
Differential Clearance of A? Species from the Brain by Brain Lymphatic Endothelial Cells in Zebrafish

 and
and 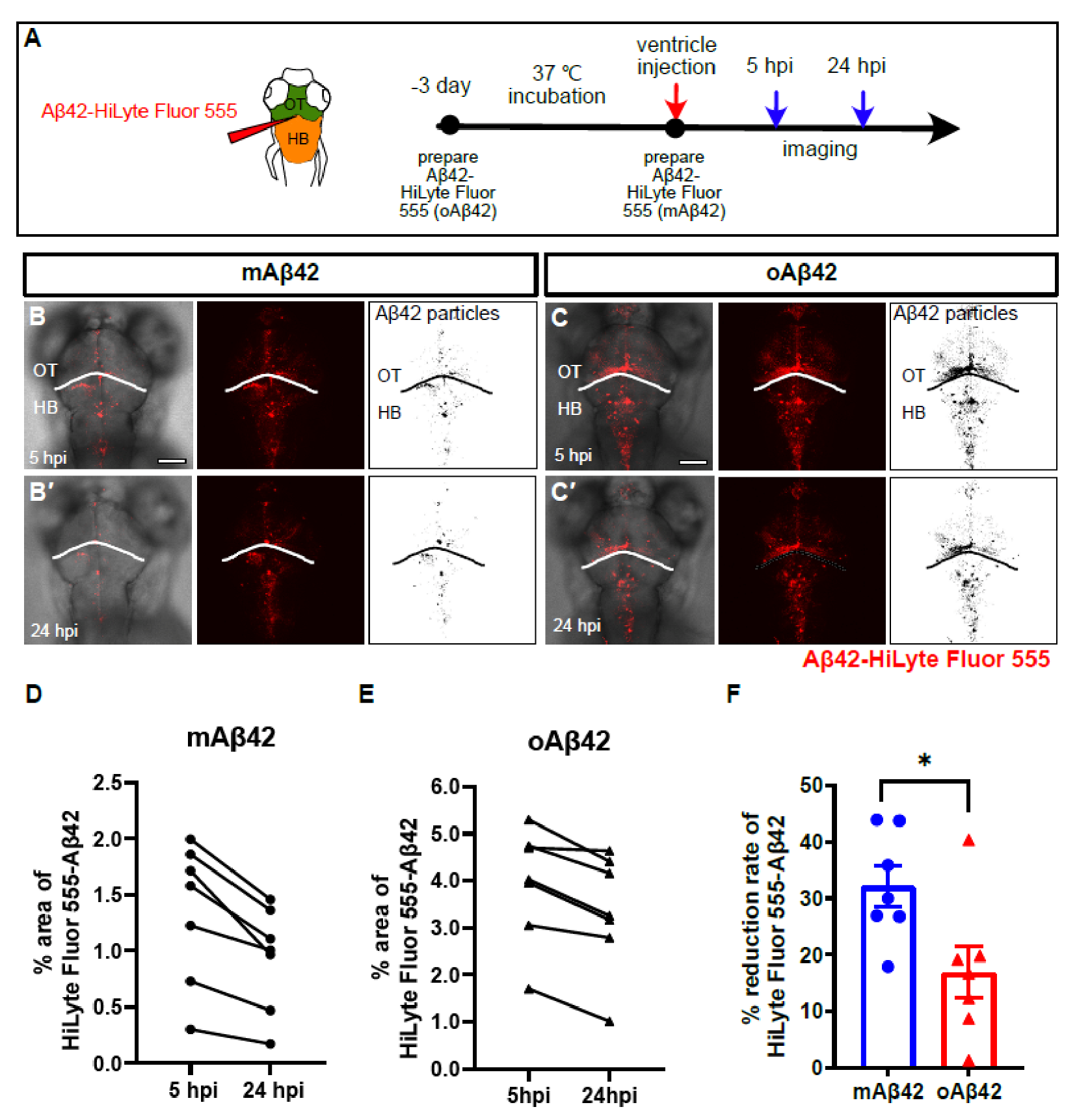{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Monomeric and Oligomeric Aβ42 Are Differentially Cleared from the Brain
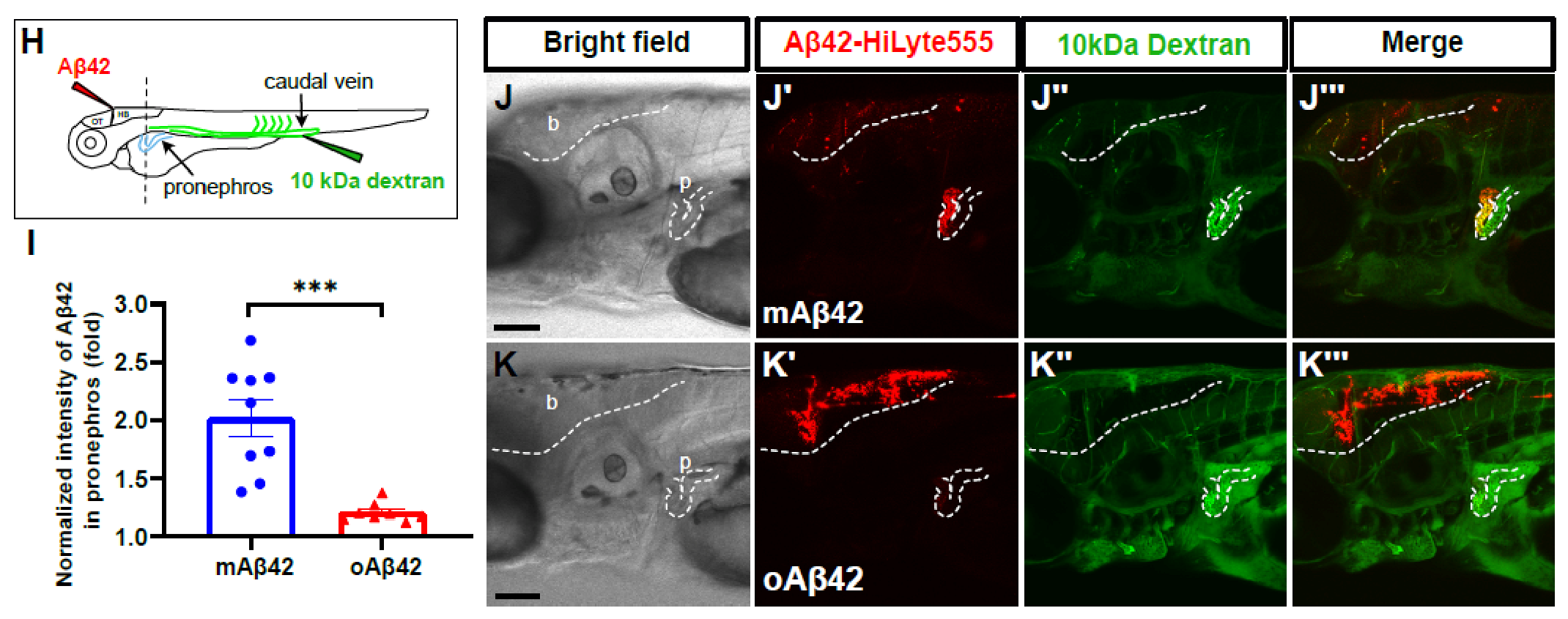
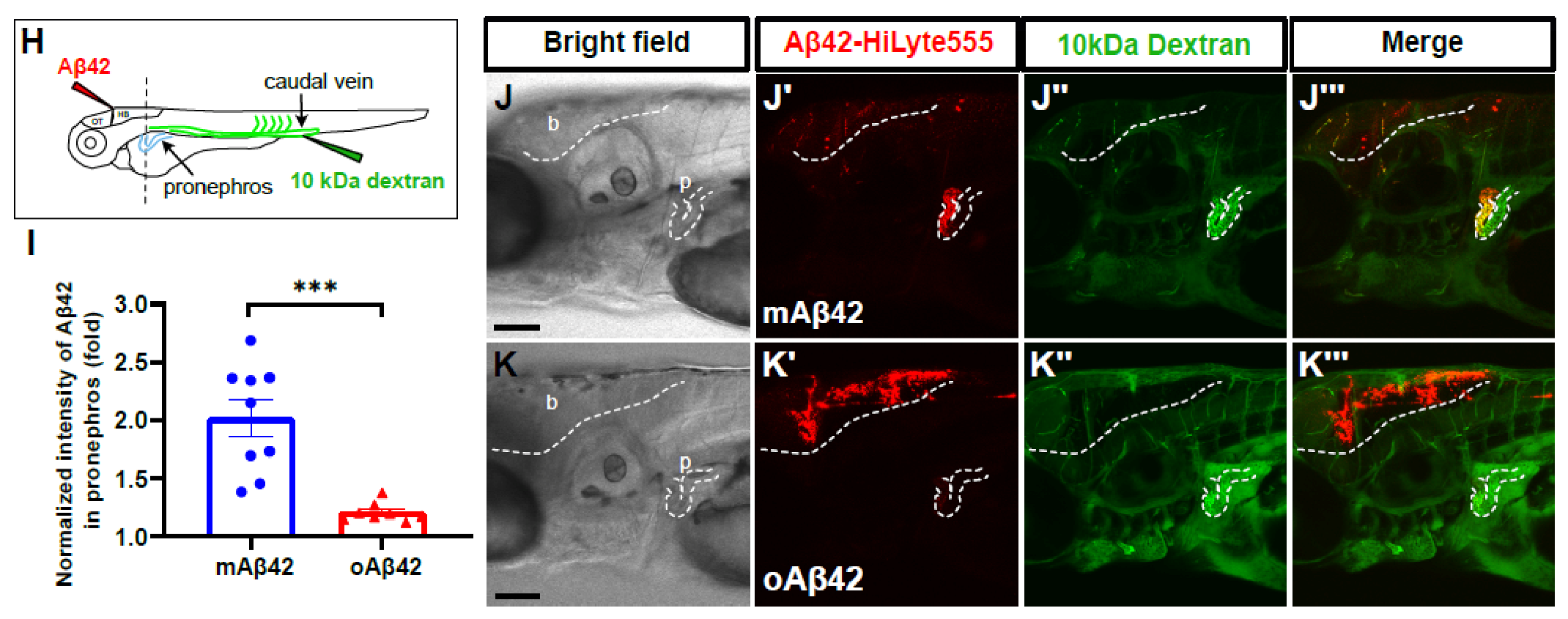
2.2. Cleared mAβ42 Accumulates in the Pronephros via Blood Flow
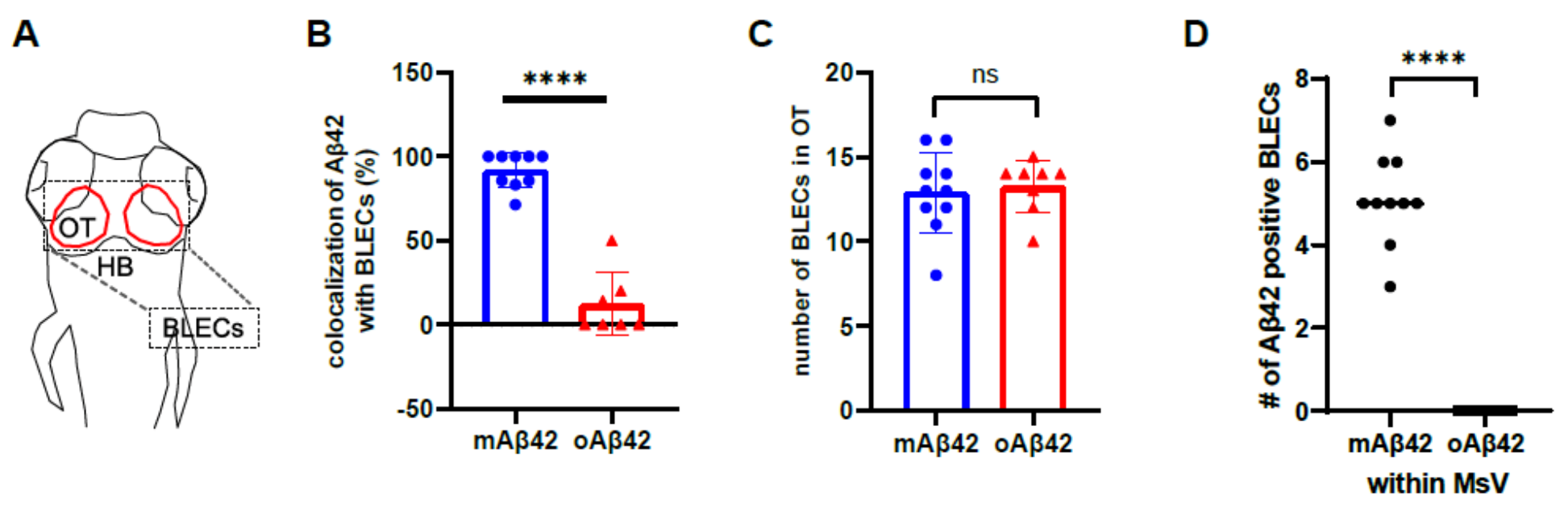
2.3. Monomeric Aβ42 Is Taken up by Brain Lymphatic Endothelial Cells
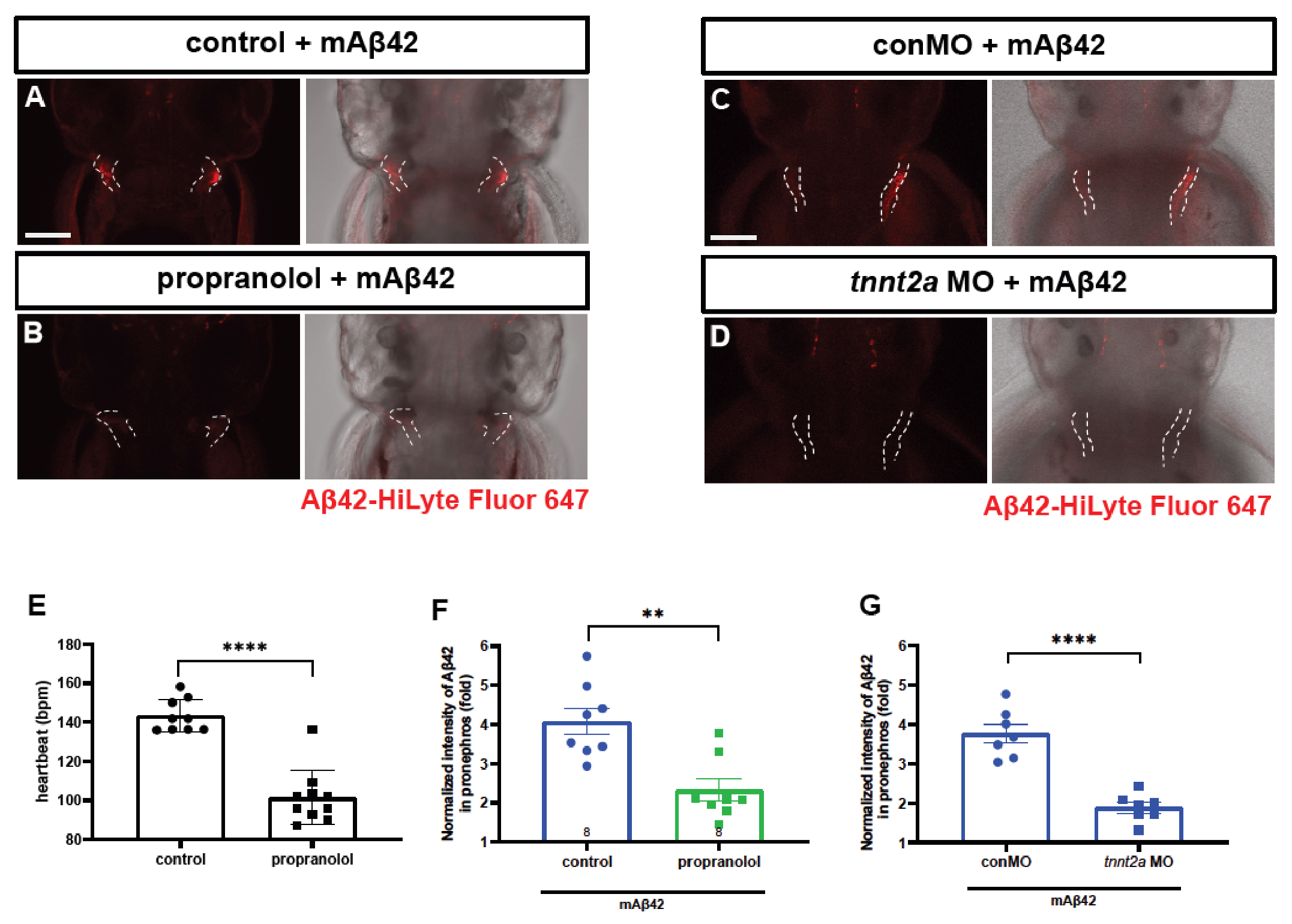
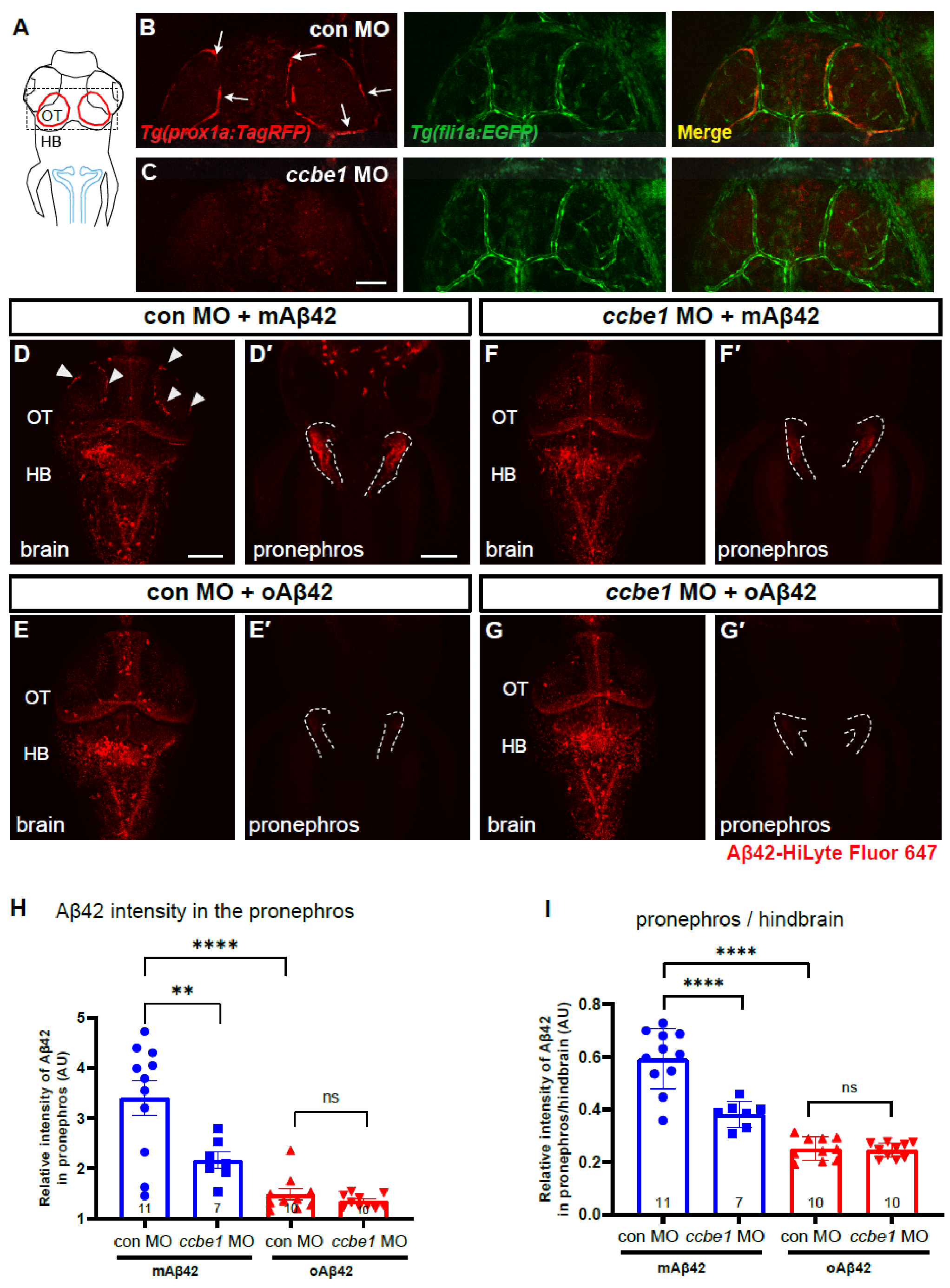
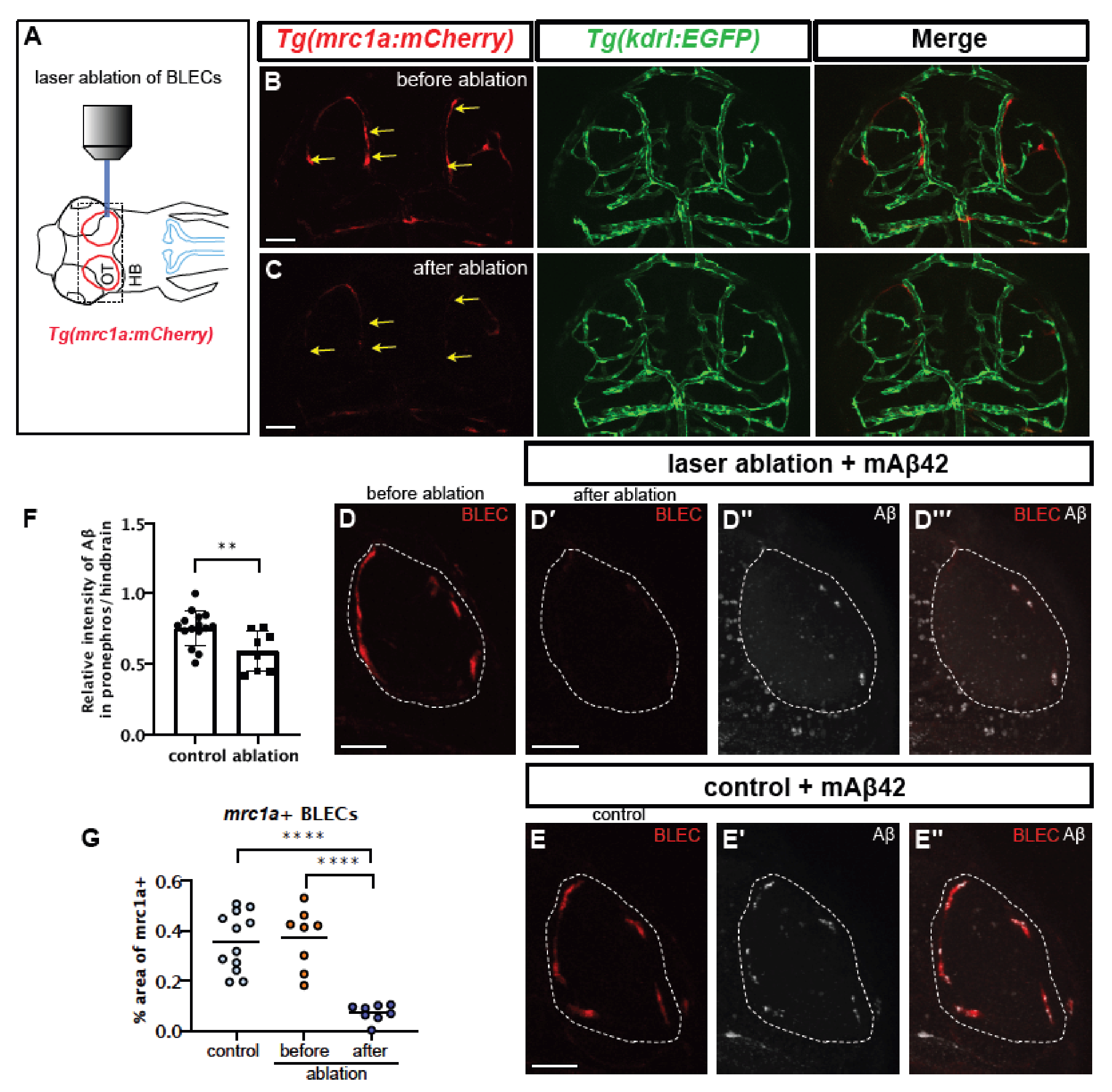
2.4. BLEC Depletion Reduces the Peripheral Transport of mAβ42 to the Pronephros
2.5. Mannan Administration Reduces mAβ42 Internalization by BLECs and Peripheral Transport
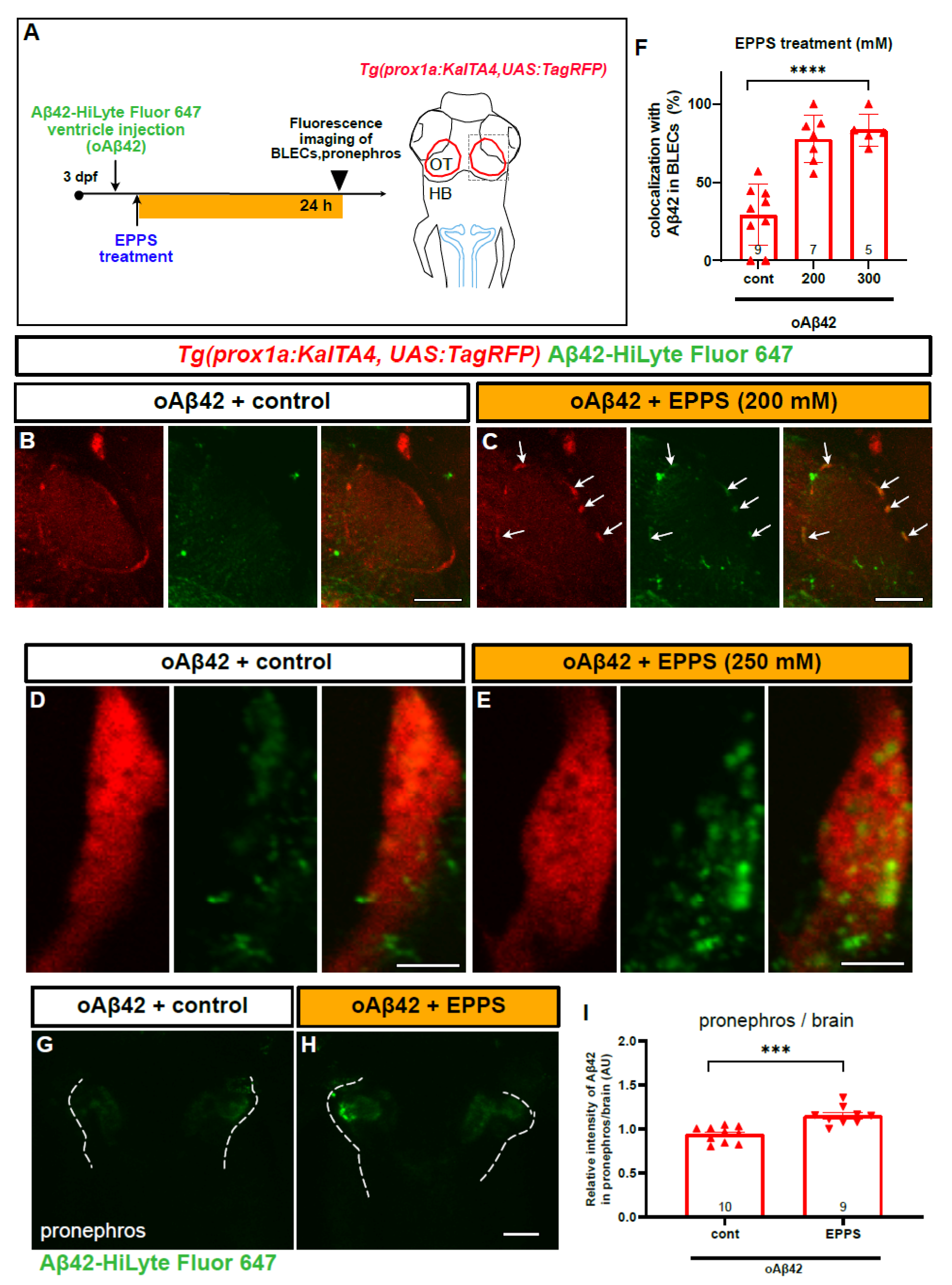
2.6. An Aβ42 Aggregation Inhibitor Promotes oAβ42 Localization into BLECs and the Peripheral Transport
3. Discussion
3.1. Cerebroventricular Injection of Aβ for Modeling Alzheimer’s Disease in Zebrafish
3.2. BLEC Is a Novel Component of the Brain Clearance System to Remove mAβ42
3.3. BLECs Selectively Clear Monomeric Aβ from Brain with a Blood Circulatory Route
4. Materials and Methods
4.1. Animals
4.2. Aβ preparation and Ventricle Microinjection
4.3. Confocal Microscopic Analyses for Aβ Clearance and Quantification
4.4. pHrodoGreen Injection
4.5. Genetic and Pharmacological Inhibition of Heartbeat
4.6. Intravenous Injection of Endocytic Tracer
4.7. Genetic and Pharmacological Inhibition of Brain Lymphatic Endothelial Cells
4.8. Laser Ablation of Brain Lymphatic Endothelial Cells
4.9. EPPS Treatment
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Aβ | amyloid beta |
| AD | Alzheimer’s disease |
| AFM | atomic force microscopy |
| BBB | blood–brain barrier |
| BLEC | brain lymphatic endothelial cells |
| CSF | cerebrospinal fluid |
| FGPs | fluorescent granule perithelial cells |
| ISF | interstitial fluid |
| mAβ42 | monomeric Aβ42 |
| oAβ42 | oligomeric Aβ42 |
References
