
Fibronectin 1B Gene Plays an Important Role in Loach Barbel Air-Breathing


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
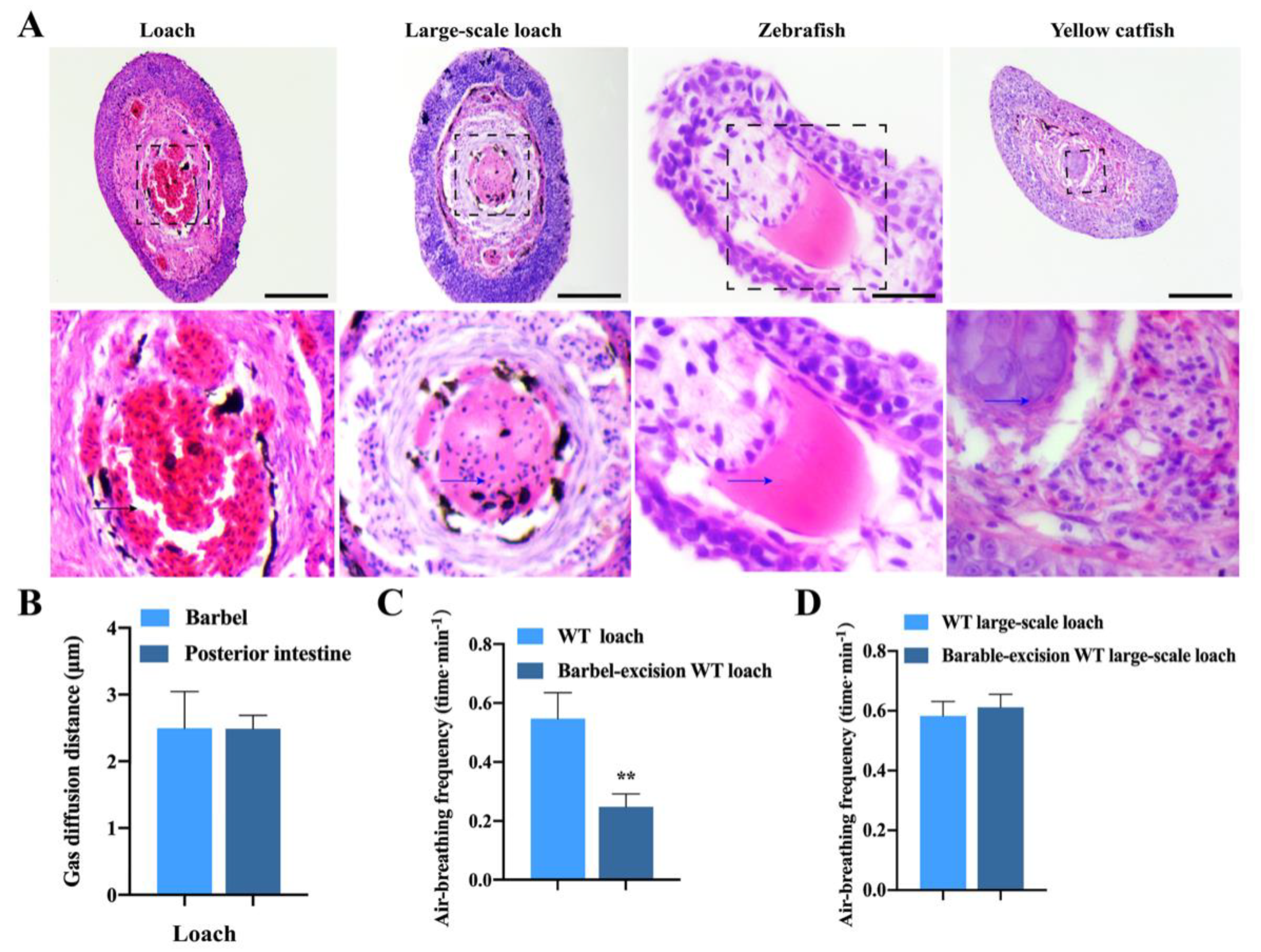
2.1. Histological Structures of Loach Barbels Suitable for Air-Breathing
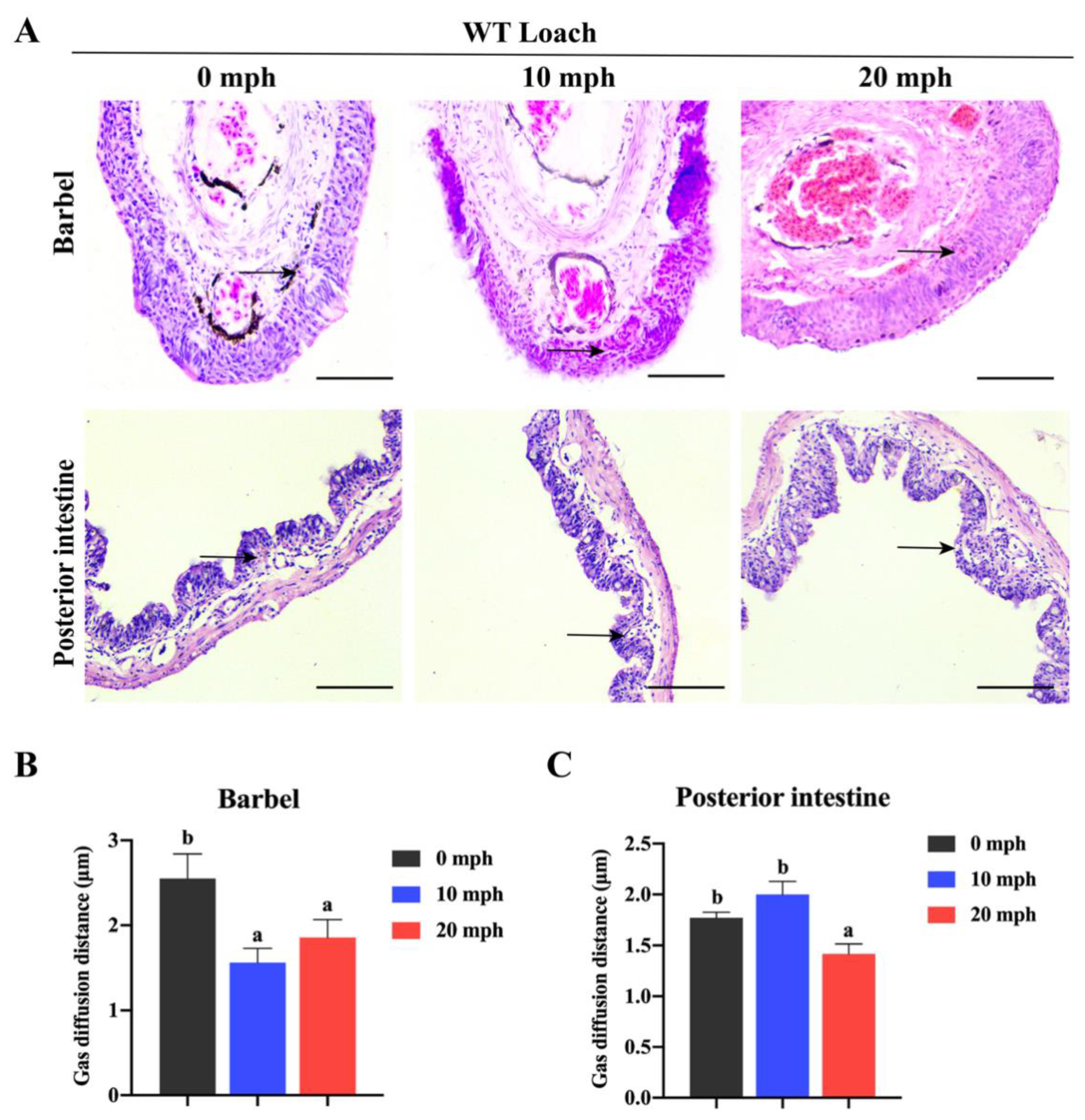
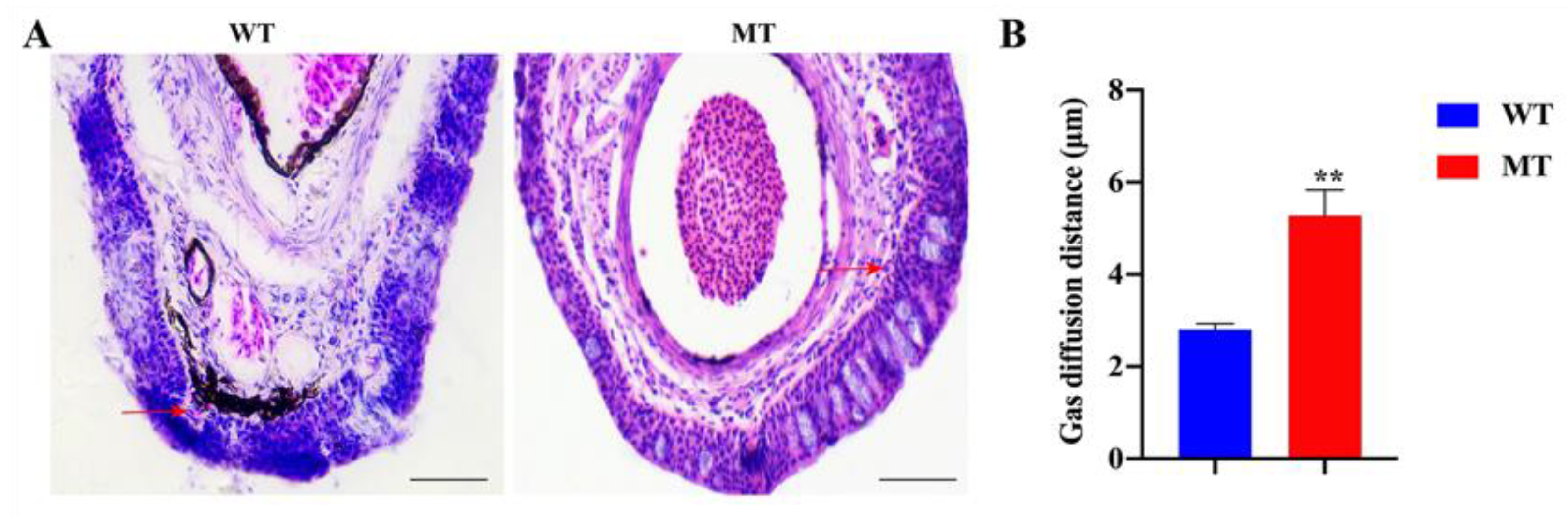
2.2. The Changes in Histological Structures and Gas Diffusion Distances of Loach Barbels in Response to Acute Hypoxia
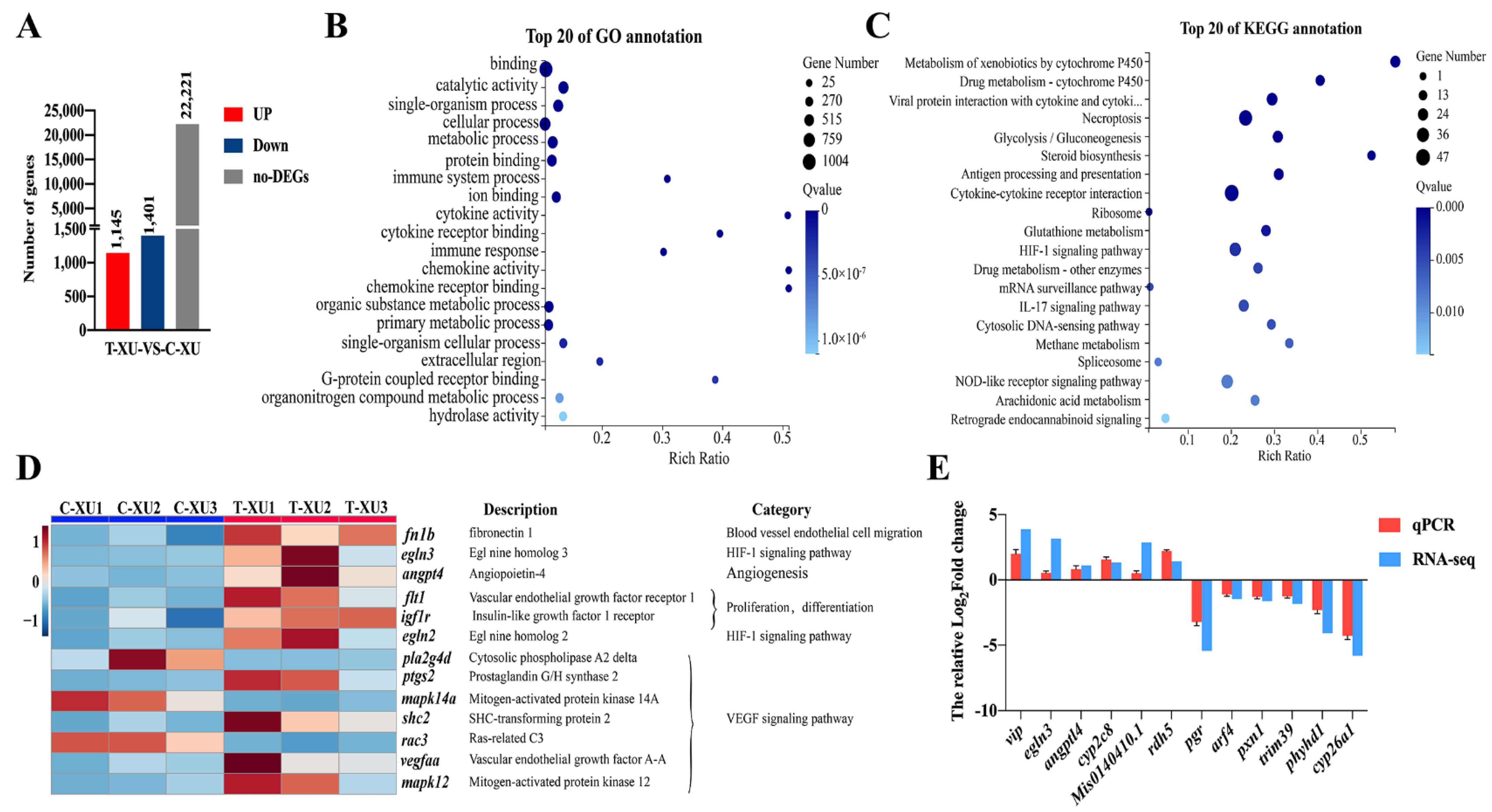
2.3. Transcriptome Analysis of Loach Barbels with Air Exposure
2.3.1. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment Analysis of Differentially Expressed Genes (DEGs)
2.3.2. Mining of DEGs Related to Barbel Air-Breathing
2.3.3. qPCR Validation of RNA-Seq Data
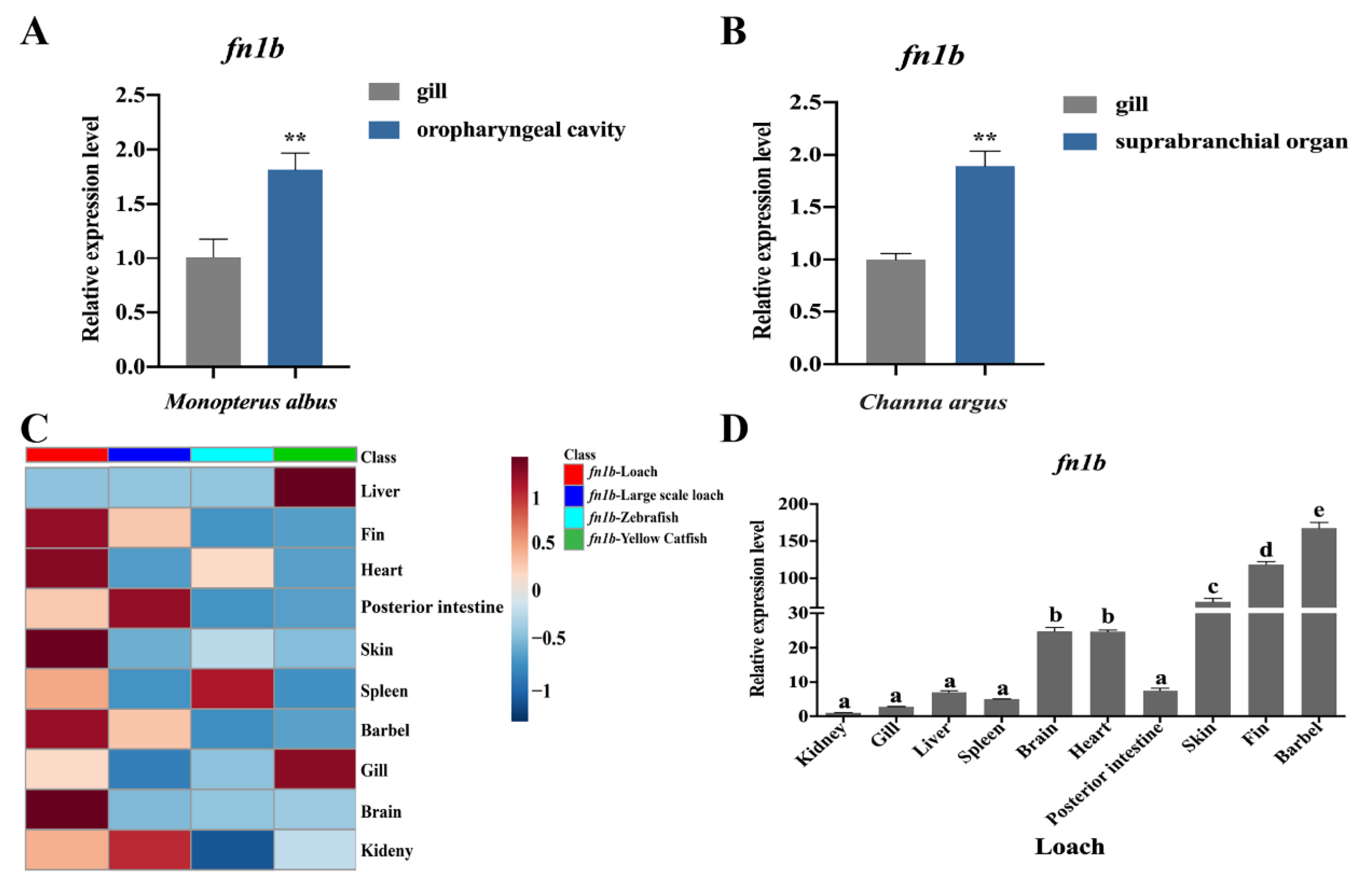
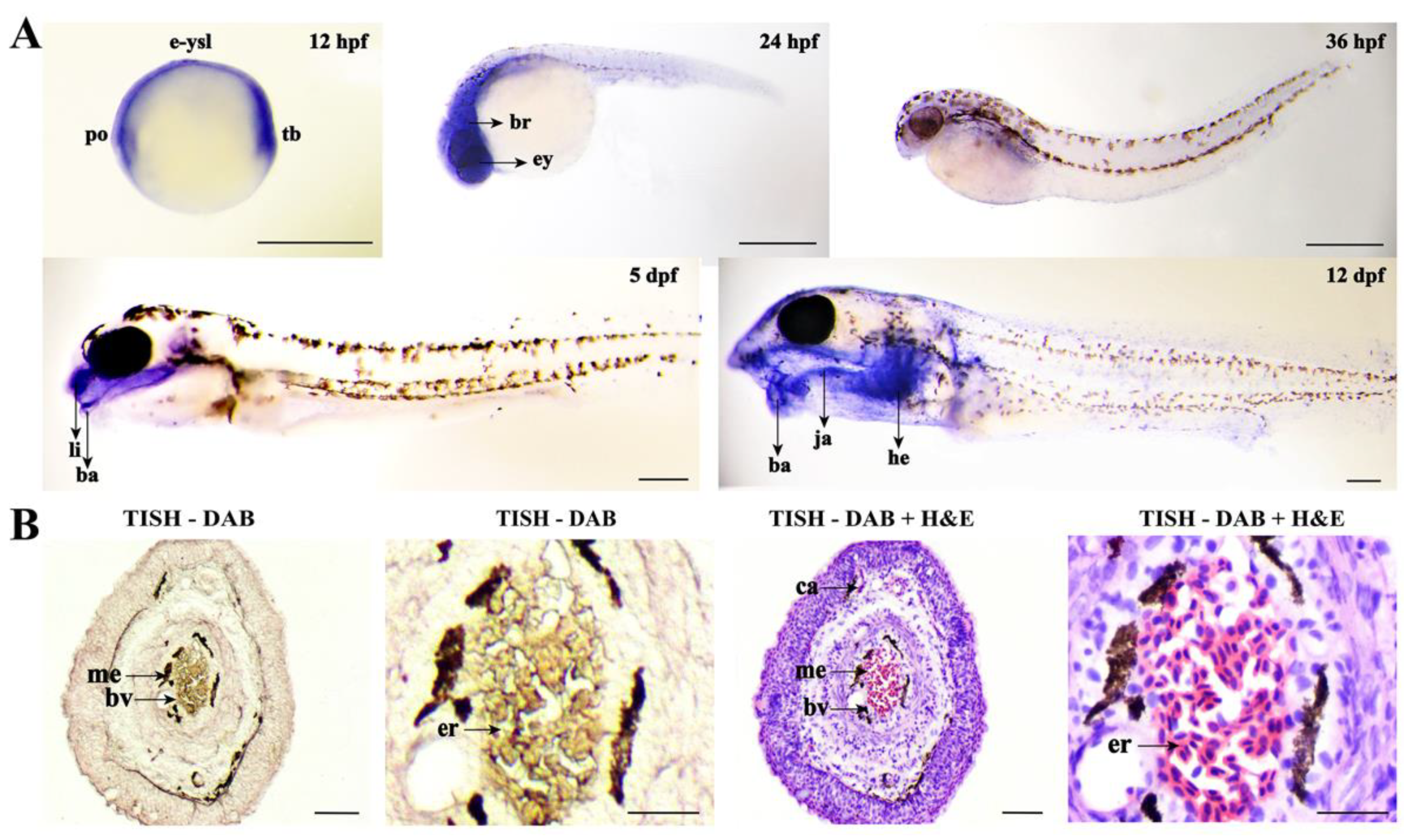
2.4. Sequence and Expression Profiles of Fn1b in Loach
2.5. The Depletion of Fn1b Weakened the Air-Breathing of Loach Barbel
3. Discussion
4. Materials and Methods
4.1. Fish Species
4.2. Morphological and Histological Observations and Estimations of Blood–Gas Diffusion Distance and Air-Breathing Frequency
