; Lu, X.; Fang, L.; Wang, K.; Ren, F. Polydopamine Mediated Assembly of Hydroxyapatite Nanoparticles and Bone Morphogenetic Protein-2 on Magnesium Alloys for Enhanced Corrosion Resistance and Bone Regeneration. J. Biomed. Mater. Res. Part A 2017, 105, 2750–2761. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Li, H.; Wang, D.; Tian, P.; Tian, Y.; Yuan, G.; Xu, D.; Liu, X. Enhanced Corrosion Resistance and Biocompatibility of Magnesium Alloy by Mg–Al-Layered Double Hydroxide. ACS Appl. Mater. Interfaces 2016, 8, 35033–35044. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhu, Y.; Meng, L.; Chen, P.; Shi, T.; Liu, X.; Zheng, Y. Electrophoretic Deposition of Colloidal Particles on MG with Cytocompatibility, Antibacterial Performance, and Corrosion Resistance. Acta Biomater. 2016, 45, 387–398. [Google Scholar] [CrossRef]
- Augustine, R.; Malik, H.N.; Singhal, D.K.; Mukherjee, A.; Malakar, D.; Kalarikkal, N.; Thomas, S. Electrospun Polycaprolactone/ZnO Nanocomposite Membranes as Biomaterials with Antibacterial and Cell Adhesion Properties. J. Polym. Res. 2014, 21, 1–17. [Google Scholar] [CrossRef]
- Bui, H.T.; Chung, O.H.; Cruz, J.D.; Park, J.S. Fabrication and Characterization of Electrospun Curcumin-Loaded Polycaprolactone-Polyethylene Glycol Nano-Fibers for Enhanced Wound Healing. Macromol. Res. 2014, 22, 1288–1296. [Google Scholar] [CrossRef]
- Lee, J.S.; Kim, J.K.; Chang, Y.H.; Park, S.R. Preparation of Collagen/Poly (L-Lactic Acid) Composite Material for Wound Dressing. Macromol. Res. 2007, 5, 205–210. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Ghosh, C.; Hwang, S.-G.; Tran, L.D.; Park, J.S. Characteristics of Curcumin-Loaded Poly (Lactic Acid) Nanofibers for Wound Healing. J. Mater. Sci. 2013, 48, 7125–7133. [Google Scholar] [CrossRef]
- Pant, H.R.; Neupane, M.P.; Pant, B.; Panthi, G.; Oh, H.-J.; Lee, M.H.; Kim, H.Y. Fabrication of Highly Porous Poly (ɛ-Caprolactone) Fibers for Novel Tissue Scaffold via Water-Bath Electrospinning. Colloids Surf. B Biointerfaces 2011, 88, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Merrell, J.G.; McLaughlin, S.W.; Tie, L.; Laurencin, C.T.; Chen, A.F.; Nair, L.S. Curcumin-Loaded Poly(ε-Caprolactone) Nanofibres: Diabetic Wound Dressing with Anti-oxidant and Anti-Inflammatory Properties. Clin. Exp. Pharmacol. Physiol. 2009, 36, 1149–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumbar, S.G.; Nukavarapu, S.P.; James, R.; Nair, L.S.; Laurencin, C.T. Electrospun Poly (Lactic Acid-Co-glycolic Acid) Scaffolds for Skin Tissue Engineering. Biomaterials 2008, 29, 4100–4107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanas, T.; Kumar, T.S.; Perumal, G.; Doble, M. Tailoring Degradation of AZ31 Alloy by Surface Pre-treatment and Electrospun PCL Fibrous Coating. Mater. Sci. Eng. C 2016, 65, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Rezk, A.; Mousa, H.M.; Lee, J.; Park, C.H.; Kim, C.S. Composite PCL/HA/Simvastatin Electrospun Nanofiber Coating on Biodegradable MG Alloy for Orthopedic Implant Application. J. Coat. Technol. Res. 2019, 16, 477–489. [Google Scholar] [CrossRef]
- Shitole, A.A.; Raut, P.W.; Sharma, N.; Giram, P.; Khandwekar, A.P.; Garnaik, B. Electrospun Polycaprolactone/Hydroxyapatite/ZnO Nanofibers as Potential Biomaterials for Bone Tissue Regeneration. J. Mater. Sci. Mater. Med. 2019, 30, 51. [Google Scholar] [CrossRef]
- Barot, K.P.; Jain, S.V.; Kremer, L.; Singh, S.; Ghate, M.D. Recent Advances and Therapeutic Journey of Coumarins: Current Status and Perspectives. Med. Chem. Res. 2015, 24, 2771–2798. [Google Scholar] [CrossRef]
- Srikumar, S.P.R.K. Therapeutic Role of Coumarins and Coumarin-Related Compounds. J. Fertil. Pestic. 2014, 5, 1000130. [Google Scholar] [CrossRef]
- Münchow, E.A.; Albuquerque, M.T.P.; Zero, B.; Kamocki, K.; Piva, E.; Gregory, R.L.; Bottino, M.C. Development and Characterization of Novel ZnO-Loaded Electrospun Membranes for Periodontal Regeneration. Dent. Mater. 2015, 31, 1038–1051. [Google Scholar] [CrossRef] [Green Version]
- Umashankar, T.; Govindappa, M.; Yarappa Lakshmikantha, R.; Padmalatha Rai, S.; Channabasava, R. Isolation and Characterization of Coumarin Isolated from Endophyte, Alternaria Species-1 of Crotalaria pallida and Its Apoptotic Action on HeLa Cancer Cell Line. J. Postgenomics Drug Biomark. Dev. 2015, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Wasserbauer, J.; Buchtík, M.; Tkacz, J.; Fintová, S.; Minda, J.; Doskočil, L. Improvement of AZ91 Alloy Corrosion Properties by Duplex NI-P Coating Deposition. Materials 2020, 13, 1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vadiveloo, P.K.; Keramidaris, E.A.; Morrison, W.; Stewart, A.G. Lipopolysaccharide-Induced Cell Cycle Arrest in Macrophages Occurs Independently of Nitric Oxide Synthase II Induction. Biochim. et Biophys. Acta (BBA)—Bioenerg. 2001, 1539, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Sheikh, Z.; Brooks, P.J.; Barzilay, O.; Fine, N.; Glogauer, M. Macrophages, Foreign Body Giant Cells and Their Response to Implantable Biomaterials. Materials 2015, 8, 5671–5701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padmanabhan, J.; Kyriakides, T.R. Nanomaterials, Inflammation, and Tissue Engineering. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2015, 7, 355–370. [Google Scholar] [CrossRef] [Green Version]
- Ratner, B.D. Reducing Capsular Thickness and Enhancing Angiogenesis around Implant Drug Release Systems. J. Control. Release 2002, 78, 211–218. [Google Scholar] [CrossRef]
