
CD38 Correlates with an Immunosuppressive Treg Phenotype in Lupus-Prone Mice

 ,
,  , , ,
, , ,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
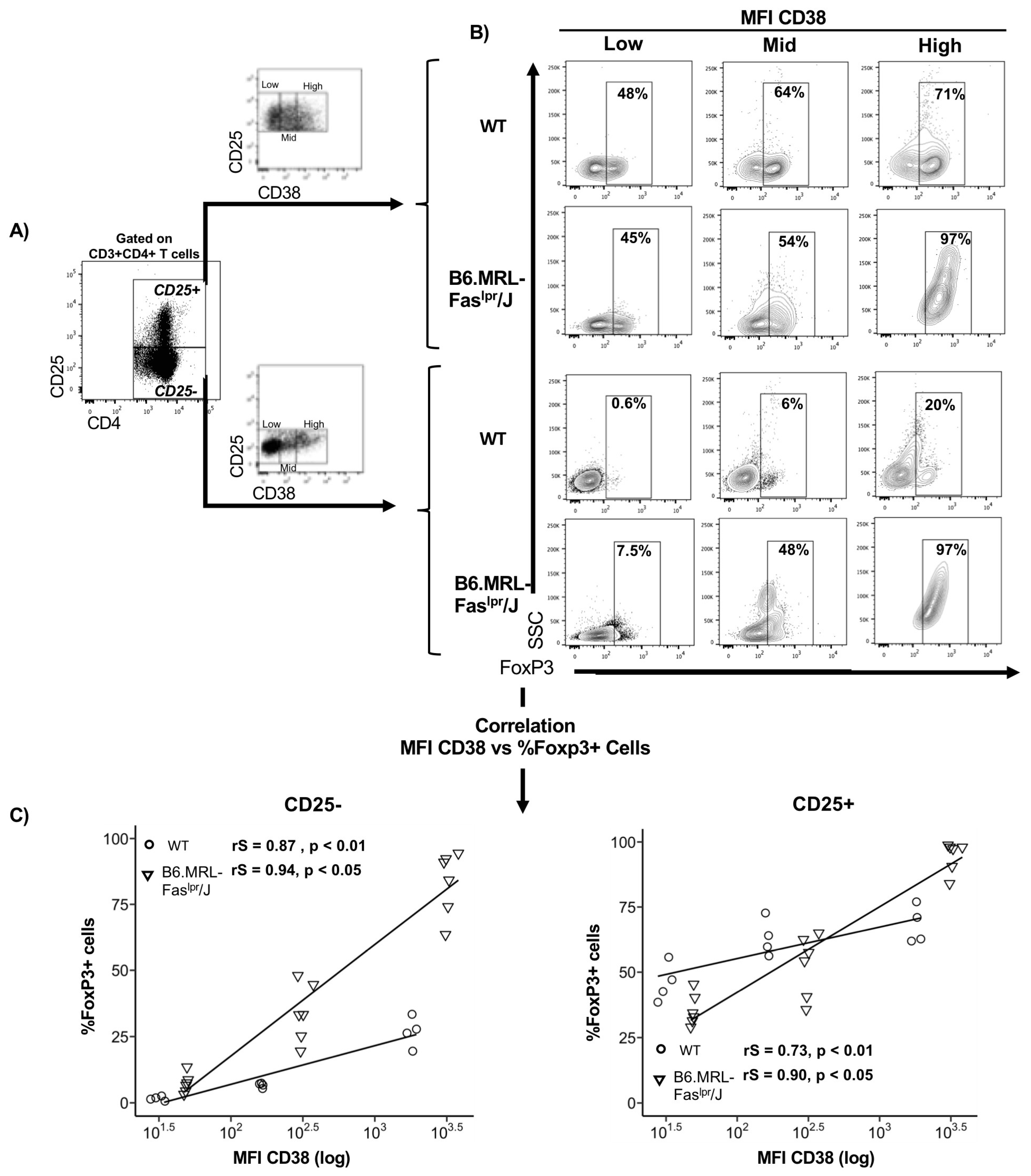
2.1. CD38 Expression Positively Correlates with FoxP3 Expression
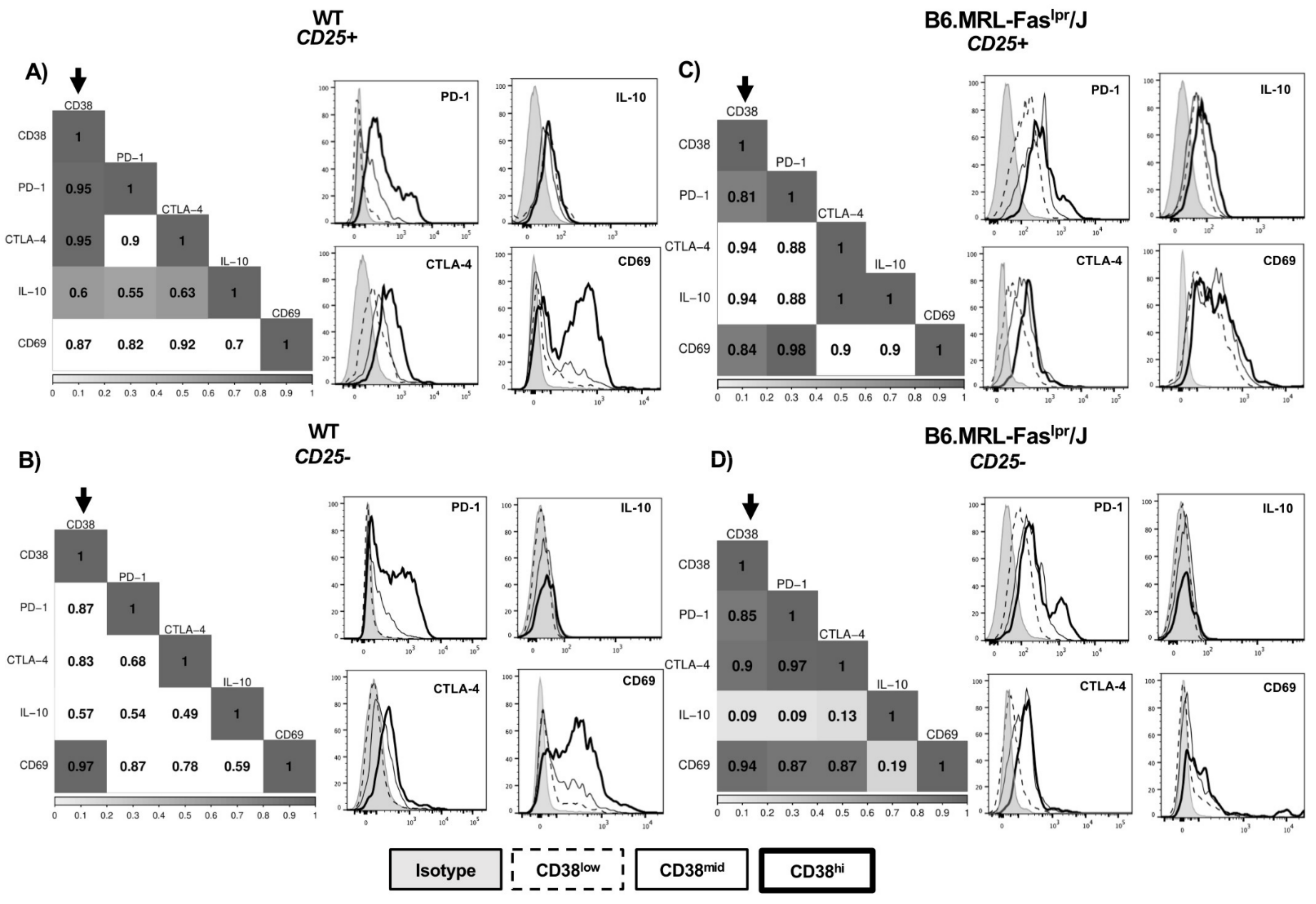
2.2. CD38 Expression Positively Correlates with Immunosuppressive Molecules
2.3. Reduced CD38+ Treg Proportion in B6.MRL-Faslpr/J Mice
2.4. Reduced Treg Expansion and IL-10 Secretion in Stimulated CD38-/- Splenocytes
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Isolation of Splenocytes
4.3. Flow Cytometry
4.4. Expansion Treg Assay
4.5. IFN-γ and IL-10 Detection
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Fava, A.; Petri, M. Systemic Lupus Erythematosus: Diagnosis and Clinical Management. J. Autoimmun. 2019, 96, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Nusbaum, J.S.; Mirza, I.; Shum, J.; Freilich, R.W.; Cohen, R.E.; Pillinger, M.H.; Izmirly, P.M.; Buyon, J.P. Sex Differences in Systemic Lupus Erythematosus: Epidemiology, Clinical Considerations, and Disease Pathogenesis. Mayo Clin. Proc. 2020, 95, 384–394. [Google Scholar] [CrossRef] [PubMed]
- Karagianni, P.; Tzioufas, A.G. Epigenetic Perspectives on Systemic Autoimmune Disease. J. Autoimmun. 2019, 104, 102315. [Google Scholar] [CrossRef] [PubMed]
- Yeoh, S.-A.; Dias, S.S.; Isenberg, D.A. Advances in Systemic Lupus Erythematosus. Medicine 2018, 46, 84–92. [Google Scholar] [CrossRef]
- Gray-McGuire, C.; Moser, K.L.; Gaffney, P.M.; Kelly, J.; Yu, H.; Olson, J.M.; Jedrey, C.M.; Jacobs, K.B.; Kimberly, R.P.; Neas, B.R.; et al. Genome Scan of Human Systemic Lupus Erythematosus by Regression Modeling: Evidence of Linkage and Epistasis. Am. J. Hum. Genet. 2000, 67, 1460–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindqvist, A.K.; Alarcón-Riquelme, M.E. The Genetics of Systemic Lupus Erythematosus. Scand. J. Immunol. 1999, 50, 562–571. [Google Scholar] [CrossRef]
- Nakagawara, K.; Mori, M.; Takasawa, S.; Nata, K.; Takamura, T.; Berlova, A.; Tohgo, A.; Karasawa, T.; Yonekura, H.; Takeuchi, T. Assignment of CD38, the Gene Encoding Human Leukocyte Antigen CD38 (ADP-Ribosyl Cyclase/Cyclic ADP-Ribose Hydrolase), to Chromosome 4p15. Cytogenet. Genome Res. 1995, 69, 38–39. [Google Scholar] [CrossRef] [PubMed]
