1. Introduction
More than 24,500 new cases of the brain & other nervous system tumors and more than 18,500 deaths are estimated to occur in 2021 in the United States [
1]. Brain and other nervous system tumors are the leading cause of cancer death among men aged <40 years and women aged <20 years worldwide [
1]. Among various brain tumor types, glioblastoma (GBM) is the most common, with an incidence of 3.7 per 100,000 persons per year [
2]. It is also the most aggressive primary brain tumor in adults, with an overall 2-year survival of 18%, 3-year survival of 11%, and 5-year survival of only 4% [
2]. Numerous factors implement the high aggressiveness of GBM cells, with the hyperactivated Wnt/β-catenin pathway being one of the most important. Thus, novel approaches to the treatment and control of GBM are urgently needed. A growing number of studies indicates that natural bioactive molecules may serve well in this purpose.
Numerous epidemiological studies demonstrated the relationship between diet and cancer and the potential of dietary components, particularly polyphenols as promising anticancer and chemopreventive agents [
3,
4]. Curcumin, also known as diferuloylmethane (1,7-bis (4-hydroxy-3-methoxyphenyl)-1,6-hepadiene-3,5-dione), is a natural polyphenol extracted from the rhizome of
Curcuma longa. It was used for centuries in traditional Indian Ayurvedic and Chinese medicine for treating respiratory and liver disorders, digestive disorders, infections, allergies, and rheumatisms [
5]. Curcumin has low inherent toxicity and various properties with significant impact and potential medical applications [
6]. It was reported to modulate the activity of kinases and transcription factors as well as to regulate the expression of genes involved in cell survival and malignant transformation [
5]. Numerous studies show that it regulates multiple cellular signaling pathways, including Wnt/β-catenin, PI3K/Akt, JAK/STAT, MAPK, p53 and NF-ĸB signaling pathways [
7]. Curcumin can also epigenetically regulate gene expression by reversing DNA methylation, altering histone modifications, and targeting several miRNAs [
8]. Curcumin can cross the blood-brain barrier, thus yielding therapeutic benefits within the CNS [
9]. However, its impact on GBM remains to be elucidated before curcumin can enter the clinical trial phase as a potential anti-GBM substance.
Sodium butyrate (NaBu), a salt of short chain fatty acid, is one of the main products of the anaerobic fermentation of indigestible polysaccharides such as dietary fiber and resistant starch produced by the microbiota in the large intestine [
10]. It is produced in the lower gastrointestinal tract and was proposed to confer a number of health benefits, including a reduced risk of colorectal cancer [
11,
12]. Moreover, through the bidirectional communication between the gastrointestinal tract and the central nervous system (CNS), NaBu exerts its neuroactive properties [
13]. Animal studies showed that NaBu and other short chain fatty acids exert widespread influence on key neurological and behavioral processes and may be involved in critical phases of neurodevelopmental and neurodegenerative disorders [
