
Atypical Renal Clearance of Nanoparticles Larger Than the Kidney Filtration Threshold



Abstract
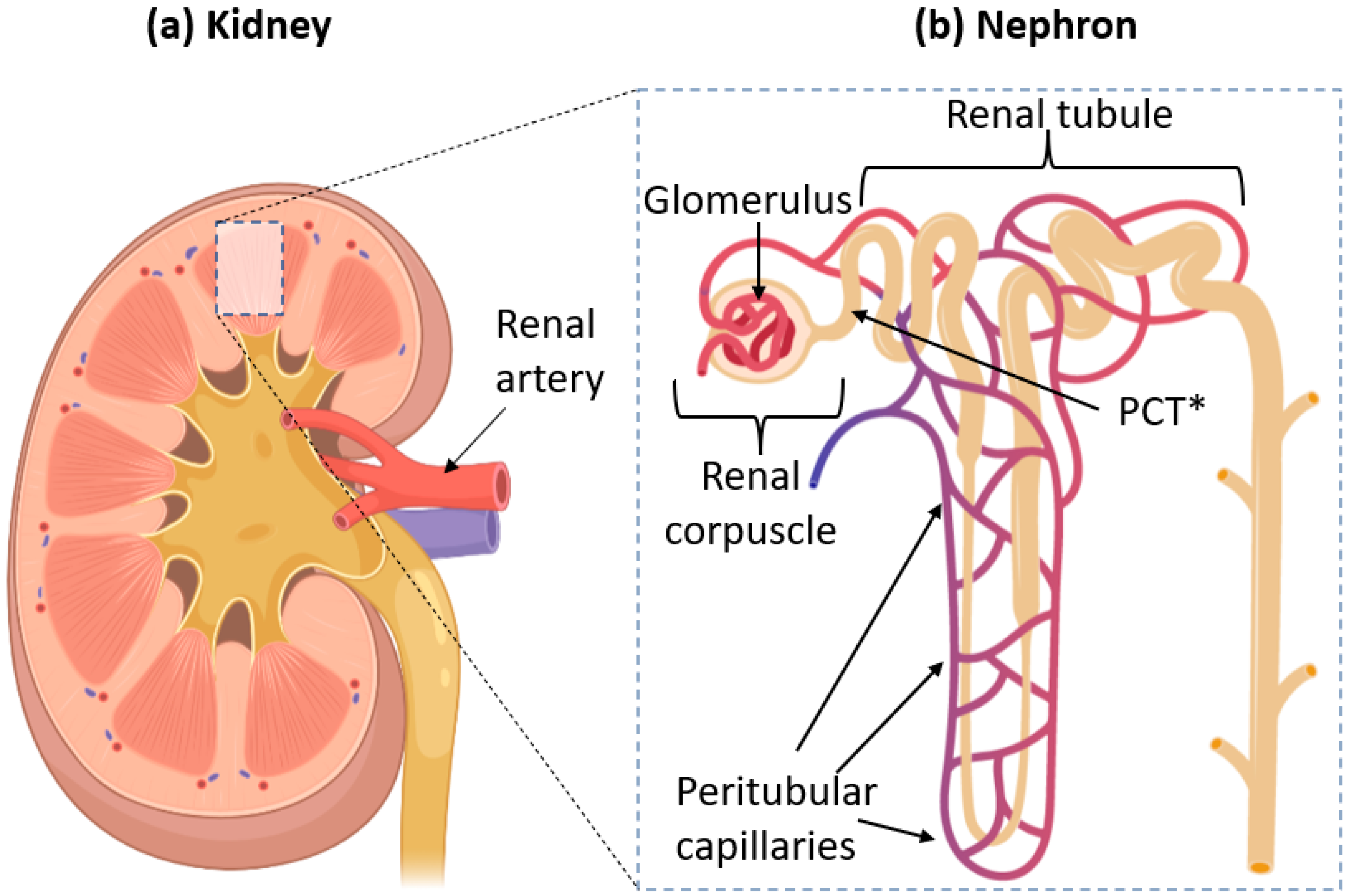
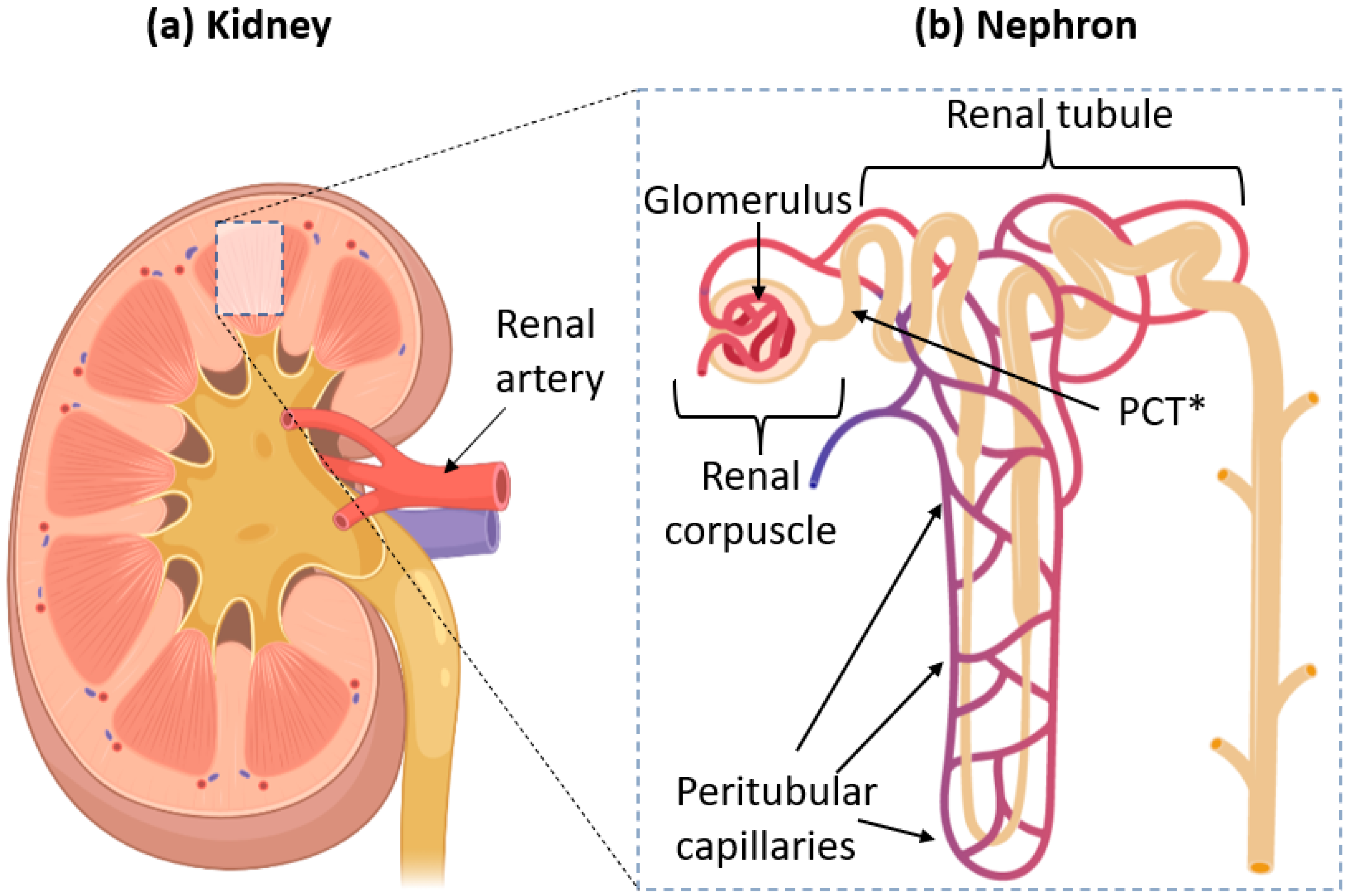
:1. Introduction
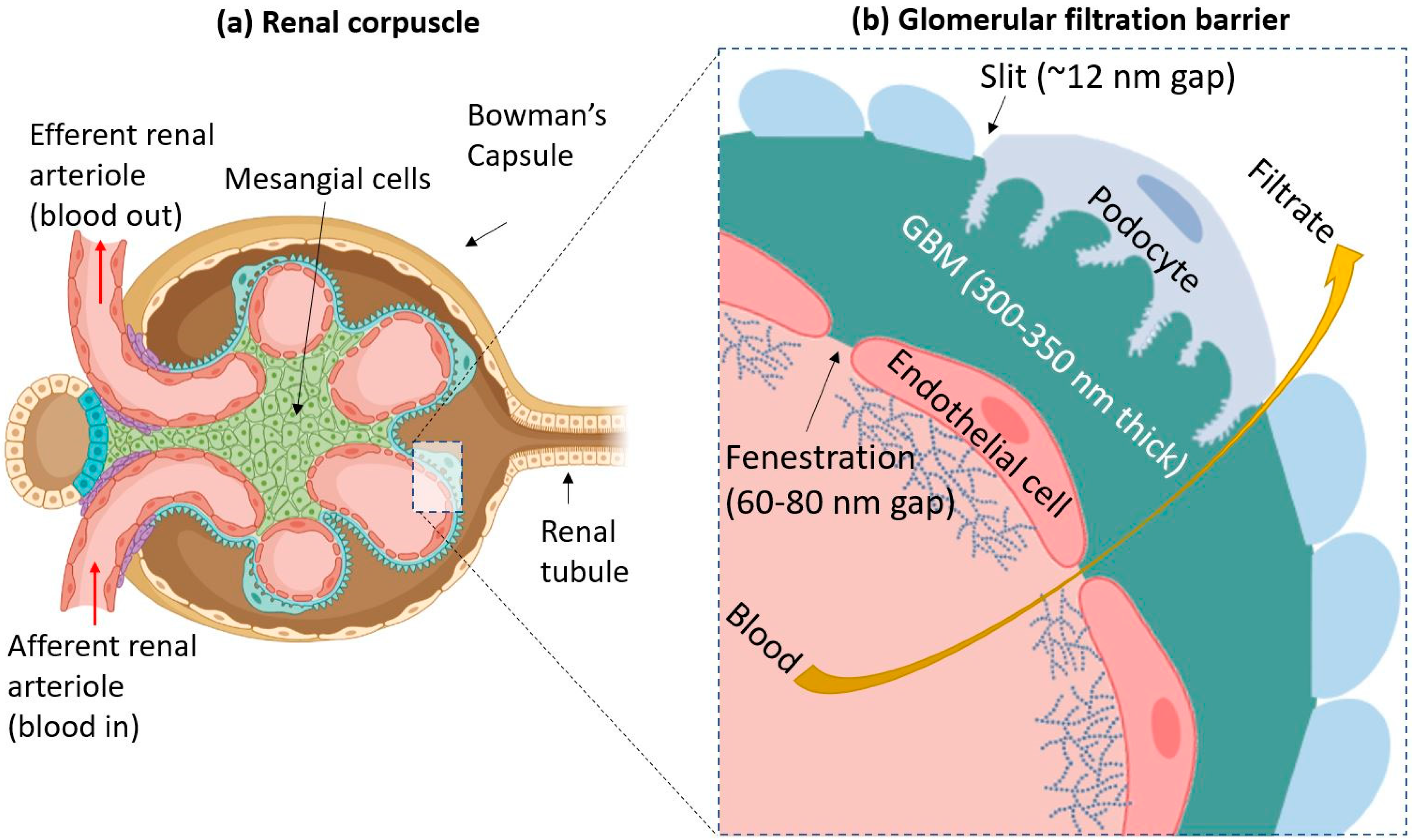
2. The Glomerular Filtration Barrier
3. Renal Clearance through Interactions with the GFB
4. Bypassing the GFB through the Proximal Convoluted Tubules (PCTs)
5. Renal Clearance through an Unknown Route
6. Possible Mechanisms of Renal Clearance
7. NP Physicochemical Properties Which Allow for Renal Excretion
8. The Effect of Nanoparticle Size on Renal Excretion
9. The Effect of Nanoparticle Charge on Renal Excretion
10. The Effect of Nanoparticle Composition on Renal Excretion
11. The Effect of Nanoparticle Surface Modifications on Renal Excretion
12. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, Applications and Toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Vert, M.; Doi, Y.; Hellwich, K.-H.; Hess, M.; Hodge, P.; Kubisa, P.; Rinaudo, M.; Schué, F. Terminology for Biorelated Polymers and Applications (IUPAC Recommendations 2012). Pure Appl. Chem. 2012, 84, 377–410. [Google Scholar] [CrossRef]
- Da Silva, F.L.O.; Marques, M.B.F.; Kato, K.C.; Carneiro, G. Nanonization Techniques to Overcome Poor Water-Solubility with Drugs. Expert Opin. Drug Discov. 2020, 15, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Gunasekaran, T.; Haile, T.; Nigusse, T.; Dhanaraju, M.D. Nanotechnology: An Effective Tool for Enhancing Bioavailability and Bioactivity of Phytomedicine. Asian Pac. J. Trop. Biomed. 2014, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Desai, P.P.; Date, A.A.; Patravale, V.B. Overcoming Poor Oral Bioavailability Using Nanoparticle Formulations-Opportunities and Limitations. Drug Discov. Today Technol. 2012, 9, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Morgen, M.; Bloom, C.; Beyerinck, R.; Bello, A.; Song, W.; Wilkinson, K.; Steenwyk, R.; Shamblin, S. Polymeric Nanoparticles for Increased Oral Bioavailability and Rapid Absorption Using Celecoxib as a Model of a Low-Solubility, High-Permeability Drug. Pharm. Res. 2012, 29, 427–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering Precision Nanoparticles for Drug Delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Rosenblum, D.; Joshi, N.; Tao, W.; Karp, J.M.; Peer, D. Progress and Challenges Towards Targeted Delivery of Cancer Therapeutics. Nat. Commun. 2018, 9, 1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Yu, J.; Lu, X.; He, X. Nanoparticle Systems Reduce Systemic Toxicity in Cancer Treatment. Nanomedicine 2015, 11, 103–106. [Google Scholar] [CrossRef]
- Paris, J.L.; Baeza, A.; Vallet-Regi, M. Overcoming the Stability, Toxicity, and Biodegradation Challenges of Tumor Stimuli-Responsive Inorganic Nanoparticles for Delivery of Cancer Therapeutics. Expert Opin. Drug Deliv. 2019, 16, 1095–1112. [Google Scholar] [CrossRef] [PubMed]