- Funaro, A.; Spagnoli, G.C.; Ausiello, C.M.; Alessio, M.; Roggero, S.; Delia, D.; Zaccolo, M.; Malavasi, F. Involvement of the Multilineage CD38 Molecule in a Unique Pathway of Cell Activation and Proliferation. J. Immunol. 1990, 145, 2390–2396. [Google Scholar]
- States, D.J.; Walseth, T.F.; Lee, H.C. Similarities in Amino Acid Sequences of Aplysia ADP-Ribosyl Cyclase and Human Lymphocyte Antigen CD38. Trends Biochem. Sci. 1992, 17, 495. [Google Scholar] [CrossRef]
- Wei, W.; Graeff, R.; Yue, J. Roles and Mechanisms of the CD38/Cyclic Adenosine Diphosphate Ribose/Ca2+ Signaling Pathway. World J. Biol. Chem. 2014, 5, 58–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Argumedo, L.; Teixera, C.; Preece, G.; Kirkham, P.; Parkhouse, R.M. A B Lymphocyte Surface Molecule Mediating Activation and Protection from Apoptosis via Calcium Channels. J. Immunol. 1993, 151, 3119–3130. [Google Scholar] [PubMed]
- Partida-Sánchez, S.; Goodrich, S.; Kusser, K.; Oppenheimer, N.; Randall, T.D.; Lund, F.E. Regulation of Dendritic Cell Trafficking by the ADP-Ribosyl Cyclase CD38: Impact on the Development of Humoral Immunity. Immunity 2004, 20, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Lund, F.E.; Muller-Steffner, H.; Romero-Ramirez, H.; Moreno-García, M.E.; Partida-Sánchez, S.; Makris, M.; Oppenheimer, N.J.; Santos-Argumedo, L.; Schuber, F. CD38 Induces Apoptosis of a Murine Pro-B Leukemic Cell Line by a Tyrosine Kinase-Dependent but ADP-Ribosyl Cyclase- and NAD Glycohydrolase-Independent Mechanism. Int. Immunol. 2006, 18, 1029–1042. [Google Scholar] [CrossRef]
- Rodríguez-Alba, J.C.; Moreno-García, M.E.; Sandoval-Montes, C.; Rosales-Garcia, V.H.; Santos-Argumedo, L. CD38 Induces Differentiation of Immature Transitional 2 B Lymphocytes in the Spleen. Blood 2008, 111, 3644–3652. [Google Scholar] [CrossRef] [Green Version]
- Alcocer-Varela, J.; Alarcón-Riquelme, M.; Laffón, A.; Sánchez-Madrid, F.; Alarcón-Segovia, D. Activation Markers on Peripheral Blood T-cells from Patients with Active or Inactive Systemic Lupus Erythematosus. Correlation with Proliferative Responses and Production of IL-2. J. Autoimmun. 1991, 4, 935–945. [Google Scholar] [CrossRef]
- al-Janadi, M.; Raziuddin, S. B Cell Hyperactivity Is a Function of T-cell Derived Cytokines in Systemic Lupus Erythematosus. J. Rheumatol. 1993, 20, 1885–1891. [Google Scholar]