13]. It has neuroprotective activity and was shown to be effective against several neuropsychiatric disorders [
14,
15]. Moreover, it was reported to exert antitumor effects in many tumors including not only colorectal cancer [
12], but also breast cancer [
16], and GBM [
17,
18]. The reports evaluating NaBu in the context of GBM cells are however scarce and the issue of whether NaBu can be considered as anti-GBM agent requires further investigation. The bidirectional axis between the intestine and the brain was recently highlighted as an important determinant of glioma biology [
18], making NaBu an interesting agent for investigation. Moreover, according to our best knowledge, up to date, no study shows the effects of the combined treatment with curcumin, or more generally, curcuminoids (CCM) and NaBu on human GBM cells.
Thus, the aim of this study was to analyze the impact of CCM and NaBu on GBM cell viability and to check whether these compounds act synergistically. Other goals were to analyze the role of CCM and NaBu in inducing cell cycle arrest and apoptosis in GBM cells and to verify if the ability to generate reactive oxygen species (ROS) plays a role in these phenomena. The analysis of DNA methylation and expression changes in genes essential in gliomagenesis (Wnt pathway antagonists, SFRP1, RUNX3 and master cell cycle regulator, RASSF1A), and the impact on the expression of the crucial epigenetic regulator UHRF1 were evaluated to further decipher the molecular mechanisms exerted by CCM and NaBu in GBM cells. Finally, to verify if the two compounds can successfully be implemented as a combination treatment targeting CNS, the permeability of CCM and NaBu through the blood-brain barrier (BBB) and the nasal cavity was analyzed.
3. Discussion
GBM is the most malignant and the most common primary brain tumor in adults, with a very poor prognosis. It is characterized by many mechanisms of treatment resistance and inevitable relapse. It shows a number of changes on the epigenetic and genetic level, leading to high invasiveness due to the infiltrative nature of growth, neoangiogenesis, and participation of stem cells resistant to cancer therapy. In addition, GBM can avoid signals leading to apoptosis, and is also characterized by hyperactivation of signaling pathways, such as Wnt/β-catenin pathway, guaranteeing continuous proliferation. Since current GBM treatments are ineffective, finding new therapeutic options are of great importance.
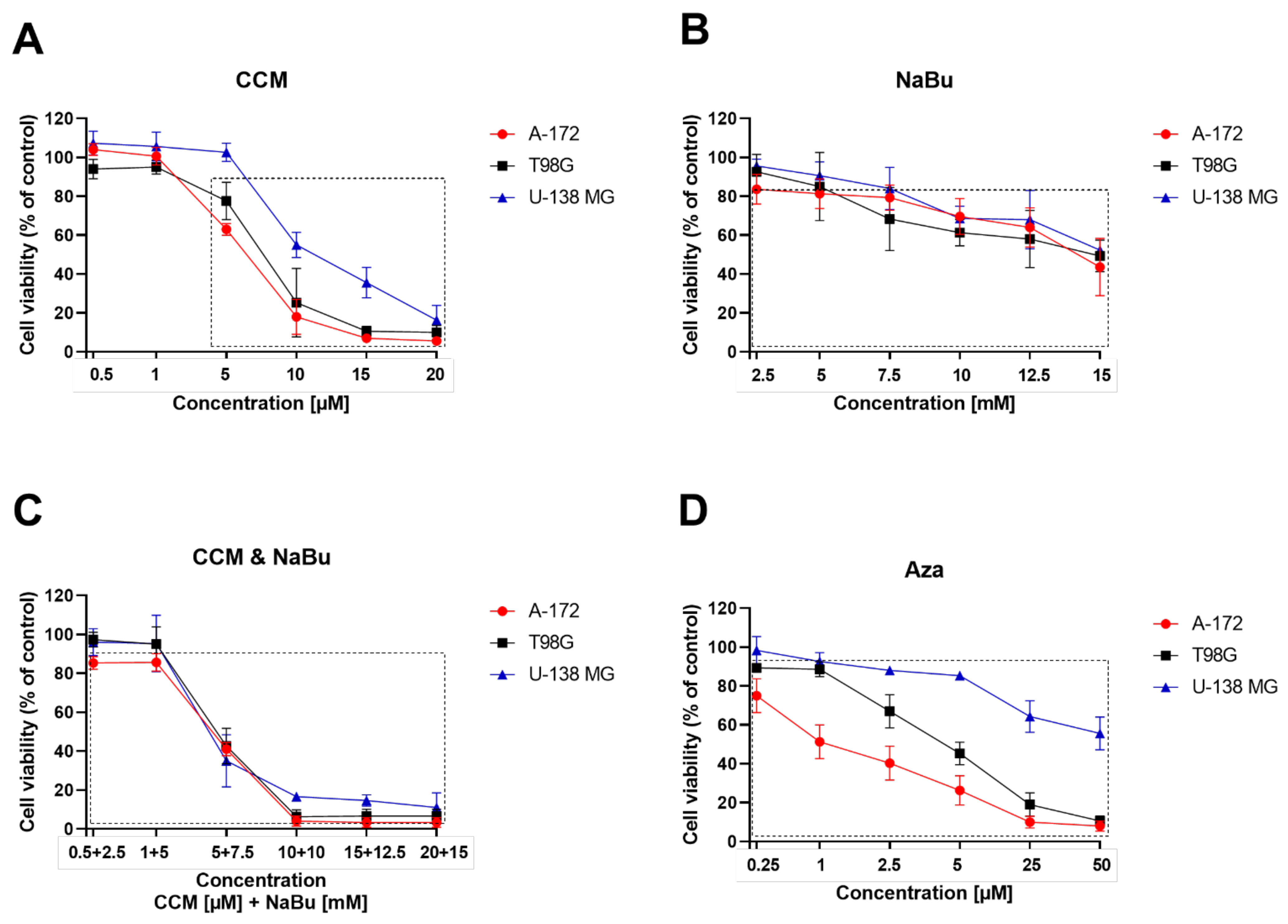
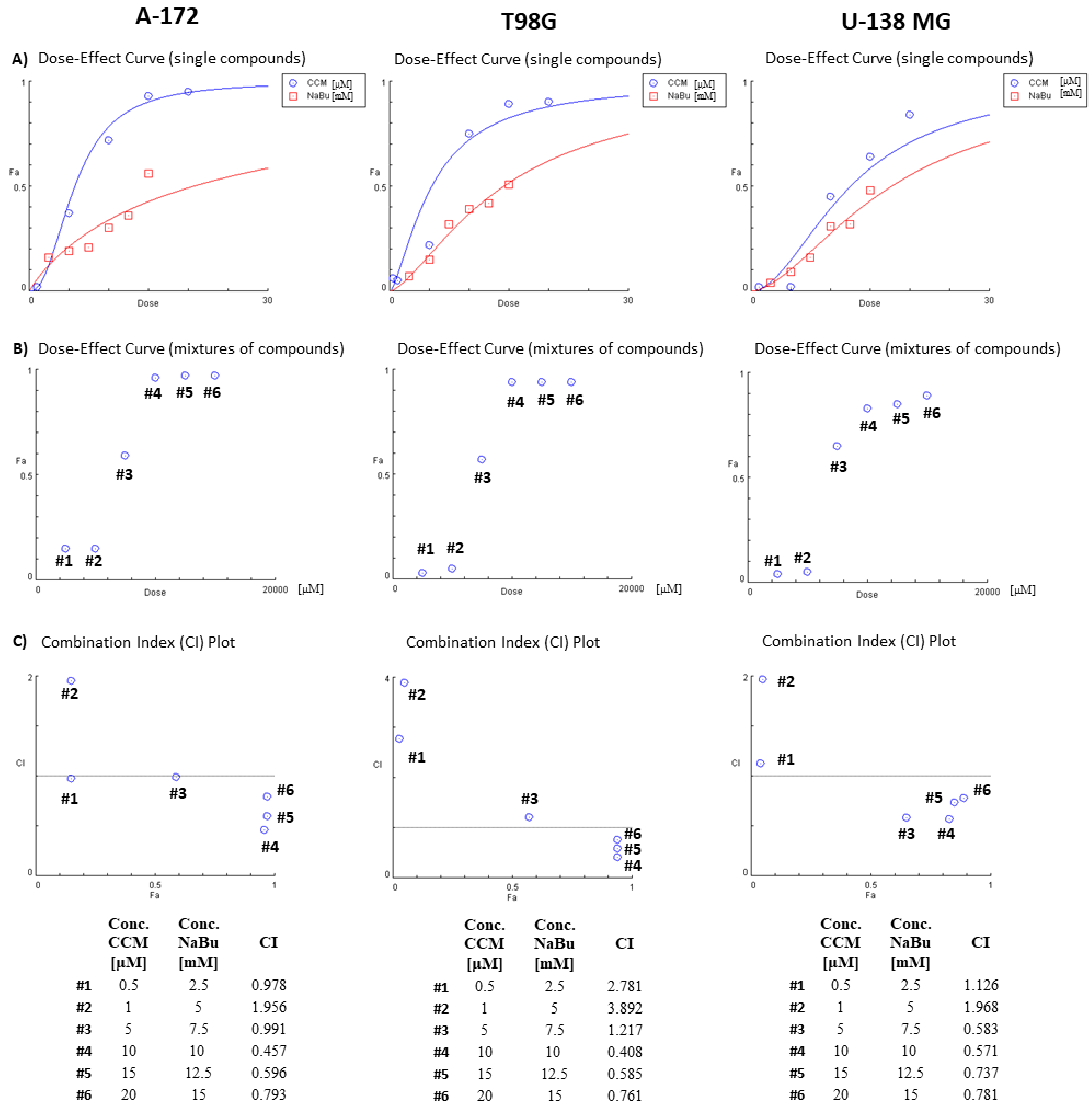
CCM and NaBu are natural compounds with anticancer effects and minimal toxicity. However, their molecular targets and mechanisms exerted in GBM cells are still unclear. In this study, we found that CCM and NaBu decreased the viability of GBM cells and these compounds, and when administered in concentrations higher than 1 µM of CCM and 5 mM of NaBu, acted synergistically. This is a novel finding, as to the best of our knowledge, the combination treatment of CCM and NaBu was not analyzed in GBM cells previously. Synergism between another polyphenol, quercetin, and NaBu for controlling growth of GBM cells was, however reported [