- Long, J.; Holtzman, D.M. Alzheimer Disease: An Update on Pathobiology and Treatment Strategies. Cell 2019, 179, 312–339. [Google Scholar] [CrossRef] [PubMed]
- Thal, D.R. Clearance of Amyloid Beta-Protein and its Role in the Spreading of Alzheimer’s Disease Pathology. Front Aging Neurosci. 2015, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J.; Hardy, J. The Amyloid Hypothesis of Alzheimer’s Disease at 25 years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Cline, E.N.; Bicca, M.A.; Viola, K.L.; Klein, W.L. The Amyloid-Beta Oligomer Hypothesis: Beginning of the Third Decade. J Alzheimers Dis. 2018, 64, S567–S610. [Google Scholar] [CrossRef] [Green Version]
- Ramusino, M.C.; Garibotto, V.; Bacchin, R.; Altomare, D.; Dodich, A.; Assal, F.; Mendes, A.; Costa, A.; Tinazzi, M.; Morbelli, S.D.; et al. Incremental Value of Amyloid-PET versus CSF in the Diagnosis of Alzheimer’s Disease. Eur. J. Nucl. Med. Mol. Imaging 2020, 47, 270–280. [Google Scholar] [CrossRef]
- Xin, S.-H.; Tan, L.; Cao, X.; Yu, J.-T.; Tan, L. Clearance of Amyloid Beta and Tau in Alzheimer’s Disease: From Mechanisms to Therapy. Neurotox. Res. 2018, 34, 733–748. [Google Scholar] [CrossRef]
- Tarasoff-Conway, J.M.; Carare, R.O.; Osorio, R.; Glodzik, L.; Butler, T.; Fieremans, E.; Axel, L.; Rusinek, H.; Nicholson, C.; Zlokovic, B.V.; et al. Clearance Systems in the Brain—Implications for Alzheimer Disease. Nat. Rev. Neurol. 2015, 11, 457–470. [Google Scholar] [CrossRef] [Green Version]
- Shibata, M.; Yamada, S.; Kumar, S.R.; Calero, M.; Bading, J.; Frangione, B.; Holtzman, D.M.; Miller, C.A.; Strickland, D.K.; Ghiso, J.; et al. Clearance of Alzheimer’s Amyloid-ss (1–40) Peptide from Brain by LDL Receptor-Related Protein-1 at the Blood-Brain Barrier. J. Clin. Investig. 2000, 106, 1489–1499. [Google Scholar] [CrossRef] [Green Version]
- Louveau, A.; Smirnov, I.; Keyes, T.J.; Eccles, J.D.; Rouhani, S.J.; Peske, J.D.; Derecki, N.C.; Castle, D.; Mandell, J.W.; Lee, K.S.; et al. Structural and Functional Features of Central Nervous System Lymphatic Vessels. Nature 2015, 523, 337–341. [Google Scholar] [CrossRef]