- Pavón, E.J.; Muñoz, P.; Navarro, M.-C.; Raya-Alvarez, E.; Callejas-Rubio, J.-L.; Navarro-Pelayo, F.; Ortego-Centeno, N.; Sancho, J.; Zubiaur, M. Increased Association of CD38 with Lipid Rafts in T-cells from Patients with Systemic Lupus Erythematosus and in Activated Normal T-cells. Mol. Immunol. 2006, 43, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Spronk, P.E.; Horst, G.; Van Der Gun, B.T.; Limburg, P.C.; Kallenberg, C.G. Anti-DsDNA Production Coincides with Concurrent B and T-cell Activation during Development of Active Disease in Systemic Lupus Erythematosus (SLE). Clin. Exp. Immunol. 1996, 104, 446–453. [Google Scholar] [CrossRef]
- Pavón, E.J.; Zumaquero, E.; Rosal-Vela, A.; Khoo, K.-M.; Cerezo-Wallis, D.; García-Rodríguez, S.; Carrascal, M.; Abian, J.; Graeff, R.; Callejas-Rubio, J.-L.; et al. Increased CD38 Expression in T-cells and Circulating Anti-CD38 IgG Autoantibodies Differentially Correlate with Distinct Cytokine Profiles and Disease Activity in Systemic Lupus Erythematosus Patients. Cytokine 2013, 62, 232–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Chen, Y.-G.; Reifsnyder, P.C.; Schott, W.H.; Lee, C.-H.; Osborne, M.; Scheuplein, F.; Haag, F.; Koch-Nolte, F.; Serreze, D.V.; et al. Targeted Disruption of CD38 Accelerates Autoimmune Diabetes in NOD/Lt Mice by Enhancing Autoimmunity in an ADP-Ribosyltransferase 2-Dependent Fashion. J. Immunol. 2006, 176, 4590–4599. [Google Scholar] [CrossRef] [Green Version]
- Viegas, M.S.; Silva, T.; Monteiro, M.M.; do Carmo, A.; Martins, T.C. Knocking out of CD38 Accelerates Development of a Lupus-like Disease in Lpr Mice. Rheumatology 2011, 50, 1569–1577. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Pantoja, M.; López-Herrera, G.; Romero-Ramírez, H.; Santos-Argumedo, L.; Chávez-Rueda, A.K.; Hernández-Cueto, Á.; Flores-Muñoz, M.; Rodríguez-Alba, J.C. CD38 Protein Deficiency Induces Autoimmune Characteristics and Its Activation Enhances IL-10 Production by Regulatory B Cells. Scand. J. Immunol. 2018, 87, e12664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horwitz, D.A. Regulatory T-cells in Systemic Lupus Erythematosus: Past, Present and Future. Arthritis Res. Ther. 2008, 10, 227. [Google Scholar] [CrossRef] [Green Version]
