
NTRK Gene Fusion Detection in Atypical Spitz Tumors

 ,
,  , , ,
, , ,  , , , ,
, , , , 
 and
and
Abstract
:1. Introduction
2. Results
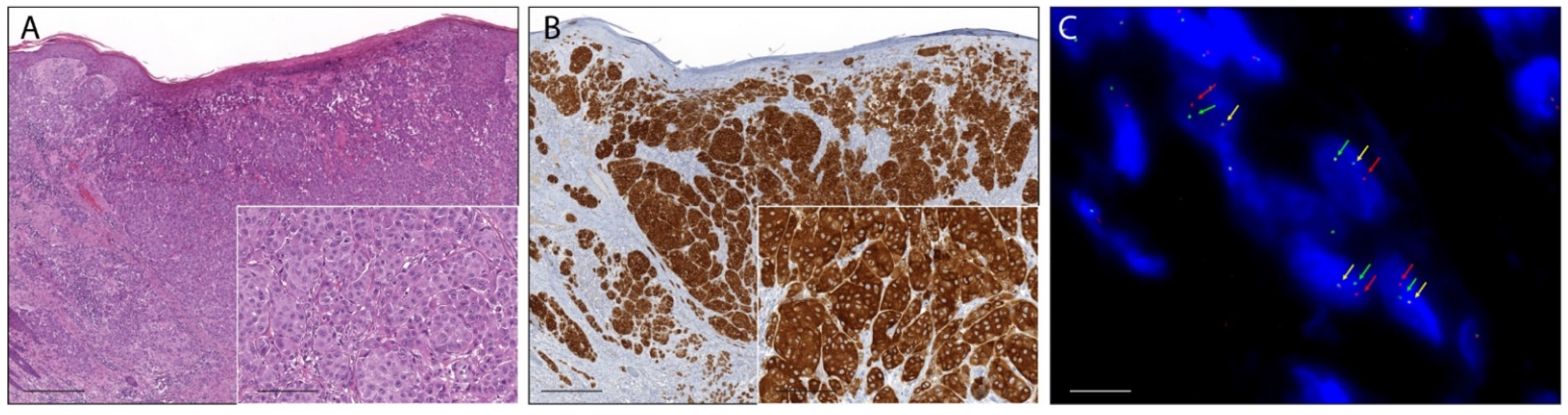
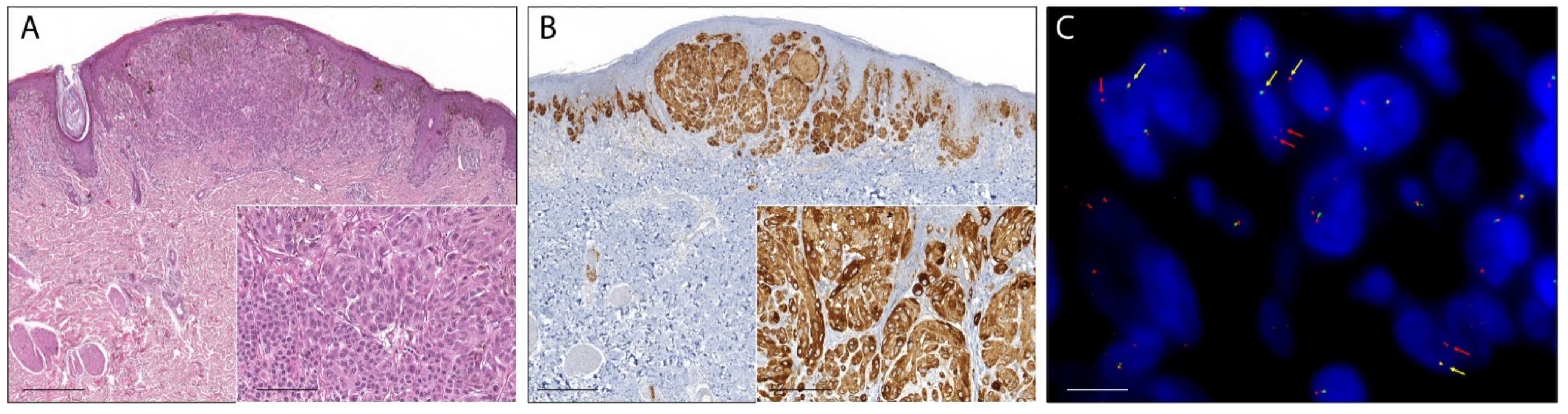
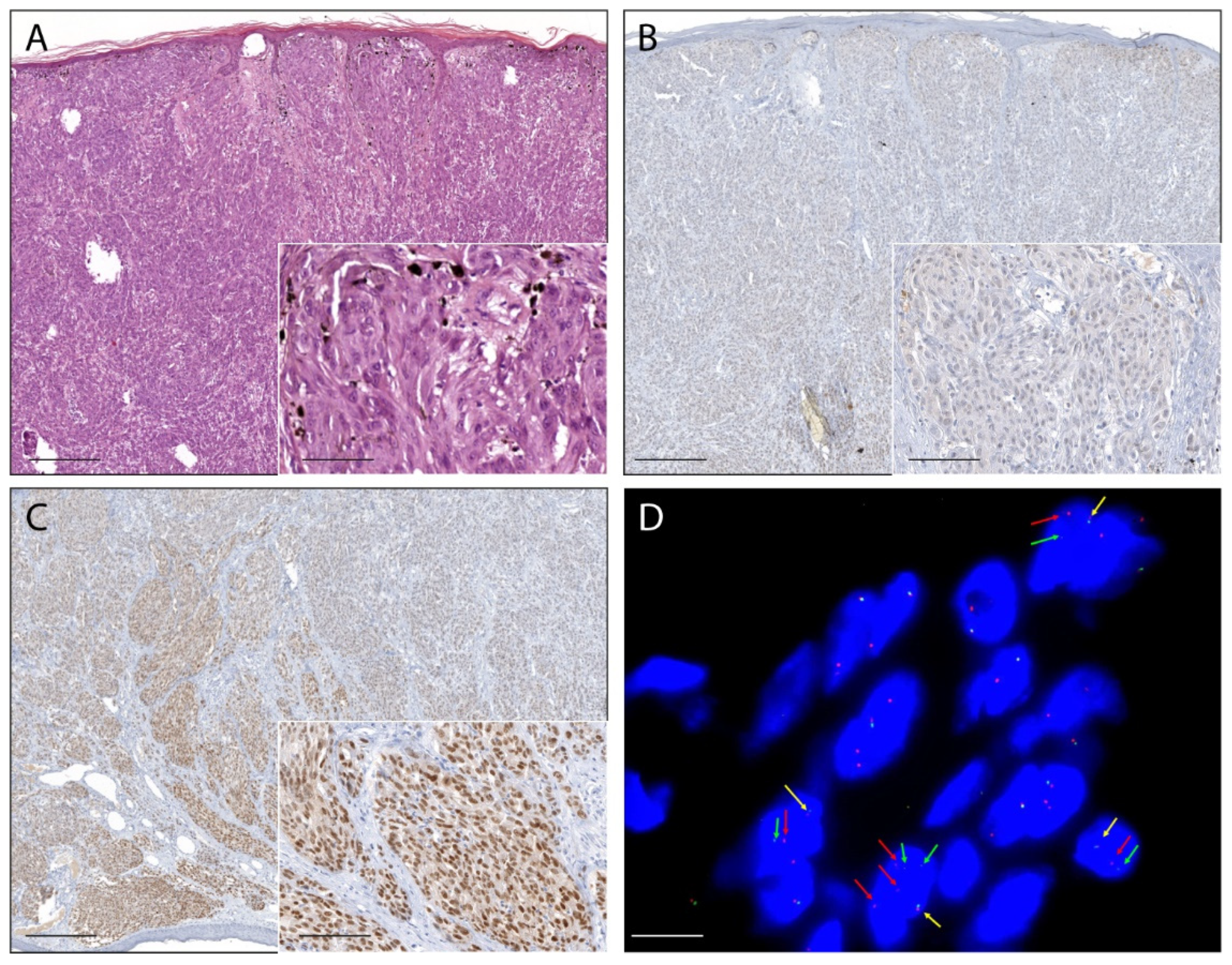
2.1. Pan-Trk Immunohistochemistry
2.2. RNA-Based NGS
2.3. Real-Time RT-PCR
2.4. Fluorescence In Situ Hybridization
3. Discussion
4. Materials and Methods
4.1. Case Selection and Tumour Specimen Collection
4.2. Immunohistochemistry
4.3. RNA Extraction and Quantification
4.4. RNA Libraries Preparation and NGS Sequencing
4.5. Sequencing Data Analysis
4.6. Real-Time-PCR
4.7. Fluorescence In Situ Hybridization
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| NTRK1 exon 9–10 | TFG-NTRK1 T5:N9 |
| NFASC-NTRK1 N20:N10 | |
| NFASC-NTRK1 N21:N10 | |
| NTRK1 exon 10 | SQSTM1-NTRK1 S4:N10 |
| TFG-NTRK1 T5:N10 | |
| TPM3-NTRK1 T7:N10 | |
| TPR-NTRK1 T21:N10 | |
| TRIM63-NTRK1 T8:N10 | |
| NTRK1 exon 11–12 del | BCAN-NTRK1 B13:N11 |
| LMNA-NTRK1 L11 del178:N11 | |
| LMNA-NTRK1 L2:N11 | |
| PPL-NTRK1 P22 del2181:N11 | |
| TPR-NTRK1 T6 del122:N12 del99 | |
| NTRK1 exon 12 | MPRIP-NTRK1 M21:N12 |
| MPRIP-NTRK1 M18:N12 | |
| MPRIP-NTRK1 M14:N12 | |
| SCYL3-NTRK1 S11:N12 | |
| TPM3-NTRK1 T7:N12 | |
| TPR-NTRK1 T6:N12 | |
| NTRK2 exon 12–15 | AFAP1-NTRK2 A13:N12 |
| VCL-NTRK2 V16:N12 | |
| TLE4-NTRK2 T7:N15 | |
| TRIM24-NTRK2 T12:N15 | |
| NTRK2 exon 16–17 | AGBL4-NTRK2 A6:N16 |
| QKI-NTRK2 Q6:N16 | |
| SQSTM1-NTRK2 S4:N16 | |
| STRN-NTRK2 S3:N16 | |
| SQSTM1-NTRK2 S4:N17 | |
| NTRK3 exon 14 | ETV6-NTRK3 E4:N14 |
| ETV6-NTRK3 E5:N14 | |
| NTRK3 exon 15 | ETV6-NTRK3 E4:N15 |
