
NECTIN4: A Novel Therapeutic Target for Melanoma


Abstract
1. Introduction
2. Results
2.1. Characteristics of the Study Cohort
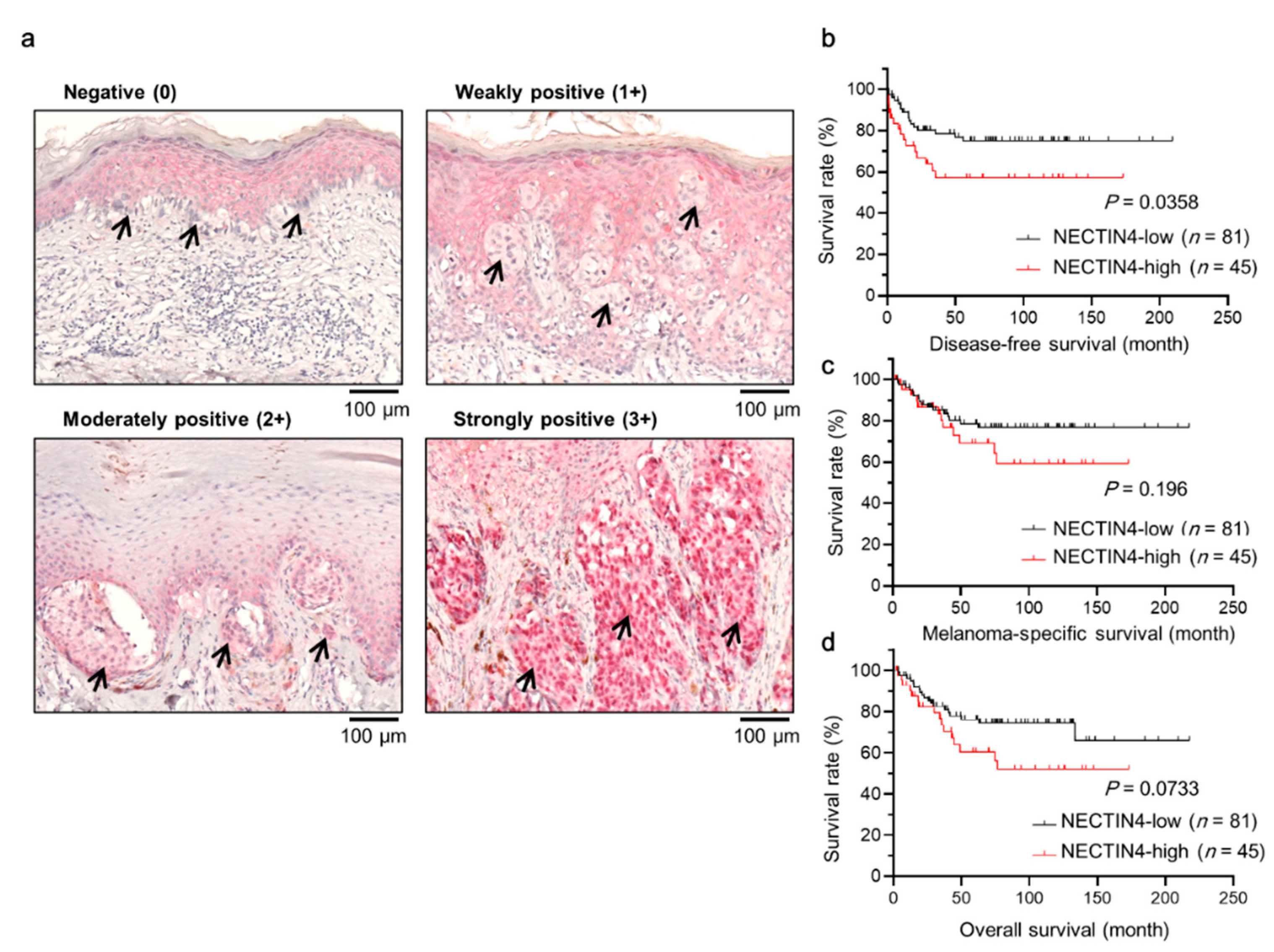
2.2. Association of NECTIN4 with Clinicopathological Factors and Prognostic Impact in Melanoma
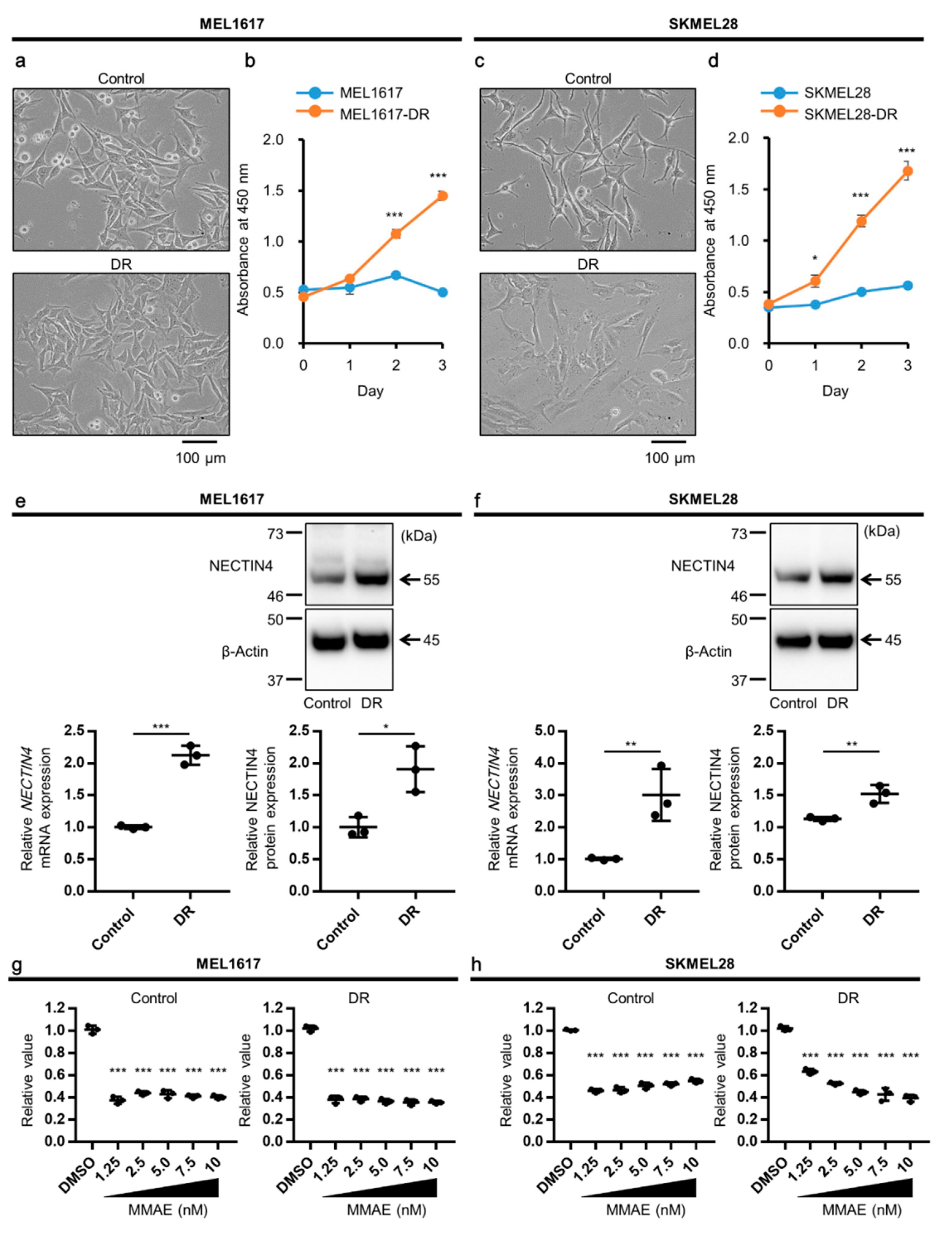
2.3. NECTIN4 Is Upregulated in BRAFi-Resistant Melanoma Cells
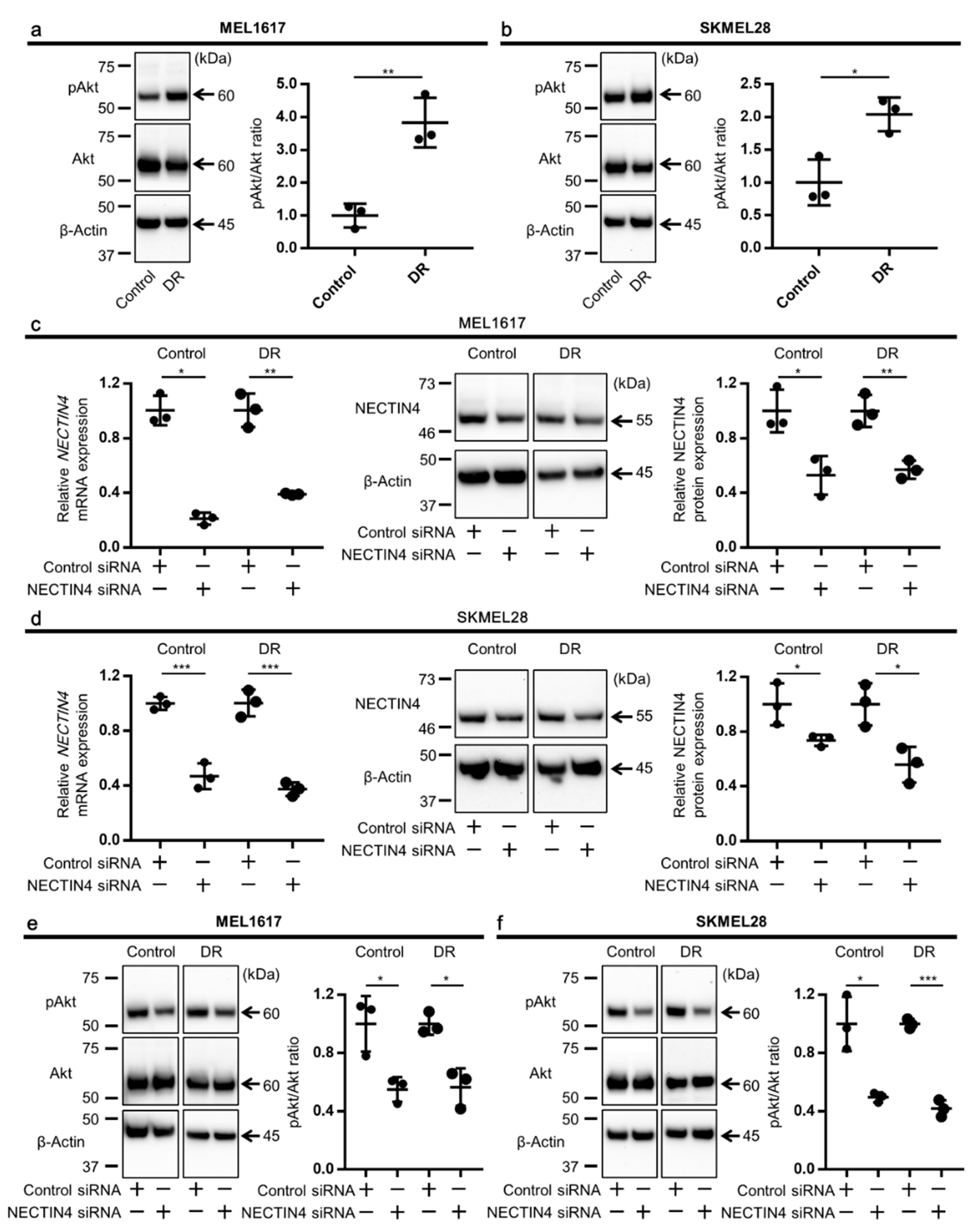
2.4. Akt Phosphorylation Is Enhanced in BRAFi-Resistant Melanoma Cells
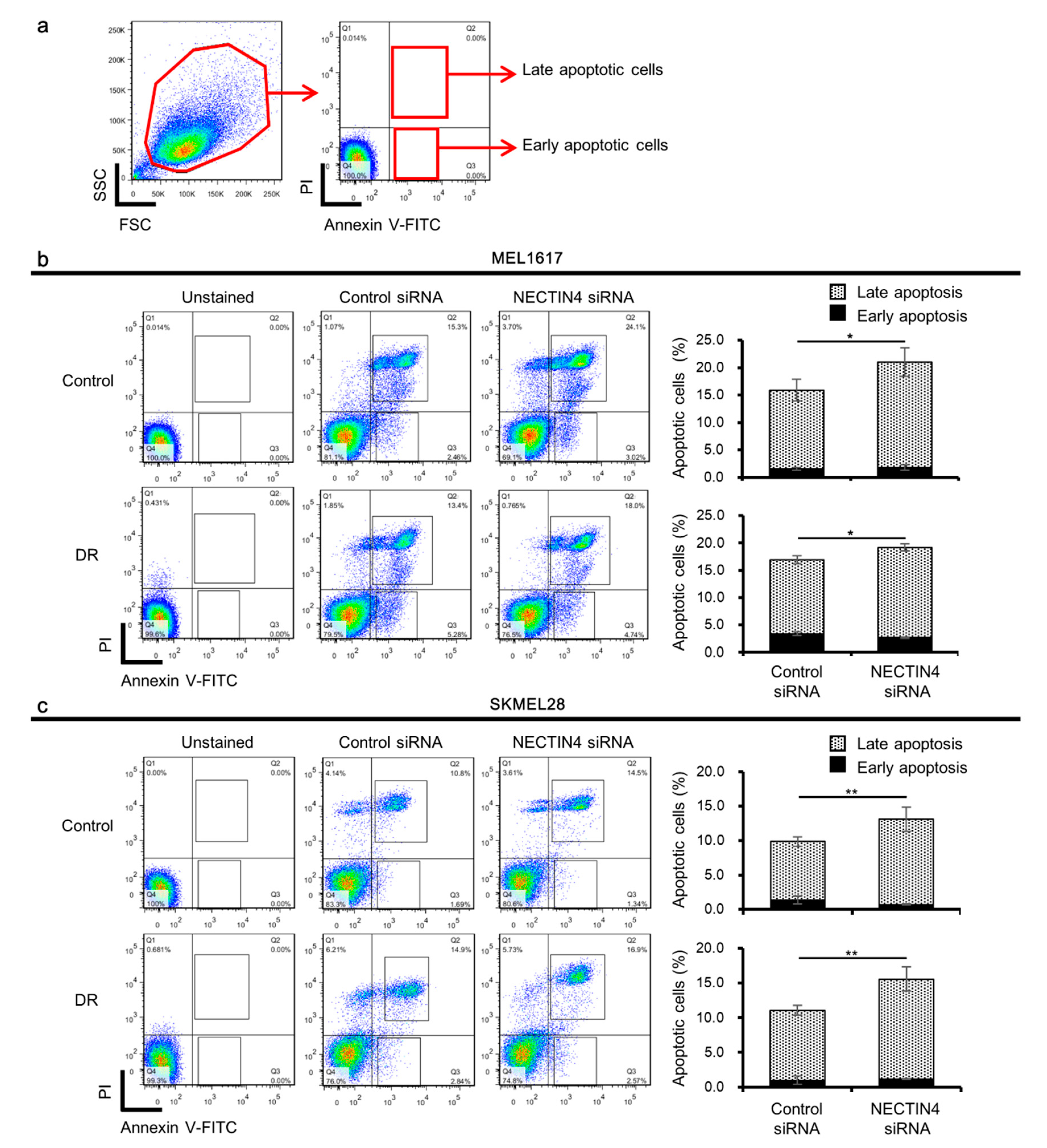
2.5. Knockdown of NECTIN4 Impairs PI3K/Akt Pathway and Increases Apoptosis of Melanoma Cells
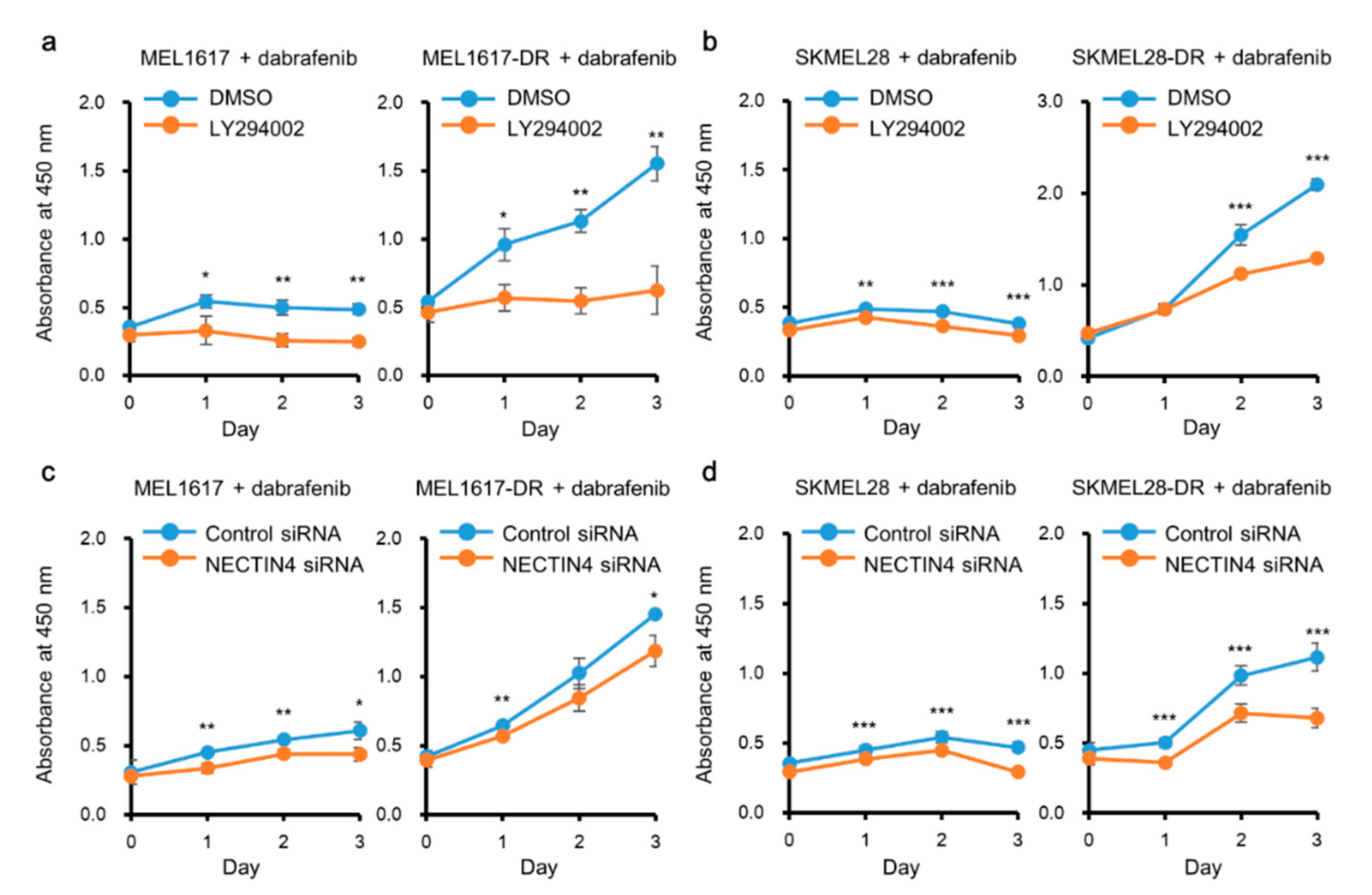
2.6. PI3K/Akt and NECTIN4 Inhibition Improved the Sensitivity of Melanoma Cells to BRAFi
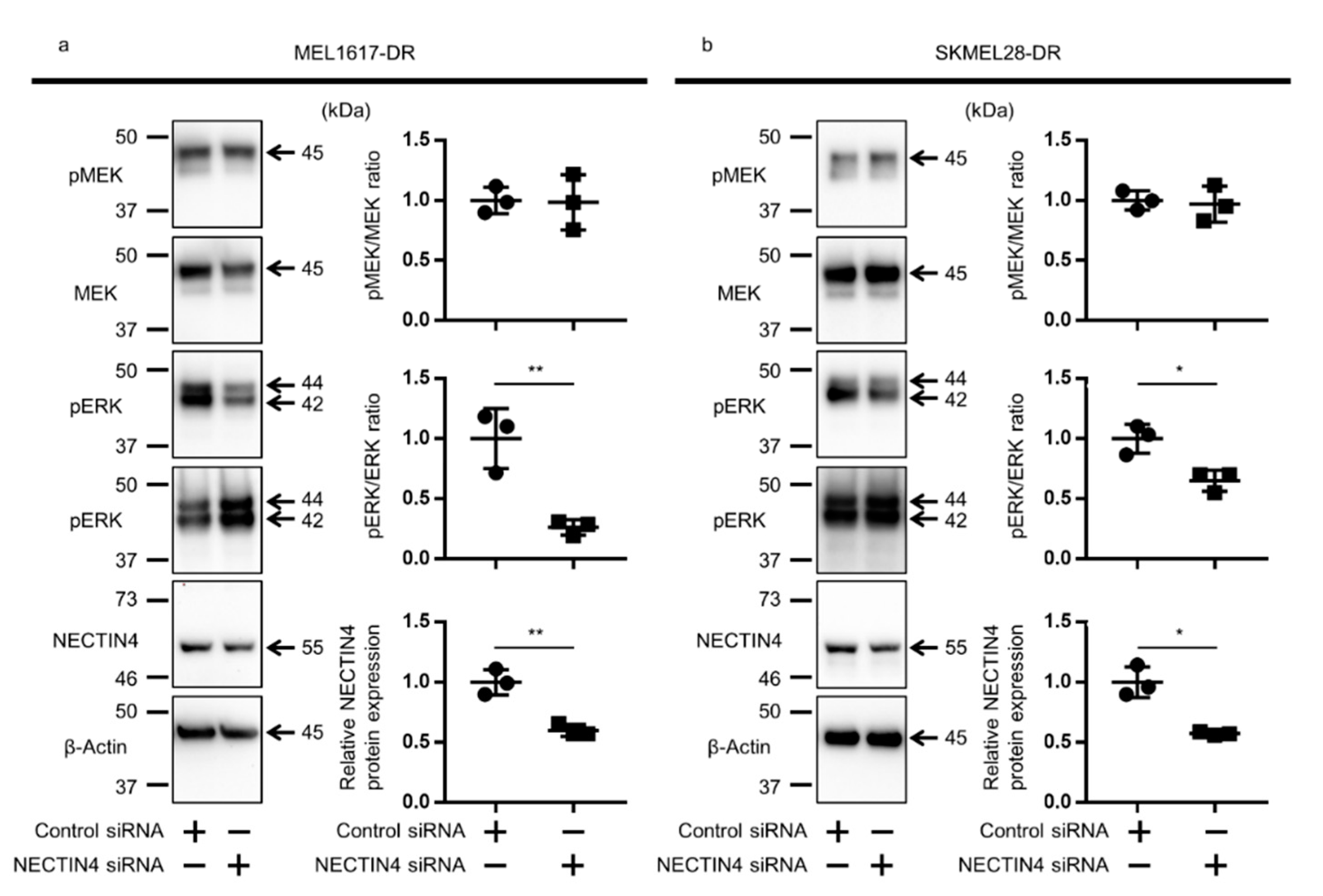
2.7. NECTIN4 Inhibition Down-Regulated ERK Signal in BRAFi-Resistant Melanoma Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Generation of BRAFi-Resistant Melanoma Cells
4.3. Cell Viability
4.4. siRNA Transfection
4.5. RNA Extraction and qRT-PCR
4.6. Western Blotting
4.7. Annexin V-PI Staining and Flow Cytometry
4.8. Patients
4.9. Immunohistological Analysis
4.10. Evaluation of NECTIN4 Immunohistochemical Staining
4.11. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NECTIN4 | NECTIN cell adhesion molecule 4 |
| BRAFi | BRAF inhibitor |
| MEKi | MEK inhibitor |
| ADC | Antibody–drug conjugate |
| PI | Propidium iodide |
| qRT-PCR | Quantitative reverse transcription polymerase chain reaction |
| DR | Dabrafenib-resistant |
| MMAE | Monomethyl auristatin E |
| DFS | Disease-free survival |
| MSS | Melanoma-specific survival |
| OS | Overall survival |
| CCK-8 | Cell Counting Kit-8 |
References
- Rastrelli, M.; Tropea, S.; Rossi, C.R.; Alaibac, M. Melanoma: Epidemiology, risk factors, pathogenesis, diagnosis, and classification. In Vivo 2014, 28, 1005. [Google Scholar] [PubMed]
- Erdei, E.; Torres, M. A new understanding in the epidemiology of melanoma. Expert Rev. Anticancer Ther. 2010, 10, 1811–1818. [Google Scholar] [CrossRef] [PubMed]
