2.1. Net Charge Is Not Enough to Explain Phosphorylation Induced Changes
Atomistic simulations of five different disordered peptides in both non-phosphorylated and phosphorylated state, shown in
Table 1, have been performed at conditions corresponding to physiological pH (approximately pH 7). The peptides were chosen based on the availability of experimental data and their size, considering computational expense.
SN15, Tau2, and bCPP all contract upon phosphorylation, as shown from the peak shift towards lower values of the distributions of radius of gyration (R
g) and end-to-end distance (R
ee) in
Figure 1, as well as the average values of R
g and R
ee presented in
Table 2. For SN15 and Tau2, the width of the distribution also decreases, while bCPP keeps the same range, only the shape of the distribution changes. Stath and Tau1 both expand, shown from a peak shift towards larger values in the distributions. For Tau1, the expansion is more clear observing the R
g distribution than the R
ee distribution, which only changes shape by the disappearance of a shoulder at lower values. This, however, causes the average R
ee, presented in
Table 2, to increase. An increase of R
ee upon phosphorylation of Tau1 has been detected by fluorescence resonance energy transfer measurements, as reported by Chin et al. [
15].
The shape factor, presented in
Figure 2, can be used as an estimate of the shape of the peptide. If it behaves as a Gaussian coil, the shape factor is approximately 6, whereas for a stiff rod, it is around 12. SN15, Tau2, and bCPP are shown to behave rather coil-like in non-phosphorylated state, while Tau1 is more stiff, and Stath more contracted. Upon phosphorylation, bCPP becomes more contracted than a Gaussian coil, while Stath expands to become more coil-like.
Comparing the induced changes of R
g and R
ee with the net charge of the non-phosphorylated peptides, it is clear that the prediction of Jin and Gräter, i.e., that net charge controls the effect of phosphorylation [
10], only holds for SN15, Tau2, and Stath. bCPP contracts despite having a negative net charge, and Tau1 expands despite the positive net charge. Note that the peptides in this study are distinctly shorter (11–43 residues) compared to the IDPs in the study by Jin and Gräter (approximately 80 residues) [
10], hence local interactions are expected to have a more direct effect on the global dimensions. To understand the effect of phosphorylation of these peptides, we therefore need to investigate changes in secondary structure and specific interactions.