- US-FDA. Nanotechnology-Over a Decade of Progress and Innovation; FDA: Silver Spring, MD, USA, 2020. [Google Scholar]
- Ventola, C. Progress in Nanomedicine: Approved and Investigational Nanodrugs. Pharm. Ther. 2017, 42, 742–755. [Google Scholar]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the Clinic: An Update. Bioeng. Transl. Med. 2019, 4, e10143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dri, D.A.; Marianecci, C.; Carafa, M.; Gaucci, E.; Gramaglia, D. Surfactants, Nanomedicines and Nanocarriers: A Critical Evaluation on Clinical Trials. Pharmaceutics 2021, 13, 381. [Google Scholar] [CrossRef] [PubMed]
- Desai, N. Challenges in Development of Nanoparticle-Based Therapeutics. AAPS J. 2012, 14, 282–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, S.; de Matos, M.B.C.; Metselaar, J.M.; Storm, G. Current Trends and Challenges in the Clinical Translation of Nanoparticulate Nanomedicines: Pathways for Translational Development and Commercialization. Front. Pharmacol. 2018, 9, 790. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.N.; Poon, W.; Tavares, A.J.; McGilvray, I.D.; Chan, W.C.W. Nanoparticle-Liver Interactions: Cellular Uptake and Hepatobiliary Elimination. J. Control. Release 2016, 240, 332–348. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, J.; Jiang, K.; Chung, E.J. Improving Kidney Targeting: The Influence of Nanoparticle Physicochemical Properties on Kidney Interactions. J. Control. Release 2021, 334, 127–137. [Google Scholar] [CrossRef]
- Simkó, M.; Mattson, M.-O. Interactions Between Nanosized Materials and the Brain. Curr. Med. Chem. 2014, 21, 4200–4214. [Google Scholar] [CrossRef] [Green Version]
- Huai, Y.; Hossen, M.N.; Wilhelm, S.; Bhattacharya, R.; Mukherjee, P. Nanoparticle Interactions with the Tumor Microenvironment. Bioconjug. Chem. 2019, 30, 2247–2263. [Google Scholar] [CrossRef]
- Du, B.; Yu, M.; Zheng, J. Transport and Interactions of Nanoparticles in the Kidneys. Nat. Rev. Mater. 2018, 3, 358–374. [Google Scholar] [CrossRef]
- Rampado, R.; Crotti, S.; Caliceti, P.; Pucciarelli, S.; Agostini, M. Recent Advances in Understanding the Protein Corona of Nanoparticles and in the Formulation of “Stealthy” Nanomaterials. Front. Bioeng. Biotechnol. 2020, 8, 166. [Google Scholar] [CrossRef]
- Park, S.J. Protein-Nanoparticle Interaction: Corona Formation and Conformational Changes in Proteins on Nanoparticles. Int. J. Nanomed. 2020, 15, 5783–5802. [Google Scholar] [CrossRef]
- Zhang, A.; Meng, K.; Liu, Y.; Pan, Y.; Qu, W.; Chen, D.; Xie, S. Absorption, Distribution, Metabolism, and Excretion of Nanocarriers In Vivo and Their Influences. Adv. Colloid Interface Sci. 2020, 284, 102261. [Google Scholar] [CrossRef]
- Corbo, C.; Molinaro, R.; Parodi, A.; Furman, N.E.T. The Impact of Nanoparticle Protein Corona on Cytotoxicity, Immunotoxicity and Target Drug Delivery. Nanomedicine 2015, 11, 81–100. [Google Scholar] [CrossRef] [Green Version]