21]. In another study, curcumin used as a single compound reduced GBM cell viability, inhibiting activation of the PI3K/Akt/mTOR pathway [
22]. Intriguingly, curcumin did not modify the phenotype of healthy astrocytes, suggesting that this natural compound selectively targets transformed cells [
22]. As far as NaBu is concerned, in a study of Nakagawa et al. [
17], its physiological concentrations (0.25–4.00 mM) dose-dependently inhibited A-172 cells proliferation and invasion. Similar results were also published using a model of U251 cells [
23]. Importantly, in that study, the decreased viability was also accompanied by the increased radiosensitivity of GBM cells.
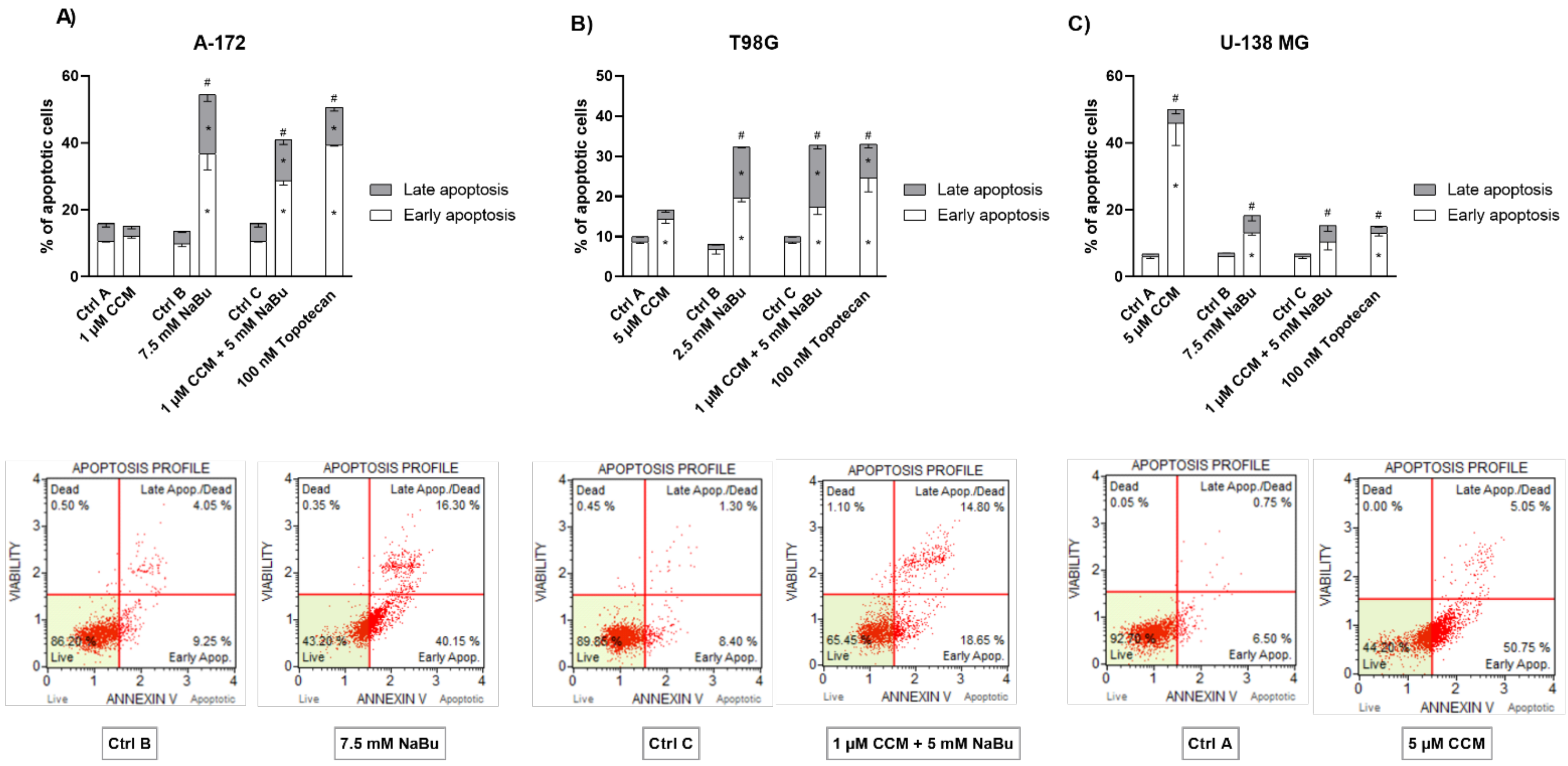
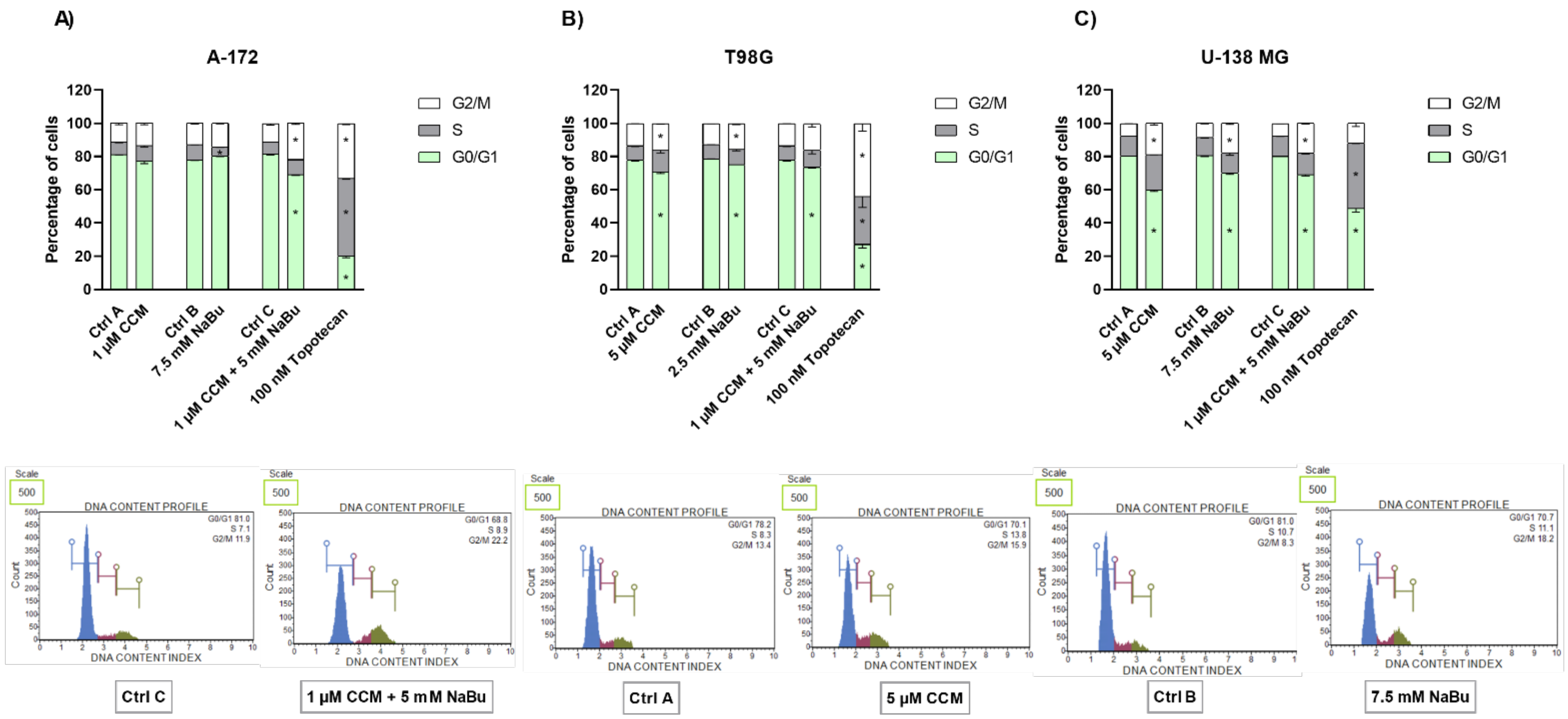
To decipher what mechanisms may be responsible for the observed viability reduction of GBM cells, we performed subsequent apoptosis and cell cycle flow cytometry analyses. We found that CCM and NaBu induce apoptotic cell death; however, the degree of apoptosis was cell line-dependent. In this regard, CCM was most active in U-138 MG cells, while in T98G, it increased the percentage of only early apoptotic cells. On the other hand, NaBu acted proapoptotically in all analyzed cells, but the degree of apoptosis varied between the cell lines. Moreover, we observed significant increases in apoptotic cell number when the combination of compounds was used, and the results for combination treatment resembled that of a reference anticancer drug—topotecan. The proapoptotic effect of the tested compounds was also accompanied or could were induced by the cell cycle distribution changes. In this regard, the combination treatment of CCM and NaBu was actively arresting the cell cycle in the G2/M phase in all cell lines, while single compounds were active in two out of three analyzed cell lines, i.e., T98G and U-138 MG cells.
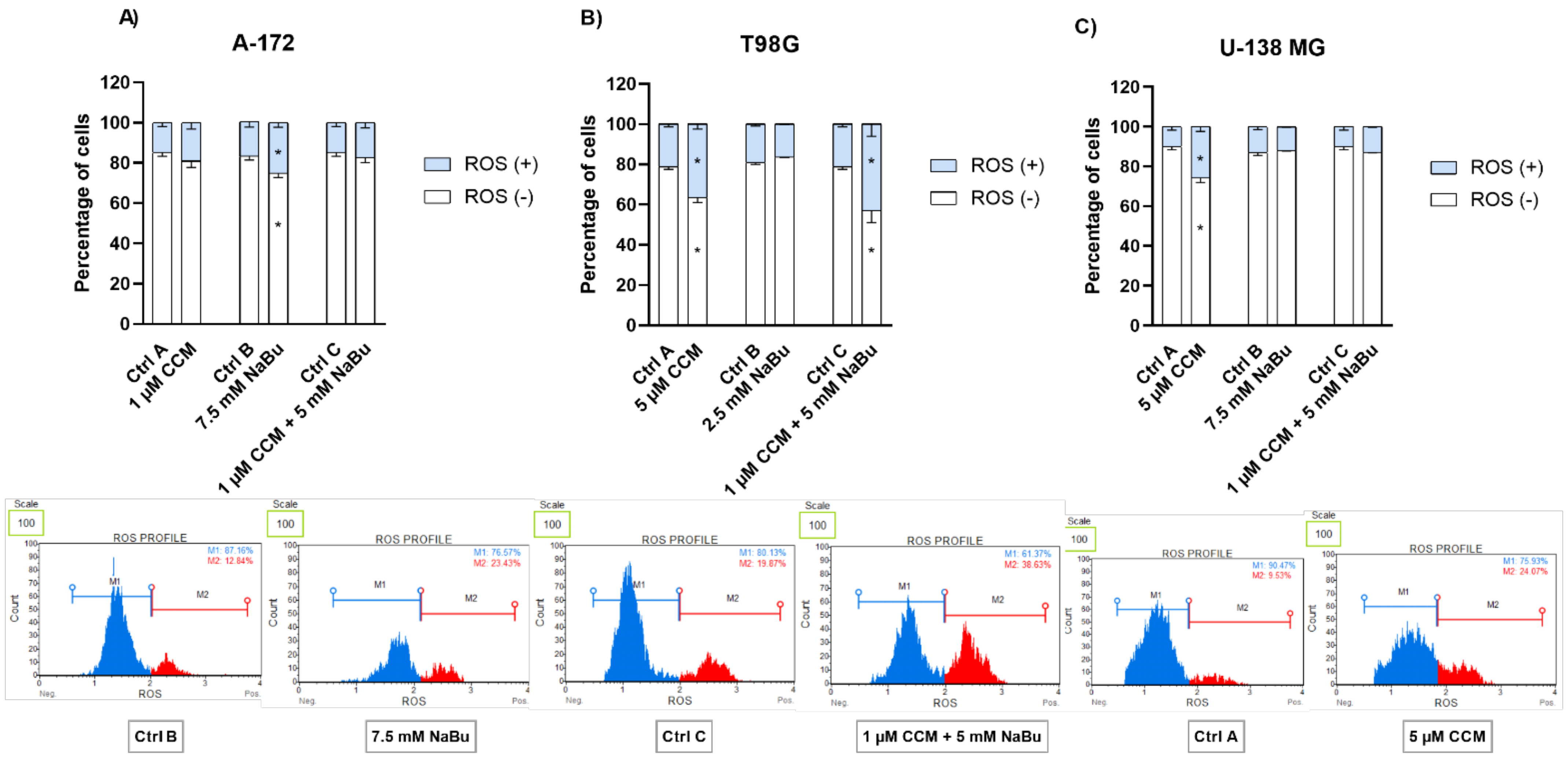
Next, in the attempt to unravel the molecular mechanisms underlying CCM and NaBu-mediated growth inhibition and apoptosis induction, we performed the oxidative stress analysis of the cells treated with the analyzed compounds and their combination. Here, we report that direct cytotoxic and proapoptotic effects of CCM in T98G and U-138 MG cells are mediated, at least in part, via ROS production. Our findings are in line with a study by Gersey et al. [
24], who found that 25 μM curcumin significantly induced ROS in GBM stem cells. Thus, these data confirm that even though curcumin is considered a natural phenolic antioxidant, it can act pro-oxidatively in cancer cells, inducing desirable anticancer effects. Moreover, in our study also NaBu-elicited apoptosis in A-172 cells was accompanied by an elevated level of ROS. Similar results were published by Nakagawa et al. [
17] and Salimi et al. [
16]; however, the latter reported pro-oxidative effects of NaBu in regard to breast cancer cells. Thus, the data suggest that ROS generation may be responsible for the observed increase in the apoptotic cell number in CCM and NaBu-treated GBM cells.
