Abstract
Although engineered cyanobacteria for the production of lipids and fatty acids (FAs) are intelligently used as sustainable biofuel resources, intracellularly overproduced FAs disturb cellular homeostasis and eventually generate lethal toxicity. In order to improve their production by enhancing FFAs secretion into a medium, we constructed three engineered Synechocystis 6803 strains including KA (a mutant lacking the aas gene), KAOL (KA overexpressing lipA, encoding lipase A in membrane lipid hydrolysis), and KAOGR (KA overexpressing quadruple glpD/rbcLXS, related to the CBB cycle). Certain contents of intracellular lipids and secreted FFAs of all engineered strains were higher than those of the wild type. Remarkably, the KAOL strain attained the highest level of secreted FFAs by about 21.9%w/DCW at day 5 of normal BG11 cultivation, with a higher growth rate and shorter doubling time. TEM images provided crucial evidence on the morphological changes of the KAOL strain, which accumulated abundant droplets on regions of thylakoid membranes throughout the cell when compared with wild type. On the other hand, BG11-N condition significantly induced contents of both intracellular lipids and secreted FFAs of the KAOL strain up to 37.2 and 24.5%w/DCW, respectively, within 5 days. Then, for the first time, we shone a spotlight onto the overexpression of lipA in the aas mutant of Synechocystis as another potential strategy to achieve higher FFAs secretion with sustainable growth.
1. Introduction
Photosynthetic cyanobacteria and microalgae are classified as third-generation renewable energy resources of biofuels, such as biodiesel, bio-oil, and biohydrogen, due to their atmospheric CO2 fixation without food source competition [1,2,3]. In cyanobacterial biomass, lipids and fatty acid (FAs) biomolecules are mainly distributed as components of the membranes [3]. In order to balance lipid homeostasis in the cyanobacterium Synechocystis sp. PCC 6803, not only FA and membrane lipid syntheses but also FFAs recycling and FFAs secretion are directly involved [4]. Increasing lipid-producing microorganisms by genetic engineering has gained attraction in recent years [4,5,6], but with limitations on productivity, such as toxicity of overproduced FAs. Excessive levels of intracellular FFAs in the engineered FFA-producing Synechococcus elongatus strain (dAS1T) ultimately generated toxicity and resulted in cell death [7]. Recently, to address the FFAs toxicity, Synechocystis sp. PCC6803, Synechococcus elongatus PCC7942, and Synechococcus sp. PCC 7002 strains secreting FFAs out of cells and into the culture medium have been identified [4,8,9,10]. This FFA-secretion property of cyanobacteria is considered an important aspect of industrial productivity, such as cost reduction of production and downstream processing, including harvesting, drying, and cell breakage for FFA product recovery [11]. Strategies for improving intracellular lipid production and FFA excretion from cyanobacteria and algae have recently been focused on physiological conditions (such as nitrogen and phosphorous deprivations), environmental stress challenges, and genetically metabolic engineering approaches [12,13,14]. Known mechanisms using metabolic engineering approaches for increasing FFA secretion in cyanobacteria were previously reported, such as weakening of cell membrane layers by manipulating the surface layer (S-layer) protein encoded by the sll1951 gene in Synechocystis 6803 [8]. This allowed FFA diffusion across the lipid layers with inactivation of acyl-ACP synthetase (aas) which prevented FFA recycling to acyl-ACP, and as a result, a secretion of produced FFAs into the culture medium [4,15,16,17]. Heterologous expression of the tesA gene encoding thioesterase cleaves the thioester bond of acyl-ACP to FFAs [8,18]. Moreover, integrated strategies were extensively implemented to attain higher FFAs secretion, such as a modified strain with both aas inactivation and tesA expression [7,9,19,20], double aac/tesA expression integrated with an inactivation of fadD gene encoding fatty acyl-CoA synthase in Escherichia coli [21,22], or deletions of genes involved in acyl-CoA synthesis (such as Δfaa genes) and beta-oxidation (such as Δpox1) in Saccharomyces cerevisiae [23,24].
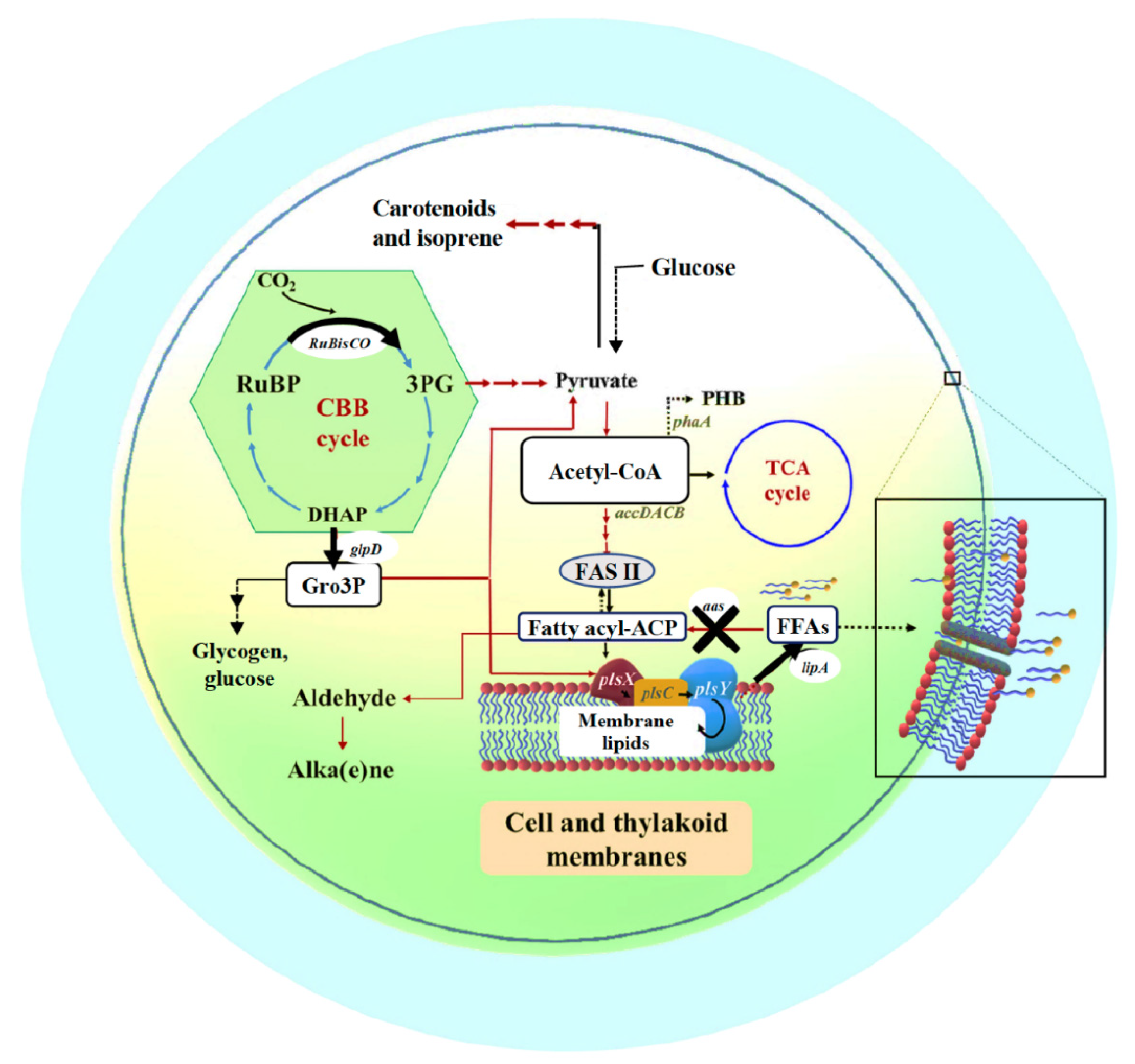
The metabolic networks of the Calvin–Bassham–Benson (CBB) cycle, fatty acid and membrane lipid biosyntheses and their linkage pathways including membrane hydrolysis and FFA recycling are depicted in Figure 1 (modified from [6]). In photosynthetic organisms, the CBB cycle, initially catalyzed by the key enzyme of RuBisCO, ribulose-1,5-bisphosphate carboxylase/oxygenase, provides many key intermediates, such as 3PG which is subsequently converted to pyruvate, and DHAP that flows through several reactions for RuBP production and partly to Gro3P via glpD (glycerol-3-phosphate dehydrogenase) catalysis [25,26]. This Gro3P, or glycerol-3-phosphate intermediate, is subsequently incorporated to many biochemical reactions, such as pyruvate production, gluconeogenesis, membrane lipid synthesis, and lycopene production [26,27]. For fatty acid and membrane lipid syntheses, acetyl-CoA is converted to malonyl-CoA, catalyzed by multi-subunit acetyl-CoA carboxylase, encoded by accDACB, and consecutively flows through fatty acid synthesis II (FASII) which may generate an intermediate fatty acyl–acyl carrier protein (fatty acyl-ACP), a substrate for membrane lipid synthesis via PlsX, PlsC and PlsY enzymes. The linkage between membrane lipids and the CBB cycle attains the glycerol backbone of lipid molecule from Gro3P, a product from glpD reaction. In terms of membrane lipid homeostasis, not only its synthesis but also degradation is substantially balanced. Final products, namely intracellular FFAs, from membrane lipid degradation or hydrolysis, are produced by lipase A catalysis encoded by lipA or sll1969 in Synechocystis 6803 [16]. In addition, the intracellular FFA level is regulated to lower its toxicity to the cells by both the FFAs recycling reaction catalyzed by fatty acyl-ACP synthetase (encoded by aas or slr1609 in Synechocystis 6803) for generating fatty acyl-ACP, and FFAs secretion [4,6]. On the other flow direction of acetyl-CoA (Figure 1), a synthesis of bioplastic polyhydroxybutyrate (or PHB) is inducible via acetyl-CoA acetyltransferase (phaA) catalysis and accumulated as neutral lipid granules in Synechocystis, in particular under nutrient deprived-conditions [28,29].
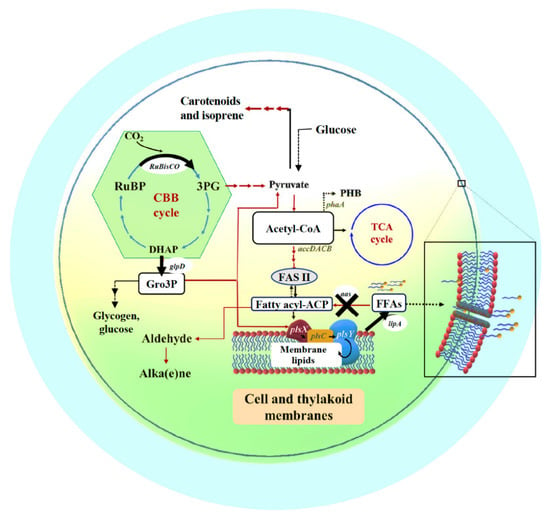
Figure 1.
Outline of intracellular lipid production and FFA secretion into cultured medium system in cyanobacteria. Key enzyme-encoding genes (represented in italic letters) associated with lipid and fatty acid synthesis, the Calvin–Benson–Bassham (CBB) cycle, polyhydroxybutyrate (PHB) synthesis, and free fatty acid (FFA) recycling, including accDACB, a multi-subunit acetyl-CoA carboxylase gene; lipA, a lipolytic enzyme-encoding gene; aas, acyl-ACP synthetase gene; plsX, plsY and plsC, putative phosphate acyl-transferases genes; and phaA, acetyl-CoA acetyltransferase gene; the RuBisCO gene cluster, including rbcLXS, encoding RuBisCO in respective order of large, chaperone and small subunits; glpD, glycerol-3-phosphate dehydrogenase gene. The abbreviated intermediates and products are: DHAP, dihydroxyacetone phosphate; fatty acyl-ACP, fatty acyl–acyl carrier protein; FFAs, free fatty acids; Gro3P, glycerol-3-phosphate; 3PG, 3-phosphoglycerate; PHB, polyhydroxybutyrate; RuBP, ribulose-1,5-bisphosphate. The thick arrows represent the overexpression (O) of indicated genes whereas the cross symbol represents knockout (K) of indicated gene.