- McNally, A.K.; Anderson, J.M. Macrophage Fusion and Multinucleated Giant Cells of Inflammation. Cannabinoids Neuropsychiatr. Disord. 2011, 713, 97–111. [Google Scholar] [CrossRef]
- Teitelbaum, S.L. Osteoclasts: What Do They Do and How Do They Do It? Am. J. Pathol. 2007, 170, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Xu, Z.; Smith, C.; Sankar, J. Recent Advances in the Development of Magnesium Alloys for Biodegradable Implants. Acta Biomater. 2014, 10, 4561–4573. [Google Scholar] [CrossRef]
- Brar, H.S.; Platt, M.O.; Sarntinoranont, M.; Martin, P.I.; Manuel, M.V. Magnesium as a Biodegradable and Bioabsorbable Material for Medical Implants. JOM 2009, 61, 31–34. [Google Scholar] [CrossRef]
- Wang, J.; Qian, S.; Liu, X.; Xu, L.; Miao, X.; Xu, Z.; Cao, L.; Wang, H.; Jiang, X. M2 Macrophages Contribute to Osteogenesis and Angiogenesis on Nanotubular TiO2 Surfaces. J. Mater. Chem. B 2017, 5, 3364–3376. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Du, Z.; Du, J.; Yao, W.; Zhang, J.; Weng, Z.; Liu, S.; Zhao, Y.; Liu, Y.; Zhang, X.; et al. A Multifaceted Coating on Titanium Dictates Osteoimmunomodulation and Osteo/Angio-Genesis Towards Ameliorative Osseointegration. Biomaterials 2018, 162, 154–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schutte, R.J.; Parisi-Amon, A.; Reichert, W.M. Cytokine Profiling Using Monocytes/Macrophages Cultured on Common Biomaterials with a Range of Surface Chemistries. J. Biomed. Mater. Res. Part A 2008, 88, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Bachhuka, A.; Han, S.; Wei, F.; Lu, S.; Visalakshan, R.M.; Vasilev, K.; Xiao, Y. Tuning Chemistry and Topography of Nanoengineered Surfaces to Manipulate Immune Response for Bone Regeneration Applications. ACS Nano 2017, 11, 4494–4506. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Jones, J.A.; Xu, Y.; Low, H.Y.; Anderson, J.M.; Leong, K.W. Characterization of Topographical Effects on Macrophage Behaviour in a Foreign Body Response Model. Biomaterials 2010, 31, 3479–3491. [Google Scholar] [CrossRef]
- Bota, P.C.S.; Collie, A.M.B.; Puolakkainen, P.; Vernon, R.B.; Sage, E.H.; Ratner, B.D.; Stayton, P.S. Biomaterial Topography Alters Healing in Vivo and Monocyte/Macrophage Activation In Vitro. J. Biomed. Mater. Res. Part A 2010, 95, 649–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wójciak-Stothard, B.; Curtis, A.; Monaghan, W.; Macdonald, K.; Wilkinson, C. Guidance and Activation of Murine Macrophages by Nanometric Scale Topography. Exp. Cell Res. 1996, 223, 426–435. [Google Scholar] [CrossRef]
- Brodbeck, W.G.; Nakayama, Y.; Matsuda, T.; Colton, E.; Ziats, N.; Anderson, J. Biomaterial Surface Chemistry Dictates Adherent Monocyte/Macrophage Cytokine Expression in Vitro. Cytokine 2002, 18, 311–319. [Google Scholar] [CrossRef]
- Godek, M.L.; Sampson, J.A.; Duchsherer, N.L.; McElwee, Q.; Grainger, D.W. Rho GTPase Protein Expression and Activation in Murine Monocytes/Macrophages Are Not Modulated by Model Biomaterial Surfaces in Serum-Containing in Vitro Cultures. J. Biomater. Sci. Polym. Ed. 2006, 17, 1141–1158. [Google Scholar] [CrossRef] [Green Version]
- Sussman, E.M.; Halpin, M.C.; Muster, J.; Moon, R.T.; Ratner, B.D. Porous Implants Modulate Healing and Induce Shifts in Local Macrophage Polarization in the Foreign Body Reaction. Ann. Biomed. Eng. 2014, 42, 1508–1516. [Google Scholar] [CrossRef]
- Dash, T.K.; Konkimalla, V.B. Poly-ε-Caprolactone Based Formulations for Drug Delivery and Tissue Engineering: A Review. J. Control. Release 2012, 158, 15–33. [Google Scholar] [CrossRef] [PubMed]
- Vandrovcova, M.; Douglas, T.E.; Mróz, W.; Musial, O.; Schaubroeck, D.; Budner, B.; Syroka, R.; Dubruel, P.; Bacakova, L. Pulsed Laser Deposition of Magnesium-Doped Calcium Phosphate Coatings on Porous Polycaprolactone Scaffolds Produced by Rapid Prototyping. Mater. Lett. 2015, 148, 178–183. [Google Scholar] [CrossRef]
- Li, L.-Y.; Cui, L.-Y.; Zeng, R.-C.; Li, S.-Q.; Chen, X.-B.; Zheng, Y.; Kannan, M.B. Advances in Functionalized Polymer Coatings on Biodegradable Magnesium Alloys—A Review. Acta Biomater. 2018, 79, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.; Luan, B. Protective Coatings on Magnesium and Its Alloys—A Critical Review. J. Alloys Compd. 2002, 336, 88–113. [Google Scholar] [CrossRef]
- Xu, L.; Yamamoto, A. Characteristics and Cytocompatibility of Biodegradable Polymer Film on Magnesium by Spin Coating. Colloids Surf. B Biointerfaces 2012, 93, 67–74. [Google Scholar] [CrossRef]
- Hanas, T.; Kumar, T.S.; Perumal, G.; Doble, M.; Ramakrishna, S. Electrospun PCL/HA Coated Friction Stir Processed AZ31/Ha Composites for Degradable Implant Applications. J. Mater. Process. Technol. 2018, 252, 398–406. [Google Scholar] [CrossRef]
- Shin, H.; Jo, S.; Mikos, A.G. Biomimetic Materials for Tissue Engineering. Biomaterials 2003, 24, 4353–4364. [Google Scholar] [CrossRef]
- Fylaktakidou, K.C.; Hadjipavlou-Litina, D.E.; Litinas, K.; Nicolaides, D.N. Natural and Synthetic Coumarin Derivatives with Anti-Inflammatory/Antioxidant Activities. Curr. Pharm. Des. 2004, 10, 3813–3833. [Google Scholar] [CrossRef]
- Sandhiutami, N.M.D.; Moordiani, M.; Laksmitawati, D.R.; Fauziah, N.; Maesaroh, M.; Widowati, W. In Vitro Assesment of Anti-inflammatory Activities of Coumarin and Indonesian Cassia Extract in RAW264.7 Murine Macrophage Cell Line. Iran. J. Basic Med. Sci. 2017, 20, 99–106. [Google Scholar]
- Chandel, P.; Kumar, A.; Singla, N.; Kumar, A.; Singh, G.; Gill, R.K. Rationally Synthesized Coumarin Based Pyrazolines Ameliorate Carrageenan Induced Inflammation through COX-2/Pro-inflammatory Cytokine Inhibition. MedChemComm 2019, 10, 421–430. [Google Scholar] [CrossRef]