- Tekie, F.S.M.; Hajiramezanali, M.; Geramifar, P.; Raoufi, M.; Dinarvand, R.; Soleimani, M.; Atyabi, F. Controlling Evolution of Protein Corona: A Prosperous Approach to Improve Chitosan-Based Nanoparticle Biodistribution and Half-Life. Sci. Rep. 2020, 10, 9664. [Google Scholar] [CrossRef]
- Li, S.D.; Huang, L. Nanoparticles Evading the Reticuloendothelial System: Role of the Supported Bilayer. Biochim. Biophys. Acta 2009, 1788, 2259–2266. [Google Scholar] [CrossRef] [Green Version]
- Watermann, A.; Brieger, J. Mesoporous Silica Nanoparticles as Drug Delivery Vehicles in Cancer. Nanomaterials 2017, 7, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.Y.; Kim, H.S.; Palanikumar, L.; Go, E.M.; Jana, B.; Park, S.A.; Kim, H.Y.; Kim, K.; Seo, J.K.; Kwak, S.K.; et al. Cloaking Nanoparticles with Protein Corona Shield for Targeted Drug Delivery. Nat. Commun. 2018, 9, 4548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loiola, L.M.D.; Batista, M.; Capeletti, L.B.; Mondo, G.B.; Rosa, R.S.M.; Marques, R.E.; Bajgelman, M.C.; Cardoso, M.B. Shielding and Stealth Effects of Zwitterion Moieties in Double-Functionalized Silica Nanoparticles. J. Colloid Interface Sci. 2019, 553, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, G. Imaging Nano-Bio Interactions in the Kidney: Toward a Better Understanding of Nanoparticle Clearance. Angew. Chem. Int. 2018, 57, 3008–3010. [Google Scholar] [CrossRef] [PubMed]
- Menon, M.C.; Chuang, P.Y.; He, C.J. The Glomerular Filtration Barrier: Components and Crosstalk. Int. J. Nephrol. 2012, 2012, 749010. [Google Scholar] [CrossRef] [Green Version]
- Fissell, W.H.; Miner, J.H. What Is the Glomerular Ultrafiltration Barrier? J. Am. Soc. Nephrol. 2018, 29, 2262–2264. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, M.G.; Altenburg, M.K.; Sanford, R.; Willett, J.D.; Bleasdale, B.; Ballou, B.; Wilder, J.; Li, F.; Miner, J.H.; Berg, U.B.; et al. Permeation of Macromolecules into the Renal Glomerular Basement Membrane and Capture by the Tubules. Proc. Natl. Acad. Sci. USA 2017, 114, 2958–2963. [Google Scholar] [CrossRef] [Green Version]
- Naumenko, V.; Nikitin, A.; Kapitanova, K.; Melnikov, P.; Vodopyanov, S.; Garanina, A.; Valikhov, M.; Ilyasov, A.; Vishnevskiy, D.; Markov, A.; et al. Intravital Microscopy Reveals a Novel Mechanism of Nanoparticles Excretion in Kidney. J. Control. Release 2019, 307, 368–378. [Google Scholar] [CrossRef]