- Sakaguchi, S.; Sakaguchi, N.; Asano, M.; Itoh, M.; Toda, M. Immunologic Self-Tolerance Maintained by Activated T-cells Expressing IL-2 Receptor Alpha-Chains (CD25). Breakdown of a Single Mechanism of Self-Tolerance Causes Various Autoimmune Diseases. J. Immunol. 1995, 155, 1151–1164. [Google Scholar]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T-cells and Immune Tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parry, R.V.; Chemnitz, J.M.; Frauwirth, K.A.; Lanfranco, A.R.; Braunstein, I.; Kobayashi, S.V.; Linsley, P.S.; Thompson, C.B.; Riley, J.L. CTLA-4 and PD-1 Receptors Inhibit T-Cell Activation by Distinct Mechanisms. Mol. Cell. Biol. 2005, 25, 9543–9553. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, P.; Penninger, J.M.; Timms, E.; Wakeham, A.; Shahinian, A.; Lee, K.P.; Thompson, C.B.; Griesser, H.; Mak, T.W. Lymphoproliferative Disorders with Early Lethality in Mice Deficient in Ctla-4. Science 1995, 270, 985–988. [Google Scholar] [CrossRef]
- Taylor, A.; Verhagen, J.; Blaser, K.; Akdis, M.; Akdis, C.A. Mechanisms of Immune Suppression by Interleukin-10 and Transforming Growth Factor-Beta: The Role of T Regulatory Cells. Immunology 2006, 117, 433–442. [Google Scholar] [CrossRef]
- Wei, F.; Zhong, S.; Ma, Z.; Kong, H.; Medvec, A.; Ahmed, R.; Freeman, G.J.; Krogsgaard, M.; Riley, J.L. Strength of PD-1 Signaling Differentially Affects T-Cell Effector Functions. Proc. Natl. Acad. Sci. USA 2013, 110, E2480–E2489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontenot, J.D.; Gavin, M.A.; Rudensky, A.Y. Foxp3 Programs the Development and Function of CD4+CD25+ Regulatory T-cells. Nat. Immunol. 2003, 4, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhang, L.; Acharya, C.; An, G.; Wen, K.; Qiu, L.; Munshi, N.C.; Tai, Y.-T.; Anderson, K.C. Targeting CD38 Suppresses Induction and Function of T Regulatory Cells to Mitigate Immunosuppression in Multiple Myeloma. Clin. Cancer Res. 2017, 23, 4290–4300. [Google Scholar] [CrossRef] [Green Version]
- Krejcik, J.; Casneuf, T.; Nijhof, I.S.; Verbist, B.; Bald, J.; Plesner, T.; Syed, K.; Liu, K.; van de Donk, N.W.C.J.; Weiss, B.M.; et al. Daratumumab Depletes CD38+ Immune Regulatory Cells, Promotes T-Cell Expansion, and Skews T-Cell Repertoire in Multiple Myeloma. Blood 2016, 128, 384–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.O.; Rudensky, A.Y. T-cell Receptor Signaling in the Control of Regulatory T-cell Differentiation and Function. Nat. Rev. Immunol. 