4.3. Transcriptome Analysis of Loach Barbel
4.3.1. Air Exposure Experiment
4.3.2. RNA Isolation and cDNA Library Constructions
4.3.3. Sequencing and Read Mapping
4.3.4. Identification of DEGs and Functional Annotations
4.3.5. qPCR Validation of Transcriptome Data
4.4. Sequence, Expression, and Location Analyses of fn1b
4.4.1. Sequence Analysis
4.4.2. qPCR Analysis
4.4.3. In Situ Hybridization
4.5. Effects of fn1b-Depletion on Loach Barbel Air-Breathing
4.5.1. fn1b+/− Loach Produced by CRISPR/Cas9
4.5.2. Histological Observations of Barbels from WT and MT Loach
4.5.3. Chronic Hypoxia Experiment
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Data Accessibility
References
- Ma, X.; Su, B.; Bangs, M.; Alston, V.; Backenstose, N.; Simora, R.M.; Wang, W.; Xing, D.; Li, S.; Ye, Z. Comparative Genomic and Transcriptomic Analyses Revealed Twenty-Six Candidate Genes Involved in the Air-Breathing Development and Function of the Bighead Catfish Clarias macrocephalus. Mar. Biotechnol. 2021, 23, 90–105. [Google Scholar] [CrossRef]