In this study, we performed an integrative design of three engineered strains of Synechocystis sp. PCC 6803. (1) KA strain, the disruption of the aas gene encoding acyl-ACP synthetase that blocks FFAs recycling, (2) KAOL strain, KA with the overexpression of lipA which increases the membrane lipid degradation, and (3) KAOGR strain, KA with the overexpression of the RuBisCO operon (rbcL, rbcX and rbcS) and glpD encoding glycerol-3-phosphate dehydrogenase, which enhance the carbon supply from the CBB cycle via Gro3P generation for fatty-acid and membrane-lipid productions (Figure 1). We reported that all single and integrated KA strains secreted a higher content of extracellular FFAs when compared to that by wild-type cells, in particular the KAOL strain has an increased production.
2. Results and Discussion
2.1. Engineered Constructs of Synechocystis sp. PCC 6803 Lacking aas (KA), and Its Hybrids with Single Overexpression of LipA (KAOL) and Quadruple Overexpression of glpD/rbcLXS (KAOGR)
Our results showed that the inactivation of acyl-ACP synthetase (aas), which prevented FFA recycling to fatty acyl-ACP, secreted more free fatty acids (FFAs) into the culture medium, in agreement with earlier reports in cyanobacteria [15,16,17]. Then, the aas-overexpression in Synechocystis sp. PCC 6803 (hereafter, Synechocystis) conversely resulted in a significant decrease of secreted FFAs content [6]. In the current study, we then aimed to improve the FFA secretion by creating Synechocystis cells lacking aas (or KA) as a base for two more constructs. We examined the impact of either directly elevated FFAs levels via the increased membrane lipid hydrolysis by single lipA-overexpression in the KA strain (or the KAOL strain), and the indirect elevated carbon-substrate supply via the CBB cycle for FFA synthesis by quadruple GlpD/rbcLXS overexpression in the KA strain (KAOGR) thereby increasing the FFA secretion into the surrounding medium (Figure 1).
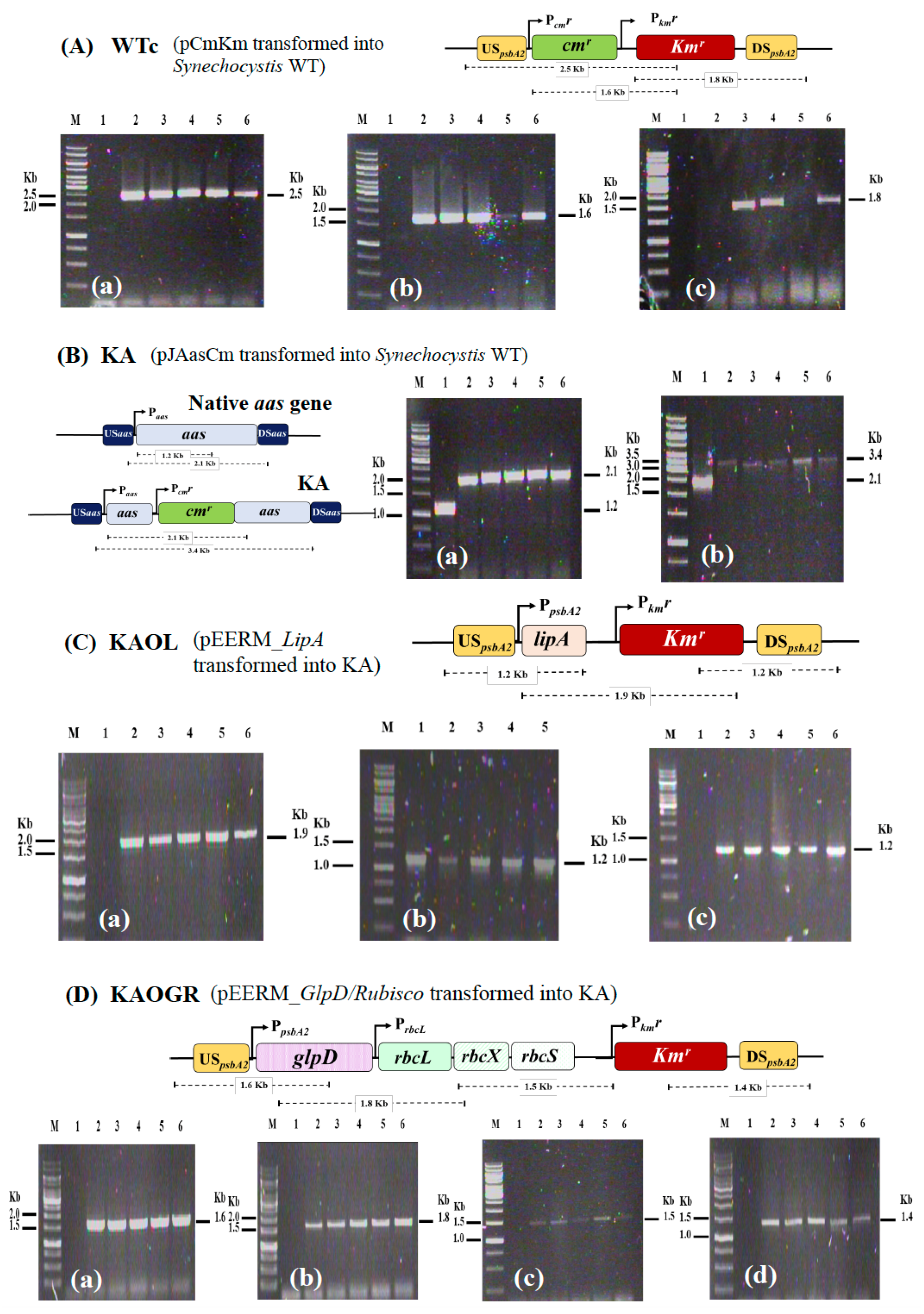
We initially constructed a Synechocystis strain lacking aas or KA (Table 1). To gain reliable results concerning antibiotic cmr and kmr effects, we constructed Synechocystis WT control (or WTc) by generating a pEERM_CmKm vector containing cassettes of cmr and kmr which was consequently transformed into Synechocystis WT cells. Those cassettes of cmr and kmr genes were integrated into Synechocystis WT genome at the native psbA2 location via double homologous recombination (Figure 2A). The KA strain was constructed by inserting cmr cassette to disrupt the aas gene (Figure 2B). Moreover, the metabolic engineering approach was employed by integrating a recombinant plasmid containing lipA or glpD/rbcLXS genes to KA host strain, generating the KAOL and KAOGR strains, respectively. Those lipA- and glpD/rbcLXS-overexpressing strains were created by transforming pEERM_LipA and pEERM_GlpD/rbcLXS recombinants into KA host cells via double homologous recombination, and generated KA hybrid strains including the KAOL and KAOGR strains, respectively (Figure 2C,D). Then, its complete segregation was confirmed by PCR analysis using specific pairs of primers (Supplementary Materials, Table S1).

Table 1.
Strains and plasmids used in this study.
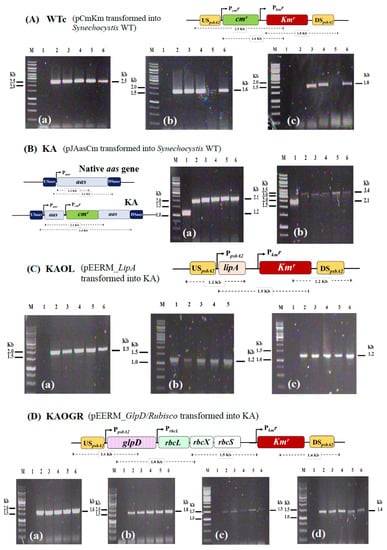
Figure 2.
Genomic maps of engineered Synechocystis PCC 6803 strains. The four modified strains are WTc (A), KA (B), KAOL (C), and KAOGR (D). The specific primers (Supplementary Materials, Table S1) were used to confirm the integration of each gene into Synechocystis genome. WTc is a wild-type control made by transforming both antibiotic chloramphenicol and kanamycin resistance cassettes (cmr and kmr, respectively) into WT strain. KA was constructed by inserting a cmr cassette to disrupt the aas gene. The integration of each strain was confirmed using PCR with genomic DNA from WT strain as the template. Lane M: GeneRuler DNA ladder (Fermentas). For (A) WTc strain; Lane 1: negative control using WTc as template (a–c), (a) Lanes 2–6: clone numbers 1 to 5 using UUSpsbA2 and Km_SR primers, (b) Lanes 2–6: clone numbers 1 to 5 using Cm_F and Km_SR primers, and (c) Lanes 2–6: clone numbers 1 to 5 using Km_FBamHI and DDSpsbA2 primers. For (B) KA strain; Lane 1: negative control using WTc as template (a,b), (a) Lanes 2–6: clone numbers 1 to 5 using Aas_F3 and aas_SR primers, and (b) Lanes 2–6: clone numbers 1 to 5 using USaas and DSaas primers. Only positive clones were selected for subsequent experiments; (A) WTc: clone numbers 2, 3, 5, (B) KA: clone numbers 1, 3, 4, 5, (C) KAOL: clone numbers 1–4, and (D) KAOGR: clone numbers 1–5.