- Peng, W.; Achariyar, T.M.; Li, B.; Liao, Y.; Mestre, H.; Hitomi, E.; Regan, S.; Kasper, T.; Peng, S.; Ding, F.; et al. Suppression of Glymphatic Fluid Transport in a Mouse Model of Alzheimer’s Disease. Neurobiol. Dis. 2016, 93, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Harrison, I.F.; Ismail, O.; Machhada, A.; Colgan, N.; Ohene, Y.; Nahavandi, P.; Ahmed, Z.; Fisher, A.; Meftah, S.; Murray, T.K.; et al. Impaired Glymphatic Function and Clearance of Tau in an Alzheimer’s Disease Model. Brain 2020, 143, 2576–2593. [Google Scholar] [CrossRef]
- Da Mesquita, S.; Louveau, A.; Vaccari, A.; Smirnov, I.; Cornelison, R.C.; Kingsmore, K.; Contarino, C.; Onengut-Gumuscu, S.; Farber, E.; Raper, D.; et al. Functional Aspects of Meningeal Lymphatics in Ageing and Alzheimer’s Disease. Nature 2018, 560, 185–191. [Google Scholar] [CrossRef]
- Ahn, J.H.; Cho, H.; Kim, J.-H.; Kim, S.H.; Ham, J.-S.; Park, I.; Suh, S.H.; Hong, S.P.; Song, J.-H.; Hong, Y.-K.; et al. Meningeal Lymphatic Vessels at the Skull Base Drain Cerebrospinal Fluid. Nature 2019, 572, 62–66. [Google Scholar] [CrossRef]
- Bower, I.N.; Koltowska, K.; Pichol-Thievend, C.; Virshup, I.; Paterson, S.; Lagendijk, A.K.; Wang, W.; Lindsey, B.W.; Bent, S.; Baek, S.; et al. Mural Lymphatic Endothelial Cells Regulate Meningeal Angiogenesis in the Zebrafish. Nat. Neurosci. 2017, 20, 774–783. [Google Scholar] [CrossRef]
- Galanternik, M.V.; Castranova, D.; Gore, A.; Blewett, N.H.; Jung, H.M.; Stratman, A.N.; Kirby, M.R.; Iben, J.; Miller, M.F.; Kawakami, K.; et al. A Novel Perivascular Cell Population in the Zebrafish Brain. eLife 2017, 6, e24369. [Google Scholar] [CrossRef]
- Van Lessen, M.; Shibata-Germanos, S.; Van Impel, A.; Hawkins, A.T.; Rihel, J.; Schulte-Merker, S. Intracellular Uptake of Macromolecules by Brain Lymphatic Endothelial Cells During Zebrafish Embryonic Development. eLife 2017, 6, e25932. [Google Scholar] [CrossRef]
- Shibata-Germanos, S.; Goodman, J.R.; Greig, A.; Trivedi, C.A.; Benson, B.C.; Foti, S.C.; Faro, A.; Castellan, R.; Correra, R.M.; Barber, M.; et al. Structural and Functional Conservation of Non-Lumenized Lymphatic Endothelial Cells in the Mammalian Leptomeninges. Acta Neuropathol. 2019, 139, 383–401. [Google Scholar] [CrossRef] [Green Version]