- Wang, K.; Wang, J.; Zhu, C.; Yang, L.; Wang, W. African lungfish genome sheds light on the vertebrate water-to-land transition. Cell 2021, 5, 1362–1376. [Google Scholar] [CrossRef]
- Dobson, G.E. Notes on the Respiration of some Species of Indian Freshwater Fishes. Proc. Zool. Soc. Lond. 1874, 42, 312–321. [Google Scholar] [CrossRef]
- Nelson, J.A. Breaking wind to survive: Fishes that breathe air with their gut. J. Fish. Biol. 2014, 84, 554–576. [Google Scholar] [CrossRef]
- Munshi, J.; Olson, K.R.; Ojha, J.; Ghosh, T.K. Morphology and vascular anatomy of the accessory respiratory organs of the air-breathing climbing perch, Anabas testudineus (Bloch). Dev. Dyn. 2010, 176, 321–331. [Google Scholar] [CrossRef]
- Johansen, K. Air breathing in Fishes. Fish. Physiol. 1970, 4, 361–411. [Google Scholar] [CrossRef]
- Huebner, E.; Chee, G. Histological and ultrastructural specialization of the digestive tract of the intestinal air breather hoplosternum thoracatum (teleost). J. Morphol. 1978, 157, 301–327. [Google Scholar] [CrossRef]