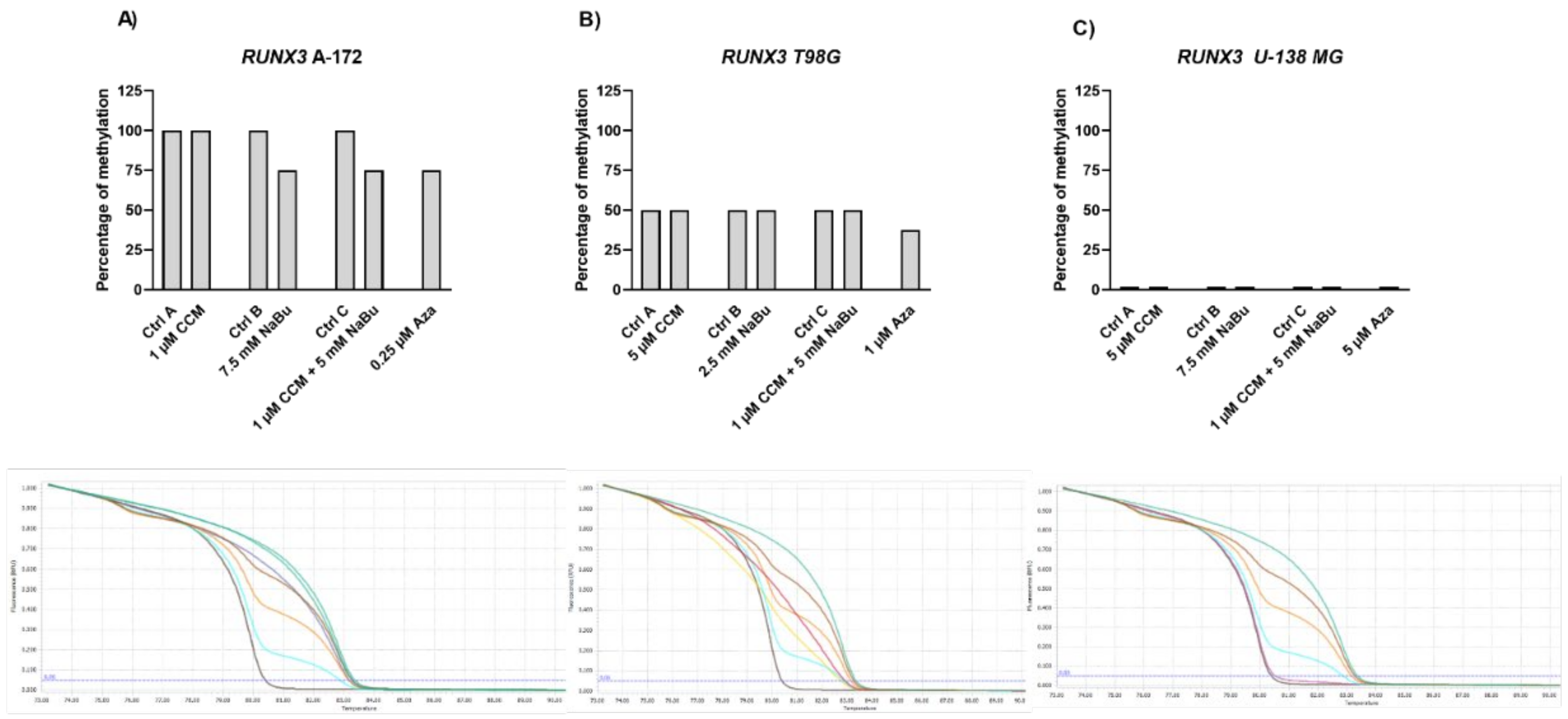
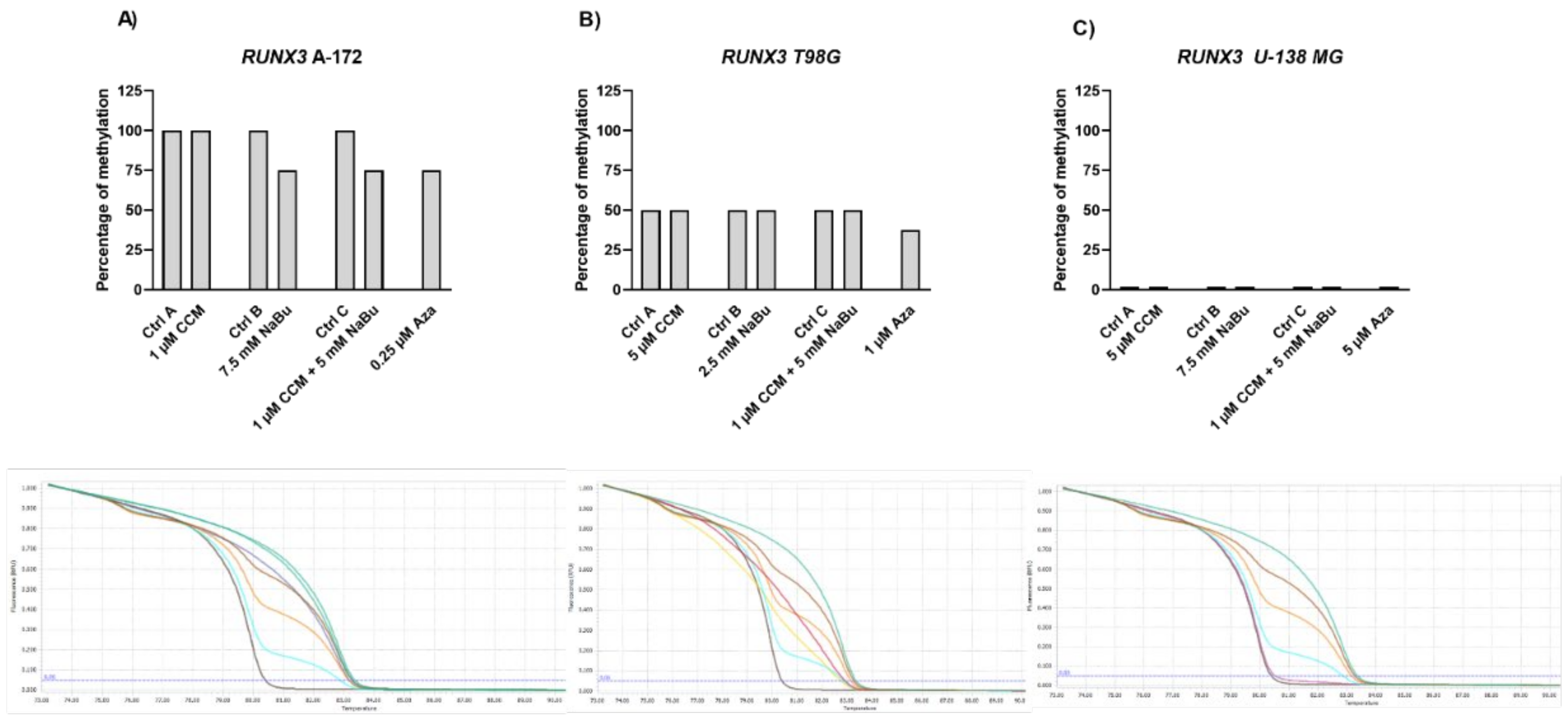
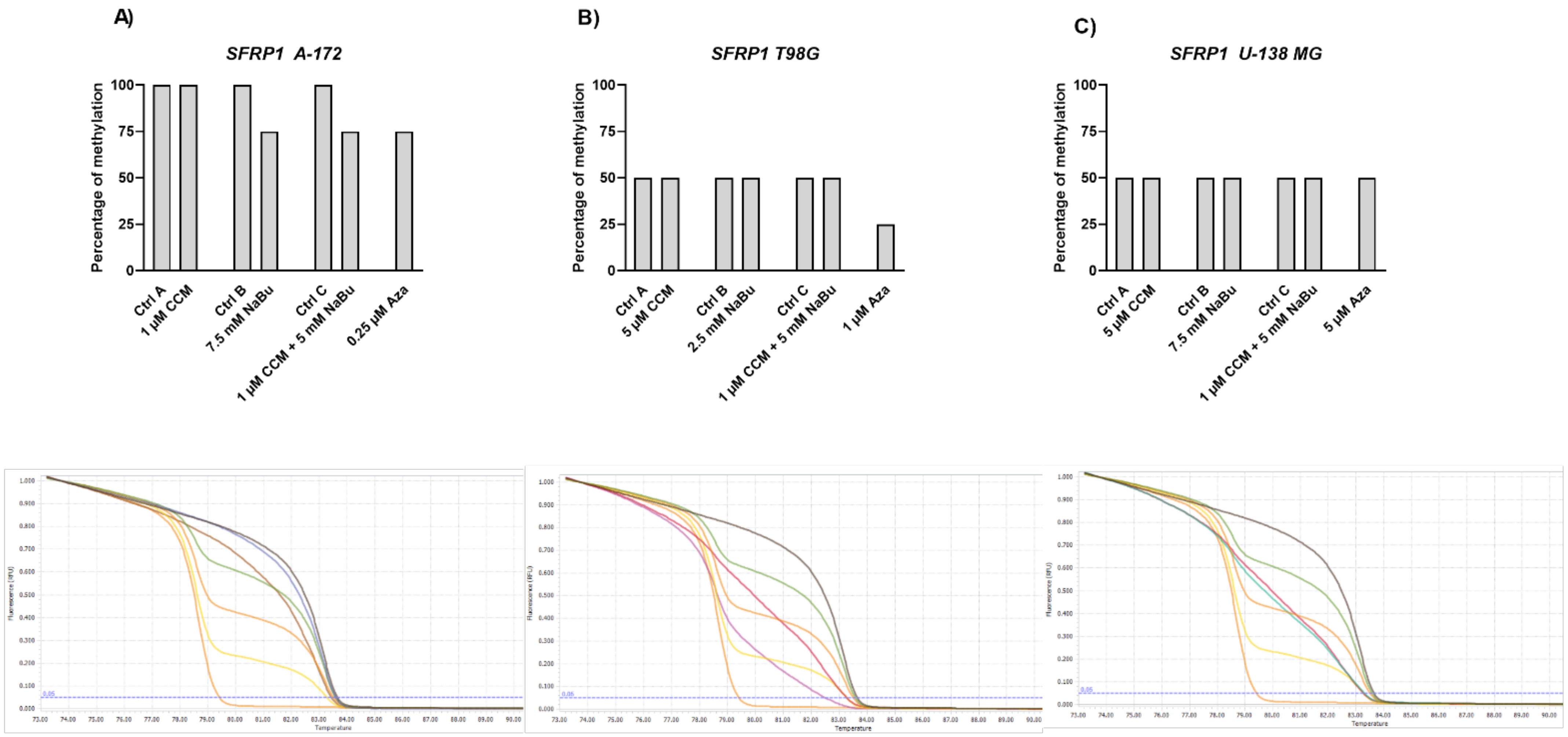
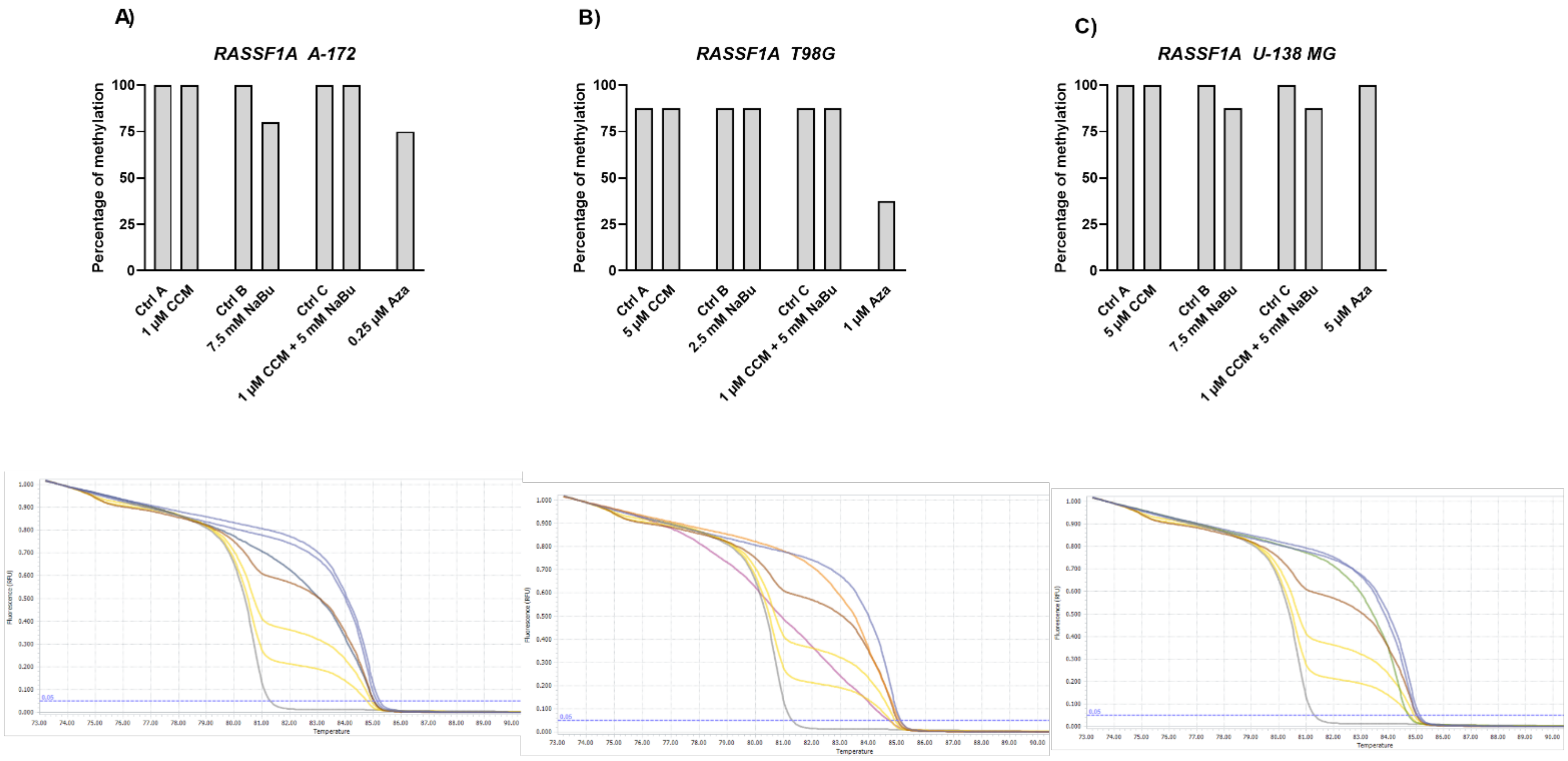
The Wnt/β-catenin signaling pathway modulates the transcription of genes linked to proliferation, differentiation, and tumor progression. Targeting this pathway is suggested as a good strategy in anti-GBM treatment, as it is often hyperactivated in GBM and contributes to malignant glioma stemness, invasiveness, therapeutic resistance, and angiogenesis. Previously, we and others found that Wnt/β-catenin signaling is hyperactivated in glioma cells due to promoter methylation of its antagonists, including
SFRP1 and
RUNX3 [
25]. Another frequent event in GBM cells is the epigenetic silencing of a master cell cycle regulator,
RASSF1A [
26]. Thus, we wanted to analyze if CCM and NaBu are able to reactivate
RUNX3,
SFRP1, and
RASSF1A gene expression and if this change in expression is accompanied by DNA promoter demetylation. Here, we report significant upregulation of
RUNX3 (in A-172 and U-138 MG) and
SFRP1 (in all three cell lines analyzed) after the treatment with NaBu and the combination treatment of CCM and NaBu. This is an important observation since NaBu was reported to modulate the Wnt signaling in a cell type- and promoter-dependent manner [
27]. For instance in SW620 colon carcinoma cells, NaBu downregulated β-catenin-dependent expression of the Tcf-TK, matrilysin, and cyclin D1 promoters [
27]. Contrarily, in another colon carcinoma cell line, HCT-116, NaBu upregulated the expression of these promoters [
27]. As the impact of NaBu on GBM cells in the context of Wnt signaling pathway was not analyzed so far, this novel finding is of great importance, especially since our previous study revealed that
SFRP1 methylation predicts shorter survival of glioma patients [
25].