2.2. Phosphorylation of Tau1 Favors Expanded Conformations
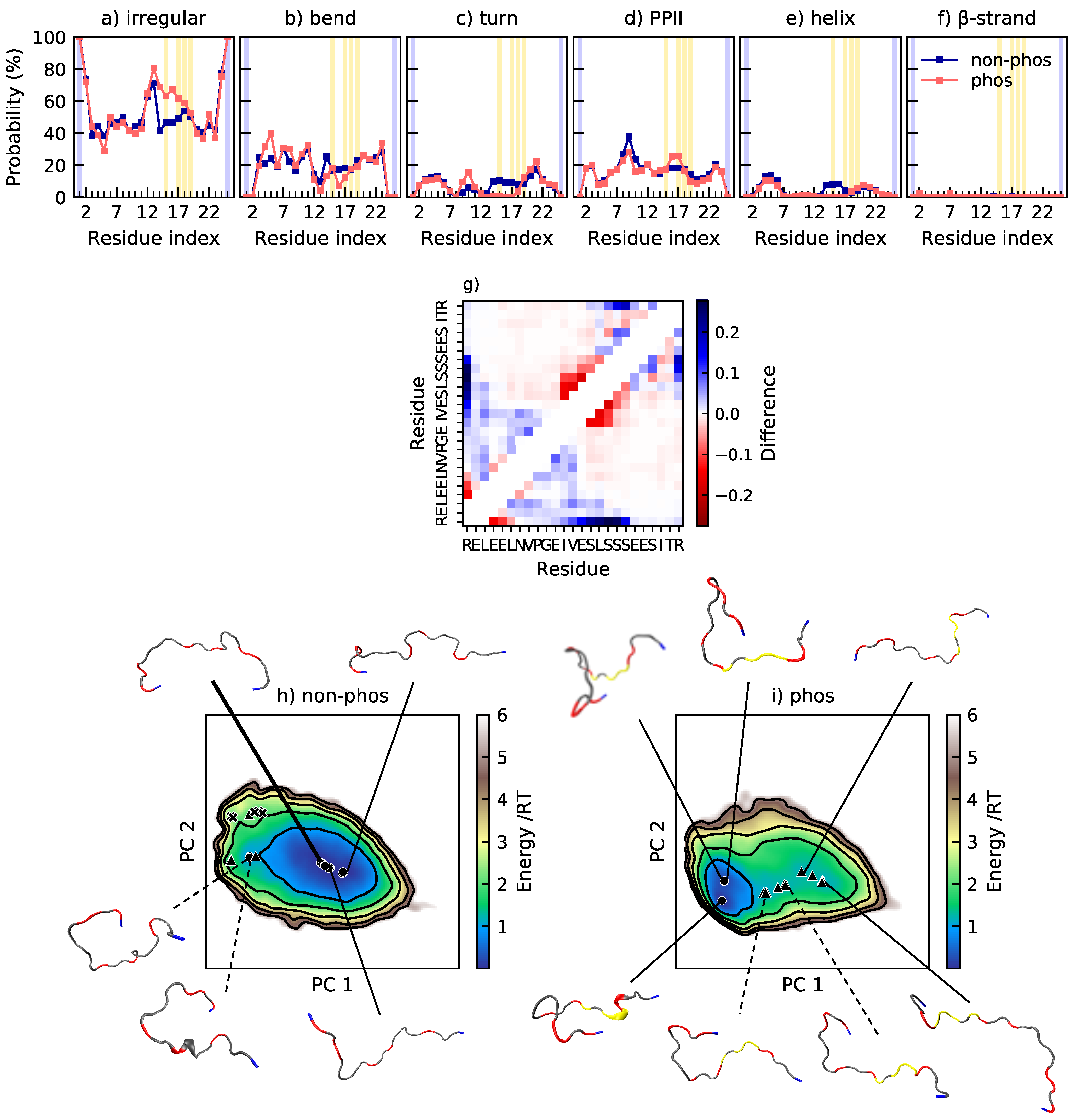
Tau1 is dominated by irregular structure and polyproline type II helix (PPII), as shown in
Figure 3a–f. It possesses 46% and 51% PPII in the non-phosphorylated and phosphorylated state, respectively. Elam et al. [
41] have predicted close to 50% PPII content in this region of Tau, and CD measurements of this segment indicate an increase of PPII content upon phosphorylation [
15]. In
Figure 3a–f, it is shown that all structural changes upon phosphorylation at T175 and T181 take place at the C-terminal end of the peptide, from residue 179 and forward. The propensity for bends and turns at residue 179–181 decreases, while the PPII content increases at residues 181–182. There is occasional salt bridge formation between the phosphothreonines and their respective neighboring lysine. Specifically, the probability of salt bridge formation is
% for pT175–K174 and
% for pT181–K180. The most occurring salt bridge is, however, formed between pT175 and the N-terminal, with a probability of
%. However, due to the close proximity between the salt bridging residues, the effect on the overall dimensions of the peptide is small. Since Tau1 is a short and rather stiff peptide, as shown by the shape factor in
Figure 2, there is limited contact between residues. The change in contact probability upon phosphorylation is also small, according to
Figure 3g, which reveals that the main change is a decrease of contact between T181 and the preceding residues A177 and P178, in agreement with the decreased probability of a bend or turn in that region, as shown by
Figure 3b,c. The conformational effects of phosphorylation of Tau are well summarized by
Figure 3h,i, showing the energy landscape and conformations of non-phosphorylated and phosphorylated Tau1. The energy landscape of non-phosphorylated Tau1 contains several minima, of which the minimum containing expanded conformations dominate, in line with the relatively high shape factor. Other less populated minima contain conformations with a kink in the C-terminal end, originating from a bend or turn. Upon phosphorylation, the minima with kinked conformations disappears, leaving only the minima with expanded conformations. This is in line with decreased contact probability and explains the change in shape of the R
g and R
ee distributions, from a peak with a preceding shoulder to a single peak.