- Emmanuel-Giota, A.A.; Fylaktakidou, K.C.; Litinas, K.E.; Nicolaides, D.N.; Hadjipavlou-Litina, D.J. Synthesis and Biological Evaluation of Several 3-(Coumarin-4-Yl)Tetrahydroisoxazole and 3-(Coumarin-4-Yl)Dihydropyrazole Derivatives. J. Heterocycl. Chem. 2001, 38, 717–722. [Google Scholar] [CrossRef]
- Akhter, M.; Akhter, N.; Alam, M.M.; Zaman, M.S.; Saha, R.; Kumar, A. Synthesis and Biological Evaluation of 2,5-Disubstituted 1,3,4-Oxadiazole Derivatives with Both COX and LOX Inhibitory Activity. J. Enzym. Inhib. Med. Chem. 2011, 26, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Grover, J.; Kumar, V.; Sobhia, M.E.; Jachak, S.M. Synthesis, Biological Evaluation and Docking Analysis of 3-Methyl-1-Phenylchromeno[4,3-C]Pyrazol-4(1H)-Ones as Potential Cyclooxygenase-2 (COX-2) Inhibitors. Bioorganic Med. Chem. Lett. 2014, 24, 4638–4642. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-H. Biological Effects of Zinc Oxide Nanoparticles on Inflammation. TANG Humanit. Med. 2016, 6, 23–24. [Google Scholar] [CrossRef] [Green Version]
- Augustine, R.; Dominic, E.A.; Reju, I.; Kaimal, B.; Kalarikkal, N.; Thomas, S. Investigation of Angiogenesis and Its Mechanism Using Zinc Oxide Nanoparticle-Loaded Electrospun Tissue Engineering Scaffolds. RSC Adv. 2014, 4, 51528–51536. [Google Scholar] [CrossRef]
- Park, K.H.; Park, B.; Yoon, D.S.; Kwon, S.-H.; Shin, D.M.; Li, J.W.; Lee, H.G.; Shim, J.-H.; Park, J.H.; Lee, J.M. Zinc Inhibits Osteoclast Differentiation by Suppression of Ca2+-Calcineurin-NFATc1 Signaling Pathway. Cell. Commun. Signal. 2013, 11, 74. [Google Scholar] [CrossRef] [Green Version]
- Reyes-López, S.Y.; Cornejo-Monroy, D.; González-García, G. A Novel Route for the Preparation of Gold Nanoparticles in Polycaprolactone Nanofibers. J. Nanomater. 2015, 2015, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Mousa, H.M.; Park, C.H.; Kim, C.S. Enhanced Corrosion Resistance and Biocompatibility of AZ31 Mg Alloy Using PCL/ZnO NPs via Electrospinning. Appl. Surf. Sci. 2017, 396, 249–258. [Google Scholar] [CrossRef]
- Wu, Y.-J.; Chen, X.-B.; Williams, G.; Scully, J.R.; Gengenbach, T.; Birbilis, N. Stifling Magnesium Corrosion via a Novel Anodic Coating. RSC Adv. 2016, 6, 43408–43417. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.-J.; Gao, L.; Liu, J.-C.; Jiang, R.-Z.; Sun, F.-Y.; Cui, L.-Y.; Zeng, R.-C.; Zhi, K.-Q.; Zeng, R.-C.; Wang, Z.-L. Corrosion Resistance and Antibacterial Activity of Hydroxyapatite Coating Induced by Ciprofloxacin-Loaded Polymeric Multilayers on Magnesium Alloy. Prog. Org. Coat. 2019, 135, 465–474. [Google Scholar] [CrossRef]
- Ji, X.-J.; Gao, L.; Liu, J.-C.; Wang, J.; Cheng, Q.; Li, J.-P.; Zeng, R.-C.; Zhi, K.-Q.; Zeng, R.-C.; Wang, Z.-L. Corrosion Resistance and Antibacterial Properties of Hydroxyapatite Coating Induced by Gentamicin-Loaded Polymeric Multilayers on Magnesium Alloys. Colloids Surf. B Biointerfaces 2019, 179, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Sebaa, M.A.; Dhillon, S.; Liu, H. Electrochemical Deposition and Evaluation of Electrically Conductive Polymer Coating on Biodegradable Magnesium Implants for Neural Applications. J. Mater. Sci. Mater. Electron. 2013, 24, 307–316. [Google Scholar] [CrossRef]
- Jin, T.; Xie, Z.; Fullston, D.; Huang, C.; Zeng, R.; Bai, R. Corrosion Resistance of Copolymerization of Acrylamide and Acrylic Acid Grafted Graphene Oxide Composite Coating on Magnesium Alloy. Prog. Org. Coat. 2019, 136, 105222. [Google Scholar] [CrossRef]
- Feyerabend, F.; Feyerabend, F.; Maier, P.; Fischer, J.; Störmer, M.; Blawert, C.; Dietzel, W.; Hort, N. Biodegradable Magnesium–Hydroxyapatite Metal Matrix Composites. Biomaterials 2007, 28, 2163–2174. [Google Scholar] [CrossRef] [Green Version]
- Wallin, R.F. A Practical Guide to ISO 10993-12: Sample Preparation and Reference Materials; MDDI: Los Angeles, CA, USA, 1998. [Google Scholar]
- Fischer, J.; Pröfrock, D.; Hort, N.; Willumeit, R.; Feyerabend, F. Improved Cytotoxicity Testing of Magnesium Materials. Mater. Sci. Eng. B 2011, 176, 830–834. [Google Scholar] [CrossRef] [Green Version]
- Neacsu, P.; Mazare, A.; Cimpean, A.; Park, J.; Costache, M.; Schmuki, P.; Demetrescu, I. Reduced Inflammatory Activity of RAW 264.7 Macrophages on Titania Nanotube Modified TI Surface. Int. J. Biochem. Cell Biol. 2014, 55, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, X.; Xu, M.; Yang, F.; Wang, W.; Niu, X. In Vitro Immunomodulation of Magnesium on Monocytic Cell toward Anti-Inflammatory Macrophages. Regen. Biomater. 2020, 7, 391–401. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Wu, J.; Yuan, G.; Chen, T.-X. In Vitro Study of the Inflammatory Cells Response to Biodegradable Mg-Based Alloy Extract. PLoS ONE 2018, 13, e0193276. [Google Scholar] [CrossRef]
- Jin, L.; Chen, C.; Li, Y.; Yuan, F.; Gong, R.; Wu, J.; Zhang, H.; Kang, B.; Yuan, G.; Zeng, H.; et al. A Biodegradable Mg-Based Alloy Inhibited the Inflammatory Response of THP-1 Cell-Derived Macrophages Through the TRPM7–PI3K–AKT1 Signaling Axis. Front. Immunol. 2019, 10, 2798. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.M. Future Challenges in the in Vitro and in Vivo evaluation of Biomaterial Biocompatibility. Regen. Biomater. 2016, 3, 73–77. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-S.; Stachelek, S.J.; Tomczyk, N.; Finley, M.J.; Composto, R.J.; Eckmann, D.M. Correlating Macrophage Morphology and Cytokine Production Resulting from Biomaterial Contact. J. Biomed. Mater. Res. Part A 2012, 101, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Refai, A.K.; Textor, M.; Brunette, D.M.; Waterfield, J.D. Effect of Titanium Surface Topography on Macrophage Activation and Secretion of Proinflammatory Cytokines and Chemokines. J. Biomed. Mater. Res. 2004, 70, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.; Zhang, Y.; Yang, Q.; Cheng, S.; Hao, J.; Zhao, X.; Jiang, Z. Genistein Suppresses LPS-Induced Inflammatory Response through Inhibiting NF-κB following AMP Kinase Activation in RAW 264.7 Macrophages. PLoS ONE 2012, 7, e53101. [Google Scholar] [CrossRef] [PubMed]