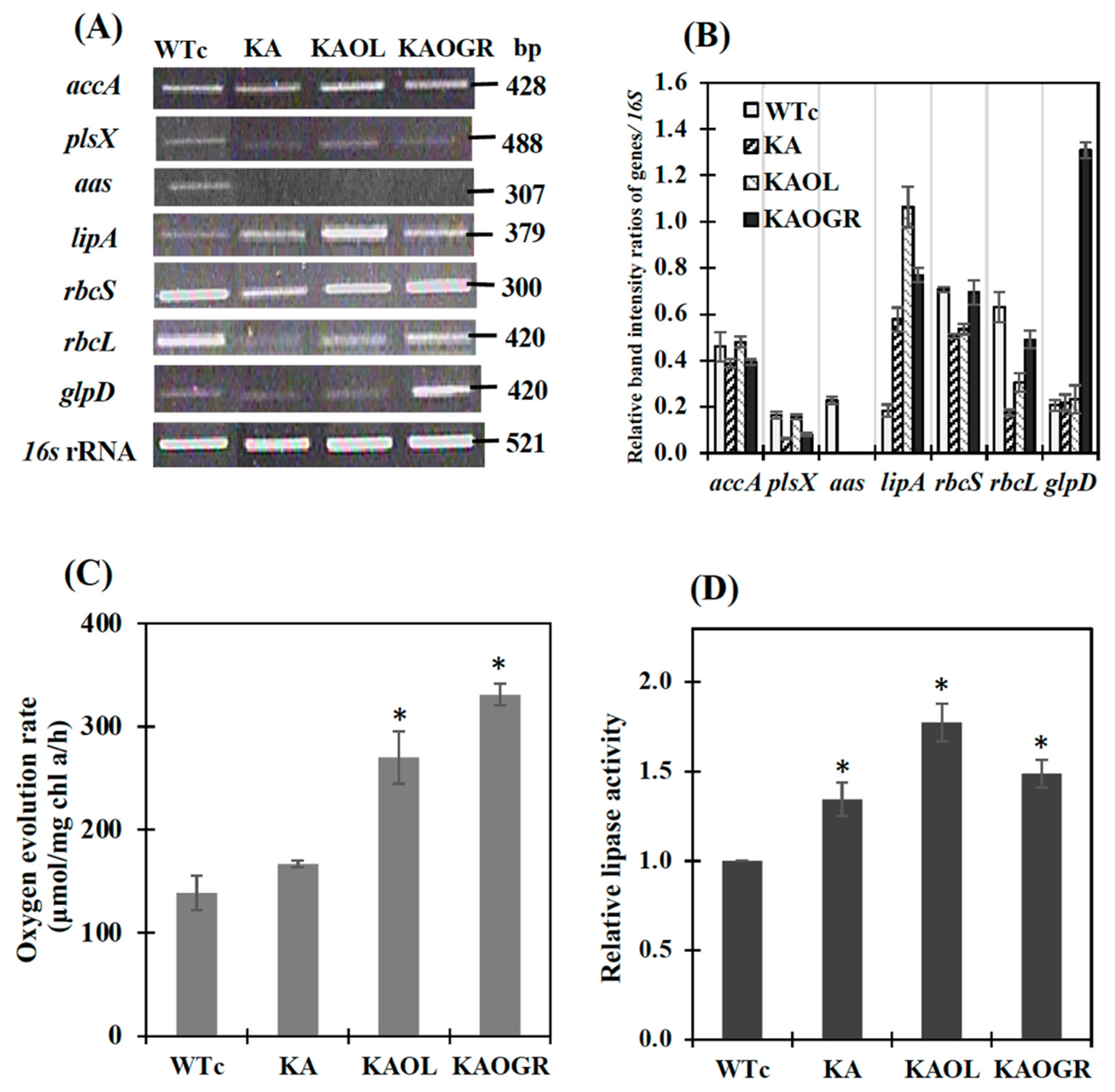
We determined the relative transcript levels by using the RT-PCR method (Figure 3A,B). The noted transcript levels of rbcS and rbcL encoding RuBisCO in the CBB cycle were decreased in KA strain (Figure 3) when compared to those of WTc and of WT cells [4]. In the KAOL strain, the transcript level of lipA was clearly increased. Likewise, the apparent increases in glpD, rbcS and rbcL transcript levels were confirmed in the KAOGR strain. Moreover, a significant increase in the transcript levels of accA, plsX, rbcS and rbcL occurred in the KAOL strain when compared with those of KA host. In contrast, only a slight increase in accA and plsX transcript levels was observed in the KAOGR strain (Figure 3A). When compared with WTc, the noted decreases in accA and plsX transcript levels in KA and KAOGR suggested the reduced effect on fatty acid and phospholipid syntheses, while the increased effect on membrane hydrolysis occurred with higher lipA transcript levels (Figure 3B). As expected, a significant increase in the oxygen evolution rate, which represents photosynthetic efficiency, was observed in the KAOGR with overexpressing RuBisCO and KAOL strains compared to the WTc (Figure 3C). The highest level of lipase activity was found in the KAOL strain with lipA overexpression (Figure 3D), in agreement with its increased transcript level (Figure 3A,B).
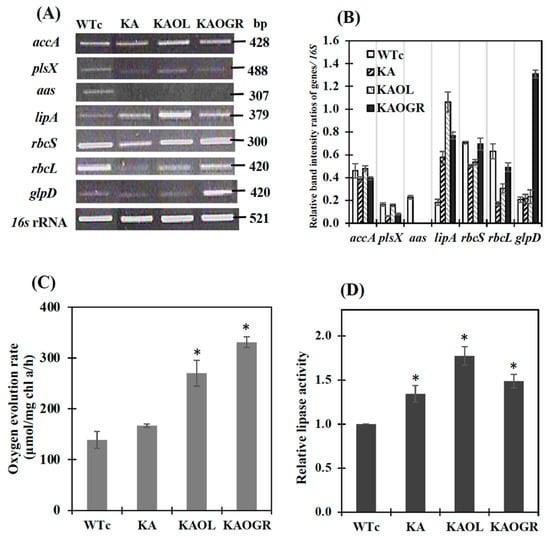
Figure 3.
The transcript levels (A) and relative transcript intensity ratios (B) of the accA, plsX, aas, lipA, rbcS, rbcL, glpD and 16S rRNA genes in Synechocystis PCC 6803 strains WTc, KA, KAOL and KAOGR. Oxygen evolution rate (C) and relative lipase activity (D) of all strains. Cells were grown in BG11 medium and analyzed at day 5 of cultivation. The error bars represent standard deviations of means (mean ± S.D., n = 3). For (A), agarose gel (1.2%) of PCR product stained by RedSafeTM nucleic acid staining solution (Intron Biotechnology Inc., Korea). For (D), control data (WTc) at 1.0 is derived from the lipase activity of 0.22 U/mg protein. For (C) and (D), the statistical differences of the results between WTc and engineered strain are indicated by an asterisk *, p < 0.05.
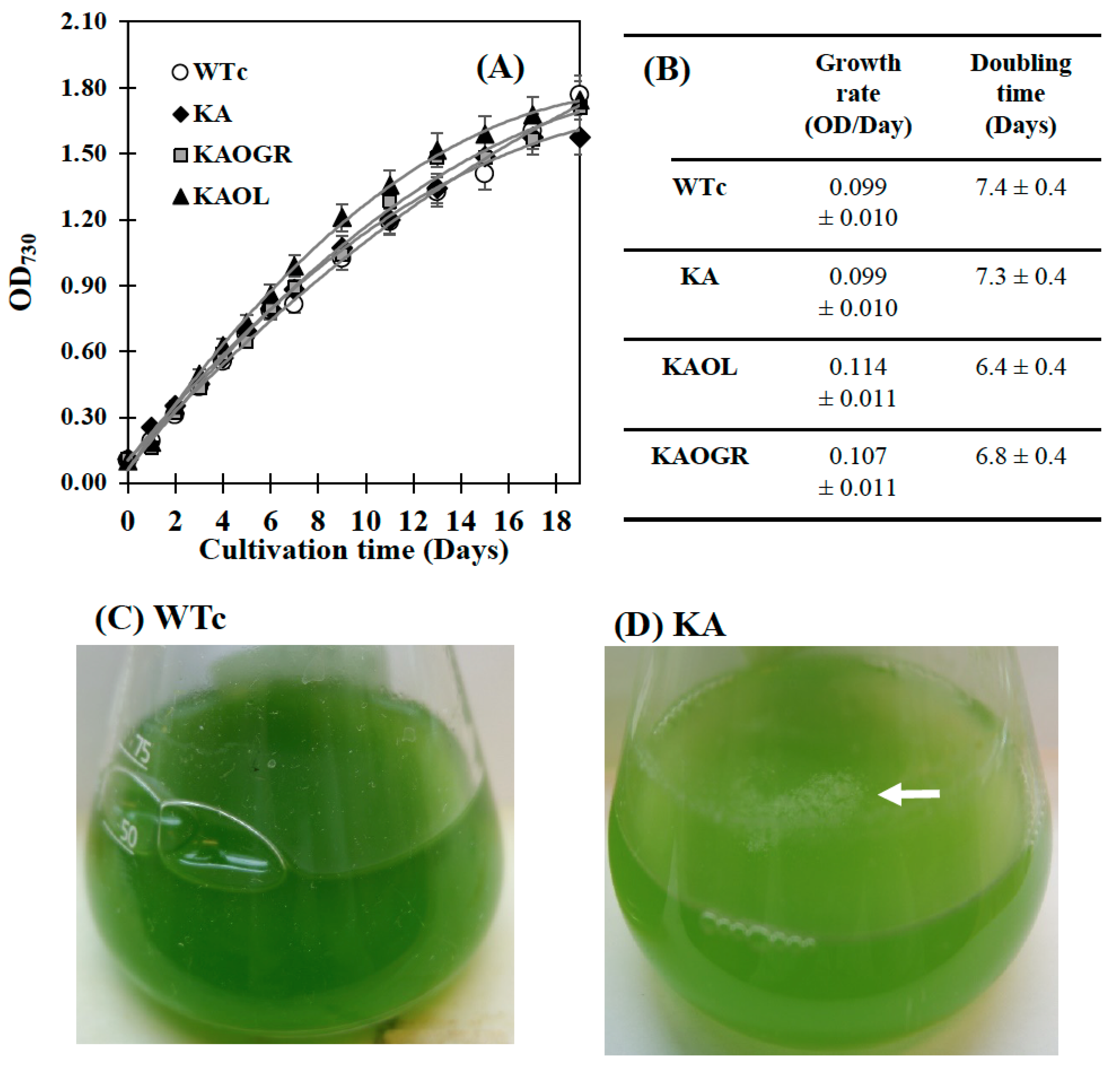
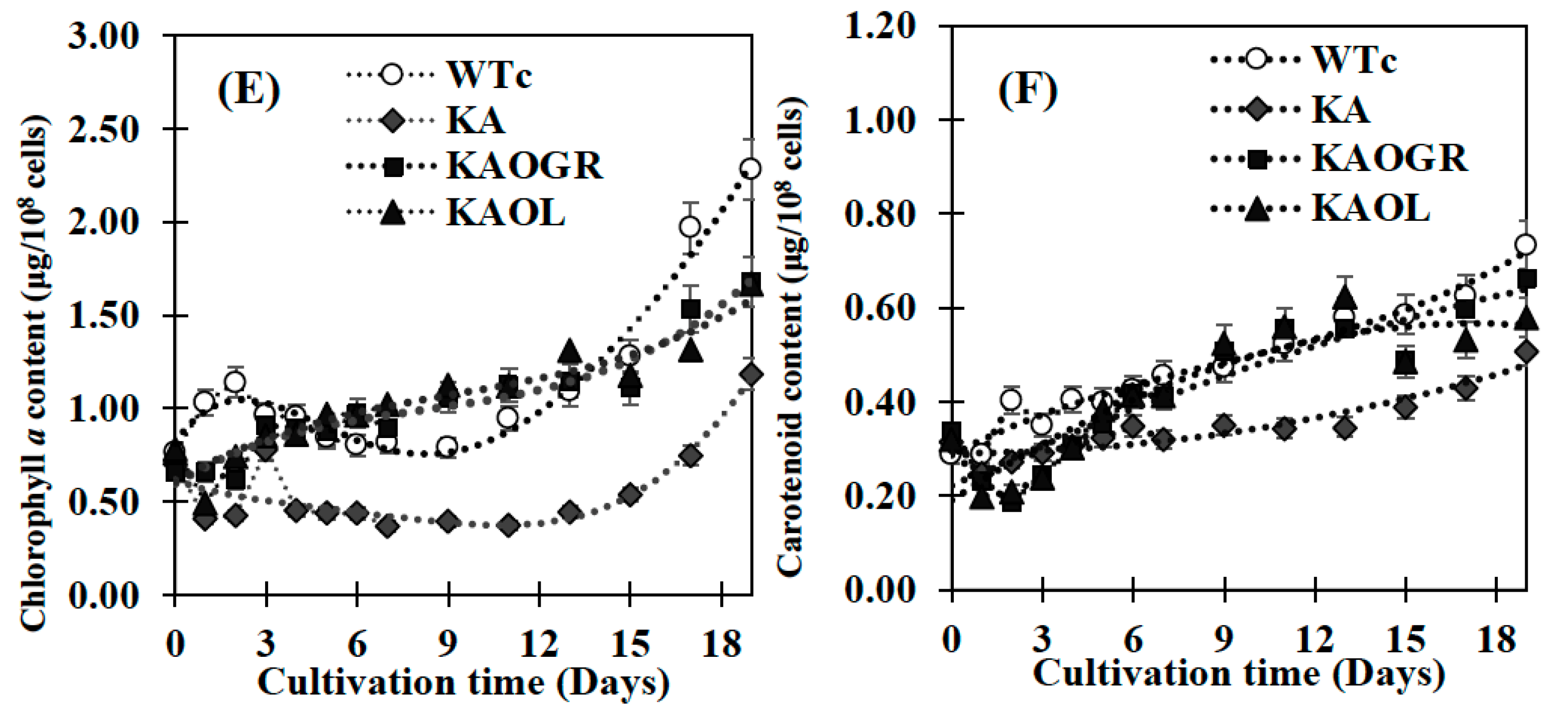
The engineered WTc strain notably contained similar properties of growth and lipid production as WT cells [4]. Although the KA strain had a similar growth tendency to WTc, its intracellular pigment contents of chlorophyll a and carotenoids were markedly decreased when compared with WTc (Figure 4A–F). For qualitative results, digital images of the KA culture flask showed a lighter green color compared with the dark green of WTc cell cultures (Figure 4C,D). In addition, the white droplets floating on the surface of KA culture medium (Figure 4D), represented secreted FFAs, clearly visualized when compared with WTc without white droplets (Figure 4C). On the other hand, it was observed that the KA strain contained lower chlorophyll a and carotenoids accumulations than those of WTc cells (Figure 4E,F), in agreement with lowered transcript levels of rbcS and rbcL encoding RuBisCO enzyme in the CBB cycle (Figure 3A). We assume that RuBisCO substantially contributed to the accumulation patterns of chlorophyll a and carotenoids, in particular, carotenoids which relate to the pyruvate precursor partly converted from Gro3P ([27], and Figure 1). However, in respect to the lowered relative rbcS and rbcL transcript levels in the KA strain, although the oxygen evolution rate of the KA strain was similar to the WTc (Figure 3C), primary or secondary effects, such as RuBisCo activity or RuBP regeneration should be clarified for better understanding of its regulation. KAOGR, and specifically the KAOL strain, had slight increases in growth when compared with the WTc (Figure 4A), the latter clearly having a shorter doubling time (Figure 4B). However, the contents of chlorophyll a and carotenoids in the KAOL and KAOGR strains were slightly lower than in the WTc under normal conditions after 12 days of cultivation (Figure 4E,F).