- Parkin, D.M.; Bray, F.; Ferlay, J.; Pisani, P. Estimating the world cancer burden: Globocan 2000. Int. J. Cancer 2001, 94, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Robert, C.; Long, G.V.; Brady, B.; Dutriaux, C.; Maio, M.; Mortier, L.; Hassel, J.C.; Rutkowski, P.; McNeil, C.; Kalinka-Warzocha, E.; et al. Nivolumab in preciously untreated melanoma without BRAF mutation. N. Engl. J. Med. 2015, 372, 320–330. [Google Scholar] [CrossRef]
- Hennequin, C.; Rio, E.; Mahé, M.A. Radiotherapy of skin cancers. Cancer Radiother. 2016, 20, S249–S255. [Google Scholar] [CrossRef]
- Franklin, C.; Livingstone, E.; Roesch, A.; Shilling, B.; Scgadendorf, D. Immunotherapy in melanoma: Recent advances and future directions. Eur. J. Surg. Oncol. 2017, 43, 604–611. [Google Scholar] [CrossRef]
- Mahoney, K.M.; Freeman, G.J.; McDermott, D.F. The next immune-checkpoint inhibitors: PD-1/PD-L1 blockade in melanoma. Clin. Ther. 2015, 37, 764–782. [Google Scholar] [CrossRef]
- Gide, T.N.; Wilmott, J.S.; Scolyer, R.A.; Long, G.V. Primary and acquired resistance to immune checkpoint inhibitors in metastatic melanoma. Clin. Cancer Res. 2017, 24, OF1–OF11. [Google Scholar] [CrossRef]
- Shergold, A.L.; Millar, R.; Nibbs, R.J.B. Understanding and overcoming the resistance of cancer PD-1/PD-LI blockade. Pharmacol. Res. 2019, 145, 104258. [Google Scholar] [CrossRef]
- Bradish, J.R.; Cheng, L. Molecular pathology of malignant melanoma: Changing the clinical practice paradigm toward a personalized approach. Hum. Pathol. 2014, 45, 1315–1326. [Google Scholar] [CrossRef]
- Davies, H.; Bignell, G.R.; Cox, C.; Stephens, P.; Edkins, S.; Clegg, S.; Teague, J.; Woffendin, H.; Garnett, M.J.; Bottomley, W.; et al. Mutations of the BRAF gene in human cancer. Nature 2002, 417, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Pavri, S.N.; Clune, J.; Ariyan, S.; Narayan, D. Malignant melanoma: Beyond the basics. Plast. Reconstr. Surg. 2016, 138, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Ascierto, P.A.; Kirkwood, J.M.; Grob, J.-J.; Simeone, E.; Grimaldi, A.M.; Maio, M.; Palmieri, G.; Testori, A.; Marincola, F.M.; Mozzillo, N. The role of BRAF V600 mutation in melanoma. J. Transl. Med. 2012, 10, 85. [Google Scholar] [CrossRef] [PubMed]