2.3. Phosphorylation Increases the Helix Propensity and Induces Salt Bridge Formation in Tau2 and SN15
Tau2 and SN15 are both mainly irregular and report an increase of helicity upon phosphorylation, see
Figure 4a–f and
Figure 5a–f, respectively. The helical region is identified as “pSpSAKSR” in Tau2 and “pSpSEEKFLR” in SN15, according to
Figure 4e and
Figure 5e. The sequences, hence, share two characteristics: (1) the helical region starts with two phoshorylation sites, and (2) three or four steps away from the phosphorylation site, a positively charged residue is positioned. Phosphorylation has been shown to stabilize α-helices if the phosphorylation site is located in the N-terminal end of the helix, by electrostatic interaction between phosphorylated serines and the macrodipole of the helix, and by hydrogen bonding with the amide backbone [
42]. With a
spacing between a phosphorylated serine and a lysine, phosphorylation also stabilizes α-helices through salt bridge formation between the side groups [
43].
For Tau2, a phosphorylation-induced increase of α-helical structure from 5 to 40% in region A239–R242 has been reported [
13]. In these simulations, the main helical increase upon phosphorylation is associated with region S237–K240, where the increase is from 4 to 26%. However, the helical increase is mainly due to 3
10-helix, since the increase of α-helix is only from 1 to 5%. Hence, the simulations are in qualitative agreement with the experiments, but the quantitative results should be treated with caution. In addition, in SN15, the larger part of the helical increase is due to 3
10-helix, and an increase of α-helix is supported by CD spectroscopy [
20], once again giving qualitative support to the findings in this study. Notice also that, while it is hard to make quantitative comparisons with CD data, our study on SN15 suggested that the simulations underestimate the structural content [
20], which is the same as observed for Tau2.
While helix formation decreases the R
g and R
ee, salt bridge formation can also contribute to the compaction observed upon phosphorylation. In Tau2, several salt bridges have been established from NMR measurements, specifically pT231–R230, pS237–K240, and pS238–R242 [
13]. pT231–R230 and pS238–R242 are indeed the two most occurring salt bridges according to
Table 3, while pS237–R242 is the third most common. Apart from the increase of helical content related to phosphorylation,
Figure 4b reveals an interesting pattern of bends after phosphorylation, where the charged residues R, K, pT, and pS are enriched in bends. The conformations in
Figure 4 illustrate how the salt bridges contribute to the formation of bends. Since the probability of a turn at A227–V229 is roughly the same as the probability of the pT231–K225 salt bridge (see
Figure 3 and
Figure 4c), and V228 is located right between K225 and pT231, we conclude that this turn is also a result of a salt bridge interaction. Hence, this peptide shows that salt bridge formation can induce bends and turns.
Comparing the energy landscapes of non-phosphorylated and phosphorylated Tau2 in
Figure 4h,i, it is shown that, for both peptides, more extended conformations, such as in the minima furthest to the right, are sampled, but to a different extent. These type of conformations are more common in the non-phosphorylated variant, while the most populated basin contains conformations with the N-terminal end folded over, to come closer to the phosphorylated residues. While K225 rarely involves in a proper salt bridge with other residues than pT231, it is still energetically favorable to be in rather close vicinity of the phosphorylated region, considering both the charged side chain and the N-terminus. These types of conformations give rise to an increased contact probability within the N-terminal part of the chain, see
Figure 4g. The increased contact probability close to the diagonal in the middle to C-terminal end corresponds to the increase of helical structure and certain salt bridges. Apart from those, there is a decrease of the probability of contacts within the C-terminal end upon phosphorylation. The two minima in the left part of the energy landscape of non-phosphorylated Tau2 in
Figure 4h are examples of conformations with a higher level of contact within the C-terminal end. They originate from the electrostatic attraction between the C-terminus and the positively charged residues. In phosphorylated Tau2, that region of the energy landscape is visited much less (see
Figure 4i), in agreement with the changes in contact probability. Notice, however, that the probability of conformations with one end folded over is much higher after phosphorylation, which explains the decrease in R
g and R
ee. The conformation corresponding to the minimum in the most populated basin for the phosphorylated peptide additionally shows a helix in the C-terminal end, which also contributes to a decreased R
g and R
ee.