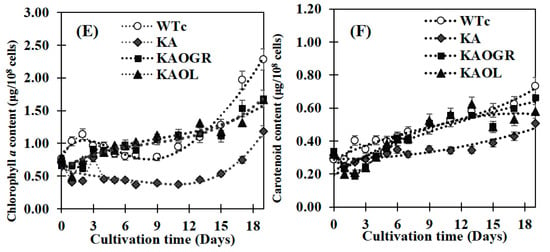
Figure 4.
Growth curve (A), growth rate and doubling time (B), digital images of cell cultures (C,D), chlorophyll a content (E) and carotenoid content (F) of the Synechocystis PCC6803 wild-type control (WTc) and engineered KA strains. Cells were grown in BG11 medium for 19 days. Error bars represent standard deviations of means (mean ± S.D., n = 3). For (C,D), images of 10-day cell culture flasks of WTc and KA were depicted. Floating white droplets in KA-cultured flask represent free fatty acids (FFAs) secreted to the medium (white arrow).
2.2. Production of Intracellular Lipids qnd Extracellular FFAs in Synechocystis sp. PCC 6803 Wild-Type Control and Engineered Strains
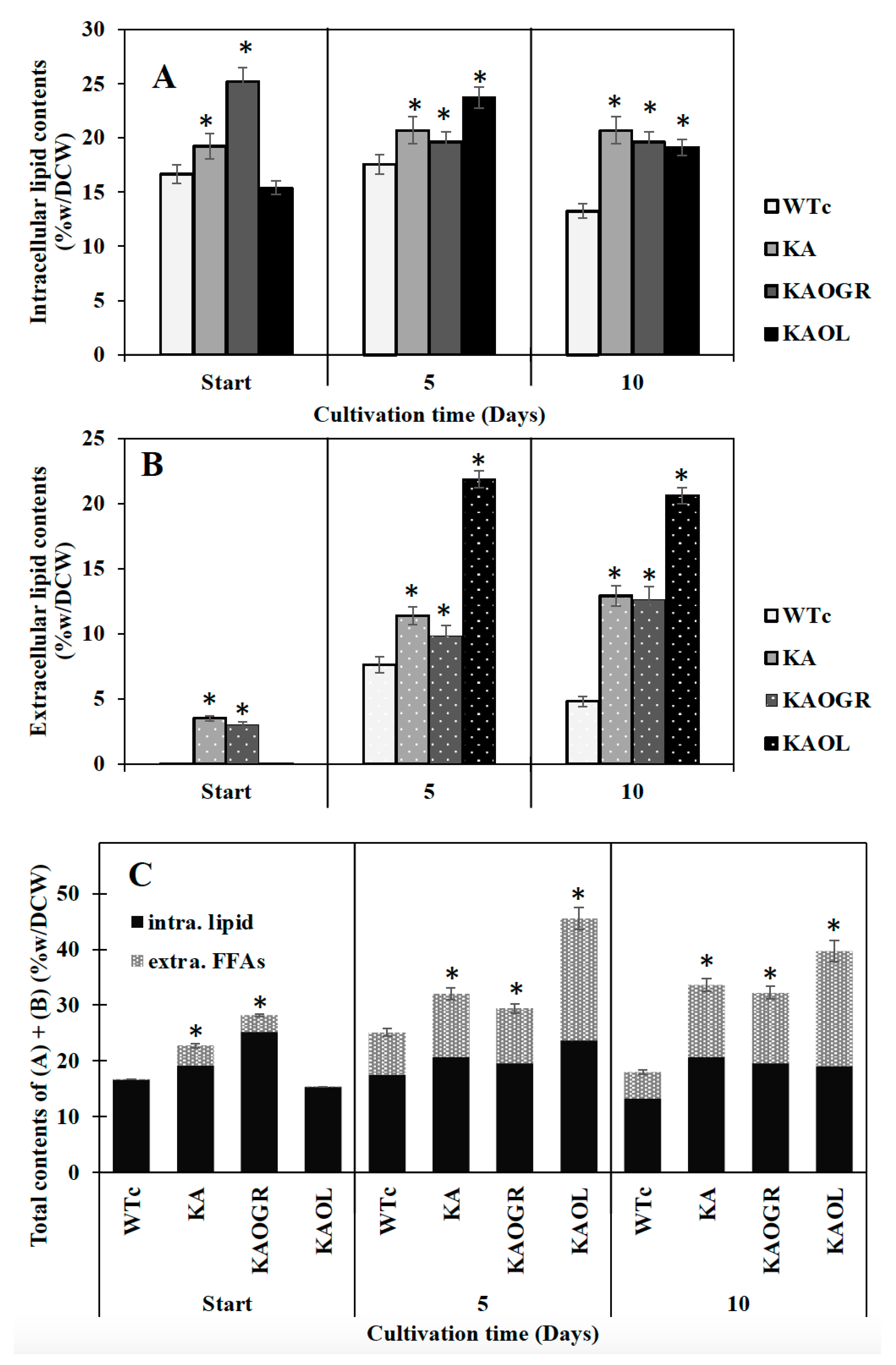
Recently, radiolabeled FAs, thin layer chromatography (TLC) and gas chromatography (GC) have been employed to establish the intracellular and extracellular fractions of Synechocystis sp. PCC6803 containing FAs. A significant release of FFAs from membrane lipids to the surrounding culture medium was demonstrated [15]. Moreover, the white deposits or droplets of secreted FFAs in the culture medium of Synechocystis sp. PCC6803 were tangibly identified by GC/GC–MS [4,8,20]. In this study, although KAOGR showed the highest amount of intracellular lipid content at the start point, we experimentally ascertained that both intracellular lipid and extracellular FFAs contents of the KA strain were practically higher than those of the WTc by a constant lipid accumulation of about 20.7%w/DCW during 5–10 days of cultivation, whereas the extracellular FFA content reached up to 12.9%w/DCW at day 10 (Figure 5A,B). Consistent parameters of titer (mg/L) and production rate (mg/L/day) of KA strain were noted, corresponding to their respective contents (%w/DCW) of intracellular lipids and extracellular FFAs (Supplementary Materials, Table S2). Moreover, when compared among the three KA strains, the KAOL strain clearly contained the highest levels of both intracellular lipids, and showed the highest secreted FFAs at 5 days of cultivation, at 23.7 and 21.9%w/DCW, respectively (Figure 5A). This also holds for total contents of both intracellular lipids and extracellular FFAs (Figure 5C). Interestingly, the titer and production rates of the KAOL strain resulted in the highest levels of extracellular FFAs fractions at days 5 and 10 of cultivation (Supplementary Materials, Table S2).
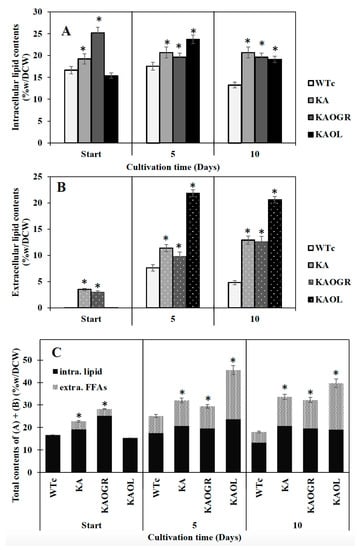
Figure 5.
Contents (%w/DCW) of intracellular lipid (A) and extracellular FFAs (B) and total contents of intracellular lipid and extracellular FFAs (C) in Synechocystis PCC 6803 strains WTc, KA, KAOL and KAOGR. Cells were grown in BG11 medium at the start of the experiment (day 0) and analysed at days 5 and day 10 of cultivation. Error bars represent standard deviations of means (mean ± S.D., n = 3). The statistical differences of the results between the WTc and engineered strains are indicated by an asterisk *, p < 0.05.
Recent observations on increased lipid synthesis have shown that high lipid content occurred during the exponential growth phase and remained steady or decreased during the late-log and stationary phases [4,30]. The interplay of many biochemical pathways to maintain lipid homeostasis includes fatty acid and membrane lipid synthases, membrane lipid degradation, FFA recycling and FFA secretion. Most lipid molecules in cyanobacteria consist of four major types including monogalactosyl diacylglycerol (MGDG), galactosyl diacylglycerol (DGDG), sulfoquinovosyl diacylglycerol (SQDG) and phosphatidylglycerol (PG), located in both the cytoplasmic and thylakoid membranes [31]. Hence, membrane lipids could be hydrolyzed into free fatty acids (FFAs) by lipase A encoded by sll1969 in Synechocystis PCC 6803 [16]. Those FFAs products are either partly secreted out of cells into the medium or are incorporated into membrane lipids’ biosynthesis via FFA recycling [32,33,34,35]. In the current study, we demonstrated that the lipA overexpression in the KA strain (KAOL) impacts the FFA secretion, with the highest level reaching 21.9%w/DCW at day 5 of cultivation (Figure 5), rather than that of KAOGR, a KA strain with overexpressing glpD/rbcLXS genes. Therefore, we reason that the regulation of FFA secretion may have a direct correlation with intracellular FFA yields, obtained either from the degradation of membrane lipids or the disturbance of FFAs recycling. This is in agreement with a previous strategy in which the prominent FFA secretion could be enhanced by weakening the cell-wall layers [8,36].
