VDAC1 Negatively Regulates Floral Transition in Arabidopsis thaliana


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
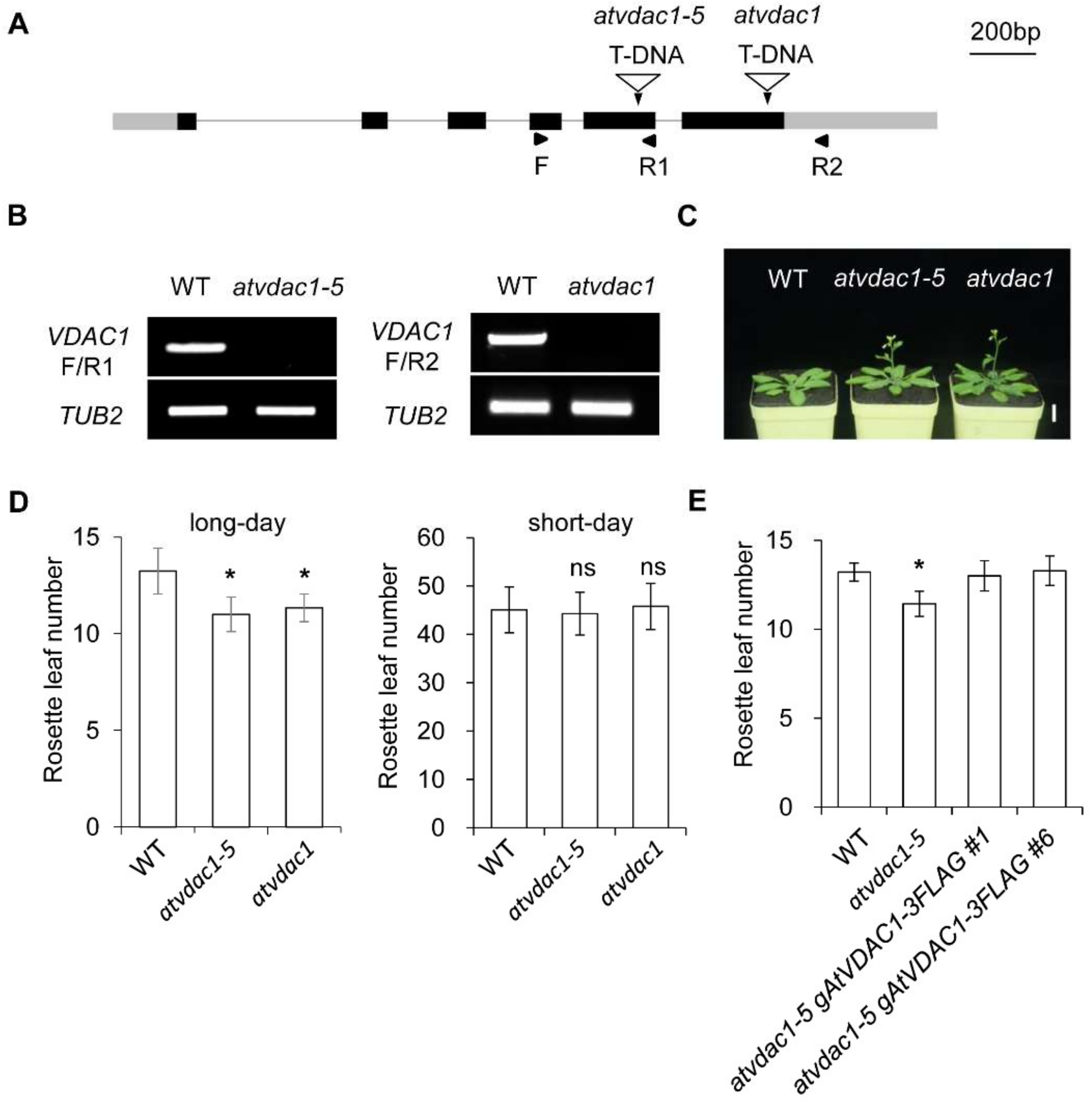
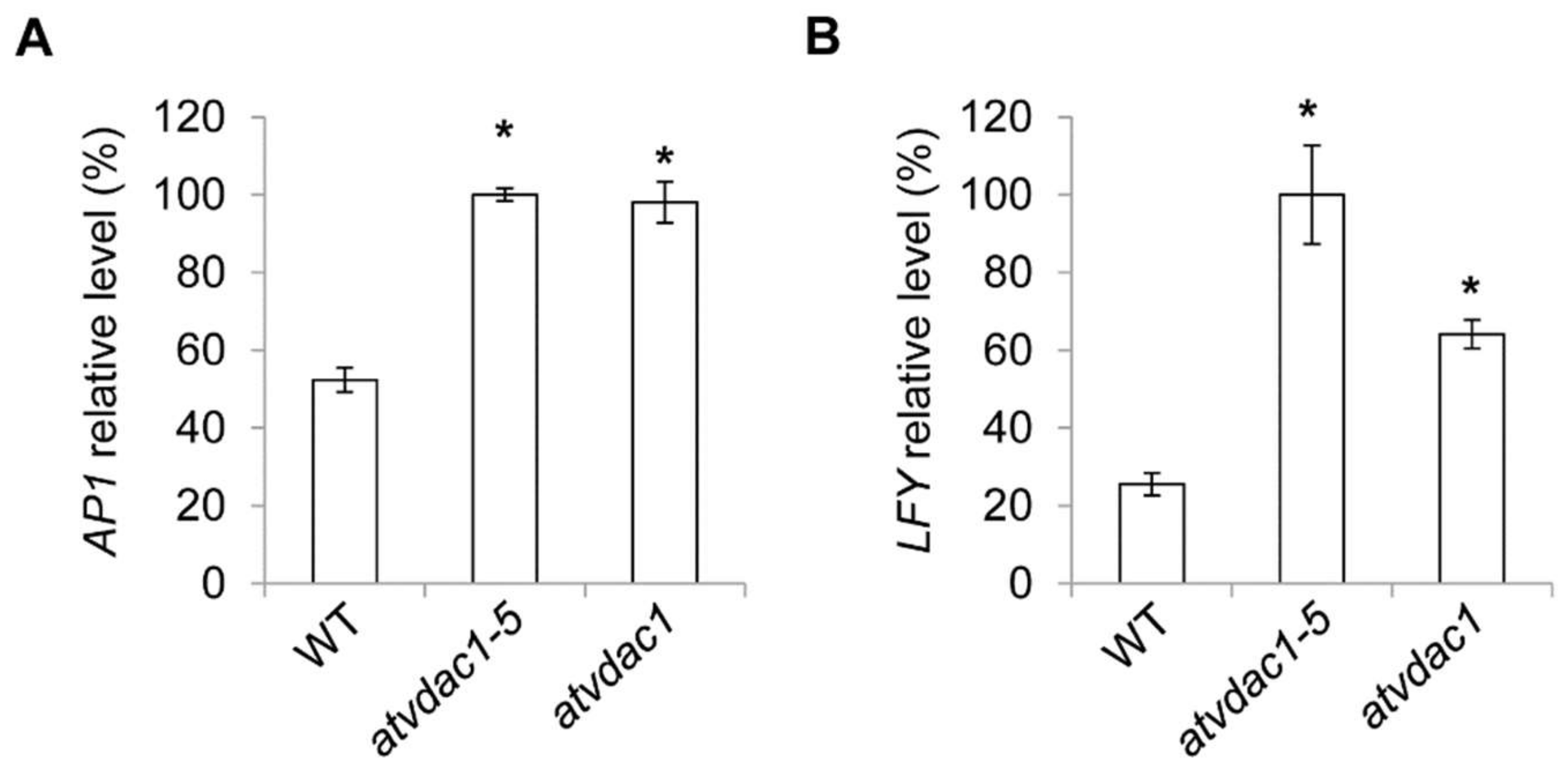
2.1. AtVDAC1 Participates in Flowering-Time Regulation in Arabidopsis
2.2. AtVDAC1 Is Expressed Ubiquitously in Arabidopsis
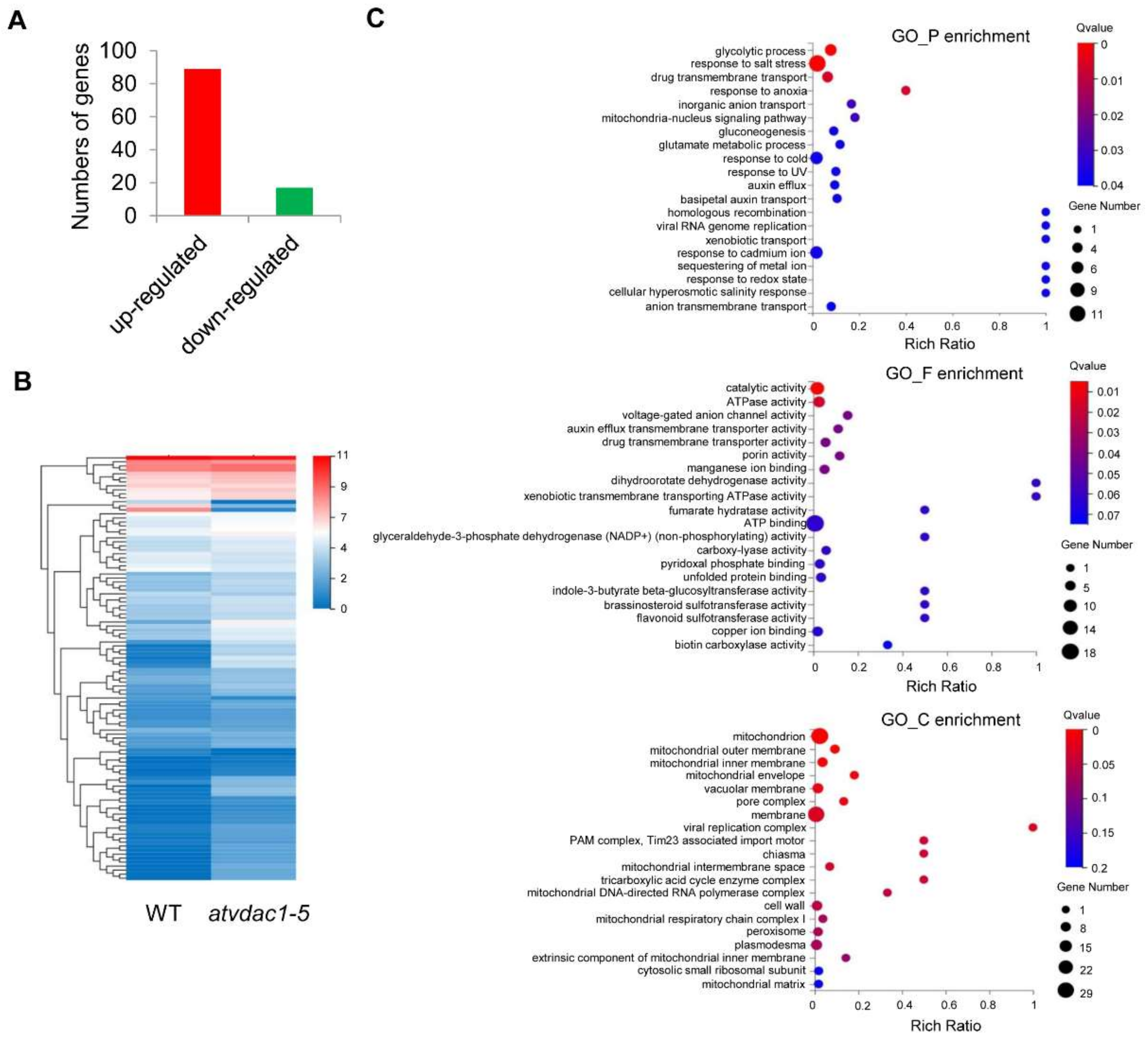
2.3. Transcriptome Profiles and Differentially Expressed Gene Identification between WT and atvdac1-5 Mutant Seedlings
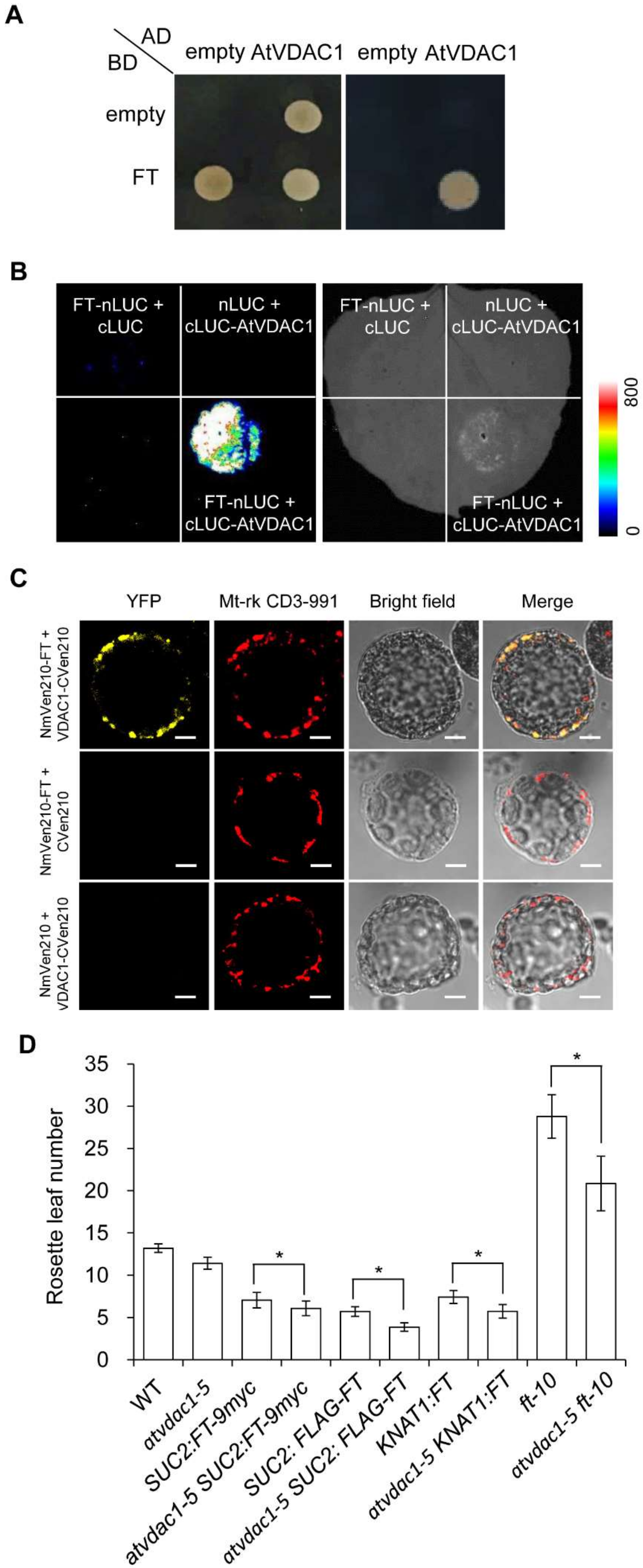
2.4. AtVDAC1 Interacts with FT
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Plasmid Construction
4.3. Yeast Two-Hybrid Assay
4.4. LCI Assays
4.5. GUS Staining
4.6. RNA-seq Analyses
4.7. Expression Analysis
4.8. Flowering Time Measurement
4.9. Arabidopsis Protoplast Isolation and Sub-Cellular Localization Assay
4.10. BiFC Analysis
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fuchs, P.; Rugen, N.; Carrie, C.; Elsasser, M.; Finkemeier, I.; Giese, J.; Hildebrandt, T.M.; Kuhn, K.; Maurino, V.G.; Ruberti, C.; et al. Single organelle function and organization as estimated from Arabidopsis mitochondrial proteomics. Plant J. 2020, 101, 420–441. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Chen, Z.; Yang, X.; Liu, G. Arabidopsis voltage-dependent anion channel 1 (AtVDAC1) is required for female development and maintenance of mitochondrial functions related to energy-transaction. PLoS ONE 2014, 9, e106941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blumenthal, A.; Kahn, K.; Beja, O.; Galun, E.; Colombini, M.; Breiman, A. Purification and Characterization of the Voltage-Dependent Anion-Selective Channel Protein from Wheat Mitochondrial Membranes. Plant Physiol. 