- Hauschild, A.; Grob, J.-J.; Demidov, L.V.; Jouary, T.; Gutzmer, R.; Millward, M.; Rutkowski, P.; Blank, C.U.; Miller Jr, W.H.; Kaempgen, E.; et al. Dabrafenib in BRAF-mutated metastatic melanoma: A multicenter, open-label, phase 3 randomised controlled trial. Lancet 2012, 380, 358–365. [Google Scholar] [CrossRef]
- Poulikakos, P.I.; Persaud, Y.; Janakiraman, M.; Kong, X.; Ng, C.; Moriceau, G.; Shi, H.; Atefi, M.; Titz, B.; Gabay, M.T.; et al. RAF inhibitor resistance is mediated by dimerization of aberrantly spliced BRAF(V600E). Nature 2012, 480, 387–390. [Google Scholar] [CrossRef]
- Hirata, E.; Girotti, M.R.; Viros, A.; Hooper, S.; Spencer-Dene, B.; Matsuda, M.; Larkin, J.; Marais, R.; Sahai, E. Intravital imaging reveals how BRAF inhibition generates drug-tolerant microenvironments with high integrin β1/FAK signaling. Cancer Cell 2015, 27, 574–588. [Google Scholar] [CrossRef]
- Samanta, D.; Almo, S.C. Nectin family of cell-adhesion molecules: Structural and molecular aspects of function and specificity. Cell Mol. Life Sci. 2015, 72, 645–658. [Google Scholar] [CrossRef]
- Rikitake, Y.; Mandai, K.; Takai, Y. The role of nectins in different types of cell-cell adhesion. J. Cell Sci. 2012, 125, 3713–3722. [Google Scholar] [CrossRef]
- Ogita, H.; Ikeda, W.; Takai, Y. Roles of cell adhesion molecules nectin and nectin-like molecule-5 in the regulation of cell movement and proliferation. J. Microsc. 2008, 231, 455–465. [Google Scholar] [CrossRef]
- Takai, Y.; Irie, K.; Shimizu, K.; Sakisaka, T.; Ikeda, W. Nectins and nectin-like molecules: Roles in cell adhesion, migration, and polarization. Cancer Sci. 2003, 94, 655–667. [Google Scholar] [CrossRef]