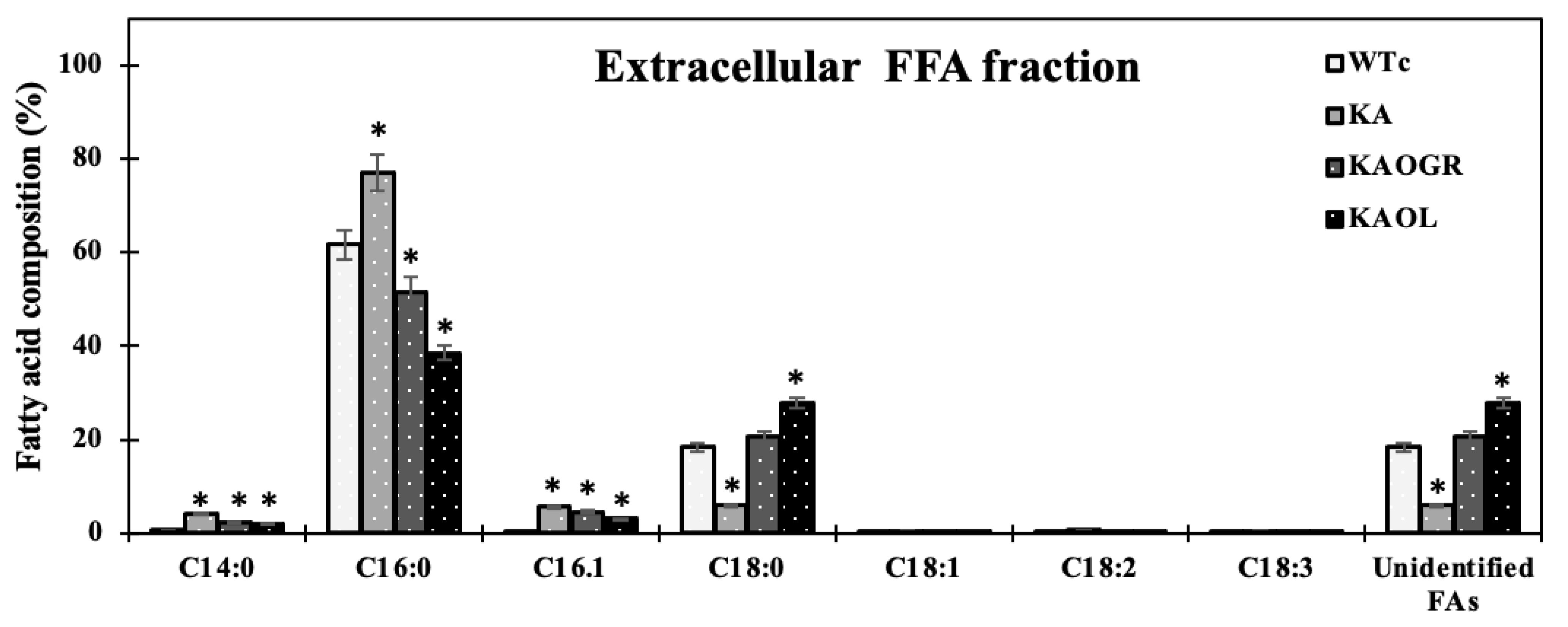
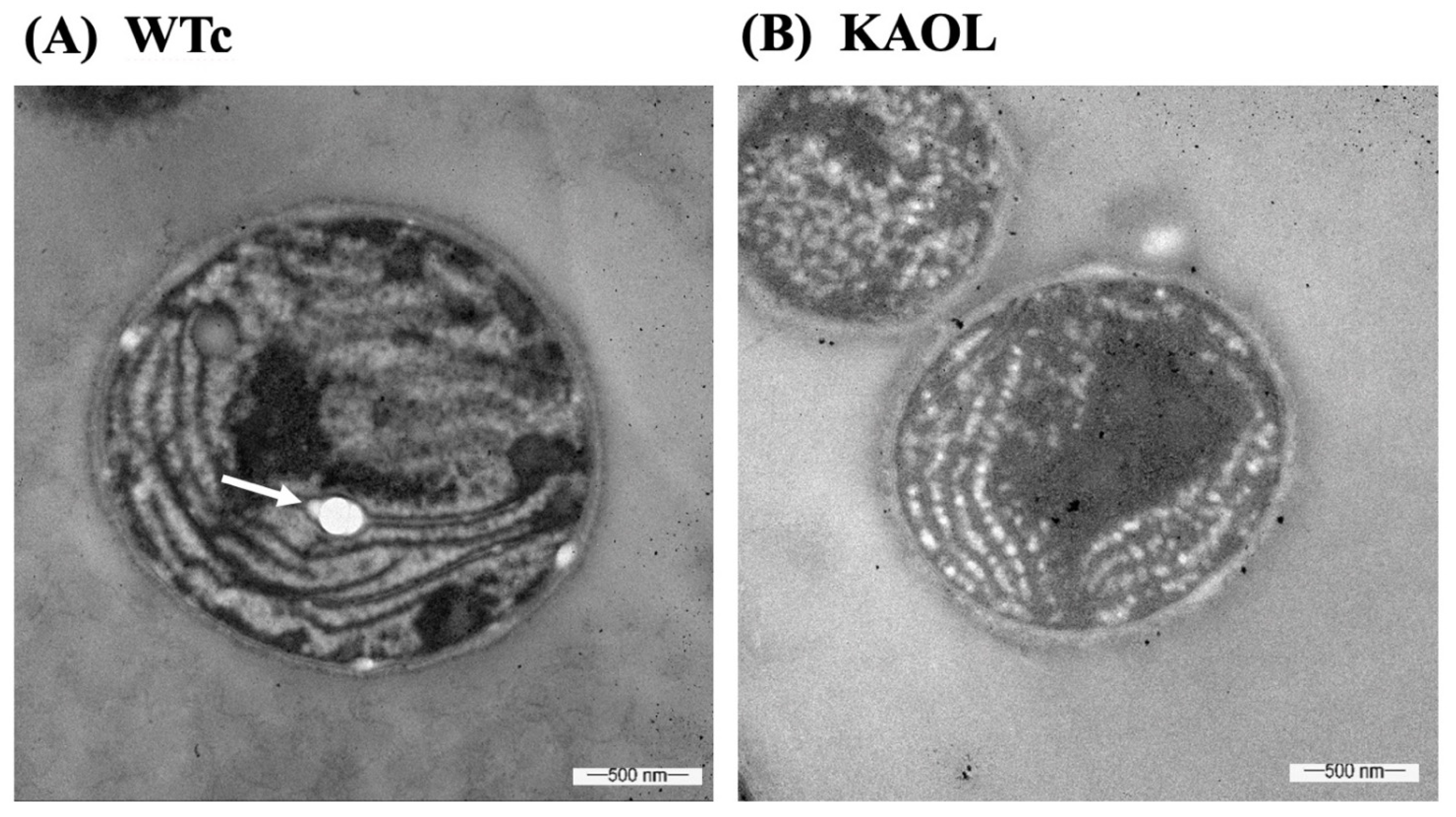
Moreover, we also measured fatty acid (FA) compositions of extracellular FFA fraction by GC. For the KA strain, higher FA proportions were noted for myristic acid (C14:0), palmitic acid (C16:0), and palmitoleic acid (C16:1) (Figure 6). With the different FA compositions in the extracellular fraction of secreted FFAs, strains KAOL and KAOGR secreted higher proportions of stearic acid (C18:0), whereas a lower proportion of palmitic acid (C16:0) was noted when compared to both the KA and WTc strains. Interestingly, the relative contents of myristic acid (C14:0), palmitoleic acid (16:1), stearic acid (C18:0), and unidentified FAs, which may be longer than C18-Fas, were secreted into the medium in higher levels than those of intracellular FA pool. Certain structures of C16:0 and C18:0 molecules are more hydrophobic, which enabled them to more easily diffuse through cytoplasmic and thylakoid membranes [4]. The higher proportion of palmitoleic acid (16:1) in the extracellular FFA fraction was previously demonstrated for Synechococcus elongatus and Synechocystis sp. PCC 6803 [18]. For the qualitative images by TEM comparing the KAOL strain (Figure 7), the highest lipid producing strain, an abundant number of droplet-like bright dots located on regions of thylakoid membranes were visualized in the cells of the KAOL strain (Figure 7A,B). Interestingly, when we observed the cell membrane layers, the KAOL strain contained irregular and rough surfaces (Figure 7B) when compared with more regular and smooth cell surfaces of WTc cells (Figure 7A). We speculate that this rough surface of KAOL cells may support FFAs secretion.
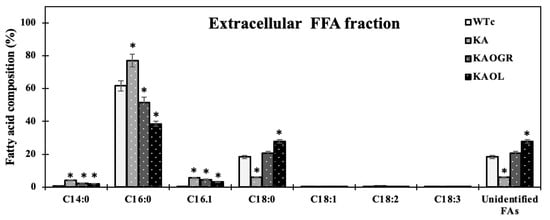
Figure 6.
Fatty acid compositions (%) in extracellular fraction analysed by a GC instrument in Synechocystis PCC 6803 WTc and all engineered strains. Cells were grown in BG11 medium for 10 days before being analysed. Error bars represent standard deviations of means (mean ± S.D., n = 3). The statistical differences of the results between the WTc and engineered strains are indicated by an asterisk *, p < 0.05.
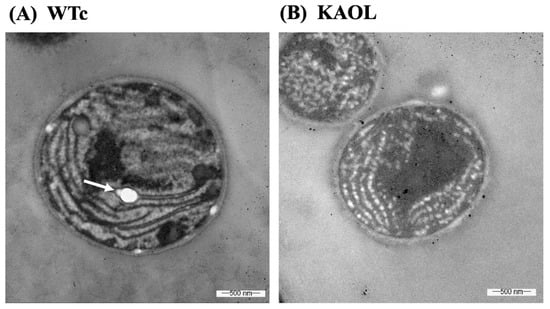
Figure 7.
Transmission electron micrographs (TEMs) of Synechocystis WTc (A) and KAOL (B) strains under normal BG11 medium for 5 days of cultivation. In (A), WTc images are shown in 40,000× magnifications. In (B), KAOL images are shown in 50,000× magnifications. In (A), the white dots (white arrow) represent the examples of electron-transparent bodies. In (B), abundant droplet-like white dots were located on regions of thylakoid membranes inside KAOL cells.
Excess of FFAs may cause toxicity and cell damage by intercalating their amphipathic structure into both cytoplasmic and thylakoid membranes [7,18]. As consequences of this, destroyed thylakoid pigments and destabilized membrane proteins detrimentally occur, as well as the disruption of the electron transport chain in photosynthesis of cyanobacteria [8,37,38]. Additionally, unsaturated fatty acids (UFAs), with double bonds, react with reactive oxygen species (ROS) and generate the toxic free-radical products which eventually damage the cell [39,40]. In this study, we observed that although the KA strain with FFA recycling blocked had similar growth and photosynthetic efficiency to the WTc, significant reductions in chlorophyll a and carotenoids occurred (Figure 4). These results may suggest that intracellular excess of FFAs which were not secreted out of cells can generate harmful effects on the intracellular pigments. However, lipA- and glpD/rbcLXS-overexpressions in KA strains, KAOL and KAOGR, may alleviate those negative consequences on the cell growth and retain regular pigment levels with higher photosynthetic efficiencies. Increased biomass yields of Synechocystis PCC 6803 were recently reported in the genetically modified strain, overexpressing either the RuBisCO operon [41], or glpD_RuBisCO [4] by generating increased carbon flow through the CBB cycle. On the other hand, it was surprising that the excess of FFAs in the KAOL strain had no negative effect on cell growth, oxygen evolution rate and intracellular pigment accumulations. Since there is no annotated fatty acid degradation pathway in Synechocystis [35], we propose that any excessive FFAs which are not secreted out of cells may be preferentially stored inside the cell by incorporation into higher intracellular lipids. Moreover, several droplet-like white granules can be identified inside the KAOL cells (Figure 7). Previously, an accumulation of lipid droplets, containing mainly saturated C16 and C18 fatty acids, was found in the cyanobacterium Nostoc punctiforme, induced when the cells consumed nitrate as their source of nitrogen during the log phase of cell growth [42]. We also demonstrated the polyhydroxybutyrate (PHB) contents and visualized Nile-red-stained PHB granules in both strains (Figure 8A and Supplementary Materials, Figure S1). It is worth noting that both WTc and KAOL cells contained less PHB granules under normal growth condition, visualized by fluorescence microscope (Supplementary Materials, Figure S1). This corresponds with a lower PHB content under normal BG11 conditions, detected by HPLC (Figure 8A). This obvious augmentation of droplet-like white spots then represents a crucial adaptation to significant membrane-lipids degradation, intracellularly generated by lipA overexpression.