1993, 101, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Elkeles, A.; Devos, K.M.; Graur, D.; Zizi, M.; Breiman, A. Multiple cDNAs of wheat voltage-dependent anion channels (VDAC): Isolation, differential expression, mapping and evolution. Plant Mol. Biol. 1995, 29, 109–124. [Google Scholar] [CrossRef]
- Elkeles, A.; Breiman, A.; Zizi, M. Functional differences among wheat voltage-dependent anion channel (VDAC) isoforms expressed in yeast. Indication for the presence of a novel VDAC-modulating protein? J. Biol. Chem. 1997, 272, 6252–6260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roosens, N.; Al Bitar, F.; Jacobs, M.; Homble, F. Characterization of a cDNA encoding a rice mitochondrial voltage-dependent anion channel and its gene expression studied upon plant development and osmotic stress. Biochim. Biophys. Acta 2000, 1463, 470–476. [Google Scholar] [CrossRef]
- Al Bitar, F.; Roosens, N.; Boxtel, J.V.; Dewaele, E.; Jacobs, M.; Homble, F. Expression of the rice vdac isoform2: Histochemical localization and expression level. Biochim. Biophys. Acta 2002, 1579, 133–141. [Google Scholar] [CrossRef]
- Al Bitar, F.; Roosens, N.; Smeyers, M.; Vauterin, M.; Van Boxtel, J.; Jacobs, M.; Homble, F. Sequence analysis, transcriptional and posttranscriptional regulation of the rice vdac family. Biochim. Biophys. Acta 2003, 1625, 43–51. [Google Scholar] [CrossRef]
- Wandrey, M.; Trevaskis, B.; Brewin, N.; Udvardi, M.K. Molecular and cell biology of a family of voltage-dependent anion channel porins in Lotus japonicus. Plant Physiol. 2004, 134, 182–193. [Google Scholar] [CrossRef] [Green Version]