- Padberg, Y.; van Impel, A.; van Lessen, M.; Bussmann, J.; Schulte-Merker, S. Meningeal Lymphatic Endothelial Cells Fulfill Scavenger Endothelial Cell Function and Employ Mrc1a for Cargo Uptake. BioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Takeda, S.; Hashimoto, T.; Roe, A.D.; Hori, Y.; Spires-Jones, T.L.; Hyman, B.T. Brain Interstitial Oligomeric Amyloid Beta Increases with Age and is Resistant to Clearance from Brain in a Mouse Model of Alzheimer’s Disease. FASEB J. 2013, 27, 3239–3248. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.Y.; Lee, D.K.; Chung, B.R.; Kim, H.V.; Kim, Y. Intracerebroventricular Injection of Amyloid-beta Peptides in Normal Mice to Acutely Induce Alzheimer-like Cognitive Deficits. J. Vis. Exp. 2016, 109, 53308. [Google Scholar]
- Jungbauer, L.M.; Yu, C.; Laxton, K.J.; Ladu, M.J. Preparation of Fluorescently-Labeled Amyloid-Beta Peptide Assemblies: The Effect of Fluorophore Conjugation on Structure and Function. J. Mol. Recognit. 2009, 22, 403–413. [Google Scholar] [CrossRef] [Green Version]
- McIntee, F.L.; Giannoni, P.; Blais, S.; Sommer, G.; Neubert, T.A.; Rostagno, A.; Ghiso, J. In vivo Differential Brain Clearance and Catabolism of Monomeric and Oligomeric Alzheimer’s Abeta protein. Front. Aging Neurosci. 2016, 8, 223. [Google Scholar] [CrossRef] [Green Version]
- Mazaheri, F.; Breus, O.; Durdu, S.; Haas, P.; Wittbrodt, J.; Gilmour, D.; Peri, F. Distinct roles for BAI1 and TIM-4 in the Engulfment of Dying Neurons by Microglia. Nat. Commun. 2014, 5, 4046. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Gu, B.J.; Masters, C.L.; Wang, Y.J. A Systemic View of Alzheimer Disease—Insights from Amyloid-Beta Metabolism Beyond the Brain. Nat. Rev. Neurol. 2017, 13, 612–623. [Google Scholar] [CrossRef]
- Tian, D.-Y.; Cheng, Y.; Zhuang, Z.-Q.; He, C.-Y.; Pan, Q.-G.; Tang, M.-Z.; Hu, X.-L.; Shen, Y.-Y.; Wang, Y.-R.; Chen, S.-H.; et al. Physiological Clearance of Amyloid-Beta by the Kidney and its Therapeutic Potential for Alzheimer’s Disease. Mol. Psychiatry 2021, 21, 1–9. [Google Scholar] [CrossRef]