The results of our study also indicate that NaBu, and its combination treatment with CCM, upregulates the critical cell cycle regulator, RASSF1A, in A-172 and T98G cells, but downregulates its mRNA level in U-138 MG cells. This finding shows a cell line-dependent manner of RASSF1A expression regulation by CCM and NaBu, and indicates that this issue should be further analyzed.
Gene expression changes are often a result of DNA methylation changes at the promoter, enhancer, or other regulatory regions. The ability of curcumin to demethylate tumor suppressor genes was previously demonstrated [
8]. On the other hand, NaBu is well known as histone deacetylase (HDAC) inhibitor, but its impact on DNA methylation was also reported [
10]. Thus, we wanted to analyze if the observed gene expression changes result from demethylating properties of the studied compounds in the context of the particular, important for gliomagenesis genes. Here, we found that CCM had no demethylating or hypomethylating properties regarding
RUNX3,
SFRP1, and
RASSF1A promoters in GBM cell lines analyzed in this study. In contrast, hypomethylating effects of NaBu were cell line- and gene-dependent, as it reduced the methylation level of selected genes in A-172 and T98G cell lines. Thus, we can conclude that the impact of CCM on the promoter methylation of
RUNX3,
SFRP1, and
RASSF1A was inconsiderable, while of NaBu—moderate. Similar results were obtained by Parashar et al. [
28], who investigated the reactivation of
PAX1 using curcumin and resveratrol in HeLa, SiHa, and Caski cell lines. They found that curcumin in HeLa and SiHa cells and resveratrol in Caski cells caused significant reactivation of
PAX1 expression; however, the reversal of promoter hypermethylation was not observed across the three cell lines. Contrarily, in a study of Shin et al. [
10] NaBu induced demethylation and histone modification at the promoter region of
SFRP1/2, restoring the SFRP expression in human gastric cancer cells. Similar to our results, in that study, NaBu did not induce complete demethylation, but generated a more sporadic pattern of demethylation.