- Miyoshi, J.; Takai, Y. Nectin and nectin-like molecules: Biology and pathology. Am. J. Nephrol 2007, 27, 590–604. [Google Scholar] [CrossRef] [PubMed]
- Nishiwada, S.; Sho, M.; Yasuda, S.; Shimada, K.; Yamato, I.; Akahori, T.; Kinoshita, S.; Nagai, M.; Konishi, N.; Nakajima, Y. Nectin-4 expression contributes to tumor proliferation, angiogenesis and patient prognosis in human pancreatic cancer. J. Exp. Clin. Cancer Res. 2015, 34, 30. [Google Scholar] [CrossRef] [PubMed]
- Fabre-Lafay, S.; Monville, F.; Garrido-Urbani, S.; Berruyer-Pouyet, C.; Ginestier, C.; Reymond, N.; Finetti, P.; Sauvan, R.; Adélaïde, J.; Geneix, J. Nectin-4 is a new histological and serological tumor associated marker for breast cancer. BMC Cancer 2007, 7, 73. [Google Scholar] [CrossRef]
- Challita-Eid, P.M.; Satpayev, D.; Yang, P.; An, Z.; Morrison, K.; Shostak, Y.; Raitano, A.; Nadell, R.; Liu, W.; Lortie, D.R.; et al. Enfortumab vedotin antibody-drug conjugate targeting nectin-4 is a highly potent therapeutic agent in multiple preclinical cancer models. Cancer Res. 2016, 76, 3003–3013. [Google Scholar] [CrossRef] [PubMed]
- Manabe, S.; Yamaguchi, Y.; Matsumoto, K.; Fuchigami, H.; Kawase, T.; Hirose, K.; Mitani, A.; Sumiyoshi, W.; Kinoshita, T.; Abe, J.; et al. Characterization of antibody products obtained through enzymatic and nonenzymatic glycosylation reactions with a glycan oxazoline and preparation of a homogeneous antibody-drug conjugate via Fc N-glycan. Bioconjug. Chem. 2019, 30, 1343–1355. [Google Scholar] [CrossRef]
- Beck, A.; Goetsch, L.; Dumontet, C.; Corvaia, N. Strategies and challenges for the next generation of antibody-drug conjugates. Nat. Rev. Drug Discov. 2017, 16, 315–337. [Google Scholar] [CrossRef]
- Carter, P.J.; Lazar, G.A. Next generation antibody drugs: Pursuit of the ‘high-hanging fruit’. Nat. Rev. Drug Discov. 2017, 17, 197–223. [Google Scholar] [CrossRef]
- Kaplon, H.; Reichert, J.M. Antibodies to watch in 2019. MAbs 2019, 11, 219–238. [Google Scholar] [CrossRef]