2016, 16, 220–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Shen, S.; Gorentla, B.K.; Gao, J.; Zhong, X.-P. Murine Regulatory T-cells Contain Hyperproliferative and Death-Prone Subsets with Differential ICOS Expression. J. Immunol. 2012, 188, 1698–1707. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Cucchetti, M.; Roncagalli, R.; Yokosuka, T.; Malzac, A.; Bertosio, E.; Imbert, J.; Nijman, I.J.; Suchanek, M.; Saito, T.; et al. The Lymphoid Lineage—Specific Actin-Uncapping Protein Rltpr Is Essential for Costimulation via CD28 and the Development of Regulatory T-cells. Nat. Immunol. 2013, 14, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, M.; Majdic, O.; Knapp, W.; Holter, W. High-Level IL-10 Production by Monoclonal Antibody-Stimulated Human T-cells. Immunology 1995, 86, 364–371. [Google Scholar]
- Yanagawa, Y.; Iwabuchi, K.; Onoé, K. Co-Operative Action of Interleukin-10 and Interferon-γ to Regulate Dendritic Cell Functions. Immunology 2009, 127, 345–353. [Google Scholar] [CrossRef]
- Saraiva, M.; O’Garra, A. The Regulation of IL-10 Production by Immune Cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Read, S.; Mauze, S.; Asseman, C.; Bean, A.; Coffman, R.; Powrie, F. CD38+ CD45RBlow CD4+ T-cells: A Population of T-cells with Immune Regulatory Activities in Vitro. Eur. J. Immunol. 1998, 28, 3435–3447. [Google Scholar] [CrossRef]
- Cohen, P.L.; Eisenberg, R.A. Lpr and Gld: Single Gene Models of Systemic Autoimmunity and Lymphoproliferative Disease. Annu. Rev. Immunol. 1991, 9, 243–269. [Google Scholar] [CrossRef]
- van Nieuwenhuijze, A.E.; Cauwe, B.; Klatt, D.; Humblet-Baron, S.; Liston, A. Lpr-Induced Systemic Autoimmunity Is Unaffected by MasT-cell Deficiency. Immunol. Cell Biol. 2015, 93, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Patton, D.T.; Wilson, M.D.; Rowan, W.C.; Soond, D.R.; Okkenhaug, K. The PI3K P110δ Regulates Expression of CD38 on Regulatory T-cells. PLoS ONE 2011, 6, e17359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drach, J.; McQueen, T.; Engel, H.; Andreeff, M.; Robertson, K.A.; Collins, S.J.; Malavasi, F.; Mehta, K. Retinoic Acid-Induced Expression of CD38 Antigen in Myeloid Cells Is Mediated through Retinoic Acid Receptor-Alpha. Cancer Res. 1994, 54, 1746–1752. [Google Scholar] [PubMed]