In SN15, the salt bridges pS2–K6, pS3–K6, pS3–R9, and pS3–R10 are the most probable and all form with an approximately 25% occurrence. From the change in contact probability displayed in
Figure 5g, it appears that the pS2–K6 and pS3–K6 salt bridges contribute to stabilize the formed helix. The pS3–R9 and pS3–R10 salt bridges are also visible in the contact map and contribute to an increase in the amount of more compact conformations after phosphorylation. In the energy landscape in
Figure 5, it is shown that phosphorylation shifts the position of the main minima in the energy landscape, from an area of more coil-like structures to a more compact state. The non-phosphorylated peptide also samples conformations that are more compact with a higher content of secondary structure, but more rarely than the phosphorylated peptide. The conformation corresponding to the minimum in the most populated basin in the phosphorylated peptide has residue pS2 and K6 close enough to be in contact; however, there is no helix, but instead a turn at residues E4–E5. This shows that it is favorable to have pS2 and K6 in contact, but that the interaction does not necessarily imply helix formation. In
Figure 5c, it was shown that the turn content in region S3–E5 also increases upon phosphorylation, not only the helix content. There is also an increase of turn content in region F7–R11, which is partly caused by occasional β-strand formation, as shown in the other conformation in
Figure 5, and partly by residues pS3 and R9 coming close to form a salt bridge, in line with the turn induced in Tau2. Both of these changes give rise to more compact conformations. We must, however, note that SAXS measurements have indicated that a compaction upon phosphorylation is plausible, but probably smaller than shown in the simulations [
20]. While Jin and Gräter found that changes in the hydration shell upon phosphorylation can hide global conformational changes in SAXS measurements, they also concluded that the force field used in this study overestimates the charge effect, thus providing two different explanations of the deviations between the simulations and experiments [
10]. Note also that the contact map reports a decrease of contact between R10 and F14, a contact probably formed due to cation–π interaction, which will be discussed further in the section regarding Stath.
2.4. Salt Bridge Formation Shifts the Conformational Ensemble of bCPP
For bCPP, the secondary structure content is dominated by an irregular structure and is highly similar in phosphorylated and non-phosphorylated states, as shown by
Figure 6a–f, in agreement with CD spectroscopy results by Farrell et al. [
25]. The small difference that occurs upon phosphorylation at S14, S17, S18, and S19 is a change from helix and turn to irregular structure in region E14–S17. The vanishing of helical content is in agreement with the conclusion of Andrew et al. that phosphorylation of a residue in the interior of a helix, without a positively charged residue within suitable distance, destabilizes the helix [
42]. Since disruption of a short helix would not cause a contraction of the peptide, the conformational changes in bCPP upon phosphorylation are not explained by secondary structure. Instead, the contraction is due to electrostatic attraction including salt bridge formation between the positively charged end residues and the phosphorylated residues, as seen in
Table 4. Although both end residues are arginines, there is a preference of R1 to interact with the phosphorylated region over R25, due to the respective charges of the termini. This is evident from the fact that the N-terminus is also involved in salt bridges with the phosphorylated residues, and further shown by the difference in contact probability in
Figure 6g. When R1 interacts with the phosphorylated residues, it causes the peptide to fold over, reducing R
g and R
ee substantially. From the energy landscapes in
Figure 6h,i, it is shown that before phosphorylation the minima with lowest energy contain more extended conformations, while after phosphorylation the minima with lowest energy instead showcase the N-terminal part being folded over.
Based only on the net charge of non-phosphorylated bCPP, it was expected that it would expand upon phosphorylation. Considering only region E13–E21, which contains the four phosphorylation sites, this effect was noticed. The average distance between the C atoms of residue 13 and 21 increases from nm to nm upon phosphorylation. However, due to the strong electrostatic interaction between the arginines and the phosphorylated region that are far apart in the sequence, the global result is compaction. Hence, the relative position of charged residues is very important to consider for the effects of phosphorylation on the overall dimensions of the peptide.