- Drummond, I.A.; Davidson, A.J. Zebrafish Kidney Development. Methods Cell Biol. 2010, 100, 233–260. [Google Scholar] [CrossRef]
- Sehnert, A.J.; Huq, A.; Weinstein, B.M.; Walker, C.; Fishman, M.; Stainier, D.Y.R. Cardiac Troponin T is Essential in Sarcomere Assembly and Cardiac Contractility. Nat. Genet. 2002, 31, 106–110. [Google Scholar] [CrossRef]
- Oltrabella, F.; Pietka, G.; Ramirez, I.B.-R.; Mironov, A.; Starborg, T.; Drummond, I.A.; Hinchliffe, K.A.; Lowe, M. The Lowe Syndrome Protein OCRL1 Is Required for Endocytosis in the Zebrafish Pronephric Tubule. PLoS Genet. 2015, 11, e1005058. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.D.; Jin, S.W. A Tale of Two Models: Mouse and Zebrafish as Complementary Models for Lymphatic Studies. Mol. Cells 2014, 37, 503–510. [Google Scholar] [CrossRef] [Green Version]
- van Impel, A.; Zhao, Z.; Hermkens, D.M.A.; Roukens, M.G.; Fischer, J.C.; Peterson-Maduro, J.; Duckers, H.; Ober, E.A.; Ingham, P.W.; Schulte-Merker, S. Divergence of Zebrafish and Mouse Lymphatic Cell Fate Specification Pathways. Development 2014, 141, 1228–1238. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.N.; Long, H.; Mu, Y.; Chew, L.Y. The Toxicity of Amyloid Beta Oligomers. Int. J. Mol. Sci. 2012, 13, 7303–7327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogan, B.; Bos, F.L.; Bussmann, J.; Witte, M.; Chi, N.C.; Duckers, H.J.; Schulte-Merker, S. ccbe1 is Required for Embryonic Lymphangiogenesis and Venous Sprouting. Nat. Genet. 2009, 41, 396–398. [Google Scholar] [CrossRef] [PubMed]
- Volpe, B.A.; Fotino, T.H.; Steiner, A.B. Confocal Microscope-Based Laser Ablation and Regeneration Assay in Zebrafish Interneuromast Cells. J. Vis. Exp. 2020, 15, e60966. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Kim, H.V.; Jo, S.; Lee, C.J.; Choi, S.Y.; Kim, D.J.; Kim, Y. EPPS Rescues Hippocampus-Dependent Cognitive Deficits In APP/PS1 Mice by Disaggregation of Amyloid-Beta Oligomers and Plaques. Nat. Commun. 2015, 6, 8997. [Google Scholar] [CrossRef] [Green Version]