- Tateda, C.; Yamashita, K.; Takahashi, F.; Kusano, T.; Takahashi, Y. Plant voltage-dependent anion channels are involved in host defense against Pseudomonas cichorii and in Bax-induced cell death. Plant Cell Rep. 2009, 28, 41–51. [Google Scholar] [CrossRef]
- Lee, S.M.; Hoang, M.H.; Han, H.J.; Kim, H.S.; Lee, K.; Kim, K.E.; Kim, D.H.; Lee, S.Y.; Chung, W.S. Pathogen inducible voltage-dependent anion channel (AtVDAC) isoforms are localized to mitochondria membrane in Arabidopsis. Mol. Cells 2009, 27, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Robert, N.; d’Erfurth, I.; Marmagne, A.; Erhardt, M.; Allot, M.; Boivin, K.; Gissot, L.; Monachello, D.; Michaud, M.; Duchêne, A.-M.; et al. Voltage-dependent-anion-channels (VDACs) in Arabidopsis have a dual localization in the cell but show a distinct role in mitochondria. Plant Mol. Biol. 2012, 78, 431–446. [Google Scholar] [CrossRef] [PubMed]
- Tateda, C.; Watanabe, K.; Kusano, T.; Takahashi, Y. Molecular and genetic characterization of the gene family encoding the voltage-dependent anion channel in Arabidopsis. J. Exp. Bot. 2011, 62, 4773–4785. [Google Scholar] [CrossRef] [Green Version]
- Tateda, C.; Kusano, T.; Takahashi, Y. The Arabidopsis voltage-dependent anion channel 2 is required for plant growth. Plant Signal Behav. 2012, 7, 31–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, Y.; Tateda, C. The functions of voltage-dependent anion channels in plants. Apoptosis Int. J. Program. Cell Death 2013, 18, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Y.; Xu, Z.S.; He, G.Y.; Yang, G.X.; Chen, M.; Li, L.C.; Ma, Y. The voltage-dependent anion channel 1 (AtVDAC1) negatively regulates plant cold responses during germination and seedling development in Arabidopsis and interacts with calcium sensor CBL1. Int. J. Mol. Sci. 2013, 14, 701–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, T. Mitochondrial Porin Isoform AtVDAC1 Regulates the Competence of Arabidopsis thaliana to Agrobacterium-Mediated Genetic Transformation. Mol. Cells 2016, 39, 705–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Luo, Q.H.; Wen, G.Q.; Wang, J.M.; Li, X.F.; Yang, Y. VDAC2 involvement in the stress response pathway in Arabidopsis thaliana. Genet. Mol. Res. 2015, 14, 15511–15519. [Google Scholar] [CrossRef]
- Singh, N.; Swain, S.; Singh, A.; Nandi, A.K. AtOZF1 Positively Regulates Defense Against Bacterial Pathogens and NPR1-Independent Salicylic Acid Signaling. Mol. Plant Microbe Interact. 2018, 31, 323–333. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; He, H.; Tong, S.; Zhang, W.; Wang, J.; Li, X.; Yang, Y. Voltage-dependent anion channel 2 of Arabidopsis thaliana (AtVDAC2) is involved in ABA-mediated early seedling development. Int. J. Mol. Sci. 2009, 10, 2476–2486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.Y.; Chen, Z.W.; Xu, T.; Qu, Z.; Pan, X.D.; Qin, X.H.; Ren, D.T.; Liu, G.Q. Arabidopsis kinesin KP1 specifically interacts with VDAC3, a mitochondrial protein, and regulates respiration during seed germination at low temperature. Plant Cell 2011, 23, 1093–1106. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Takano, T.; Liu, S.; Zhang, X. Arabidopsis mitochondrial voltage-dependent anion channel 3 (AtVDAC3) protein interacts with thioredoxin m2. FEBS Lett. 2015, 589, 1207–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Liu, S.; Takano, T.; Zhang, X. The interaction between AtMT2b and AtVDAC3 affects the mitochondrial membrane potential and reactive oxygen species generation under NaCl stress in Arabidopsis. Planta 2019, 249, 417–429. [Google Scholar] [CrossRef]
- Michaud, M.; Ubrig, E.; Filleur, S.; Erhardt, M.; Ephritikhine, G.; Marechal-Drouard, L.; Duchene, A.M. Differential targeting of VDAC3 mRNA isoforms influences mitochondria morphology. Proc. Natl. Acad. Sci. USA 2014, 111, 8991–8996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouain, N.; Korte, A.; Satbhai, S.B.; Nam, H.I.; Rhee, S.Y.; Busch, W.; Rouached, H. Systems genomics approaches provide new insights into Arabidopsis thaliana root growth regulation under combinatorial mineral nutrient limitation. PLoS Genet. 2019, 15, e1008392. [Google Scholar] [CrossRef] [Green Version]
- Taoka, K.I.; Ohki, I.; Tsuji, H.; Kojima, C.; Shimamoto, K. Structure and function of florigen and the receptor complex. Trends Plant Sci. 2013, 18, 287–294. [Google Scholar] [CrossRef]
- Taoka, K.; Ohki, I.; Tsuji, H.; Furuita, K.; Hayashi, K.; Yanase, T.; Yamaguchi, M.; Nakashima, C.; Purwestri, Y.A.; Tamaki, S.; et al. 14-3-3 proteins act as intracellular receptors for rice Hd3a florigen. Nature 2011, 476, 332–335. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Cheng, L.; Zhang, H.; He, M.; Shen, J.; Zhang, Y.; Wu, P. OsGatB, the Subunit of tRNA-Dependent Amidotransferase, Is Required for Primary Root Development in Rice. Front. Plant Sci. 2016, 7, 599. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xue, Z.; Lin, J.; Wang, Y.; Ying, H.; Lv, Q.; Hua, C.; Wang, M.; Chen, S.; Zhou, B. Proline improves cardiac remodeling following myocardial infarction and attenuates cardiomyocyte apoptosis via redox regulation. Biochem. Pharmacol. 2020, 178, 114065. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.H.; Shen, S.C.; Lee, L.Y.; Lee, S.H.; Chan, M.T.; Lin, C.S. Tape-Arabidopsis Sandwich—A simpler Arabidopsis protoplast isolation method. Plant Methods 2009, 5, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, B.K.; Cai, X.; Nebenfuhr, A. A multicolored set of in vivo organelle markers for co-localization studies in Arabidopsis and other plants. Plant J. 2007, 51, 1126–1136. [Google Scholar] [CrossRef] [PubMed]
- Gookin, T.E.; Assmann, S.M. Significant reduction of BiFC non-specific assembly facilitates in planta assessment of heterotrimeric G-protein interactors. Plant J. 2014, 80, 553–567. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Zhang, Y.; Ren, H.; Yu, R.; Yuan, C.; Hu, Y.; Xu, R.; Wang, X.; Qin, C. VDAC1 Negatively Regulates Floral Transition in Arabidopsis thaliana. Int. J. Mol. Sci. 2021, 22, 11603. https://doi.org/10.3390/ijms222111603
Xu J, Zhang Y, Ren H, Yu R, Yuan C, Hu Y, Xu R, Wang X, Qin C. VDAC1 Negatively Regulates Floral Transition in Arabidopsis thaliana. International Journal of Molecular Sciences. 2021; 22(21):11603. https://doi.org/10.3390/ijms222111603
Chicago/Turabian StyleXu, Jingya, Yuzhen Zhang, Hongjia Ren, Runyi Yu, Chen Yuan, Yikai Hu, Rumeng Xu, Xuming Wang, and Cheng Qin. 2021. "VDAC1 Negatively Regulates Floral Transition in Arabidopsis thaliana" International Journal of Molecular Sciences 22, no. 21: 11603. https://doi.org/10.3390/ijms222111603