- Mucida, D.; Pino-Lagos, K.; Kim, G.; Nowak, E.; Benson, M.J.; Kronenberg, M.; Noelle, R.J.; Cheroutre, H. Retinoic Acid Can Directly Promote TGF-Beta-Mediated Foxp3(+) Treg Cell Conversion of Naive T-cells. Immunity 2009, 30, 472–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Xiao, Y.; Zeng, Z.; Shi, Y.; Tang, B.; Long, H.; Kanekura, T.; Wang, J.; Wu, H.; Zhao, M.; et al. All-Trans Retinoic Acid Induces CD4+CD25+FOXP3+ Regulatory T-cells by Increasing FOXP3 Demethylation in Systemic Sclerosis CD4+ T-cells. J. Immunol. Res. 2018, 2018, 8658156. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Liu, Y. The Nature of Increased Circulating CD4+CD25−Foxp3+ T-cells in Patients with Systemic Lupus Erythematosus: A Novel Hypothesis. Open Rheumatol. J. 2009, 3, 22–24. [Google Scholar] [CrossRef] [PubMed]
- Zelenay, S.; Lopes-Carvalho, T.; Caramalho, I.; Moraes-Fontes, M.F.; Rebelo, M.; Demengeot, J. Foxp3+ CD25- CD4 T-cells Constitute a Reservoir of Committed Regulatory Cells That Regain CD25 Expression upon Homeostatic Expansion. Proc. Natl. Acad. Sci. USA 2005, 102, 4091–4096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonelli, M.; Göschl, L.; Blüml, S.; Karonitsch, T.; Steiner, C.-W.; Steiner, G.; Smolen, J.S.; Scheinecker, C. CD4+CD25−Foxp3+ T-cells: A Marker for Lupus Nephritis? Arthritis Res. Ther. 2014, 16, R104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, R.C.; Simons, H.Z.; Thompson, W.S.; Rainbow, D.B.; Yang, X.; Cutler, A.J.; Oliveira, J.; Castro Dopico, X.; Smyth, D.J.; Savinykh, N.; et al. Cells with Treg-Specific FOXP3 Demethylation but Low CD25 Are Prevalent in Autoimmunity. J. Autoimmun. 2017, 84, 75–86. [Google Scholar] [CrossRef]
- Han, Y.; Guo, Q.; Zhang, M.; Chen, Z.; Cao, X. CD69+ CD4+ CD25− T-cells, a New Subset of Regulatory T-cells, Suppress T-cell Proliferation through Membrane-Bound TGF-Beta 1. J. Immunol. 2009, 182, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altman, A.; Theofilopoulos, A.N.; Weiner, R.; Katz, D.H.; Dixon, F.J. Analysis of T-cell Function in Autoimmune Murine Strains. Defects in Production and Responsiveness to Interleukin 2. J. Exp. Med. 1981, 154, 791–808. [Google Scholar] [CrossRef] [Green Version]