| ETV6-NTRK3 E5:N15 |
References
- Massi, D.; De Giorgi, V.; Mandalà, M. The complex management of atypical Spitz tumours. Pathology 2016, 48, 132–141. [Google Scholar] [CrossRef]
- Ferrara, G.; Cavicchini, S.; Corradin, M.T. Hypopigmented atypical Spitzoid neoplasms (atypical Spitz nevi, atypical Spitz tumors, Spitzoid melanoma): A clinicopathological update. Derm. Pr. Concept 2015, 30, 45–52. [Google Scholar] [CrossRef]
- Gerami, P.; Busam, K.; Cochran, A.; Cook, M.G.; Duncan, L.M.; Elder, D.E.; Fullen, D.R.; Guitart, J.; LeBoit, P.E.; Mihm, M.C., Jr.; et al. Histomorphologic assessment and interobserver diagnostic reproducibility of atypical spitzoid melanocytic neoplasms with long-term follow-up. Am. J. Surg. Pathol. 2014, 38, 934–940. [Google Scholar] [CrossRef]
- Cerroni, L.; Barnhill, R.; Elder, D.; Gottlieb, G.; Heenan, P.; Kutzner, H.; LeBoit, P.E.; Mihm, M., Jr.; Rosai, J.; Kerl, H. Melanocytic tumors of uncertain malignant potential: Results of a tutorial held at the XXIX Symposium of the International Society of Dermatopathology in Graz, October 2008. Am. J. Surg. Pathol. 2010, 34, 314–326. [Google Scholar] [CrossRef] [PubMed]
- Gill, M.; Renwick, N.; Silvers, D.N.; Celebi, J.T. Lack of BRAF mutations in Spitz nevi. J. Investig. Dermatol. 2004, 122, 1325–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmedo, G.; Hantschke, M.; Rütten, A.; Mentzel, T.; Hügel, H.; Flaig, M.J.; Yazdi, A.S.; Sander, C.A.; Kutzner, H. The T1796A mutation of the BRAF gene is absent in Spitz nevi. J. Cutan. Pathol. 2004, 31, 266–270. [Google Scholar] [CrossRef]