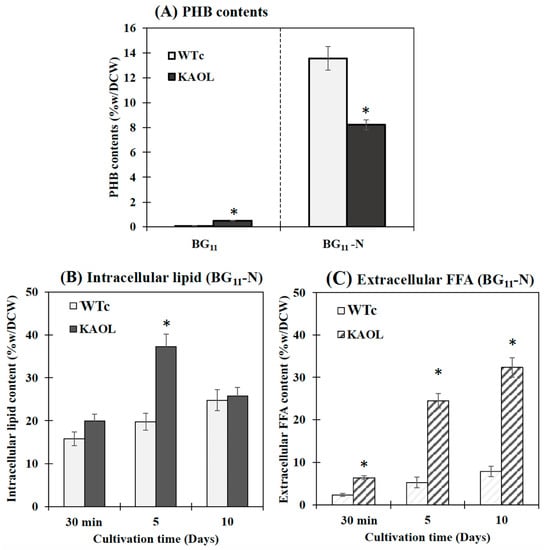
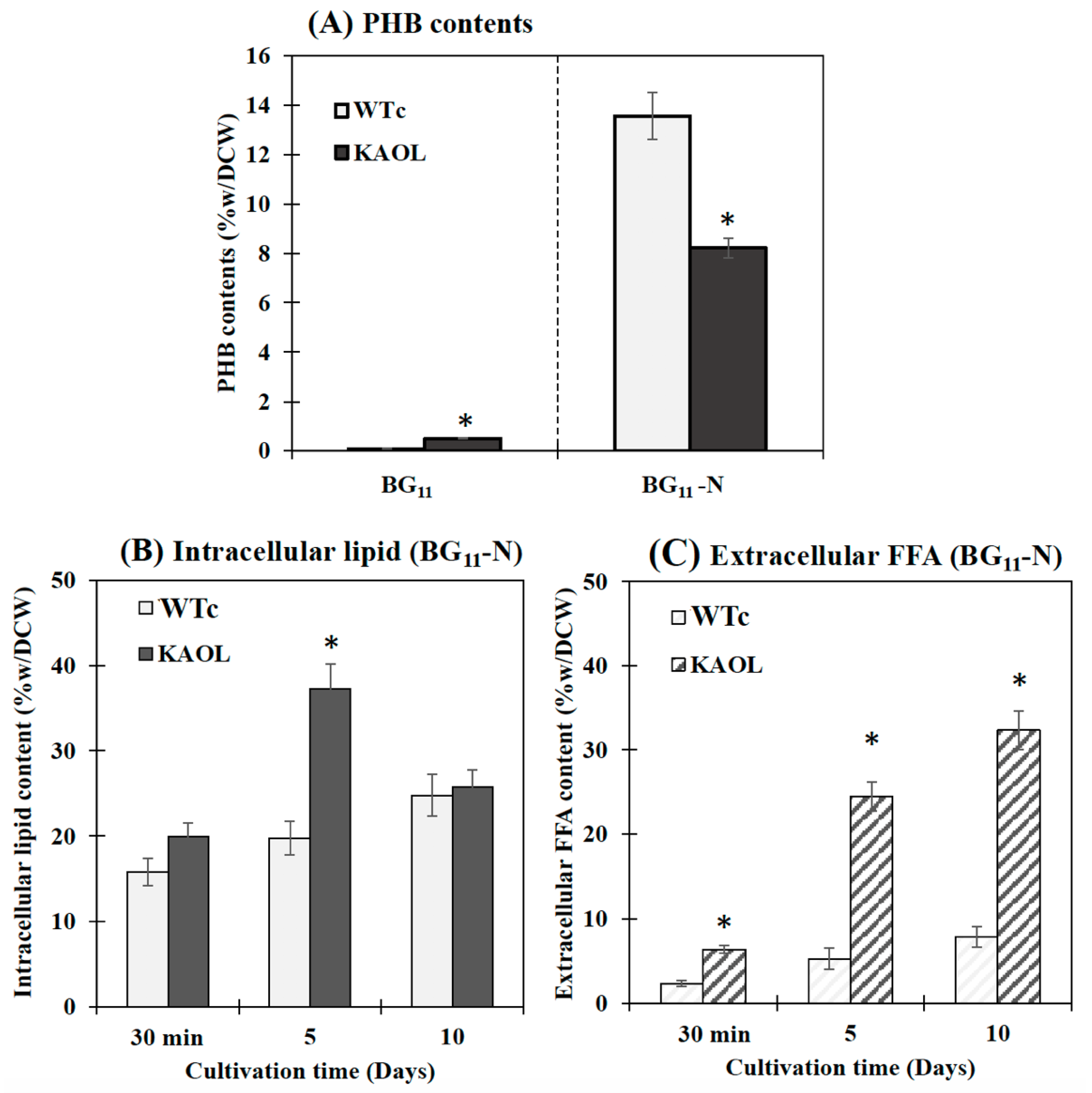
Figure 8.
Contents (%w/DCW) of PHB (A), intracellular lipid (B) and extracellular FFAs (C) in Synechocystis PCC 6803 WTc and KAOL strains under BG11-N condition. For (A), cells were adapted for 5 days under BG11 and BG11-N conditions, respectively, before being analysed. For (B,C), cells were grown in BG11 medium lacking NaNO3 (BG11-N) at 30 min, day 5 and day 10 of cultivations. Error bars represent standard deviations of means (mean ± S.D., n = 3). The statistical difference of the results between the WTc and KAOL strains is indicated by an asterisk at * p < 0.05.
2.3. NaNO3 Depletion Induces Higher Levels of Intracellular Lipids and Extracellular FFAs
It was noted that Synechocystis PCC 6803 WTc and KAOL cell cultures grown in BG11 lacking NaNO3 (or BG11-N) showed higher PHB production when compared to BG11 condition, demonstrated by an increased number of PHB granules (Supplementary Materials, Figure S1) and PHB content by about 13.6 and 8.2%w/DCW, respectively (Figure 8). This well-known strategy of nutrient deprivation was previously reported for PHB induction in Synechocystis sp. PCC 6803 [28,29]. However, it is also revealed that the KAOL strain showed lower PHB content than the WTc under BG11-N condition after 5 days (Figure 8A). We propose that this lower PHB content in the KAOL strain may partially inverse with a higher lipid content flow (see Figure 1). In short, KAOL cells showed higher contents of both intracellular lipids and extracellular FFAs under the BG11-N condition when compared to those observed in WTc cells (Figure 8B,C). The highest intracellular lipid level of the KAOL strain was 37.2%w/DCW at day 5, a 1.6-fold increase compared to that under BG11 condition (23.7%w/DCW). On the other hand, the secreted FFAs content of the KAOL strain significantly increased at days 5 and 10, reaching 24.5 and 32.3%w/DCW, respectively. This represents a 1.1- and 1.6-fold increase compared to those under BG11 condition (Figure 8C). Our findings support a conclusion that N-deprived conditions induce higher accumulations of intracellular lipids and secreted FFA in both Synechocystis wild-type and engineered strains, with overexpression of lipA in combination with a disruption of aas. Additionally, glycogen assimilation and degradation should be considered as competing for acetyl-CoA utilization, specifically in the KAOGR strain. An increased level of glgX encoding a glycogen debranching enzyme occurred in a high PHB-overproducing Synechocystis strain with overexpression of native pha gene [29].
3. Materials and Methods
3.1. Construction of Recombinant Plasmids
The recombinant plasmids, including pEERM_CmKm, pEERM_GlpD/rbcLXS, pJAasCm and pEERM_lipA (Table 1) were used to generate all strains including an aas-knock out Synechocystis (KA), glpD/rbcLXS-overexpressing KA (KAOGR) and lipA-overexpressing KA (KAOL). The recombinant pJAasCm vector was created by firstly ligating the aas gene fragment amplified by PCR with genomic DNA as a template, using aas_F3 and aas_R3 as primers (Supplementary Materials, Table S1) into the pJET1.2 blunt cloning vector. Secondly, the antibiotic cmr cassette fragment was amplified by cm_F and cm_R primers (Supplementary Materials, Table S1) using a pEERM plasmid [43] as the template. This cmr gene fragment was subsequently inserted onto the AvrII restriction site of the aas gene which earlier ligated into the pJET1.2 blunt cloning vector. On the other hand, the construction of the recombinant pEERM_LipA plasmid was performed by inserting the lipA gene fragment amplified by PCR using lipA_F and lipA_R as primers (Supplementary Materials, Table S1) inbetween the restriction sites of XbaI and SpeI in the pEERM_Km vector [4]. After that, the recombinant plasmids were confirmed by sequencing using specific primers as shown in Supplementary Materials, Table S1.
3.2. Natural Transformation of Recombinant Plasmids into Synechocystis Cells
Initially, the recombinant pEERM_CmKm and pJAasCm plasmids were separately transformed into the Synechocystis sp. PCC 6803 wild-type (WT) strain by natural transformation [6] in order to obtain the control WT (WTc) and an aas-knockout (KA) strains, respectively. Those WTc and KA strains were subsequently used as host strains. For pEERM_glpD/rbcLXS and pEERM_lipA recombinant plasmids, they were separately transformed into the KA host strain and finally the KAOGR and KAOL strains were obtained, respectively. The host cells were cultured in fresh BG11 medium and grown until the optical density at 730 nm (OD730) was about 0.3–0.5. Then, 50 mL of cell culture was harvested by centrifugation at 6000 rpm (4025 × g) for 10 min, and cell pellets were collected. To condense the cell suspension, fresh BG11 medium (0.5 mL) was subsequently added to suspended cell pellets. Next, 10 µg of recombinant plasmid was added into that condensed cell suspension and mixed gently by pipetting. The mixture was incubated at 28 °C for 6 h by inverting the mixture tube every 2 h under a continuous light intensity at 15 µE/m2s. After incubation, that mixture was smeared on a 0.45 µm sterile nitrocellulose membrane which was placed over a BG11 agar plate and incubated overnight, and then the membrane was transferred to be placed upon BG11 agar containing 35 µg/mL kanamycin, alone or combined with 35 µg/mL chloramphenicol. The surviving colonies were randomly picked from those plates after several weeks of incubating under normal growth conditions, and their gene locations and segregation were confirmed by PCR using specific pairs of primers, as shown in Supplementary Materials, Table S1.
3.3. Determinations of Cells Growth and Pigment Contents
Synechocystis WT and all engineered strains were initially cultivated on normal BG11 agar plate and selective BG11 agar plate containing antibiotic 35 µg/mL kanamycin and 35 µg/mL chloramphenicol, respectively. Normal growth condition was performed, at 28–32 °C under a continuous light illumination with an intensity of 40–50 µE/m2s. In order to prepare stock culture, colonies were transferred into a 250 mL-flask containing 100 mL of fresh liquid BG11 medium, and cultured until OD730 reaching 0.5–0.7 by shaking at 160 rpm on a rotary shaker. Cell culture was harvested by centrifugation at 6000 rpm (4025× g) for 10 min. Cell pellets were resuspended into fresh BG11 medium with initial OD730 at 0.1 and were subsequently cultured under normal growth condition for 16 days. One mL of cultured cells was collected to measure cell growth by spectrophotometrically measuring at OD730 [44]. For N-deprived conditions, cells at the mid-log phase of growth were harvested by centrifugation at 6000 rpm (4025× g) for 10 min and transferred into normal and modified media including normal BG11 medium (BG11 containing 17.6 mM NaNO3) and BG11 without NaNO3 (BG11-N). For pigment extraction, another 1 mL of cell culture was centrifuged at 6000 rpm (4025 g) for 10 min, and cell pellets were collected for pigment extraction using N,N-dimethylformamide (DMF). The DMF-extracted samples were subsequently measured for their absorbances at 461, 625 and 664 nm, respectively, by a spectrophotometer. The pigment contents were calculated and normalized by equations according to [45,46].