- Rosenberg, J.E.; O’Donnell, P.H.; Balar, A.V.; McGregor, B.A.; Heath, E.I.; Yu, E.Y.; Galsky, M.D.; Hahn, N.M.; Gartner, E.M.; Pinelli, J.M.; et al. Pivotal trial of enfortumab vedotin in urothelial carcinoma after platinum and anti-programmed death 1/programmed death ligand 1 therapy. J. Clin. Oncol. 2019, 37, 2592–2600. [Google Scholar] [CrossRef]
- Mollo, M.R.; Antonini, D.; Mitchell, K.; Fortugno, P.; Costanzo, A.; Dixon, J.; Brancati, F.; Missero, C. P63-dependent and independent mechanisms of nectin-1 and nectin-4 regulation in the epidermis. Exp. Dermatol. 2015, 24, 114–119. [Google Scholar] [CrossRef]
- Brancati, F.; Fortugno, P.; Bottillo, I.; Lopez, M.; Josselin, E.; Boudghene-Stambouli, O.; Agolini, E.; Bernardini, L.; Bellacchio, E.; Iannicelli, M.; et al. Mutations in PVRL4, encoding cell adhesion molecule nectin-4, cause ectodermal dysplasia-syndactyly syndrome. Am. J. Hum. Genet. 2010, 87, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Ito, T.; Tanaka, Y.; Kaku-Ito, Y.; Furue, M. NECTIN4 expression in extramammary Paget’s disease: Implication of a new therapeutic target. Int. J. Mol. Sci. 2020, 21, 5891. [Google Scholar] [CrossRef] [PubMed]
- McClelland, R.A.; Finlay, P.; Walker, K.J.; Nicholson, D.; Robertson, J.F.; Blamey, R.W.; Nicholson, R.I. Automated quantification of immunocytochemically localized estrogen receptors in human breast cancer. Cancer Res. 1990, 50, 3545–3550. [Google Scholar] [PubMed]
- Kakadia, S.; Yarlagadda, N.; Awad, R.; Kundranda, M.; Niu, J.; Naraev, B.; Mina, L.; Dragovich, T.; Gimbel, M.; Mahmoud, F. Mechanisms of resistance to BRAF and MEK inhibitors and clinical update of US Food and Drug Administration-approved targeting therapy in advanced melanoma. Onco. Targets Ther. 2018, 11, 7095–7107. [Google Scholar] [CrossRef] [PubMed]
- Torres-Collado, A.X.; Knott, J.; Jazirehi, A.R. Reversal of resistance in targeted therapy of metastatic melanoma: Lessons learned from Vemurafenib (BRAFV600E-specific inhibitor). Cancers 2018, 10, 157. [Google Scholar] [CrossRef] [PubMed]