- Satora, L. Histological and ultrastructural study of the stomach of the air-breathing Ancistrus multispinnis (Siluriformes, Teleostei). Can. J. Zool. 1998, 76, 83–86. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Li, X.H.; Zhao, J.R.; Wang, Z.J. Effect of Intestinal Air-breathing Restriction on Respiratory Metabolism and Antioxidant Capability of Loach (Paramisgurnus dabryanus). Chin. J. Zool. 2017, 5, 857–864. [Google Scholar] [CrossRef]
- Hughes, G.M.; Singh, B.N. Respiration in an air-breathing fish, the climbing perch Anabas testudineus. J. Exp. Biol. 1970, 53, 265–280. [Google Scholar] [CrossRef]
- Hakim, A.; Datta Munshi, J.S.; Hughes, G.M. Morphometries of the respiratory organs of the Indian green snake-headed fish. Channa punctata. J. Zool. 1978, 184, 519–543. [Google Scholar] [CrossRef]
- Yadav, A.N.; Singh, B.R. The gut of an intestinal air-breathing fish, Lepidocephalus guntea (Ham). Arch. Biol. 1980, 91, 413–422. [Google Scholar]
- Weibel, E.R. Morphological basis of alveolar-capillary gas exchange. Physiol. Rev. 1973, 53, 419–495. [Google Scholar] [CrossRef] [PubMed]
- García, L. Histology, ultrastructure and immunohistochemistry of the respiratory organs in non mammalian vertebrates. Biochem. Educ. 1995, 10, 125. [Google Scholar] [CrossRef]