- Wofsy, D.; Murphy, E.D.; Roths, J.B.; Dauphinée, M.J.; Kipper, S.B.; Talal, N. Deficient Interleukin 2 Activity in MRL/Mp and C57BL/6J Mice Bearing the Lpr Gene. J. Exp. Med. 1981, 154, 1671–1680. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhao, Y. The Regulation of Foxp3 Expression in Regulatory CD4(+)CD25(+)T-cells: Multiple Pathways on the Road. J. Cell. Physiol. 2007, 211, 590–597. [Google Scholar] [CrossRef]
- Horenstein, A.L.; Chillemi, A.; Zaccarello, G.; Bruzzone, S.; Quarona, V.; Zito, A.; Serra, S.; Malavasi, F. A CD38/CD203a/CD73 Ectoenzymatic Pathway Independent of CD39 Drives a Novel Adenosinergic Loop in Human T Lymphocytes. Oncoimmunology 2013, 2, e26246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, R.; Hou, J.; Li, Y.; Bian, J.; Deng, X.; Zhu, X.; Yang, T. Adenosine Promotes Foxp3 Expression in Treg Cells in Sepsis Model by Activating JNK/AP-1 Pathway. Am. J. Transl. Res. 2016, 8, 2284–2292. [Google Scholar] [PubMed]
- Ohta, A.; Kini, R.; Ohta, A.; Subramanian, M.; Madasu, M.; Sitkovsky, M. The Development and Immunosuppressive Functions of CD4(+) CD25(+) FoxP3(+) Regulatory T-cells Are under Influence of the Adenosine-A2A Adenosine Receptor Pathway. Front. Immunol. 2012, 3, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubert, S.; Rissiek, B.; Klages, K.; Huehn, J.; Sparwasser, T.; Haag, F.; Koch-Nolte, F.; Boyer, O.; Seman, M.; Adriouch, S. Extracellular NAD+ Shapes the Foxp3+ Regulatory T-cell Compartment through the ART2–P2X7 Pathway. J. Exp. Med. 2010, 207, 2561–2568. [Google Scholar] [CrossRef]
- Aswad, F.; Kawamura, H.; Dennert, G. High Sensitivity of CD4+CD25+ Regulatory T-cells to Extracellular Metabolites Nicotinamide Adenine Dinucleotide and ATP: A Role for P2X7 Receptors. J. Immunol. 2005, 175, 3075–3083. [Google Scholar] [CrossRef] [Green Version]
- Aswad, F.; Dennert, G. P2X7 Receptor Expression Levels Determine Lethal Effects of a Purine Based Danger Signal in T Lymphocytes. Cell. Immunol. 2006, 243, 58–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, P.; Navarro, M.-C.; Pavón, E.J.; Salmerón, J.; Malavasi, F.; Sancho, J.; Zubiaur, M. CD38 Signaling in T-cells Is Initiated within a Subset of Membrane Rafts Containing Lck and the CD3-Zeta Subunit of the T-cell Antigen Receptor. J. Biol. Chem. 2003, 278, 50791–50802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zubiaur, M.; Izquierdo, M.; Terhorst, C.; Malavasi, F.; Sancho, J. CD38 Ligation Results in Activation of the Raf-1/Mitogen-Activated Protein Kinase and the CD3-Zeta/Zeta-Associated Protein-70 Signaling Pathways in Jurkat T Lymphocytes. J. Immunol. 1997, 159, 193–205. [Google Scholar] [PubMed]