- Turajlic, S.; Furney, S.; Stamp, G.; Rana, S.; Ricken, G.; Oduko, Y.; Saturno, G.; Springer, C.; Hayes, A.; Gore, M.; et al. Whole-genome sequencing reveals complex mechanisms of intrinsic resistance to BRAF inhibition. Ann. Oncol. 2014, 25, 959–967. [Google Scholar] [CrossRef]
- Luebker, S.A.; Koepsell, S.A. Diverse mechanisms of BRAF inhibitor resistance in melanoma inhibitor resistance in melanoma identified in clinical and preclinical studies. Front. Oncol. 2019, 9, 268. [Google Scholar] [CrossRef]
- Takahashi, S.; Uemura, M.; Kimura, T.; Kawasaki, Y.; Takamoto, A.; Yamaguchi, A.; Melhem-Bertrandt, A.; Gartner, E.M.; Inoue, T.; Akazawa, R.; et al. A phase I study of enfortumab vedotin in Japanese patients with locally advanced or metastatic urothelial carcinoma. Investig. New Drug 2020, 38, 1056–1066. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, S.; Wang, L.; Wu, Y.; Hao, J.; Wang, Z.; Lu, W.; Wang, X.-A.; Zhang, F.; Cao, Y.; et al. A novel PI3K/AKT signaling axis mediated Nectin-4-induced gallbladder cancer cell proliferation, metastasis and tumor growth. Cancer Lett. 2016, 375, 179–189. [Google Scholar] [CrossRef]
- Siddharth, S.; Goutam, K.; Das, S.; Nayak, A.; Nayak, D.; Sethy, C.; Wyatt, M.D.; Kundu, C.N. Nectin-4 is a breast cancer stem cell marker that induces WNT/β-catenin signaling via Pi3k/Akt axis. Int. J. Biochem. Cell Biol. 2017, 89, 85–94. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, P.; Yin, W.; Ji, Y.; Shen, Q.; Ni, Q. Nectin-4 promotes gastric cancer progression via the PI3K/AKT signaling pathway. Hum. Pathol. 2018, 72, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Hao, R.-T.; Zheng, C.; Wu, C.-Y.; Xia, E.-J.; Zhou, X.-F.; Quan, R.-D.; Zhang, X.-H. NECTIN4 promotes papillary thyroid cancer cell proliferation, migration, and invasion and triggers EMT by activating AKT. Cancer Manag. Res. 2019, 11, 2565–2578. [Google Scholar] [CrossRef]