- De Strooper, B.; Karran, E. The Cellular Phase of Alzheimer’s Disease. Cell 2016, 164, 603–615. [Google Scholar] [CrossRef] [Green Version]
- Yeung, J.H.Y.; Palpagama, T.H.; Tate, W.P.; Peppercorn, K.; Waldvogel, H.J.; Faull, R.L.M.; Kwakowsky, A. The Acute Effects of Amyloid-Beta1–42 on Glutamatergic Receptor and Transporter Expression in the Mouse Hippocampus. Front. Neurosci. 2019, 13, 1427. [Google Scholar] [CrossRef] [Green Version]
- Javed, I.; Peng, G.; Xing, Y.; Yu, T.; Zhao, M.; Kakinen, A.; Faridi, A.; Parish, C.; Ding, F.; Davis, T.P.; et al. Inhibition of Amyloid Beta Toxicity in Zebrafish with a Chaperone-Gold Nanoparticle Dual Strategy. Nat. Commun. 2019, 10, 378. [Google Scholar] [CrossRef]
- Bhattarai, P.; Thomas, A.K.; Cosacak, M.I.; Papadimitriou, C.; Mashkaryan, V.; Froc, C.; Reinhardt, S.; Kurth, T.; Dahl, A.; Zhang, Y.; et al. IL4/STAT6 Signaling Activates Neural Stem Cell Proliferation and Neurogenesis upon Amyloid-beta42 Aggregation in Adult Zebrafish Brain. Cell Rep. 2016, 17, 941–948. [Google Scholar] [CrossRef] [Green Version]
- Özcan, G.G.; Lim, S.; La Leighton, P.; Allison, W.T.; Rihel, J. Sleep is Bi-Directionally Modified by Amyloid Beta Oligomers. eLife 2020, 9, 3995. [Google Scholar] [CrossRef]
- Castranova, D.; Samasa, B.; Galanternik, M.V.; Jung, H.M.; Pham, V.N.; Weinstein, B.M. Live Imaging of Intracranial Lymphatics in the Zebrafish. Circ. Res. 2021, 128, 42–58. [Google Scholar] [CrossRef]
- Mato, M.; Ookawara, S.; Aikawa, E.; Kawasaki, K. Studies on Fluorescent Granular Perithelium (F.G.P.) of Rat Cerebral Cortex —Especially Referring to Morphological Changes in Aging. Anat. Anzeiger. 1981, 149, 486–501. [Google Scholar]
- Kierdorf, K.; Masuda, T.; Jordão, M.J.C.; Prinz, M. Macrophages at CNS interfaces: Ontogeny and Function in Health and Disease. Nat. Rev. Neurosci. 2019, 20, 547–562. [Google Scholar] [CrossRef]
- Weller, R.O.; Subash, M.; Preston, S.D.; Mazanti, I.; Carare, R.O. Perivascular Drainage of Amyloid-Beta Peptides from the Brain and its Failure in Cerebral Amyloid Angiopathy and Alzheimer’s Disease. Brain Pathol. 2008, 18, 253–266. [Google Scholar] [CrossRef]