- Zubiaur, M.; Guirado, M.; Terhorst, C.; Malavasi, F.; Sancho, J. The CD3-Gamma Delta Epsilon Transducing Module Mediates CD38-Induced Protein-Tyrosine Kinase and Mitogen-Activated Protein Kinase Activation in Jurkat T-cells. J. Biol. Chem. 1999, 274, 20633–20642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zubiaur, M.; Fernández, O.; Ferrero, E.; Salmerón, J.; Malissen, B.; Malavasi, F.; Sancho, J. CD38 Is Associated with Lipid Rafts and upon Receptor Stimulation Leads to Akt/Protein Kinase B and Erk Activation in the Absence of the CD3-Zeta Immune Receptor Tyrosine-Based Activation Motifs. J. Biol. Chem. 2002, 277, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burlock, B.; Richardson, G.; García-Rodríguez, S.; Guerrero, S.; Zubiaur, M.; Sancho, J. The Role of CD38 on the Function of Regulatory B Cells in a Murine Model of Lupus. Int. J. Mol. Sci. 2018, 19, 2906. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Lara, J.C.; Espinosa, E.; Santos-Argumedo, L.; Romero-Ramírez, H.; López-Herrera, G.; García-García, F.; Sandoval-Montes, C.; Ortiz-Navarrete, V.; Flores-Muñoz, M.; Rodríguez-Alba, J.C. CD38 Correlates with an Immunosuppressive Treg Phenotype in Lupus-Prone Mice. Int. J. Mol. Sci. 2021, 22, 11977. https://doi.org/10.3390/ijms222111977
Pérez-Lara JC, Espinosa E, Santos-Argumedo L, Romero-Ramírez H, López-Herrera G, García-García F, Sandoval-Montes C, Ortiz-Navarrete V, Flores-Muñoz M, Rodríguez-Alba JC. CD38 Correlates with an Immunosuppressive Treg Phenotype in Lupus-Prone Mice. International Journal of Molecular Sciences. 2021; 22(21):11977. https://doi.org/10.3390/ijms222111977
Chicago/Turabian StylePérez-Lara, Jocelyn C., Enrique Espinosa, Leopoldo Santos-Argumedo, Héctor Romero-Ramírez, Gabriela López-Herrera, Fabio García-García, Claudia Sandoval-Montes, Vianney Ortiz-Navarrete, Mónica Flores-Muñoz, and Juan C. Rodríguez-Alba. 2021. "CD38 Correlates with an Immunosuppressive Treg Phenotype in Lupus-Prone Mice" International Journal of Molecular Sciences 22, no. 21: 11977. https://doi.org/10.3390/ijms222111977