- Villanueva, J.; Vultur, A.; Lee, J.T.; Somasundaram, R.; Fukunaga-Kalabis, M.; Cipolla, A.K.; Wubbenhorst, B.; Xu, X.; Gimotty, P.A.; Kee, D.; et al. Acquired resistance to BRAF inhibitors mediated by a RAF kinase switch in melanoma can be overcome by co-targeting MEK and IGF-1R/PI3K. Cancer Cell 2010, 18, 683–695. [Google Scholar] [CrossRef]
- Sosman, J.A.; Kim, K.B.; Schuchter, L.; Gonzalez, R.; Pavlick, A.C.; Weber, J.S.; McArthur, G.A.; Hutson, T.E.; Moschos, S.J.; Flaherty, K.T.; et al. Survival in BRAF V600-mutant advanced melanoma treated with vemurafenib. N. Engl. J. Med. 2012, 366, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Flaherty, K.T.; Infante, J.R.; Daud, A.; Gonzalez, R.; Kefford, R.F.; Sosman, J.; Hamid, O.; Schuchter, L.; Cebon, J.; Ibrahim, N.; et al. Combined BRAF and MEK inhibition in melanoma with V600 mutations. N. Engl. J. Med. 2012, 367, 1694–1703. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Uchi, H.; Ito, T.; Furue, M. Indirubin-pregnane X receptor-JNK axis accelerates skin wound healing. Sci. Rep. 2019, 9, 18174. [Google Scholar] [CrossRef]
- Ito, T.; Kaku-Ito, Y.; Murata, M.; Furue, K.; Shen, C.-H.; Oda, Y.; Furue, M. Immunohistochemical BRAF V600E expression and intratumor BRAF V600E heterogeneity in acral melanoma: Implication in melanoma-specific survival. J. Clin. Med. 2020, 9, 690. [Google Scholar] [CrossRef]
- Ito, T.; Kaku-Ito, Y.; Murata, M.; Ichiki, T.; Kuma, Y.; Tanaka, Y.; Ide, T.; Ohno, F.; Wada-Ohno, M.; Yamada, Y.; et al. Intra- and inter-tumor BRAF heterogeneity in acral melanoma: An immunohistochemical analysis. Int. J. Mol. Sci. 2019, 20, 6191. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Number | |
|---|---|---|
| Age | Mean | 63.7 |
| Median | 69 | |
| Range | 16–88 | |
| Sex | Male | 55 (43.7%) |
| Female | 71 (56.3%) | |
| Tumor site | Head and neck | 12 (9.5%) |
| Trunk | 5 (4.0%) | |
| Upper limb | 3 (2.4%) | |
| Hand | 25 (19.8%) | |
| Lower limb | 7 (5.6%) | |
| Foot | 74 (58.7%) | |
| Histopathological subtype | ALM | 94 (74.6%) |
| SSM | 15 (11.9%) | |
| LLM | 10 (7.9%) | |