- Dogra, P.; Adolphi, N.L.; Wang, Z.; Lin, Y.S.; Butler, K.S.; Durfee, P.N.; Croissant, J.G.; Noureddine, A.; Coker, E.N.; Bearer, E.L.; et al. Establishing the Effects of Mesoporous Silica Nanoparticle Properties on In Vivo Disposition Using Imaging-based Pharmacokinetics. Nat. Commun. 2018, 9, 4551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Campaña, C.; Gómez-Vallejo, V.; Puigivila, M.; Martín, A.; Calvo-Fernández, T.; Moya, S.E.; Ziolo, R.F.; Reese, T.; Llop, J. Biodistribution of Different Sized Nanoparticles Assessed by Positron Emission Tomography: A General Strategy for Direct Activation of Metal Oxide Particles. ACS Nano 2013, 7, 3498–3505. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.W.; Pippin, J.W.; Eng, D.G.; Lv, S.; Shankland, S.J.; Pun, S.H. Nanoparticles Exhibit Greater Accumulation in Kidney Glomeruli during Experimental Glomerular Kidney Disease. Physiol. Rep. 2020, 8, e14545. [Google Scholar] [CrossRef]
- Williams, R.M.; Shah, J.; Ng, B.D.; Minton, D.R.; Gudas, L.J.; Park, C.Y.; Heller, D.A. Mesoscale Nanoparticles Selectively Target the Renal Proximal Tubule Epithelium. Nano Lett. 2015, 15, 2358–2364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, R.M.; Shah, J.; Tian, H.S.; Chen, X.; Geissmann, F.; Jaimes, E.A.; Heller, D.A. Selective Nanoparticle Targeting of the Renal Tubules. Hypertension 2018, 71, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Croissant, J.G.; Fatieiev, Y.; Khashab, N.M. Degradability and Clearance of Silicon, Organosilica, Silsesquioxane, Silica Mixed Oxide, and Mesoporous Silica Nanoparticles. Adv. Mater. 2017, 29, 1604634. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, I.I.; Kapralov, A.A.; Michael, Z.P.; Burkert, S.C.; Shurin, M.R.; Star, A.; Shvedova, A.A.; Kagan, V.E. Enzymatic Oxidative Biodegradation of Nanoparticles: Mechanisms, Significance and Applications. Toxicol. Appl. Pharmacol. 2016, 299, 58–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallabi, P. Some Information about the Morphology and Anatomy of the Human Kidney. J. Morphol. Anat. 2018, 2, 1–4. [Google Scholar]
- Wallace, M.A. Anatomy and Physiology of the Kidney. AORN J. 1998, 68, 799–820. [Google Scholar] [CrossRef]
- Scott, R.P.; Quaggin, S.E. Review Series: The Cell Biology of Renal Filtration. J. Cell Biol. 2015, 209, 199–210. [Google Scholar] [CrossRef]
- Edwards, J.G. The Formation of Urine. Arch. Intern. Med. 1940, 65, 800–824. [Google Scholar] [CrossRef]
- Chmielewsky, C. Renal Anatomy and Overview of Nephron Function. Nephrol. Nurs. J. 2003, 30, 185–190. [Google Scholar]
- Emelianova, Y.P.; Kuznetsov, A.P.; Laugesen, J.L.; Mosekilde, E.; Holstein-Rathlou, N. Oscillator Suppression in the Blood Flow Regulation of Interacting, Non-Identical Nephrons. J. Hypertens. 2013, 3, 1–9. [Google Scholar]
- Bertram, J.F.; Douglas-Denton, R.N.; Diouf, B.; Hughson, M.D.; Hoy, W.E. Human Nephron Number: Implications for Health and Disease. Pediatr. Nephrol. 2011, 26, 1529–1533. [Google Scholar] [CrossRef] [PubMed]
- Robinson, P.G.; Newman, D.; Reitz, C.L.; Vaynberg, L.Z.; Bahga, D.K.; Levitt, M.H. A Large Drawing of a Nephron for Teaching Medical Students Renal Physiology, Histology, and Pharmacology. Adv. Physiol. Educ. 2018, 42, 192–199. [Google Scholar] [CrossRef]