- Lin, N.; Hu, H.; Sung, L.; Lin-Gibson, S. Quantification of Cell Response to Polymeric Composites Using a Two-Dimensional Gradient Platform. Comb. Chem. High Throughput Screen. 2009, 12, 619–625. [Google Scholar] [CrossRef]
- Berghaus, L.J.; Moore, J.N.; Hurley, D.J.; Vandenplas, M.L.; Fortes, B.P.; Wolfert, M.A.; Boons, G.-J. Innate Immune Responses of Primary Murine Macrophage-Lineage Cells and RAW 264.7 Cells to Ligands of Toll-Like Receptors 2, 3, and 4. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Karlan, D.S.; Zinman, J. Microcredit in Theory and Practice: Using Randomized Credit Scoring for Impact Evaluation. Science 2011, 332, 1278–1284. [Google Scholar] [CrossRef] [Green Version]
- McWhorter, F.Y.; Wang, T.; Nguyen, P.; Chung, T.; Liu, W.F. Modulation of Macrophage Phenotype by Cell Shape. Proc. Natl. Acad. Sci. USA 2013, 110, 17253–17258. [Google Scholar] [CrossRef] [Green Version]
- Somasundaram, V.; Gilmore, A.C.; Basudhar, D.; Palmieri, E.M.; Scheiblin, D.A.; Heinz, W.F.; Cheng, R.; Ridnour, L.A.; Altan-Bonnet, G.; Lockett, S.J.; et al. Inducible Nitric Oxide Synthase-Derived Extracellular Nitric Oxide Flux Regulates Proinflammatory Responses at the Single Cell Level. Redox Biol. 2020, 28, 101354. [Google Scholar] [CrossRef]
- Anderson, J.M. Multinucleated Giant Cells. Curr. Opin. Hematol. 2000, 7, 40–47. [Google Scholar] [CrossRef]
- Anderson, J.M. Biological Responses to Materials. Annu. Rev. Mater. Res. 2001, 31, 81–110. [Google Scholar] [CrossRef]
- Luttikhuizen, D.T.; Harmsen, M.C.; Van Luyn, M.J. Cellular and Molecular Dynamics in the Foreign Body Reaction. Tissue Eng. 2006, 12, 1955–1970. [Google Scholar] [CrossRef] [PubMed]
- Gretzer, C.; Emanuelsson, L.; Liljensten, E.; Thomsen, P. The Inflammatory Cell Influx and Cytokines Changes during Transition from Acute Inflammation to Fibrous Repair around Implanted Materials. J. Biomater. Sci. Polym. Ed. 2006, 17, 669–687. [Google Scholar] [CrossRef] [PubMed]
- Jenney, C.R.; Anderson, J.M. Adsorbed Serum Proteins Responsible for Surface Dependent Human Macrophage Behaviour. J. Biomed. Mater. Res. 2000, 49, 435–447. [Google Scholar] [CrossRef]
- Brodbeck, W.G.; Colton, E.; Anderson, J.M. Effects of Adsorbed Heat Labile Serum Proteins and Fibrinogen on Adhesion and Apoptosis of Monocytes/Macrophages on Biomaterials. J. Mater. Sci. Mater. Electron. 2003, 14, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Mariani, E.; Lisignoli, G.; Borzì, R.M.; Pulsatelli, L. Biomaterials: Foreign Bodies or Tuners for the Immune Response? Int. J. Mol. Sci. 2019, 20, 636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, C.J.; Clegg, R.E.; Leavesley, D.I.; Pearcy, M.J. Mediation of Biomaterial–Cell Interactions by Adsorbed Proteins: A Review. Tissue Eng. 2005, 11, 1–18. [Google Scholar] [CrossRef]
- Xia, Z.; Triffitt, J. A Review on Macrophage Responses to Biomaterials. Biomed. Mater. 2006, 1, R1–R9. [Google Scholar] [CrossRef] [PubMed]
- Takebe, J.; Champagne, C.; Offenbacher, S.; Ishibashi, K.; Cooper, L. Titanium Surface Topography Alters Cell Shape and Modulates Bone Morphogenetic Protein 2 Expression in the j774a.1 Macrophage Cell Line. J. Biomed. Mater. Res. Part A 2003, 64, 207–216. [Google Scholar] [CrossRef]
- Labow, R.S.; Sa, D.; Matheson, L.A.; Santerre, J.P. Polycarbonate-Urethane Hard Segment Type Influences Esterase Substrate Specificity for Human-Macrophage-Mediated Biodegradation. J. Biomater. Sci. Polym. Ed. 2005, 16, 1167–1177. [Google Scholar] [CrossRef]
- Tamimi, F.; Sheikh, Z.; Barralet, J. Dicalcium Phosphate Cements: Brushite and Monetite. Acta Biomater. 2012, 8, 474–487. [Google Scholar] [CrossRef]
- Xia, Z.D.; Zhu, T.B.; Du, J.Y.; Zheng, Q.X.; Wang, L.; Li, S.-P.; Chang, C.Y.; Fang, S.Y. Macrophages in Degradation of Collagen/Hydroxylapatite(CHA), Beta-Tricalcium Phosphate Ceramics (TCP) Artificial Bone Graft. An In Vivo Study. Chin. Med. J. 1994, 107, 845–849. [Google Scholar] [PubMed]
- Kao, W.J. Evaluation of Protein-Modulated Macrophage Behavior on Biomaterials: Designing Biomimetic Materials for Cellular Engineering. Biomaterials 1999, 20, 2213–2221. [Google Scholar] [CrossRef]
- DeFife, K.M.; Jenney, C.R.; Colton, E.; Anderson, J.M. Cytoskeletal and Adhesive Structural Polarizations Accompany IL-13-Induced Human Macrophage Fusion. J. Histochem. Cytochem. 1999, 47, 65–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, W.; Reeve, J.L.; Liu, Y.; Teitelbaum, S.L.; Ross, F.P. DAP12 Couples c-Fms Activation to the Osteoclast Cytoskeleton by Recruitment of Syk. Mol. Cell 2008, 31, 422–431. [Google Scholar] [CrossRef] [Green Version]
- Helming, L.; Gordon, S. Molecular Mediators of Macrophage Fusion. Trends Cell Biol. 2009, 19, 514–522. [Google Scholar] [CrossRef]
- Vignery, A. Macrophage Fusion. J. Exp. Med. 2005, 202, 337–340. [Google Scholar] [CrossRef] [Green Version]