Interestingly, in our study, even Aza, the positive control with DNMT inhibitory properties, was not found to be effective in causing complete demethylation of CpG sites under consideration, suggesting the promoter regions to be resistant towards hypomethylating effects. We discovered that Aza was able to reduce the DNA methylation levels of
RUNX3,
SFRP1, and
RASSF1A of only ~25%, and did not completely demethylate any of the analyzed sequences. In this context, we can conclude that NaBu and its combination treatment with CCM had similar hypomethylating properties as Aza. Incomplete demethylation at various genomic regions after the treatment with Aza and another hypomethylating agent, 5-aza-2′-deoxycytidine (5-Aza-dc) were also reported by others [
28,
29]. It was shown that Aza treatment does not result in demethylation of every CpG in DNA, as some Aza-treated regions can remain methylated, even to a large extent. For instance, the presence of methylated DNA in samples treated with 5-Aza-dc was reported for genes such as
Axin1,
CXCL12,
DAPK1, and others [
29]. These findings can explain why curcumin (the major component of CCM used in our study), despite being frequently reported as a potent DNMT inhibitor, was not effective in demethylating the genes chosen in our experimental setting.
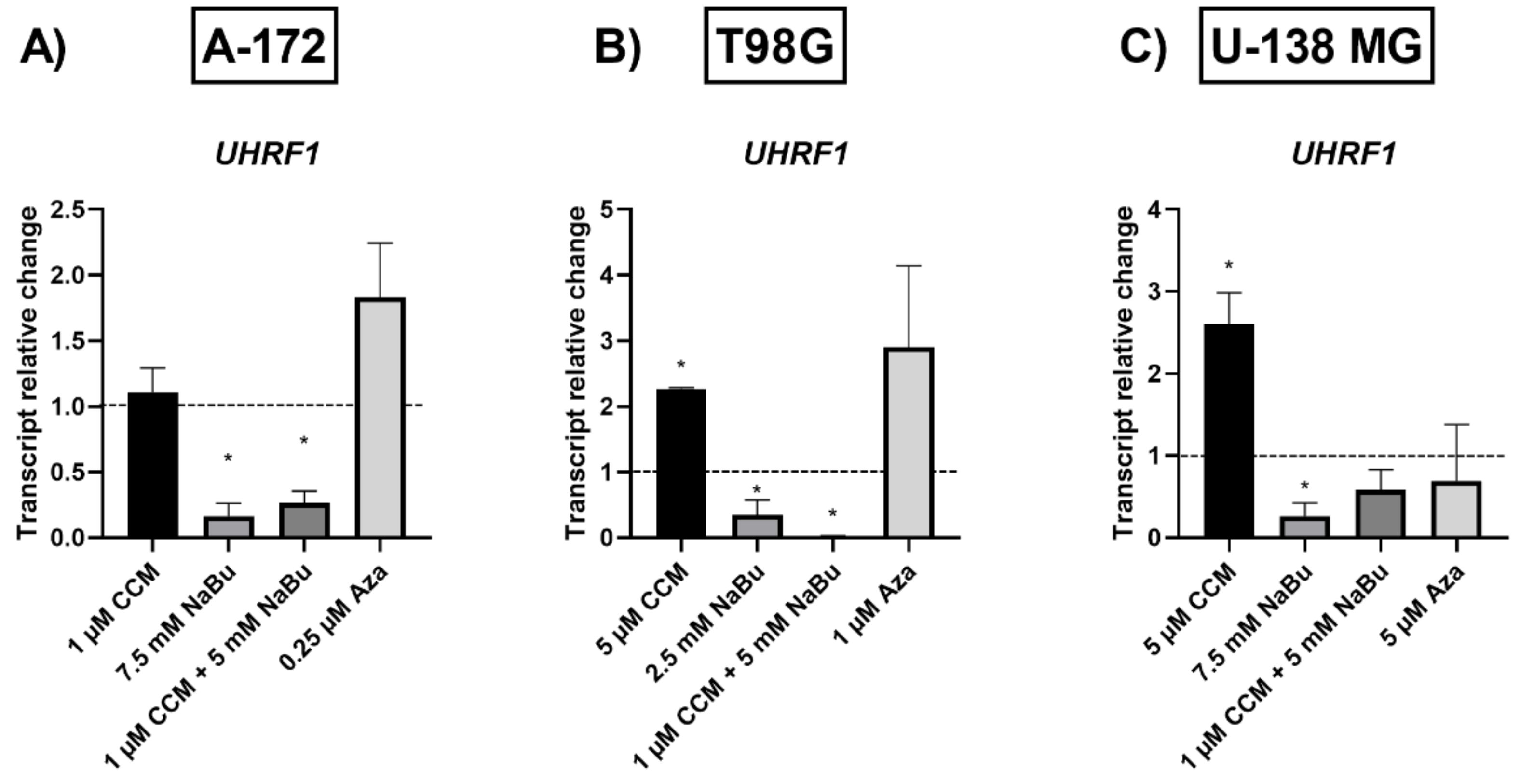
Since the gene expression changes observed in our study were not caused by complete promoter demethylation, we wanted to verify, if the analyzed compounds had an impact on the key epigenetic regulator—UHRF1 (ubiquitin-like with plant homeodomain and really interesting new gene finger domain 1). UHRF1 is involved in a macro-molecular protein complex called ‘Epigenetic Code Replication Machinery’ (ECREM), which is coordinating DNA methylation and histone modifications [
30]. Importantly, UHRF1 is regarded as a potential target in anticancer therapy since its expression is upregulated in multiple tumors, and its downregulation was reported to lead to growth arrest and apoptosis of cancer cells [
30]. Here we report that NaBu and the combination treatment of CCM and NaBu downregulated the expression of
UHRF1 in GBM cells. Contrarily, CCM upregulated its expression. Other research groups already showed that UHRF1 can be targeted by phytocompounds, including curcumin, but both downregulation and upregulation were reported, depending on the cell line analyzed and the time of exposure to the compound [
28]. Regarding the complex role played by UHRF1, and the ambiguous data from different experimental setups, further studies are urgently needed to fully elucidate the impact of CCM on this epigenetic regulator and the subsequent phenomena controlled by it. Nevertheless, NaBu and the combination treatment of CCM and NaBu can be regarded as promising compounds downregulating
UHRF1 in GBM cells. We hypothesize that the anticancer effects including reduced viability, apoptosis induction, and cell cycle arrest accompanying the treatment with NaBu and the combination treatment of CCM and NaBu, can be, at least in part, related to
UHRF1 downregulation.