- Pollak, M.R.; Quaggin, S.E.; Hoenig, M.P.; Dworkin, L.D. The Glomerulus: The Sphere of Influence. Clin. J. Am. Soc. Nephrol. 2014, 9, 1461–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlondorff, D.; Banas, B. The Mesangial Cell Revisited: No Cell is an Island. J. Am. Soc. Nephrol. 2009, 20, 1179–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latta, H. An Approach to the Structure and Function of the Glomerular Mesangium. J. Am. Soc. Nephrol. 1992, 2, 65. [Google Scholar] [CrossRef] [PubMed]
- Haraldsson, B.; Jeansson, M. Glomerular Filtration Barrier. Curr. Opin. Nephrol. Hypertens. 2009, 18, 331–335. [Google Scholar] [CrossRef]
- Arif, E.; Nihalani, D. Glomerular Filtration Barrier Assembly: An Insight. Postdoc. J. 2013, 1, 33–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Satchell, S.C.; Neal, C.R.; McKenzie, E.A.; Tooke, J.E.; Mathieson, P.W. Glomerular Endothelial Glycocalyx Constitutes a Barrier to Protein Permeability. J. Am. Soc. Nephrol. 2007, 18, 2885–2893. [Google Scholar] [CrossRef] [Green Version]
- Satchell, S.C.; Braet, F. Glomerular Endothelial Cell Fenestrations: An Integral Component of the Glomerular Filtration Barrier. Am. J. Physiol.-Ren. Physiol. 2009, 296, 947–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satchell, S.C. The Glomerular Endothelium Emerges as a Key Player in Diabetic Nephropathy. Kidney Int. 2012, 82, 949–951. [Google Scholar] [CrossRef] [Green Version]
- Suleiman, H.; Zhang, L.; Roth, R.; Heuser, J.E.; Miner, J.H.; Shaw, A.S.; Dani, A. Nanoscale Protein Architecture of the Kidney Glomerular Basement Membrane. eLife 2013, 2, e01149. [Google Scholar] [CrossRef]
- Miner, J.H. The Glomerular Basement Membrane. Exp. Cell Res. 2012, 318, 973–978. [Google Scholar] [CrossRef] [Green Version]
- Reiser, J.; Lee, H.W.; Gupta, V.; Altintas, M.M. A High-Content Screening Technology for Quantitatively Studying Podocyte Dynamics. Adv. Chronic Kidney Dis. 2017, 24, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Reiser, J.; Altintas, M.M. Podocytes. F1000Research 2016, 5, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Gagliardini, E.; Conti, S.; Benigni, A.; Remuzzi, G.; Remuzzi, A. Imaging of the Porous Ultrastructure of the Glomerular Epithelial Filtration Slit. J. Am. Soc. Nephrol. 2010, 21, 2081–2089. [Google Scholar] [CrossRef] [Green Version]
- Zuckerman, J.E.; Davis, M.E. Targeting Therapeutics to the Glomerulus with Nanoparticles. Adv. Chronic Kidney Dis. 2013, 20, 500–507. [Google Scholar] [CrossRef]
- Fan, K.; Zeng, L.; Guo, J.; Xie, S.; Yu, Y.; Chen, J.; Cao, J.; Xiang, Q.; Zhang, S.; Luo, Y.; et al. Visualized Podocyte-Targeting and Focused Ultrasound Responsive Glucocorticoid Nano-Delivery System against Immune-Associated Nephropathy without Glucocorticoid Side Effect. Theranostics 2021, 11, 2670–2690. [Google Scholar] [CrossRef]
- Ware, T. Nephrotic Syndrome. InnovAiT Educ. Inspir. Gen. Pract. 2020, 13, 159–163. [Google Scholar] [CrossRef]