- Schachtner, H.; Calaminus, S.D.J.; Thomas, S.G.; Machesky, L.M. Podosomes in Adhesion, Migration, Mechanosensing and Matrix Remodeling. Cytoskeleton 2013, 70, 572–589. [Google Scholar] [CrossRef]
- Arjonen, A.; Kaukonen, R.; Ivaska, J. Filopodia and Adhesion in Cancer Cell Motility. Cell Adhes. Migr. 2011, 5, 421–430. [Google Scholar] [CrossRef] [Green Version]
- Jeremic, A.; Kelly, M.; Cho, J.A.; Cho, S.-J.; Hörber, J.; Jena, B.P. Calcium Drives Fusion of SNARE-Apposed Bilayers. Cell Biol. Int. 2004, 28, 19–31. [Google Scholar] [CrossRef]
- Chapekar, M.S. Tissue Engineering: Challenges and Opportunities. J. Biomed. Mater. Res. 2000, 53, 617–620. [Google Scholar] [CrossRef]
- Sirelkhatim, A.; Mahmud, S.; Seeni, A.; Kaus, N.H.M.; Ann, L.C.; Bakhori, S.K.M.; Hasan, H.; Mohamad, D. Review on Zinc Oxide Nanoparticles: Antibacterial Activity and Toxicity Mechanism. Nano-Micro Lett. 2015, 7, 219–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Pi, J.; Cai, J. The Advancing of Zinc Oxide Nanoparticles for Biomedical Applications. Bioinorg. Chem. Appl. 2018, 2018, 1062562. [Google Scholar] [CrossRef]
- Nagajyothi, P.C.; Cha, S.J.; Yanh, I.J.; Sreekanth, T.V.M.; Kim, K.J.; Shin, H.M. Antioxidant and Anti-Inflammatory Activities of Zinc Oxide Nanoparticles Synthesise Using Polygala Tenuifolia Root Extract. J. Photochem. Photobiol. B Biol. 2015, 146, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Ilves, M.; Palomäki, J.; Vippola, M.; Lehto, M.; Savolainen, K.; Savinko, T.; Alenius, H. Topically Applied ZnO Nanoparticles Suppress Allergen Induced Skin Inflammation but Induce Vigorous IgE Production in the Atopic Dermatitis Mouse Model. Part. Fibre Toxicol. 2014, 11, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.-F.; Jung, J.-Y.; Jung, Y.-J.; Choi, J.-H.; Jeong, W.-S.; Song, Y.-S.; Kang, J.-S.; Bi, K.; Kim, M.-J. Anti-Inflammatory Activities of Coumarins Isolated from Angelica gigas Nakai on LPS-stimulated RAW 264.7 Cells. Prev. Nutr. Food Sci. 2009, 14, 179–187. [Google Scholar] [CrossRef]
- Lee, E.-J.; Kang, M.-K.; Kim, Y.-H.; Kim, D.Y.; Oh, H.; Kim, S.-I.; Oh, S.Y.; Na, W.; Kang, Y.-H. Coumarin Ameliorates Impaired Bone Turnover by Inhibiting the Formation of Advanced Glycation End Products in Diabetic Osteoblasts and Osteoclasts. Biomolecules 2020, 10, 1052. [Google Scholar] [CrossRef]
- Tzeng, H.-E.; Huang, P.-H.; Tsai, C.-H.; Tsay, G.J.; Lee, Y.-J.; Huang, T.-J.; Lin, T.-H.; Chiu, Y.-M.; Wu, Y.-Y. Isosteviol Derivative Inhibits Osteoclast Differentiation and Ameliorates Ovariectomy-Induced Osteoporosis. Sci. Rep. 2018, 8, 11190. [Google Scholar] [CrossRef] [Green Version]
- Kohli, N.; Ho, S.; Brown, S.J.; Sawadkar, P.; Sharma, V.; Snow, M.; García-Gareta, E. Bone Remodelling In Vitro: Where are We Headed? Bone 2018, 110, 38–46. [Google Scholar] [CrossRef]
- Loi, F.; Córdova, L.A.; Pajarinen, J.; Lin, T.-H.; Yao, Z.; Goodman, S.B. Inflammation, Fracture and Bone Repair. Bone 2016, 86, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Kelava, T.; Sucur, A.; Kuzmac, S.; Katavic, V. Interactions between Bone and Immune Systems: A Focus on the Role of Inflammation in Bone Resorption and Fracture Healing. Period. Biol. 2014, 116, 45–52. [Google Scholar]
- Oryan, A.; Alidadi, S.; Moshiri, A. Current concerns regarding healing of bone defects. Hard Tissue 2013, 2, 1–12. [Google Scholar] [CrossRef]
- Einhorn, A.T.; Majeska, R.J.; Rush, E.B.; Levine, P.M.; Horowitz, M.C. The Expression of Cytokine Activity by Fracture Callus. J. Bone Miner. Res. 2009, 10, 1272–1281. [Google Scholar] [CrossRef]
- Rundle, C.H.; Wang, H.; Yu, H.; Chadwick, R.B.; Davis, E.I.; Wergedal, J.E.; Lau, K.-H.W.; Mohan, S.; Ryaby, J.T.; Baylink, D.J. Microarray Analysis of Gene Expression during the Inflammation and Endochondral Bone Formation Stages of Rat Femur Fracture Repair. Bone 2006, 38, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, M.; Rhee, C.; Utsunomya, T.; Zhang, N.; Ueno, M.; Yao, Z.; Goodman, S.D. Modulation of the Inflammatory Response and Bone Healing. Front. Endocrinology 2020, 11, 386. [Google Scholar] [CrossRef]
- Wu, A.C.; Raggatt, L.J.A.; Alexander, K.; Pettit, A.R. Unraveling Macrophage Contributions to Bone Repair. Bone Key Rep. 2013, 2, 373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsell, R.A.; Einhorn, T. The Biology of Fracture Healing. Injury 2011, 42, 551–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindeler, A.; McDonald, M.M.; Bokko, P.; Little, D.G. BOne Remodeling during Fracture Repair: The Cellular Picture. Semin. Cell Dev. Biol. 2008, 19, 459–466. [Google Scholar] [CrossRef]
- Longoni, A.; Knežević, L.; Schepers, K.; Weinans, H.; Rosenberg, A.J.W.P.; Gawlitta, D. The Impact of Immune Response on Endochondral Bone Regeneration. Npj Regen. Med. 2018, 3, 22. [Google Scholar] [CrossRef]
- Schmidt-Bleek, K.; Schell, H.; Schulz, N.; Hoff, P.; Perka, C.; Buttgereit, F.; Volk, H.-D.; Lienau, J.; Duda, G.N. Inflammatory Phase of Bone Healing Initiates the Regenerative Healing Cascade. Cell Tissue Res. 2011, 347, 567–573. [Google Scholar] [CrossRef]