3.4. Lipid Extraction
During cultivation, 25 mL of cell culture of each strain was sampled and harvested by centrifugation at 6000 rpm (4025× g) for 10 min. The intracellular lipids and extracellular FFAs were separately obtained from cell pellets and supernatant fractions, respectively. The chloroform and methanol solvent extraction method was performed (modified from [47]). One mL of CHCL3: MeOH (ratio 2:1) solution was added into the cell pellet fraction whereas 5 mL of that solution was added into the supernatant fraction. The mixture was mixed by vortexing and incubated at 37 °C on a shaker for 2 h. Then, 0.5 mL of 0.88% w/v potassium chloride was added and the mixture was subsequently vortexed and left at room temperature for 5 min. The mixture was then centrifuged at 6000 rpm (4025× g) for 5 min, and the upper aqueous phase was removed, and the lower chloroform phase was collected and transferred to new glass tube. The lipid-containing chloroform phase was evaporated in fume hood at room temperature overnight to obtain total lipids.
3.5. Determinations of Intracellular Lipid and Extracellular FFA Contents
The extracted total lipids and secreted FFAs content were determined by the potassium dichromate oxidation reaction method [48]. Sequential additions of 0.18 M K2Cr2O7 (0.5 mL) and concentrated sulfuric acid (0.5 mL) were carried out and mixed with each sample by vortexing. The mixture was heated on a heat box at 100–105 °C for 30 min. After cooling down the reaction mixture to room temperature, distilled water (0.5 mL) was subsequently added. The absorbance of the reaction mixture was spectrophotometrically measured at 600 nm. Canola oil was used as a standard and prepared in the same way as the samples. The unit of lipid content was represented as % (w/DCW). Dried cell weight (DCW) was performed by drying the harvested cells in a 60–70 °C oven until obtaining a constant dry weight.
3.6. Determination of Fatty Acid (FA) Composition
Gas Chromatography (GC) was used for analyzing the fatty acid compositions from extracted lipids and FFAs in this study [4]. The fatty-acid methyl esters (FAMEs) were firstly prepared by adding 1 mL of 5%v/v hydrochloric acid in MeOH. The mixture reaction occurred by heating on a heat box at 85 °C for 2 h, and was then cooled down to room temperature. Distilled water (1 mL) was added and later vortexed for few seconds before adding 0.5 mL of hexane. In order to separate the reaction mixture, it was centrifuged at 6000 rpm (4025× g) for 5 min. The upper fraction of the hexane phase containing FAMEs was collected and transferred to GC vial tube. For analysis of fatty acid compositions, GC chromatograms were interpreted by comparing with standard equations. In this study, the commercial standard of fatty acid mixtures (F.A.M.E. mixed C8-C24 from SUPELCO©) was prepared in the same method as samples.
3.7. Total RNAs Extraction and Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
Total RNAs were extracted from each strain using TRIzol® Reagent (Invitrogen, Life Technologies Corporation, Carlsbad, CA, USA). The contaminated DNA was removed by treating with RNase-free DNaseI enzyme and DNaseI buffer before starting the reverse transcription step. The purified RNAs were calculated and further converted to cDNA using SuperScript™III First strand synthesis SuperMix Kit (Invitrogen, Carlsbad, CA, USA). After that, the cDNA of each stain was used as the template for PCR amplification of genes including accA, plsX, aas, lipA, glpD, rbcS, rbcL and 16S rRNA (as a reference) by corresponding pairs of RT-PCR primers (Supplementary Materials, Table S1). The PCR conditions were sequentially performed by initial denaturing at 95 °C for 3 min, followed by suitable cycles of each gene (Supplementary Materials, Table S1) at 95 °C for 30 s, primer melting temperature (Tm, Supplementary Materials, Table S1) for 30 s and 72 °C for 30 s, and then final extension at 72 °C for 5 min. Then, the PCR products were checked by 1.2% agarose gel electrophoresis. Quantification of band intensity was detected by Syngene® Gel Documentation (Syngene, Frederick, MD, USA).
3.8. Ultrastructure Visualization by Transmission Electron Microscopy (TEM)
The control WT (WTc) and engineered KAOL cells were visualized by transmission electron microscopy (TEM). Cells cultured in BG11 medium at day 5 of cultivation were harvested and sequentially fixed by adding 1 mL of the fixative solution containing 2.5% glutaraldehyde in 0.1 M phosphate buffer pH 7.0, and incubated at 4 °C for 2 h. After removing the fixative solution, 0.1 M phosphate buffer was added, and the reaction mixture was left at room temperature for 15–20 min. Next, the phosphate buffer was removed, and the cells were treated with 1% OsO4 in 0.1 M phosphate buffer for 1–2 h. The fixed pellets were washed by distilled water and subsequently coated with 2% agarose. After that, the coated cells were cut into blocks for dehydration and embedding. The embedded samples were again cut into sections with a thickness of about 80 nm by ultramicrotome and coated on the copper grid. These sections were stained with uranyl acetate and lead citrate solutions. The ultrastructure images were visualized by using transmission electron microscopy-JEM-1400Flash Electron Microscope operated at 80 kV.
3.9. Determination of PHB Contents by HPLC
Cell cultures (5–10 mL) of each condition were harvested by centrifugation at 6000 rpm (4025× g) for 10 min. To prepare the samples, 100 µL of 20 mg/mL of adipic acid (internal standard) and 800 µL of concentrated H2SO4 were added into cell pellets and further boiled at 100 °C for 1 h for digesting PHB to crotonic acid (modified from [29]). After that, 50 µL of the reaction mixture was diluted with 1.20 mL of ultrapure water. Then, 1 mL of solution was filtered through PP Syringe filter (0.45 µm, 13 mm), and collected in a glass vial for HPLC analysis (Shimadzu HPLC LGE System, Kyoto, Japan) using carbon-18 column with inert sustain of 3 µm (GL-Sciences, Tokyo, Japan). The running buffer was 30% (v/v) acetonitrile in 10 mM KH2PO4 at pH 2.3, with a flow rate of about 1.0 mL/min. The volume of crotonic acid was detected at 210 nm. Authentic commercial PHB (Sigma-Aldrich, Inc., St. Louis, MO, USA) as standard was prepared in the same way as the samples.
3.10. Fluorescence Microscopy of Nile-Red Stained Cells
Neutral lipids including PHB in Synechocystis cells were visualized by Nile-red staining dye (modified from [29]). Then, 100 µL of cell culture was harvested by centrifugation at 6000 rpm (4025× g) for 10 min. The cell pellets were washed by distilled water and stained by 30 µg/mL of Nile-red solution containing 0.9% (w/v) NaCl. The mixture was incubated in darkness overnight before observation under a fluorescent microscope (Olympus DP72, Tokyo, Japan), using a filter cube with 535 nm excitation wavelength.
3.11. Determination of Oxygen Evolution Rate
Five mL of cell culture was harvested by centrifugation at 6000 rpm (4025× g) for 10 min. The pellets were resuspended in 2 mL of fresh BG11 medium and incubated in darkness for 30 min. Thereafter, oxygen evolution of cells was measured under saturated light illumination by a Clark-type oxygen electrode (Hansatech instruments Ltd., King’s Lynn, UK) at 25 °C [6]. The oxygen evolution rate is presented as µmol/mg chlorophyll a/h.
3.12. Protein Extraction and Determination of Lipase Activity
To obtain crude protein, cell harvesting (20 mL culture) was carried out by centrifugation at 6000 rpm (4025× g), 4 °C for 10 min followed by cell extraction. Then, 200 µL of TB buffer (50 mM HEPES-NaOH (pH 7.0), 5 mM MgCl2, 25 mM CaCl2 and 10% (v/v) glycerol) was added to resuspend the cell pellets. Thereafter, glass beads were added into the cell suspension and the mixture was vigorously vortexed for 30 s and placed on ice for 15 s, repeated at least two times. After centrifugation at 6000 rpm (4025× g), at 4 °C for 5 min, the supernatant containing extracted proteins was transferred to a new tube. All supernatant fractions were pooled and subsequently centrifuged at 12,000 rpm (16,099× g), at 4 °C for 30 min to completely remove particles. The protein content was analyzed by the Bradford method [49] using commercial Sigma AldrichTM Bovine Serum Albumin, BSA as standard.
Lipase activity was determined by measuring the amount of p-nitrophenol (p-NP) released from p-NP ester substrate (modified from [50,51]). Crude extracted protein (20 μg) was added to 50 mM NaOAc (pH 8.0) containing 0.5 mM p-nitrophenylpalmitate (p-NPP), 10 mM isopropanol and 0.1% (v/v) Triton-X100. The reaction mixture was incubated at 37 °C for 60 min. The release of p-NP was measured spectrophotometrically at 405 nm. One unit of enzyme activity was defined as the amount of enzyme releasing 1 μmol of p-NP per minute.
4. Conclusions
Given the metabolic engineering approach that relies on FFA recycling, degradation of membrane lipids, and CBB cycle for improving FFA secretion of Synechocystis sp. PCC 6803, our findings from this study are expected to have broader implementations. We successfully constructed three engineered cyanobacterial strains including KA, KAOL and KAOGR strains, all with significantly increased levels of intracellular lipids and secreted FFAs, in particular the KAOL strain. The KAOL strain, a lipA-overexpressing KA, contained abundant white droplets and showed both distinct intracellular lipid production and FFA secretion, with a growth rate and an oxygen evolution rate higher than that of WT control cells. This novel, FFA-producing, cell factory continuously secretes fatty acids, and is a highly promising applied biotechnological finding, advancing the bioenergy research field.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/ijms222111468/s1.
Author Contributions
Conceptualization, A.I., P.L. and S.J.; Formal analysis, K.E.; Funding acquisition, S.J.; Investigation, K.E.; Methodology, S.J.; Project administration, S.J.; Resources, A.I., P.L. and S.J.; Supervision, S.J.; Validation, S.J.; Visualization, K.E. and S.J.; Writing—Original draft, K.E. and S.J.; Writing—Review and editing, S.J. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by grants from Chulalongkorn University Second Century Fund, C2F to S.J. and K.E., the Development and Promotion of Science and Technology Talents Project (DPST)’s scholarship to K.E., and Chulalongkorn University: CU_GR_62_25_23_08 fund to S.J.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
None of the authors have any financial conflict of interest that might be constructed to influence the results or interpretation of this manuscript.