- Frommel, A.Y.; Kwan, G.T.; Prime, K.J.; Tresguerres, M.; Brauner, C.J. Changes in gill and air-breathing organ characteristics during the transition from waterto air-breathing in juvenile Arapaima gigas. J. Exp. Zool. A Ecol. Integr. Physiol. 2021, 42, 312–321. [Google Scholar] [CrossRef]
- Bi, X.; Wang, K.; Yang, L.; Pan, H.; Jiang, H.; Wei, Q.; Fang, M.; Yu, H.; Zhu, C.; Cai, Y. Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes. Cell 2021, 5, 1377–1391. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, B.; Koner, D.; Hasan, R.; Saha, N. Molecular characterization and ornithine-urea cycle genes expression in air-breathing magur catfish (Clarias magur) during exposure to high external ammonia. Genomics 2019, 112, 2247–2260. [Google Scholar] [CrossRef]
- Li, N.; Bao, L.; Zhou, T.; Yuan, Z.; Liu, S.; Dunham, R.; Li, Y.; Wang, K.; Xu, X.; Jin, Y. Genome sequence of walking catfish (Clarias batrachus) provides insights into terrestrial adaptation. BMC Genom. 2018, 19, 952. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.W.; Cao, X.J.; Xu, X.; Huang, S.Q.; Liu, C.S.; Tomljanovic, T. Developmental transcriptome analysis and identification of genes involved in formation of intestinal air-breathing function of Loach, Misgurnus anguillicaudatus. Sci Rep. 2016, 6, 31845. [Google Scholar] [CrossRef]
- Huang, S.Q.; Cao, X.J.; Tian, X. Transcriptomic Analysis of Compromise Between Air-Breathing and Nutrient Uptake of Posterior Intestine in Loach (Misgurnus anguillicaudatus), an Air-Breathing Fish. Mar. Biotechnol. 2016, 18, 1–13. [Google Scholar] [CrossRef]
- Jiang, Y.; Feng, S.; Xu, J.; Zhang, S.; Li, S.; Sun, X.; Xu, P. Comparative transcriptome analysis between aquatic and aerial breathing organs of Channa argus to reveal the genetic basis underlying bimodal respiration. Mar. Genom. 2016, 29, 89–96. [Google Scholar] [CrossRef]
- Yang, Y.J. Comparative Transcriptome Analysis of Catfish Swimbladder Reveals Expression Signatures in Chamber Formation and their Response to Low Oxygen Stress. Ph.D. Dissertation, Auburn University, Auburn, AL, USA, 2018. Available online: http://hdl.handle.net/10415/6301 (accessed on 25 August 2021).