- Xing, Z.; Lu, C.; Hu, D.; Miclau, T.; Marcucio, R. Rejuvenation of the Inflammatory System Stimulates Fracture Repair in Aged Mice. J. Orthop. Res. 2010, 28, 1000–1006. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-K.; Lorenzo, J. Cytokines Regulating Osteoclast Formation and Function. Curr. Opin. Rheumatol. 2006, 18, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Kon, T.; Cho, T.-J.; Aizawa, T.; Yamazaki, M.; Nooh, N.; Graves, D.T.; Gerstenfeld, L.C.; Einhorn, T.A. Expression of Osteoprotegerin, Receptor Activator of NF-κB Ligand (Osteoprotegerin Ligand) and Related Proinflammatory Cytokines During Fracture Healing. J. Bone Miner. Res. 2001, 16, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Mountziaris, P.M.; Mikos, A.G. Modulation of the Inflammatory Response for Enhanced Bone Tissue Regeneration. Tissue Eng. Part B Rev. 2008, 14, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Gerstenfeld, L.C.; Thiede, M.; Seibert, K.; Mielke, C.; Phippard, D.; Svagr, B.; Cullinane, D.A.; Einhorn, T. Differential Inhibition of Fracture Healing by Non-selective and Cyclooxygenase-2 Selective Non-Steroidal Anti-Inflammatory Drugs. J. Orthop. Res. 2003, 21, 670–675. [Google Scholar] [CrossRef]
- Mullis, B.H.; Copland, S.T.; Weinhold, P.S.; Miclau, T.; Lester, G.E.; Bos, G.D. Effect of COX-2 Inhibitors and Non-Steroidal Anti-Inflammatory Drugs on a Mouse Fracture Model. Injury 2006, 37, 827–837. [Google Scholar] [CrossRef]
- Costantino, M.; Schuster, A.; Helmholz, H.; Meyer-Rachner, A.; Willumeit-Römer, R.; Luthringer-Feyerabend, B.J. Inflammatory Response to Magnesium-Based Biodegradable Implant Materials. Acta Biomater. 2020, 101, 598–608. [Google Scholar] [CrossRef]
- Glass, G.E.; Chan, J.K.; Freidin, A.; Feldmann, M.; Horwood, N.J.; Nanchahal, J. TNF-Promotes Fracture Repair by Augmenting the Recruitment and Differentiation of Muscle-Derived Stromal Cells. Proc. Natl. Acad. Sci. USA 2011, 108, 1585–1590. [Google Scholar] [CrossRef] [Green Version]
- Gerstenfeld, L.; Cho, T.-J.; Kon, T.; Aizawa, T.; Tsay, A.; Fitch, J.; Barnes, G.L.; Graves, D.T.A.; Einhorn, T. Impaired Fracture Healing in the Absence of TNF-α Signaling: The Role of TNF-α in Endochondral Cartilage Resorption. J. Bone Miner. Res. 2003, 18, 1584–1592. [Google Scholar] [CrossRef]
- Lange, J.; Sapozhnikova, A.; Lu, C.; Hu, D.; Li, X.; Miclau, T.; Marcucio, R. Action of IL-1β During Fracture Healing. J. Orthop. Res. 2009, 28, 778–784. [Google Scholar] [CrossRef] [Green Version]
- Bonaventura, P.; Benedetti, G.; Albarède, F.; Miossec, P. Zinc and Its Role in Immunity and Inflammation. Autoimmun. Rev. 2015, 14, 277–285. [Google Scholar] [CrossRef]
- Hanley, C.; Thurber, A.; Hanna, C.; Punnoose, A.; Zhang, J.; Wingett, D.G. The Influences of Cell Type and ZnO Nanoparticle Size on Immune Cell Cytotoxicity and Cytokine Induction. Nanoscale Res. Lett. 2009, 4, 1409–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heng, B.C.; Zhao, X.; Xiong, S.; Ng, K.W.; Boey, F.Y.-C.; Loo, J.S.-C. Toxicity of Zinc Oxide (ZnO) Nanoparticles on Human Bronchial Epithelial Cells (BEAS-2B) Is Accentuated by Oxidative Stress. Food Chem. Toxicol. 2010, 48, 1762–1766. [Google Scholar] [CrossRef] [PubMed]
- Mayer, L.S.; Uciechowski, P.; Meyer, S.; Schwerdtle, T.; Rink, L.; Haase, H. Differential Impact of Zinc Deficiency on Phagocytosis, Oxidative Burst, and Production of Pro-Inflammatory Cytokines by Human Monocytes. Metals 2014, 6, 1288–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ralston, S.H.; Ho, L.-P.; Helfrich, M.H.; Grabowski, P.S.; Johnston, P.W.; Benjamin, N. Nitric Oxide: A Cytokine-Induced Regulator of Bone Resorption. J. Bone Miner. Res. 2009, 10, 1040–1049. [Google Scholar] [CrossRef]
- Zamora, R.; Vodovotz, Y.; Billiar, T.R. Inducible Nitric Oxide Synthase and Inflammatory Diseases. Mol. Med. 2000, 6, 347–373. [Google Scholar] [CrossRef] [Green Version]
- Wimalawansa, S.J. Nitric Oxide: Novel Therapy for Osteoporosis. Expert Opin. Pharmacother. 2008, 9, 3025–3044. [Google Scholar] [CrossRef]
- Kwon, D.W.; Jeong, J.W.; Choi, E.O.; Lee, H.W.; Lee, K.W.; Kim, K.Y.; Kim, S.G.; Hong, S.H.; Kim, G.Y.; Park, C.; et al. Inhibitory Effects on the Production of Inflammatory Mediators and Reactive Oxygen Species by Mori Folium in Lipopolysaccharide-Stimulated Macrophages and Zebrafish. An. Acad. Bras. Cienc. 2017, 89, 661–674. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Chen, C.; Jia, G.; Li, Y.; Zhang, J.; Huang, H.; Kang, B.; Yuan, G.; Zeng, H.; Chen, T.-X. The Bioeffects of Degradable Products Derived from a Biodegradable Mg-Based Alloy in Macrophages via Heterophagy. Acta Biomater. 2020, 106, 428–438. [Google Scholar] [CrossRef]
- Lee, J.A.; Song, H.Y.; Ju, S.M.; Lee, S.J.; Kwon, H.-J.; Eum, W.S.; Jang, S.H.; Choi, S.Y.; Park, J. Differential Regulation of Inducible Nitric Oxide Synthase and Cyclooxygenase-2 Expression by Superoxide Dismutase in Lipopolysaccharide Stimulated RAW 264.7 Cells. Exp. Mol. Med. 2009, 41, 629–637. [Google Scholar] [CrossRef]
- Lee, M.-C.; Yoshino, F.; Shoji, H.; Takahashi, S.; Todoki, K.; Shimada, S.; Kuse-Barouch, K. Characterization by Electron Spin Resonance Spectroscopy of Reactive Oxygen Species Generated by Titanium Dioxide and Hydrogen Peroxide. J. Dent. Res. 2005, 84, 178–182. [Google Scholar] [CrossRef]
- Lee, H.-P.; Lin, D.-J.; Yeh, M.-L. Phenolic Modified Ceramic Coating on Biodegradable Mg Alloy: The Improved Corrosion Resistance and Osteoblast-Like Cell Activity. Materials 2017, 10, 696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bubols, G.B.; Vianna, D.D.R.; Remon, A.M.; Von Poser, G.L.; Lamuela-Raventos, R.M.; Eifler-Lima, V.L.; Garcia, S.C. The Antioxidant Activity of Coumarins and Flavonoids. Mini-Rev. Med. Chem. 2013, 13, 318–334. [Google Scholar] [CrossRef] [PubMed]
- Marreiro, D.D.N.; Cruz, K.J.C.; Morais, J.B.S.; Beserra, J.B.; Severo, J.S.; De Oliveira, A.R.S. Zinc and Oxidative Stress: Current Mechanisms. Antioxidants 2017, 6, 24. [Google Scholar] [CrossRef]