We previously showed that the addition of 150 mM NaCl had negligible effects on the salt bridges and global conformational properties of phosphorylated bCPP [
31]. The same applies to non-phosphorylated bCPP, as presented in
Supplementary File S1, Figures S1 and S2. However, although the average values of R
g at 0 and 150 mM are within error, there is a slight increase in the phosphorylated variant and decrease in the non-phosphorylated variant, see
Table 5. Hence, at 150 mM NaCl, the difference observed in R
g between the two variants vanishes, considering the associated error. Note, however, that the distributions still have distinctly different shapes, hence we argue that the conformational ensembles are still different. The same trend is observed in the average R
ee values, although a difference with respect to phosphorylation state still remains at 150 mM NaCl, see
Table 5. In addition, in the calculated scattering curve (
Supplementary File S1, Figure S2), the effect of salt is smaller than the effect of phosphorylation. The difference between the form factor of non-phosphorylated and phosphorylated bCPP is, however, still rather small, so we suspect that it can be hard to detect experimentally with SAXS. Based on the fraction of charged residues and level of charge separation, we expect the other peptides in this study to show smaller effects in regard to salt concentration than bCPP. Hence, we expect the results observed here to be also valid at 150 mM NaCl.
2.5. Arginine—Phosphoserine Interactions Outshines Arginine—Tyrosine Interactions in Stath
Upon phosphorylation of Stath, the three largest changes in secondary structure are a decrease of β-strand structure, an increase of helical structure, and an increase of turns, according to
Figure 7a–f. The increase of helical structure is in the same region as observed for the N-terminal fragment SN15.
Figure 7f implies that residues R10, Y18, Y21, and Y41 are of extra importance for the formation of β-sheet. The cation–π interaction that can occur between aromatic residues, such as tyrosine, and cationic residues, such as arginine, have been shown to be common in proteins [
44]. A correlation between β-strands and cation–π interactions have also been established [
45].
Table 6 show that the cation–π interaction indeed is more occurring in non-phosphorylated Stath than in phosphorylated Stath, suggesting that it drives the formation of β-strands. The conformations in
Figure 7I–III show examples of the cation–π interaction in non-phosphorylated Stath. Although the aromatic–cation interactions are more common in non-phosphorylated Stath, they still occur in phosphorylated Stath, as exemplified by
Figure 7. Upon phosphorylation, the occurrence of cation–π interaction decreases substantially, while salt bridge formation appears according to
Table 7. Notice that R10, which was shown to interact with tyrosines, is involved in one of the most probable salt bridges, pS3–R10. Hence, the arginine–phosphoserine interaction is deemed stronger than the arginine–tyrosine interaction. The replacement of arginine–tyrosine interaction with arginine–phosphoserine causes the β-strands to vanish, which explains the observed expansion.
As presented above, SN15, which is the first fifteen residues of Stath, contracts upon phosphorylation, which was explained by the increased helicity and formation of salt bridges.
Supplementary File S1, Figure S3 shows that, in phosphorylated Stath, the global dimensions of the first fifteen residues, Stath
1–15 agree with those of the fragment (SN15). In the non-phosphorylated variant, the distributions are also rather similar, except for a sharp peak in both the R
g and R
ee distributions, which corresponds to a basin in the energy landscape with the conformation shown in
Supplementary File S1, Figure S3c. Regarding the secondary structure, according to
Supplementary File S1, Figure S4, the largest difference between SN15 and Stath
1–15 is caused by β-strand not forming in SN15, due to lacking its partner further on in the sequence. There are also some differences in bends and turns, but the increase of helical propensity is similar. Hence, overall, the first fifteen residues of Stath behave rather similarly in the full peptide and as a standalone fragment, although especially the presence of the rest of the sequence induces β-strand formation. Despite this discrepancy, we can conclude that phosphorylation of Stath causes a contraction of the first fifteen residues, but an expansion of the full peptide, due to disruption of β-sheets.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}