- Norris, K.E. Growth, Fecundity, and Diet of Oriental Weatherloach Misgurnus Anguillicaudatus in the Chicago Area Waterways; Western Illinois University: Macomb, IL, USA, 2015. [Google Scholar] [CrossRef]
- Mcmahon, R. Resriatory physiology of intestinal air breathing in the teleost fish Misgurnus anguillicaudatus. J. Exp. Biol. 1987, 133, 371–393. [Google Scholar] [CrossRef]
- Huang, S.Q.; Cao, X.J.; Tian, X.; Wang, W.M. High-Throughput Sequencing Identifies MicroRNAs from Posterior Intestine of Loach (Misgurnus anguillicaudatus) and Their Response to Intestinal Air-Breathing Inhibition. PLoS ONE 2016, 11, e0149123. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.W.; Liang, X.; Huang, S.Q.; Cao, X.J. Molecular cloning, expression analysis and miRNA prediction of vascular endothelial growth factor A (VEGFAa and VEGFAb) in pond loach Misgurnus anguillicaudatus, an air-breathing fish. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2016, 202, 39–47. [Google Scholar] [CrossRef]
- Leclair, E.E.; Jacek, T. Methods for the Study of the Zebrafish Maxillary Barbel. J. Vis. Exp. 2009, 33, 1494–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leclair, E.E.; Jacek, T.; Bruce, R. Development and regeneration of the zebrafish maxillary barbel: A novel study system for vertebrate tissue growth and repair. PLoS ONE 2010, 5, e8737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duszynski, R.J.; Topczewski, J.; Leclair, E.E. Divergent requirements for FGF signaling in zebrafish maxillary barbel and caudal fin regeneration. Dev. Growth Differ. 2013, 55, 282–300. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Li, N.; Jin, Y.; Zeng, Q.; Prabowo, W.; Liu, Y.; Tian, C.; Bao, L.; Liu, S.; Yuan, Z. Chemokine C-C motif ligand 33 is a key regulator of teleost fish barbel development. Proc. Nat. Acad. Sci. USA 2018, 115, 5018–5027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.X.; Yang, C.; Huang, L.F.; Zeng, K.; Cao, X.J. Inefficient ATP synthesis by inhibiting mitochondrial respiration causes lipids to decrease in MSTN-lacking muscles of loach Misgurnus anguillicaudatus. Funct. Integr. Genom. 2019, 19, 889–900. [Google Scholar] [CrossRef] [PubMed]
- Jayaram, K.C. Functional responses of catfish barbels. Bull. Zool. Surv. India 1978, 1, 77–80. [Google Scholar]
- Haynes, G.D.; Gongora, J.; Gilligan, D.M.; Grewe, P.; Moran, C.; Nicholas, F.W. Cryptic hybridization and introgression between invasive Cyprinid species Cyprinus carpio and Carassius auratus in Australia: Implications for invasive species management. Anim. Conserv. 2012, 15, 83–94. [Google Scholar] [CrossRef]
- Olson, K.R.; Roy, P.K.; Ghosh, T.K.; Munshi, J. Microcirculation of gills and accessory respiratory organs from the air-breathing snakehead fish, Channa punctata, C. gaucha and C. marulius. Anat. Rec. 2010, 238, 92–107. [Google Scholar] [CrossRef]
- Niva, B.; Ojha, J.; Munshi, J. Morphometrics of the respiratory organs of an estuarine goby, Boleophthalmus boddaerti. Japan J. Ichthyol. 1981, 27, 316–326. [Google Scholar] [CrossRef]
- Wu, C.L.; Dicks, A.; Steward, N.; Tang, R.; Guilak, F. Single cell transcriptomic analysis of human pluripotent stem cell chondrogenesis. Nat. Commun. 2021, 12, 362. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, R.; Wang, X.; Zhu, H.; Tian, Z. Transcriptome analysis reveals molecular mechanisms responsive to acute cold stress in the tropical stenothermal fish tiger barb (Puntius tetrazona). BMC Genom. 2020, 21, 737. [Google Scholar] [CrossRef]
- Le, A.T.; Stainier, D. Fibronectin regulates epithelial organization during myocardial migration in zebrafish. Dev. Cell 2004, 6, 371–382. [Google Scholar] [CrossRef] [Green Version]
- Wijelath, E.S. Novel Vascular Endothelial Growth Factor Binding Domains of Fibronectin Enhance Vascular Endothelial Growth Factor Biological Activity. Circ. Res. 2002, 91, 25–31. [Google Scholar] [CrossRef] [Green Version]
- George, E.L.; Baldwin, H.S.; Hynes, R.O. Fibronectins are essential for heart and blood vessel morphogenesis but are dispensable for initial specification of precursor cells. Blood 1997, 90, 3073–3081. [Google Scholar] [CrossRef]
- Mcdougal, B.A. A Comparative Study of Tissue Oxygen Consumption Correlated with the Molting Cycle in the Fresh Water Crayfish, Procambarus Simulans; Texas Tech University: Lubbock, TX, USA, 1965; Available online: http://hdl.handle.net/2346/1 (accessed on 25 August 2021).