- Mato, M.; Ookawara, S. Influences of Age and Vasopressin on the Uptake Capacity of Fluorescent Granular Perithelial Cells (FGP) of Small Cerebral Vessels of the Rat. Am. J. Anat. 1981, 162, 45–53. [Google Scholar] [CrossRef]
- Mato, M.; Ookawara, S.; Kurihara, K. Uptake of Exogenous Substances and Marked Infoldings of the Fluorescent Granular Pericyte in Cerebral Fine Vessels. Am. J. Anat. 1980, 157, 329–332. [Google Scholar] [CrossRef]
- Hawkes, C.A.; McLaurin, J. Selective Targeting of Perivascular Macrophages for Clearance of Beta-Amyloid in Cerebral Amyloid Angiopathy. Proc. Natl. Acad. Sci. USA 2009, 106, 1261–1266. [Google Scholar] [CrossRef] [Green Version]
- Mawuenyega, K.G.; Sigurdson, W.; Ovod, V.; Munsell, L.; Kasten, T.; Morris, J.C.; Yarasheski, K.E.; Bateman, R.J. Decreased Clearance of CNS b-Amyloid in Alzheimer’s Disease. Science 2010, 330, 1774. [Google Scholar] [CrossRef] [Green Version]
- Verghese, P.B.; Castellano, J.M.; Garai, K.; Wang, Y.; Jiang, H.; Shah, A.; Bu, G.; Frieden, C.; Holtzman, D.M. ApoE Influences Amyloid-Beta (Abeta) Clearance Despite Minimal Apoe/Abeta Association in Physiological Conditions. Proc. Natl. Acad. Sci. USA 2013, 110, E1807–E1816. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Sagare, A.P.; Ma, Q.; Halliday, M.R.; Kong, P.; Kisler, K.; Winkler, E.A.; Ramanathan, A.; Kanekiyo, T.; Bu, G.; et al. Central role for PICALM in Amyloid-Beta Blood-Brain Barrier Transcytosis and Clearance. Nat. Neurosci. 2015, 18, 978–987. [Google Scholar] [CrossRef] [Green Version]
- Michaud, J.-P.; Bellavance, M.-A.; Préfontaine, P.; Rivest, S. Real-Time In Vivo Imaging Reveals the Ability of Monocytes to Clear Vascular Amyloid Beta. Cell Rep. 2013, 5, 646–653. [Google Scholar] [CrossRef] [Green Version]
- Zagrean, A.-M.; Hermann, D.M.; Opris, I.; Zagrean, L.; Popa-Wagner, A. Multicellular Crosstalk Between Exosomes and the Neurovascular Unit after Cerebral Ischemia. Therapeutic Implications. Front. Neurosci. 2018, 12, 811. [Google Scholar] [CrossRef]