Abbreviations
| AAS | acyl–acyl carrier protein synthetase |
| ACP | acyl carrier protein |
| Car | carotenoids |
| Chl a | chlorophyll a |
| CO2 | carbon dioxide |
| DCW | dry cell weight |
| DMF | N,N-dimethylformamide |
| E. coli | Escherichia coli |
| FFA | free fatty acid |
| h | hour |
| lipA | lipase A |
| m | meter |
| μg | microgram |
| mL | milliliter |
| mg | milligram |
| min | minute |
| nm | nanometer |
| OD | optical density |
| PCR | polymerase chain reaction |
| plsX | putative acyltransferase |
| p-NP | p-nitrophenol |
| p-NPP | p-nitrophenylpalmitate |
| rbc | RuBisCO |
| rpm | revolutions per minute |
| s | second |
| TEM | transmission electron microscopy |
| WT | wild type |
| WTc | wild-type control |
References
- Huntley, M.E.; Redalje, D.G. CO2 mitigation and renewable oil from photosynthetic microbes: A new appraisal. Mitig. Adapt. Strateg. Glob. Chang. 2007, 12, 573–608. [Google Scholar] [CrossRef]
- Li, Y.; Horsman, M.; Wu, N.; Lan, C.Q.; Dubois-Calero, N. Biofuels from microalgae. Biotechnol. Prog. 2008, 24, 815–820. [Google Scholar] [CrossRef]
- Quintana, N.; Van der Kooy, F.; Van de Rhee, M.D.; Voshol, G.P.; Verpoorte, R. Renewable energy from cyanobacteria: Energy production optimization by metabolic pathway engineering. Appl. Microbiol. Biotechnol. 2011, 91, 471–490. [Google Scholar] [CrossRef] [Green Version]
- Eungrasamee, K.; Incharoensakdi, A.; Lindblad, P.; Jantaro, S. Synechocystis sp. PCC 6803 overexpressing genes involved in CBB cycle and free fatty acid cycling enhances the significant levels of intracellular lipids and secreted free fatty acids. Sci. Rep. 2020, 10, 4515. [Google Scholar] [CrossRef] [PubMed]
- Janßen, H.J.; Steinbüchel, A. Fatty acid synthesis in Escherichia coli and its applications towards the production of fatty acid based biofuels. Biotechnol. Biofuels 2014, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Eungrasamee, K.; Miao, R.; Incharoensakdi, A.; Lindblad, P.; Jantaro, S. Improved lipid production via fatty acid biosynthesis and free fatty acid recycling in engineered Synechocystis sp. PCC 6803. Biotechnol. Biofuels 2019, 12, 8. [Google Scholar] [CrossRef]
- Kato, A.; Use, K.; Takatani, N.; Ikeda, K.; Matsuura, M.; Kojima, K.; Aichi, M.; Maeda, S.; Omata, T. Modulation of the balance of fatty acid production and secretion is crucial for enhancement of growth and productivity of the engineered mutant of the cyanobacterium Synechococcus elongatus. Biotechnol. Biofuels 2016, 9, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Sheng, J.; Curtiss, R. III. Fatty acid production in genetically modified cyanobacteria. Proc. Natl. Acad. Sci. USA 2011, 108, 6899–6904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruffing, A.M.; Jones, H.D. Physiological effects of free fatty acid production in genetically engineered Synechococcus elongatus PCC 7942. Biotechnol. Bioeng. 2012, 109, 2190–2199. [Google Scholar] [CrossRef] [Green Version]
- Ruffing, A.M. Improved free fatty acid production in cyanobacteria with Synechococcus sp. PCC 7002 as host. Front. Bioeng. Biotechnol. 2014, 2, 17. [Google Scholar] [CrossRef] [Green Version]
- Molina Grima, E.; Belarbi, E.H.; Acién Fernández, F.G.; Robles Medina, A.; Chisti, Y. Recovery of microalgal biomass and metabolites: Process options and economics. Biotechnol. Adv. 2003, 20, 491–515. [Google Scholar] [CrossRef]
- Yang, F.; Xiang, W.; Li, T.; Long, L. Transcriptome analysis for phosphorus starvation-induced lipid accumulation in Scenedesmus sp. Sci. Rep. 2018, 8, 16420. [Google Scholar] [CrossRef] [PubMed]
- Towijit, U.; Songruk, N.; Lindblad, P.; Incharoensakdi, A.; Jantaro, S. Co-overexpression of native phospholipid-biosynthetic genes plsX and plsC enhances lipid production in Synechocystis sp. PCC 6803. Sci. Rep. 2018, 8, 13510. [Google Scholar] [CrossRef] [PubMed]
- Minyuk, G.; Sidorov, R.; Solovchenki, A. Effect of nitrogen source on the growth, lipid, and valuable carotenoid production in the green microalga Chromochloris zofingiensis. J. Appl. Phycol. 2020, 32, 923–935. [Google Scholar] [CrossRef]
- Kaczmarzyk, D.; Fulda, M. Fatty acid activation in cyanobacteria mediated by acyl-acyl carrier protein synthetase enables fatty acid recycling. Plant Physiol. 2010, 152, 1598–1610. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Tan, X.; Lü, X. Characterization of a key gene in membrane lipid cycle in Synechocystis sp. PCC6803. Chin. J. Biotechnol. 2012, 28, 1473–1481. [Google Scholar]
- Takatani, N.; Use, K.; Kato, A.; Ikeda, K.; Kojima, K.; Aichi, M.; Maeda, S.-I.; Omata, T. Essential role of acyl-ACP synthetase in acclimation of the cyanobacterium Synechococcus elongatus strain PCC 7942 to high-light conditions. Plant Cell Physiol. 2015, 56, 1608–1615. [Google Scholar] [CrossRef] [Green Version]
- Kato, A.; Takatani, N.; Ikeda, K.; Maeda, S.-I.; Omata, T. Removal of the product from the culture medium strongly enhances free fatty acid production by genetically engineered Synechococcus elongatus. Biotechnol. Biofuels 2017, 10, 141. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, A.; Sato, Y.; Saito, Y.; Kaneko, Y.; Takimura, Y.; Hagihara, H.; Hihara, Y. Free fatty acid production in the cyanobacterium Synechocystis sp. PCC 6803 is enhanced by deletion of the cyAbrB2 transcriptional regulator. J. Biotechnol. 2016, 220, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Afrin, S.; Khan, M.; Zhang, W.; Wang, Y.; Zhang, W.; He, L.; Ma, G. Membrane-located expression of thioesterase from Acinetobacter baylyi enhances free fatty acid production with decreased toxicity in Synechocystis sp. PCC 6803. Front. Microbiol. 2018, 9, 2842. [Google Scholar] [CrossRef]
- Lu, X.; Vora, H.; Khosla, C. Overproduction of free fatty acids in E. coli: Implications for biodiesel production. Metab. Eng. 2008, 10, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Shang, H.; Zheng, Y.; Zhang, Z. Free fatty acid secretion by an engineered strain of Escherichia coli. Biotechnol. Lett. 2013, 35, 2099–2103. [Google Scholar] [CrossRef]
- Leber, C.; Polson, B.; Fernandez-Moya, R.; Da Silva, N.A. Overproduction and secretion of free fatty acids through disrupted neutral lipid recycle in Saccharomyces cerevisiae. Metab. Eng. 2015, 28, 54–62. [Google Scholar] [CrossRef]
- Ferreira, R.; Teixeira, P.G.; Siewers, V.; Nielsen, J. Redirection of lipid flux toward phospholipids in yeast increases fatty acid turnover and secretion. Proc. Natl. Acad. Sci. USA 2018, 115, 1262–1267. [Google Scholar] [CrossRef] [Green Version]
- Michelet, L.; Zaffagnini, M.; Morisse, S.; Sparla, F.; Pérez-Pérez, M.E.; Francia, F.; Danon, A.; Marchand, C.H.; Fermani, S.; Trost, P.; et al. Redox regulation of the Calvin–Benson cycle: Something old, something new. Front. Plant Sci. 2013, 4, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Xiong, X.; Sa, N.; Roje, S.; Chen, S. Metabolic engineering of enhanced glycerol-3-phosphate synthesis to increase lipid production in Synechocystis sp. PCC 6803. Appl. Microbiol. Biotechnol. 2016, 100, 6091–6101. [Google Scholar] [CrossRef]
- Farmer, W.R.; Liao, J.C. Precursor balancing for metabolic engineering of lycopene production in Escherichia coli. Biotechnol. Prog. 2001, 17, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Panda, B.; Mallick, N. Enhanced poly-beta-hydroxybutyrate accumulation in a unicellular cyanobacterium Synechocystis sp. PCC 6803. Lett. Appl. Microbiol. 2007, 44, 194–198. [Google Scholar] [CrossRef]
- Khetkorn, W.; Incharoensakdi, A.; Lindblad, P.; Jantaro, S. Enhancement of poly-3-hydroxybutyrate production in Synechocystis sp. PCC 6803 by overexpression of its native biosynthetic genes. Bioresour. Technol. 2016, 214, 761–768. [Google Scholar] [CrossRef]
- Mansour, M.P.; Volkman, J.K.; Blackburn, S.I. The effect of growth phase on the lipid class, fatty acid and sterol composition in the marine dinoflagellate, Gymnodinium sp. in batch culture. Phytochemistry 2003, 63, 145–153. [Google Scholar] [CrossRef]
- Los, D.A.; Mironov, K.S.; Allakhberdiev, S.I. Regulatory role of membrane fluidity in gene expression and physiological functions. Photosynth. Res. 2013, 116, 489–509. [Google Scholar] [CrossRef]
- Campbell, J.W.; Morgan-Kiss, R.M.; Cronan, J.E., Jr. A new Escherichia coli metabolic competency: Growth on fatty acids by a novel anaerobic beta-oxidation pathway. Mol. Microbiol. 2003, 47, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Matsuoka, H.; Hirooka, K. Regulation of fatty acid metabolism in bacteria. Mol. Microbiol. 2007, 66, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Parsons, J.B.; Rock, C.O. Bacterial lipids: Metabolism and membrane homeostasis. Prog. Lipid Res. 2013, 52, 249–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beld, J.; Abbriano, R.; Finzel, K.; Hildebrand, M.; Burkart, M.D. Probing fatty acid metabolism in bacteria, cyanobacteria, green microalgae and diatoms with natural and unnatural fatty acids. Mol. BioSyst. 2016, 12, 1299. [Google Scholar] [CrossRef]
- Trautner, C.; Vermaas, W.F. The sll1951 gene encodes the surface layer protein of Synechocystis sp. strain PCC 6803. J. Bacteriol. 2013, 195, 5370–5380. [Google Scholar] [CrossRef]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 162942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lennen, R.M.; Pfleger, B.F. Engineering Escherichia coli to synthesize free fatty acids. Trends Biotechnol. 2012, 30, 659–667. [Google Scholar] [CrossRef] [Green Version]
- Dunlop, M.J.; Keasling, J.D.; Mukhopadhyay, A.A. Model for improving microbial biofuel production using a synthetic feedback loop. Syst. Synth. Biol. 2010, 4, 95–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruffing, A.M. RNA-Seq analysis and targeted mutagenesis for improved free fatty acid production in an engineered cyanobacterium. Biotechnol. Biofuels 2013, 6, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, F.; Lindblad, P. Synechocystis PCC 6803 overexpressing RuBisCO grow faster with increased photosynthesis. Metab. Eng. Commun. 2017, 4, 29–36. [Google Scholar] [CrossRef]
- Peramuna, A.; Summers, M.L. Composition and occurrence of lipid droplets in the cyanobacterium Nostoc punctiforme. Arch. Microbiol. 2014, 196, 881–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Englund, E.; Andersen-Ranberg, J.; Miao, R.; Hamberger, B.; Lindberg, P. Metabolic engineering of Synechocystis sp. PCC 6803 for production of the plant diterpenoid manoyl oxide. ACS Synth. Biol. 2015, 4, 1270–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jantaro, S.; Mäenpää, P.; Mulo, P.; Incharoensakdi, A. Content and biosynthesis of polyamines in salt and osmotically stressed cells of Synechocystis sp. PCC 6803. FEMS Microbiol. Lett. 2003, 228, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Moran, R. Formulae for determination of chlorophyllous pigments extracted with N,N-dimethylformamide. Plant Physiol. 1982, 69, 1376–1381. [Google Scholar] [CrossRef] [Green Version]
- Chamovitz, D.; Sandmann, G.; Hirschberg, J. Molecular and biochemical characterization of herbicide-resistant mutants of cyanobacteria reveals that phytoene desaturation is a rate-limiting step in carotenoid biosynthesis. J. Biol. Chem. 1993, 268, 17348–17353. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fales, M.F. Evaluation of a spectrophotometric method for determination of total fecal lipid. Clin. Chem. 1971, 17, 1103–1108. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Winkler, U.K.; Stuckmann, M. Glycogen, hyaluronate, and some other polysaccharides greatly enhance the formation of exolipase by Serratia marcescens. J. Bacteriol. 1979, 138, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Dang, G.; Li, H.; Li, T.; Yue, Z.; Li, N.; Liu, Y.; Liu, S.; Chen, L. Identification and characterization of lipase activity and immunogenicity of LipL from Mycobacterium tuberculosis. PLoS ONE 2015, 10, e0138151. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).