- Couser, W.G. Primary Membranous Nephropathy. Clin. J. Am. Soc. Nephrol. 2017, 12, 983–997. [Google Scholar] [CrossRef]
- Thomas, M.C.; Brownlee, M.; Susztak, K.; Sharma, K.; Jandeleit-Dahm, K.A.; Zoungas, S.; Rossing, P.; Groop, P.H.; Cooper, M.E. Diabetic Kidney Disease. Nat. Rev. Dis. Primers 2015, 1, 15018. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.H.; Zuckerman, J.E.; Webster, P.; Davis, M.E. Targeting Kidney Mesangium by Nanoparticles of Defined Size. Proc. Natl. Acad. Sci. USA 2011, 108, 6656–6661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushwaha, S.K.S.; Ghoshal, S.; Rai, A.K.; Singh, S. Carbon Nanotubes as a Novel Drug Delivery System for Anticancer Therapy: A Review. Braz. J. Pharm. Sci. 2013, 49, 629–643. [Google Scholar] [CrossRef] [Green Version]
- Ruggiero, A.; Villa, C.H.; Bander, E.; Rey, D.A.; Bergkvist, M.; Batt, C.A.; Manova-Todorova, K.; Deen, W.M.; Scheinberg, D.A.; McDevitt, M.R. Paradoxical Glomerular Filtration of Carbon Nanotubes. Proc. Natl. Acad. Sci. USA 2010, 107, 12369–12374. [Google Scholar] [CrossRef] [Green Version]
- Curthoys, N.P.; Moe, O.W. Proximal Tubule Function and Response to Acidosis. Clin. J. Am. Soc. Nephrol. 2014, 9, 1627–1638. [Google Scholar] [CrossRef] [Green Version]
- Venkatesh, S.; Schrier, R.W.; Andreoli, T.E. Mechanisms of Tubular Sodium Chloride Transport. Ren. Fail. 1998, 20, 783–799. [Google Scholar] [CrossRef] [Green Version]
- Henry, C.; Hartsock, R.J.; Kirk, Z.; Behrer, R. Detection of Viruria in Cytomegalovirus-Infected Infants by Electron Microscopy. Am. J. Clin. Pathol. 1978, 69, 435–439. [Google Scholar] [CrossRef]
- Wyss, P.P.; Lamichhane, S.P.; Abed, A.; Vonwil, D.; Kretz, O.; Huber, T.B.; Sarem, M.; Shastri, V.P. Renal Clearance of Polymeric Nanoparticles by Mimicry of Glycan Surface of Viruses. Biomaterials 2020, 230, 119643. [Google Scholar] [CrossRef]
- Alidori, S.; Akhavein, N.; Thorek, D.L.; Behling, K.; Romin, Y.; Queen, D.; Beattie, B.J.; Manova-Todorova, K.; Bergkvist, M.; Scheinberg, D.A.; et al. Targeted Fibrillar Nanocarbon RNAi Treatment of Acute Kidney Injury. Sci. Transl. Med. 2016, 8, 331ra39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, C.; Liu, T.; Li, L.; Liu, H.; Chen, D.; Tang, F. The Absorption, Distribution, Excretion and Toxicity of Mesoporous Silica Nanoparticles in Mice Following Different Exposure Routes. Biomaterials 2013, 34, 2565–2575. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Nie, H.; Wang, K.; Tan, W.; Wu, X.; Zhang, P. In Vivo Study of Biodistribution and Urinary Excretion of Surface Modified Silica Nanoparticles. Anal. Chem. 2008, 80, 9597–9603. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Li, L.; Liu, T.; Hao, N.; Liu, H.; Chen, D.; Tang, F. The Shape Effect of Mesoporous Silica Nanoparticles on Biodistribution, Clearance, and Biocompatibility In Vivo. ACS Nano 2011, 5, 5390–5399. [Google Scholar] [CrossRef] [PubMed]