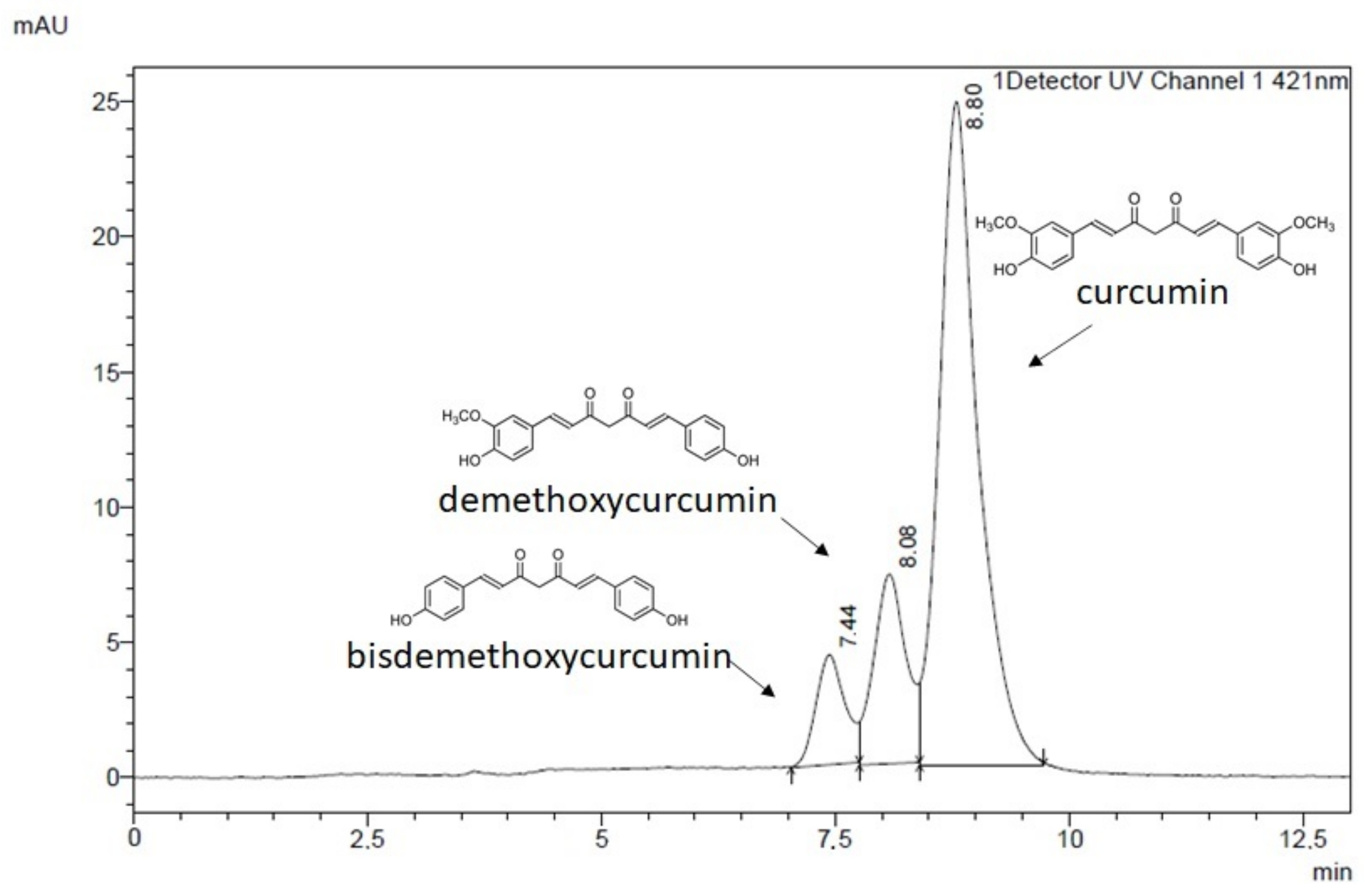
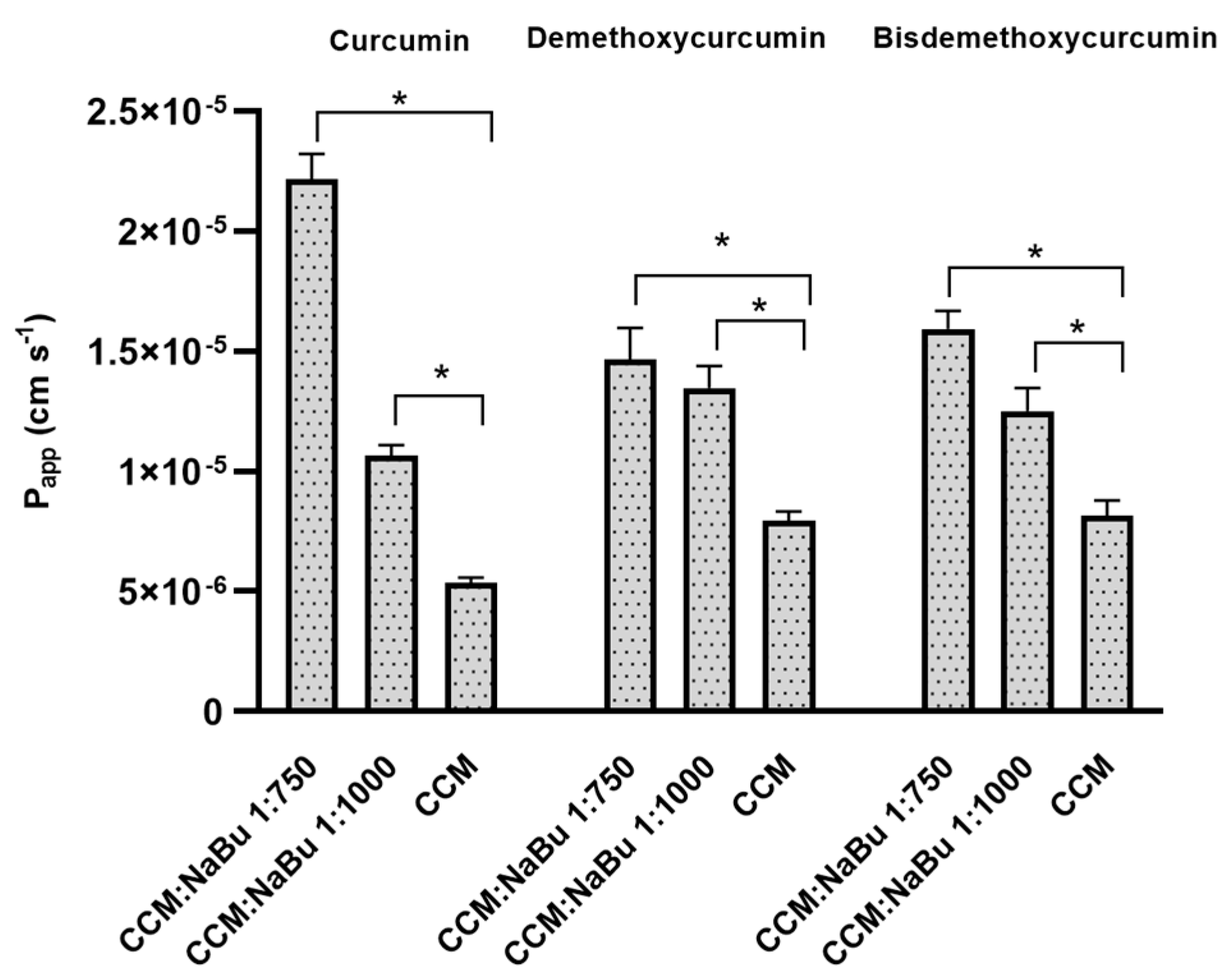
Finally, to verify if the two compounds can successfully be implemented as a combination treatment targeting CNS, the permeability of CCM and NaBu through the BBB and the nasal cavity was analyzed. Curcumin used as a single compound was already found to be able to cross the BBB [
5], yielding therapeutic benefits within the CNS. NaBu is also able to reach CNS via the gut-brain axis. In humans, colonic butyrate can range from 10 to 20 mM [
31]. Physiologically, NaBu is detectable in the human cerebrospinal fluid (CSF), typically in the range of 0–2.8 μM, and an average concentration of 17.0 pmol/mg in the human brain was reported. Thus, the physiological concentration of this microbiota-derived metabolite is far lower than that required for anticancer effects [
13]. Novel delivery systems are therefore needed to expose GBM cells to the increased concentrations of both compounds. Interestingly, in this study, we found that CCM permeability through the BBB and through the nasal cavity is increased, when it is administered together with NaBu. In fact, the combination of CCM with NaBu increased the permeability of all three curcuminoids, including curcumin, demethoxycurcumin, and bisdemethoxycurcumin. This finding paves the way for novel oral, or nose-to-brain delivery systems, with better bioavailability and improved efficacy. Intranasal drug delivery systems seem especially promising, as they are not only effective in bypassing the BBB but also very convenient to use and painless. To conclude, the findings of this study give rationale for further pre-clinical and clinical analysis of the combination treatment of CCM and NaBu and suggest the great value of the implementation of CCM and NaBu-based drug candidates in anti-GBM treatment.
4. Materials and Methods
4.1. Chemicals
CCM, NaBu, Aza, topotecan, dimethylsulfoxide (DMSO), antibiotics solution (10,000 units penicillin and 10 mg streptomycin/mL), Eagle’s Minimum Essential Medium (EMEM), Dulbecco’s modified Eagle medium (DMEM), fetal bovine serum (FBS), glutamine, nonessential amino acids (NEAA), sodium pyruvate (NaP), ribonuclease A (RNase A), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), trypsin, Tris, and all other compounds were purchased from Sigma–Aldrich (St. Louis, MO, USA).
CCM and Aza were dissolved with DMSO to make 10 mM stock solutions, aliquoted and frozen at −20 °C, and −80 °C, respectively. Only one freeze-thaw cycle was used regarding Aza to ensure maximal stability. For the same reason, NaBu was at all times dissolved ex tempore in PBS to make a 100 mM stock solution.
4.2. HPLC Analysis
The HPLC separation was carried out on a Phenomenex-C18 column (250 mm × 4.6 mm; 5 µm), the temperature of the column set at 303 K, using a mobile phase consisted of 1% acetic acid and acetonitrile (45:55, v/v), with the flow rate 1.0 mL/min (1). The injection volume was 20 µL, and maximum absorption was measured using Diode-Array (DAD) detector set at 421 nm. As reference standards, curcumin (purity > 99.5%), demethoxycurcumin (purity > 95.0%) and bisdemethoxycurcumin (purity > 98%) were used.
4.3. Cell Lines and Culture
Human GBM cell lines, A-172 and U-138 MG were obtained from American Type Culture Collection (ATCC), whereas human GBM T98G cell line was bought from the European Collection of Authenticated Cell Cultures (ECACC). The cells were maintained in the recommended media: ATCC-formulated Dulbecco’s modified Eagle’s medium (DMEM) (Merck, Darmstadt, Germany) in case of A-172 cell line, and ATCC-formulated Eagle’s Minimum Essential Medium (EMEM) (Merck, Germany), in case of T98G and U-138 MG. The media were supplemented with FBS (Biowest, Nuaillé, France) to a final concentration of 10%, as well as antibiotics (penicillin and streptomycin) (Merck, Darmstadt, Germany) to the final concentrations of 1%. For the experiments, the amount of FBS was reduced to 5%. Additionally, the medium for T98G was supplemented with 2 mM glutamine, 1% non-essential amino acids, and 1% sodium pyruvate (all purchased from Merck, Germany). The cells were propagated at 37 °C and 5% CO2 in a humidified incubator (Memmert, Schwabach, Germany).