- Gordon, M.S.; Chin, H.G.; Vojkovich, M. Energetics of swimming in fishes using different methods of locomotion: I. Labriform swimmers. Fish Physiol. Biochem. 1989, 6, 341–352. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, X.; Collodi, P. Identification and characterization of a novel fibronectin in zebrafish. Exp. Cell Res. 2001, 268, 211–219. [Google Scholar] [CrossRef]
- Chang, Y.; Shiu, J.; Chuang, W. Design, Structure Determination, and Biological Evaluation of Potent Integrin Alpha5beta1-Specific Antagonist Using the Ninth and Tenth Module of Fibronectin Type III Domain. Ph.D. Dissertation, National Cheng Kung University, Taiwan, 2013. [Google Scholar]
- Cao, X.J.; Wang, W.M. Histology and Mucin Histochemistry of The Digestive Tract of Yellow Catfish, Pelteobagrus fulvidraco. Anat. Histol. Embryol. 2010, 38, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Dewey, C.N.; Li, B. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Mao, X.; Huang, J.; Yang, D.; Wu, J.; Dong, S.; Lei, K.; Ge, G.; Li, C.Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.X.; Zhai, Y.H.; Gui, J.F. Molecular characterization and expression pattern of AFPIV during embryogenesis in gibel carp (Carassiu auratus gibelio). Mol. Biol. Rep. 2009, 36, 2011–2018. [Google Scholar] [CrossRef]
- Randilini, A.; Fujikawa, K.; Shibata, S. An in situ hybridization study of decorin and biglycan mRNA in mouse osteoblasts in vivo. Anat. Sci. Int. 2021, 96, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Mateos, M.A.; Vejnar, C.E.; Beaudoin, J.D.; Fernandez, J.P.; Mis, E.K.; Khokha, M.K.; Giraldez, A.J. CRISPRscan: Designing highly efficient sgRNAs for CRISPR-Cas9 targeting in vivo. Nat. Methods 2015, 12, 982–988. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.X.; Ren, T.Y.; Li, X.; Cao, X.J.; Gao, J. Polyunsaturated fatty acids synthesized by freshwater fish: A new insight to the roles of elovl2 and elovl5 in vivo. Biochem. Biophys. Res. Commun. 2020, 532, 414–419. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, B.; Huang, S.; Huang, L.; Yang, L.; Gao, J.; Cao, X. Fibronectin 1B Gene Plays an Important Role in Loach Barbel Air-Breathing. Int. J. Mol. Sci. 2021, 22, 11928. https://doi.org/10.3390/ijms222111928
Sun B, Huang S, Huang L, Yang L, Gao J, Cao X. Fibronectin 1B Gene Plays an Important Role in Loach Barbel Air-Breathing. International Journal of Molecular Sciences. 2021; 22(21):11928. https://doi.org/10.3390/ijms222111928
Chicago/Turabian StyleSun, Bing, Songqian Huang, Longfei Huang, Lijuan Yang, Jian Gao, and Xiaojuan Cao. 2021. "Fibronectin 1B Gene Plays an Important Role in Loach Barbel Air-Breathing" International Journal of Molecular Sciences 22, no. 21: 11928. https://doi.org/10.3390/ijms222111928
APA StyleSun, B., Huang, S., Huang, L., Yang, L., Gao, J., & Cao, X. (2021). Fibronectin 1B Gene Plays an Important Role in Loach Barbel Air-Breathing. International Journal of Molecular Sciences, 22(21), 11928. https://doi.org/10.3390/ijms222111928