- Glabe, C.G. Structural Classification of Toxic Amyloid Oligomers. J. Biol. Chem. 2008, 283, 29639–29643. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Lee, S.W.; Lee, G.; Yoon, D.S. Atomic Force Microscopy Analysis of EPPS-Driven Degradation and Reformation of Amyloid-beta Aggregates. J. Alzheimers Dis. Rep. 2018, 2, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.M.; Castranova, D.; Swift, M.R.; Pham, V.N.; Galanternik, M.V.; Isogai, S.; Butler, M.G.; Mulligan, T.S.; Weinstein, B.M. Development of the Larval Lymphatic System in the Zebrafish. Development 2017, 144, 2070–2081. [Google Scholar] [CrossRef] [Green Version]
- Kwan, K.M.; Fujimoto, E.; Grabher, C.; Mangum, B.D.; Hardy, M.E.; Campbell, D.; Parant, J.M.; Yost, H.J.; Kanki, J.P.; Chien, C.-B. The Tol2kit: A Multisite Gateway-Based Construction Kit Fortol2 Transposon Transgenesis Constructs. Dev. Dyn. 2007, 236, 3088–3099. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, Y.-M.; Lee, J.-G.; Cho, H.-J.; Lee, W.S.; Jeong, J.; Lee, J.-S. Differential Clearance of A? Species from the Brain by Brain Lymphatic Endothelial Cells in Zebrafish. Int. J. Mol. Sci. 2021, 22, 11883. https://doi.org/10.3390/ijms222111883
Jeong Y-M, Lee J-G, Cho H-J, Lee WS, Jeong J, Lee J-S. Differential Clearance of A? Species from the Brain by Brain Lymphatic Endothelial Cells in Zebrafish. International Journal of Molecular Sciences. 2021; 22(21):11883. https://doi.org/10.3390/ijms222111883
Chicago/Turabian StyleJeong, Yun-Mi, Jae-Geun Lee, Hyun-Ju Cho, Wang Sik Lee, Jinyoung Jeong, and Jeong-Soo Lee. 2021. "Differential Clearance of A? Species from the Brain by Brain Lymphatic Endothelial Cells in Zebrafish" International Journal of Molecular Sciences 22, no. 21: 11883. https://doi.org/10.3390/ijms222111883
APA StyleJeong, Y.-M., Lee, J.-G., Cho, H.-J., Lee, W. S., Jeong, J., & Lee, J.-S. (2021). Differential Clearance of A? Species from the Brain by Brain Lymphatic Endothelial Cells in Zebrafish. International Journal of Molecular Sciences, 22(21), 11883. https://doi.org/10.3390/ijms222111883