4.4. Cytotoxic Activity
The cytotoxic activity of CCM, NaBu, and their combinations, as well as Aza, a demethylating agent used as a control for methylation analysis, was performed using the MTT assay. Briefly, cells were cultured, harvested, and plated in 96-well plate at a seeding density of 10,000 cells/well. After 24 h of preincubation, increased concentrations of CCM (0.5–20 µM), NaBu (2.5–15 mM), CCM and NaBu (0.5 µM + 2.5 mM − 20 µM + 15 mM of CCM and NaBu, respectively) or Aza (0.25–50 µM) were added, and cells were grown for 48 h at 37 °C. Appropriate controls with DMSO (for CCM, the combination treatment of CCM and NaBu, and Aza) and PBS (for NaBu) were performed. Afterwards, the cells were washed with 200 µL of PBS, followed by incubation with 3-(4,5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide (MTT) solution (Merck, Darmstadt, Germany) in 10% FBS medium (0.5 mg/mL) for 3 h. Finally, the formazan crystals were dissolved in acidic isopropanol, and the absorbance was measured at λ = 570 nm and λ = 690 nm on the microplate reader (Tecan Infinite M200, Grödig, Austria). All the experiments were repeated three times with four measurements per assay.
4.5. The Combinatorial Effects of the Compounds on Cytotoxic Activity (Combination Index)
The combinatorial effect of the compounds on cell viability was determined by the evaluation of the Combination Index (CI) using the CompuSyn software (
www.combosyn.com, accessed on 18 August 2021) [
32]. The synergistic action of the chemicals in combinations was identified when CI ˂ 1.
4.6. Apoptosis Analysis
The flow cytometry apoptosis analysis using the Muse® Annexin V & Dead Cell Kit (Merck, Darmstadt, Germany) was performed following the protocol provided by the manufacturer. The principle of this method relies on Annexin V binding to phosphatidylserine—the marker of early apoptosis, while 7-Aminoactinomycin binding to DNA in late apoptotic/dead cells. Briefly, 100,000 cells of A-172 and T98G cell lines, and 150,000 cells of U-138 MG cell line were seeded on 6-well plates and incubated for 24 h. Afterward, the analyzed compounds were added in concentrations based on MTT results, and the cells were further incubated for 48 h. Cells treated with the vehicle (DMSO or PBS) and 100 nM topotecan were used as negative and positive controls, respectively. The subsequent analysis was performed on Muse™ Cell Analyzer (Merck, Darmstadt, Germany).
4.7. Cell Cycle Distribution Analysis
The cell cycle distribution analysis using the Muse™ Cell Cycle Kit and Muse™ Cell Analyzer (Merck, Darmstadt, Germany) was used to check the number of cells in each phase of the cell cycle. The assay utilizes propidium iodide-based staining of DNA content to discriminate and measure the percentage of cells in each cell cycle phase (G0/G1, S, and G2/M). The analysis was performed according to the protocol provided by the manufacturer and was previously described [
33]. Briefly, after 48 h incubation with the analyzed compounds, the cells were trypsinized, washed with PBS buffer, fixed in ice-cold 70% ethanol, and stored until staining at −20 °C. Before the analysis, the fixed cells were washed with PBS buffer, stained, and subjected to 0.5 h incubation at room temperature in the darkness. Cells treated with the vehicle (DMSO or PBS) and 100 nM topotecan were used as negative and positive controls, respectively. The subsequent analysis was performed on Muse™ Cell Analyzer (Merck, Germany).
4.8. Oxidative Stress
The intracellular detection of superoxide radicals was performed using The Muse® Oxidative Stress Kit (Merck, Darmstadt, Germany), according to the manufacturer’s recommendations. The principle of the method is based on dihydroethidium (DHE), which is cell permeable and upon reaction with superoxide anions undergoes oxidation to form the DNA-binding fluorophore. The fluorophore intercalates with DNA which results in red fluorescence. Briefly, the cells were treated with the analyzed compounds/mixes, in the concentrations allowing >70% of cell survival. After 48 h the cells were trypsinized, washed with PBS buffer, and resuspended in Assay Buffer containing Working Solution of Muse® Oxidative Stress Reagent. The cells were incubated at 37 °C for 30 min, and then run on Muse® Cell Analyzer (Merck, Darmstadt, Germany). Cells treated with the vehicle (DMSO or PBS) were used as negative control.
4.9. cDNA Synthesis & qPCR
The total RNA was isolated using Universal RNA Purification Kit (EURx, Gdańsk, Poland) and subsequently subjected to reverse transcription using The Revert-Aid First Strand cDNA Synthesis kit (Fermentas, Burlington, Canada). Quantitative real-time PCR (qPCR) was performed using Hot FIREPol EvaGreen qPCR Mix Plus (Solis Biodyne, Tartu, Estonia) and LightCycler 96 (Roche Diagnostics GmbH, Mannheim, Germany). Primer sequences were previously published [
3,
28,
34,
35], and their synthesis was performed at the Institute of Biochemistry and Biophysics, Warsaw, Poland. The protocol started with 15 min enzyme activation at 95 °C, followed by 40 cycles of 95 °C for 15 s; appropriate annealing temperature for 20 s; 72 °C for 40 s and the final elongation at 72 °C for 5 min which was followed by melting curve analysis to confirm the generation of a single amplification product. TATA box-binding protein (
TBP), a component of the DNA-binding protein complex TFIID, served as the endogenous RNA control, and each sample was normalized on the basis of its
TBP content. The results were expressed as N-fold differences in gene expression relative to the
TBP gene. Each sample was analyzed in triplicate, and the experiments were repeated twice.
4.10. Sodium Bisulfite Treatment & Methylation-Sensitive High-Resolution Melting Analysis (MS-HRM)
EZ DNA Methylation Kit (Zymo Research, Irvine, CA, USA) was used for sodium bisulfite treatment, i.e., deamination of unmethylated cytosines within CpG dinucleotides to uracil while leaving the 5-methylcytosines intact. Such bisulfite-converted DNA was subsequently subjected to MS-HRM analysis. All the methodological details of the MS-HRM analyses including the reaction and primer sequences can be found elsewhere [
3,
34], and we previously validated the MS-HRM protocols used in this study with pyrosequencing, which is considered the gold standard technique for DNA methylation investigations. Briefly, the standard solutions of completely methylated genomic DNA, i.e., CpG Methylated HeLa Genomic DNA (New England Biolabs, Ipswich, MA, USA) and completely unmethylated human genomic DNA, i.e., CpGenome Universal Unmethylated DNA Set (Merck, Darmstadt, Germany) were used to generate melting curves of known methylation level (100%, 75%, 50%, 25%, and 0% methylated DNA). To semiquantitatively estimate the sample methylation levels, their sample melting profiles were compared to those of standards. Samples were run in duplicate in each experiment, and each experiment was repeated twice. The protocol involved 15 min of preincubation at 95 °C and 40 cycles of three-step amplification (15 s/95 °C, 20 s/Ta, 20 s/72 °C), and obtained amplicons were melted in a temperature gradient to max 95 °C. The obtained melting curves were normalized automatically by the calculation of the “line of the best fit” in between two normalization regions before and after the significant fluorescence decrease. The methylation level of each sample was assessed by comparison of the PCR product normalized melting curve/peak with the normalized melting curves/peaks of the standard solutions.
4.11. PAMPA Model
The impact of NaBu on the permeability of CCM (curcumin, and two other curcuminoids—demethoxycurcumin and bisdemethoxycurcumin) through the BBB and the wall of nasal cavity was investigated using the Parallel Artificial Membrane Permeability Assay model (PAMPA). The model defines the permeability basing on the passive diffusion across the BBB and nasal mucosa. The PAMPA set consists of donor and acceptor plates, separated by a 120 μm-thick microfilter disc coated with a 20% (
w/v) dodecane solution of a lecithin mixture (Pion Inc., Billerica, MA, USA). Prisma buffers (Prisma HT, Pion Inc.) of pH 7.4 and pH 6.0 were used in the BBB permeability and nasal cavity permeability tests, respectively. The samples containing curcuminoids were studied alone and with NaBu in molar ratio CCM:NaBu 1:750; and 1:1000. The samples were distributed to donor wells with appropriate Prisma buffer, and the acceptor wells for nasal and BBB permeability were filled with Acceptor Sink Buffer, and Brain Skin Buffer (Pion Inc., Billerica, MA, USA), respectively. Then, the plates were combined to form a sandwich. The nasal PAMPA plate was incubated for 1 h, while PAMPA BBB was incubated for 4 h in a humidity-saturated atmosphere at the temperature set at 37 °C. After incubation, the plates were separated to investigate curcuminoids concentrations in donor and acceptor chambers using the HPLC-DAD method, as described above (
Section 4.1). The apparent permeability coefficient (P
app) was calculated due to the equation:
where
VD—donor volume,
VA—acceptor volume,
Cequilibrium—equilibrium concentration
,
S—membrane area,
t—incubation time (in seconds).
4.12. Statistical Analysis
All data are present as the mean and standard error of the mean (SEM). Differences among experimental groups were determined by the unpaired t-test with two-tailed distribution. In all comparisons, statistical significance was set at p < 0.05.


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}