- Busher, J.T. Serum Albumin and Globulin. In Clinical Methods: The History, Physical, and Laboratory Examinations, 3rd ed.; Walker, H.K., Hall, W.D., Hurst, J.W., Eds.; Butterworths: Boston, MA, USA, 1990; pp. 497–499. [Google Scholar]
- Tojo, A.; Kinugasa, S. Mechanisms of Glomerular Albumin Filtration and Tubular Reabsorption. Int. J. Nephrol. 2012, 2012, 481520. [Google Scholar] [CrossRef] [Green Version]
- Dickson, L.E.; Wagner, M.C.; Sandoval, R.M.; Molitoris, B.A. The Proximal Tubule and Albuminuria: Really! J. Am. Soc. Nephrol. 2014, 25, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.L.; Chaudhury, C.; Kim, J.; Bronson, C.L.; Wani, M.A.; Mohanty, S. Perspective-FcRn Transports Albumin: Relevance to Immunology and Medicine. Trends Immunol. 2006, 27, 343–348. [Google Scholar] [CrossRef]
- Pallone, T.L.; Cao, C. Renal Cortical and Medullary Microcirculations: Structure and Function. In Seldin and Giebisch’s The Kidney, 4th ed.; Alpern, R.J., Hebert, S.C., Eds.; Elsevier: Amsterdam, The Netherlands, 2008; Volume 1, pp. 627–670. [Google Scholar]
- Dobrinskikh, E.; Okamura, K.; Kopp, J.B.; Doctor, R.B.; Blaine, J. Human Podocytes Perform Polarized, Caveolae-Dependent Albumin Endocytosis. Am. J. Physiol. Ren. Physiol. 2014, 306, 941–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castrop, H.; Schiessl, I.M. Novel Routes of Albumin Passage across the Glomerular Filtration Barrier. Acta Physiol. 2017, 219, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Hoshyar, N.; Gray, S.; Han, H.; Bao, G. The Effect of Nanoparticle Size on In Vivo Pharmacokinetics and Cellular Interaction. Nanomedicine 2016, 11, 673–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, R.M.; Hsu, H.-J.; Bugno, J.; Hong, S. Understanding Nano-Bio Interactions to Improve Nanocarriers for Drug Delivery. MRS Bull. 2014, 39, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Moriyama, T.; Karasawa, K.; Nitta, K. The Role of Caveolae on Albumin Passage through Glomerular Endothelial and Epithelial Cells: The New Etiology of Urinary Albumin Excretion. Contrib. Nephrol. 2018, 195, 1–11. [Google Scholar] [PubMed]
- Longmire, M.; Choyke, P.L.; Kobayashi, H. Clearance Properties of Nano-Sized Particles and Molecules as Imaging Agents: Considerations and Caveats. Nanomedicine 2008, 3, 703–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzanares, D.; Cena, V. Endocytosis: The Nanoparticle and Submicron Nanocompounds Gateway into the Cell. Pharmaceutics 2020, 12, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Chen, M.; Mao, H.; Wang, N.; Zhang, B.; Zhao, X.; Qian, J.; Xing, C. Albumin-Based Nanoparticles as Methylprednisolone Carriers for targeted Delivery towards the Neonatal Fc Receptor in Glomerular Podocytes. Int. J. Mol. Med. 2017, 39, 851–860. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NP System | Size(s) | Renal Clearance Route | Reference |