| NM | 7 (5.6%) | |
| Ulceration | Present | 52 (41.3%) |
| Absent | 70 (55.6%) | |
| Unknown | 4 (3.2%) | |
| T category | Tis | 29 (23.0%) |
| T1 | 20 (15.9%) | |
| T2 | 13 (10.3%) | |
| T3 | 21 (16.7%) | |
| T4 | 41 (32.5%) | |
| Unknown | 2 (1.6%) | |
| N category | N0 | 85 (67.5%) |
| N1 | 15 (11.9%) | |
| N2 | 11 (8.7%) | |
| N3 | 14 (11.1%) | |
| Unknown | 1 (0.8%) | |
| M category | M0 | 115 (91.3%) |
| M1 | 11 (8.7%) | |
| VE1 staining | Positive | 38 (30.2%) |
| Negative | 88 (69.8%) | |
| Parameter | NECTIN4 Expression | p-Value | ||
|---|---|---|---|---|
| Low | High | |||
| Age | <70 | 46 | 25 | >0.999 |
| ≥70 | 35 | 20 | ||
| Sex | Male | 35 | 20 | >0.999 |
| Female | 46 | 25 | ||
| Histopathological subtype | ALM | 62 | 32 | 0.527 |
| Others | 19 | 13 | ||
| Ulceration | Present | 33 | 19 | 0.849 |
| Absent | 46 | 24 | ||
| T category | Tis, T1, T2 | 39 | 20 | >0.999 |
| T3, T4 | 42 | 23 | ||
| N category | N0 | 55 | 30 | 0.843 |
| N1–3 | 25 | 15 | ||
| M category | M0 | 74 | 41 | >0.999 |
| M1 | 7 | 4 | ||
| VE1 staining | Positive | 16 | 22 | 0.0011 * |
| Negative | 65 | 23 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, Y.; Murata, M.; Shen, C.-H.; Furue, M.; Ito, T. NECTIN4: A Novel Therapeutic Target for Melanoma. Int. J. Mol. Sci. 2021, 22, 976. https://doi.org/10.3390/ijms22020976
Tanaka Y, Murata M, Shen C-H, Furue M, Ito T. NECTIN4: A Novel Therapeutic Target for Melanoma. International Journal of Molecular Sciences. 2021; 22(2):976. https://doi.org/10.3390/ijms22020976
Chicago/Turabian StyleTanaka, Yuka, Maho Murata, Che-Hung Shen, Masutaka Furue, and Takamichi Ito. 2021. "NECTIN4: A Novel Therapeutic Target for Melanoma" International Journal of Molecular Sciences 22, no. 2: 976. https://doi.org/10.3390/ijms22020976
APA StyleTanaka, Y., Murata, M., Shen, C.-H., Furue, M., & Ito, T. (2021). NECTIN4: A Novel Therapeutic Target for Melanoma. International Journal of Molecular Sciences, 22(2), 976. https://doi.org/10.3390/ijms22020976