- Vignesh, K.S.; Deepe, G.S., Jr. Metallothioneins: Emerging Modulators in Immunity and Infection. Int. J. Mol. Sci. 2017, 18, 2197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, O.H.; Panicker, L.M.; Lu, Q.; Chae, J.J.; Feldman, R.A.; Elisseeff, J.H. Human iPSC-Derived Osteoblasts and Osteoclasts Together Promote Bone Regeneration in 3d Biomaterials. Sci. Rep. 2016, 6, 26761. [Google Scholar] [CrossRef] [PubMed]
- Steffi, C.; Shi, Z.; Kong, C.H.; Wang, W. Modulation of Osteoclast Interactions with Orthopaedic Biomaterials. J. Funct. Biomater. 2018, 9, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stover, M. Distal Femoral Fractures: Current Treatment, Results and Problems. Injury 2001, 32, 3–13. [Google Scholar] [CrossRef]
- Syed, A.; Agarwal, M.; Giannoudis, P.; Matthews, S.; Smith, R. Distal Femoral Fractures: Long-Term Outcome Following Stabilisation with LPS. Injury 2004, 35, 599–607. [Google Scholar] [CrossRef]
- Bolhofner, B.R.; Carmen, B.; Clifford, P. The Results of Open Reduction and Internal Fixation of Distal Femur Fractures Using a Biologic (Indirect) Reduction Technique. J. Orthop. Trauma 1996, 10, 372–377. [Google Scholar] [CrossRef]
- Tsurukai, T.; Udagawa, N.; Matsuzaki, K.; Takahashi, N.; Suda, T. Roles of Macrophage-Colony Stimulating Factor and Osteoclast Differentiation Factor in Osteoclastogenesis. J. Bone Miner. Metab. 2000, 18, 177–184. [Google Scholar] [CrossRef]
- Tanaka, S.; Takahashi, N.; Udagawa, N.; Tamura, T.; Akatsu, T.; Stanley, E.R.; Kurokawa, T.; Suda, T. Macrophage Colony-Stimulating Factor Is Indispensable for Both Proliferation and Differentiation of Osteoclast Progenitors. J. Clin. Investig. 1993, 91, 257–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadowska, J.M.; Ginebra, M.-P. Inflammation and Biomaterials: Role of the Immune Response in Bone Regeneration by Inorganic Scaffolds. J. Mater. Chem. B 2020, 8, 9404–9427. [Google Scholar] [CrossRef] [PubMed]
- Jimi, E.; Nakamura, I.; Amano, H.; Taguchi, Y.; Tsurukai, T.; Tamura, M. Osteoclast Function Is Activated by Osteoblastic Cells through a Mechanism Involving Cell-To-Cell Contact. Endocrinology 1996, 137, 2187–2190. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Windle, J.J.; Koop, B.A.; Mundy, G.R.; Bonewald, L.F. Establishment of an Osteocyte-Like Cell Line, MLO-Y4. J. Bone Miner. Res. 2010, 12, 2014–2023. [Google Scholar] [CrossRef]
- Zhao, S.; Kato, Y.; Zhang, Y.K.Y.; Harris, S.; Ahuja, S.S.; Bonewald, L.F. MLO-Y4 Osteocyte-Like Cells Support Osteoclast Formation and Activation. J. Bone Miner. Res. 2002, 17, 2068–2079. [Google Scholar] [CrossRef]
- Xing, L. Regulation of Osteoclast Precursors in Inflammatory Bone Loss. Curr. Opin. Investig. Drugs 2009, 10, 1195–1203. [Google Scholar]
- Takahashi, N.; Udagawa, N.; Suda, T. A New Member of Tumour Necrosis Factor Ligand Family, ODF/OPGL/TRANCE/RANKL, Regulates Osteoclast Differentiation and Function. Biochem. Biophys. Res. Commun. 1999, 256, 449–455. [Google Scholar] [CrossRef]
- Tanakas, S.; Nakamura, I.; Inoue, J.-I.; Oda, H.; Nakamura, K. Signal Transduction Pathways Regulating Osteoclast Differentiation and Function. J. Bone Miner. Metab. 2003, 21, 123–133. [Google Scholar] [CrossRef]
- Wong, B.R.; Josien, R.; Lee, S.Y.; Vologodskaia, M.; Steinman, R.M.; Choi, Y. The TRAF Family of Signal Transducers Mediates NF-KappaB Activation by the TRANCE Receptor. J. Biol. Chem. 1998, 273, 28355–28359. [Google Scholar] [CrossRef] [Green Version]
- Tanakas, S.; Nakamura, K.; Takahasi, N.; Suda, T. Role of RANKL In Physiological and Pathological Bone Resorption and Therapeutics Targeting The RANKL-RANK Signaling System. Immunol. Rev. 2005, 208, 30–49. [Google Scholar] [CrossRef]
- Honma, M.; Ikebuchi, Y.; Kariya, Y.; Suzuki, H. Regulatory Mechanisms of RANKL Presentation to Osteoclast Precursors. Curr. Osteoporos. Rep. 2014, 12, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Asagiri, M.; Sato, K.; Usami, T.; Ochi, S.; Nishina, H.; Yoshida, H.; Morita, I.; Wagner, E.F.; Mak, T.W.; Serfling, E.; et al. Autoamplification of NFATc1 Expression Determines Its Essential Role in Bone Homeostasis. J. Exp. Med. 2005, 202, 1261–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Chang, E.J.; Ryu, J.; Lee, Y.; Kim, H.H. Induction of c-Fos and NAFTc1 during RANKL-Stimulated Osteoclast Differentiation Is Mediated by the p38 Signalling Pathway. Biochem. Biophys. Res. Commun. 2006, 351, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Kirstein, B.; Chambers, T.J.; Fuller, K. Secretion of Tartrate-Resistant Acid Phosphatase by Osteoclasts Correlates with Resorptive Behavior. J. Cell. Biochem. 2006, 98, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Hayman, A.R. Tartrate-Resistant Acid Phosphatase (TRAP) and the Osteoclast/Immune Cell Dichotomy. Autoimmunity 2008, 41, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Qu, X.; Li, H.; Yang, K.; Wan, P.; Tan, L.; Ouyang, Z.; Liu, X.; Tian, B.; Xiao, F.; et al. The Effect of Metallic Magnesium Degradation Products on Osteoclast-Induced Osteolysis and Attenuation of NF-κB and NFATc1 signaling. Biomaterials 2014, 35, 6299–6310. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.R.; Peters, C.; Saftig, P.; Brömme, D. Cathepsin K Activity-Dependent Regulation of Osteoclast Actin Ring Formation and Bone Resorption. J. Biol. Chem. 2009, 284, 2584–2592. [Google Scholar] [CrossRef] [Green Version]
- Takito, J.; Inoue, S.; Nakamura, M. The Sealing Zone in Osteoclasts: A Self-Organized Structure on the Bone. Int. J. Mol. Sci. 2018, 19, 984. [Google Scholar] [CrossRef] [Green Version]