|---|---|---|---|
| Small interfering RNA and cyclodextrin-containing polymer (siRNA/CDP) NPs | 70 nm | Directly cross the GFB, but accumulate in the GBM | Zuckerman and Davis (2013) [64] |
| Sodium borohydride (NaBH4) NPs with GSH surface modification | 45 kDa | Directly cross the GFB | Lawrence et al. (2017) [34] |
| Sodium thiocyanate (NaSCN) oligoclusters with GSH surface modification | 66 and 300 kDa | Directly cross the GFB | Lawrence et al. (2017) [34] |
| Single-walled carbon nanotubes functionalized with ammonium and siRNA (fCNT/siRNA) | Length 300 nm, diameter around 1 nm | Directly cross the GFB, most of them cleared into urine but some reabsorbed by PCT | Alidori et al. (2016) [76] |
| Nano-delivery system consists of liposome containing PFP and Dex. Surface functionalized with PEG and BMS-α | 190 nm | (Presumably) directly cross GFB, taken up by podocytes | Fan et al. (2021) [65] |
| PEGylated gold nanoparticles (Au-PEG NPs) | 75 ± 25 nm | Cross endothelium and accumulate in mesangium | Choi et al. (2011) [69] |
| Polystyrene PEG-carboxylate NPs | 20 and 100 nm | Accumulate in glomerulus, some in renal tubules | Liu et al. (2020) [38] |
| Iron oxide nanocubes and nanoclusters | 140 nm | Through PCT | Naumenko et al. (2019) [35] |
| Poly(lactic-co-glycolic acid) particles conjugated with PEG (PLGA-PEG) | 350–400 nm | Through PCT | Williams et al. (2018) [40] |
| PLGA-based and they were functionalized with glycosaminoglycan | 130–180 nm | (Presumably) through PCT | Wyss et al. (2019) [75] |
| MSN-PEG functionalized with TMS, PEI or QA | 32–162 nm | Unknown | Dogra et al. (2018) [36] |
| Radioactively-labelled aluminum oxide nanoparticles (13N-labelled Al2O3 NPs) | 10 nm, 40 nm and 150 nm | Unknown | Pérez-Campaña et al. (2013) [37] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adhipandito, C.F.; Cheung, S.-H.; Lin, Y.-H.; Wu, S.-H. Atypical Renal Clearance of Nanoparticles Larger Than the Kidney Filtration Threshold. Int. J. Mol. Sci. 2021, 22, 11182. https://doi.org/10.3390/ijms222011182
Adhipandito CF, Cheung S-H, Lin Y-H, Wu S-H. Atypical Renal Clearance of Nanoparticles Larger Than the Kidney Filtration Threshold. International Journal of Molecular Sciences. 2021; 22(20):11182. https://doi.org/10.3390/ijms222011182
Chicago/Turabian StyleAdhipandito, Christophorus F., Siu-Hung Cheung, Yu-Han Lin, and Si-Han Wu. 2021. "Atypical Renal Clearance of Nanoparticles Larger Than the Kidney Filtration Threshold" International Journal of Molecular Sciences 22, no. 20: 11182. https://doi.org/10.3390/ijms222011182
APA StyleAdhipandito, C. F., Cheung, S.-H., Lin, Y.-H., & Wu, S.-H. (2021). Atypical Renal Clearance of Nanoparticles Larger Than the Kidney Filtration Threshold. International Journal of Molecular Sciences, 22(20), 11182. https://doi.org/10.3390/ijms222011182