- Saltel, F.; Destaing, O.; Bard, F.; Eichert, D.; Jurdic, P. Apatite-Mediated Actin Dynamics in Resorbing Osteoclasts. Mol. Biol. Cell 2004, 15, 5231–5241. [Google Scholar] [CrossRef]
- Naaldijk, Y.; Friedrich-Stöckigt, A.; Sethe, S.; Stolzing, A. Comparison of Different Cooling Rates for Fibroblast and Keratinocyte Cryopreservation. J. Tissue Eng. Regen. Med. 2013, 10, E354–E364. [Google Scholar] [CrossRef]
- Han, G.; Zuo, J.; Holliday, L.S. Specialized Roles for Actin in Osteoclasts: Unanswered Questions and Therapeutic Opportunities. Biomolecules 2019, 9, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Luthringer-Feyerabend, B.J.; Feyerabend, F.; Schilling, A.F.; Willumeit, R. Effects of Extracellular Magnesium on the Differentiation and Function of Human Osteoclasts. Acta Biomater. 2014, 10, 2843–2854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Feyerabend, F.; Schilling, A.F.; Willumeit-Römer, R.; Luthringer-Feyerabend, B.J. Effects of Extracellular Magnesium Extract on the Proliferation and Differentiation of Human Osteoblasts and Osteoclasts in Coculture. Acta Biomater. 2015, 27, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Totea, G.; Ionita, D.; Demetrescu, I. ICP-MS Determinations in Sustaining Corrosion Data of 316 Stainless Steels in Bioliquids. UPB Sci. Bull. B 2014, 76, 57–66, ISSN 1454-2331. [Google Scholar]
- Esmaily, M.; Svensson, J.E.; Fajardo, S.; Birbilis, N.; Frankel, G.S.; Virtanen, S.; Arrabal, R.; Thomas, S.; Johansson, L.G. Fundamentals and Advances in Magnesium Alloy Corrosion. Prog. Mater. Sci. 2017, 89, 92–193. [Google Scholar] [CrossRef]
- Neacsu, P.; Staras, A.; Voicu, S.I.; Ionascu, I.; Soare, T.; Uzun, S.; Cojocaru, D.V.; Pandele, A.-M.; Croitoru, S.M.; Miculescu, F.; et al. Characterization and In Vitro and In Vivo Assessment of a Novel Cellulose Acetate-Coated Mg-Based Alloy for Orthopedic Applications. Materials 2017, 10, 686. [Google Scholar] [CrossRef] [Green Version]
- Dascălu, C.-A.; Maidaniuc, A.; Pandele, A.M.; Voicu, S.I.; Machedon-Pisu, T.; Stan, G.E.; Cîmpean, A.; Mitran, V.; Antoniac, I.V.; Miculescu, F. Synthesis and Characterization of Biocompatible Polymer-Ceramic Film Structures as Favorable Interface in Guided Bone Regeneration. Appl. Surf. Sci. 2019, 494, 335–352. [Google Scholar] [CrossRef]
- Ion, R.; Mazare, A.; Dumitriu, C.; Pirvu, C.; Schmuki, P.; Cimpean, A. Nanochannelar Topography Positively Modulates Osteoblast Differentiation and Inhibits Osteoclastogenesis. Coatings 2018, 8, 294. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | O1s (at.%) | C1s (at.%) | Mg1s (at.%) | Al2p (at.%) |
|---|---|---|---|---|
| Mg | 46 | 33 | 19 | 2 |
| Mg-PCL | 22 | 78 | − | − |
| Mg-PCL-CM | 21 | 79 | − | − |
| Mg-PCL-CM-ZnO | 24 | 76 | − | − |
| Mg-PCL-ZnO | 23 | 77 | − | − |
| Sample | Tafel Slope Method | Polarisation Resistance Method | ||||
|---|---|---|---|---|---|---|
| Ecorr, V | icorr, μA/cm2 | Kg, g/m2h | P, mm/an | RP, Ω | icorr, μA/cm2 | |
| Mg | −1.61 ± 0.02 | 144 ± 1.45 | 1.639 ± 0.02 | 1.751 ± 0.02 | 195 ± 4.5 | 114.52 ± 1.12 |
| Mg-PCL | −1.52 ± 0.02 | 63 ± 0.67 | 0.717 ± 0.01 | 0.766 ± 0.07 | 865 ± 7 | 27.61 ± 0.25 |
| Mg-PCL-CM | −1.48 ± 0.01 | 36.6 ± 0.11 | 0.416 ± 001 | 0.445 ± 0.002 | 923 ± 6 | 28.03 ± 0.16 |
| Mg-PCL-CM-ZnO | −1.46 ± 0.01 | 11.4 ± 0.05 | 0.129 ± 0.001 | 0.138 ± 0.001 | 2898 ± 8 | 8.21 ± 0.04 |
| Mg-PCL-ZnO | −1.49 ± 0.01 | 10.5 ± 0.04 | 0.119 ± 0.001 | 0.127 ± 0.001 | 3327 ± 9 | 7.15 ± 0.03 |
| 100 ng/mL LPS | Total Number of Nuclei | Nuclei in Multinuclear Cells | Multinuclear Index (%) |
|---|---|---|---|
| TCPS | 1110 | 90 | 8.1 |
| Mg Alloy | 343 | 71 | 21.3 |
| Mg-PCL | 774 | 69 | 8.9 |
| Mg-PCL-ZnO | 845 | 15 | 1.75 |
| Mg-PCL-CM | 790 | 81 | 10.2 |
| Mg-PCL-CM-ZnO | 869 | 27 | 3.1 |
| Component | NaCl | KCl | Na2HPO4 | KH2PO4 |
|---|---|---|---|---|
| Concentration, g/L | 8 | 0.2 | 1.144 | 0.2 |
| Code | Composition | ||
|---|---|---|---|
| PCL (wt.%) | CM (mg mL−1) | ZnO (mg mL−1) | |
| Mg-PCL | 14.25 | − | − |
| Mg-PCL-CM | 5.32 | − | |
| Mg-PCL-CM-ZnO | 5.32 | 0.25 | |
| Mg-PCL-ZnO | − | 0.25 | |
| Code | Voltage (kV) | Flow Rate (µL min−1) |
|---|---|---|
| Mg-PCL | 23 | 8 |
| Mg-PCL-CM | 23 | 8 |
| Mg-PCL-CM-ZnO | 25 | 12 |
| Mg-PCL-ZnO | 25 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Negrescu, A.-M.; Necula, M.-G.; Gebaur, A.; Golgovici, F.; Nica, C.; Curti, F.; Iovu, H.; Costache, M.; Cimpean, A. In Vitro Macrophage Immunomodulation by Poly(ε-caprolactone) Based-Coated AZ31 Mg Alloy. Int. J. Mol. Sci. 2021, 22, 909. https://doi.org/10.3390/ijms22020909
Negrescu A-M, Necula M-G, Gebaur A, Golgovici F, Nica C, Curti F, Iovu H, Costache M, Cimpean A. In Vitro Macrophage Immunomodulation by Poly(ε-caprolactone) Based-Coated AZ31 Mg Alloy. International Journal of Molecular Sciences. 2021; 22(2):909. https://doi.org/10.3390/ijms22020909
Chicago/Turabian StyleNegrescu, Andreea-Mariana, Madalina-Georgiana Necula, Adi Gebaur, Florentina Golgovici, Cristina Nica, Filis Curti, Horia Iovu, Marieta Costache, and Anisoara Cimpean. 2021. "In Vitro Macrophage Immunomodulation by Poly(ε-caprolactone) Based-Coated AZ31 Mg Alloy" International Journal of Molecular Sciences 22, no. 2: 909. https://doi.org/10.3390/ijms22020909
APA StyleNegrescu, A. -M., Necula, M. -G., Gebaur, A., Golgovici, F., Nica, C., Curti, F., Iovu, H., Costache, M., & Cimpean, A. (2021). In Vitro Macrophage Immunomodulation by Poly(ε-caprolactone) Based-Coated AZ31 Mg Alloy. International Journal of Molecular Sciences, 22(2), 909. https://doi.org/10.3390/ijms22020909