- Van Dijk, M.C.; Bernsen, M.R.; Ruiter, D.J. Analysis of mutations in B-RAF, N-RAS, and H-RAS genes in the differential diagnosis of Spitz nevus and spitzoid melanoma. Am. J. Surg. Pathol. 2005, 29, 1145–1151. [Google Scholar] [CrossRef]
- Pollock, P.M.; Harper, U.L.; Hansen, K.S.; Yudt, L.M.; Stark, M.; Robbins, C.M.; Moses, T.Y.; Hostetter, G.; Wagner, U.; Kakareka, J.; et al. High frequency of BRAF mutations in nevi. Nat. Genet. 2003, 33, 19–20. [Google Scholar] [CrossRef]
- Bauer, J.; Curtin, J.A.; Pinkel, D.; Bastian, B.C. Congenital melanocytic nevi frequently harbor NRAS mutations but no BRAF mutations. J. Investig. Dermatol. 2007, 127, 179–182. [Google Scholar] [CrossRef]
- Bastian, B.C.; LeBoit, P.E.; Pinkel, D. Mutations and copy number increase of HRAS in Spitz nevi with distinctive histopathological features. Am. J. Pathol. 2000, 157, 967–972. [Google Scholar] [CrossRef] [Green Version]
- Botton, T.; Yeh, I.; Nelson, T.; Vemula, S.S.; Sparatta, A.; Garrido, M.C.; Allegra, M.; Rocchi, S.; Bahadoran, P.; McCalmont, T.H.; et al. Recurrent BRAF kinase fusions in melanocytic tumors offer an opportunity for targeted therapy. Pigment. Cell Melanoma Res. 2013, 26, 845–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, I.; Botton, T.; Talevich, E.; Shain, A.H.; Sparatta, A.J.; de la Fouchardiere, A.; Mully, T.W.; North, J.P.; Garrido, M.C.; Gagnon, A.; et al. Activating MET kinase rearrangements in melanoma and Spitz tumours. Nat. Commun. 2015, 6, 7174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiesner, T.; He, J.; Yelensky, R.; Esteve-Puig, R.; Botton, T.; Yeh, I.; Lipson, D.; Otto, G.; Brennan, K.; Murali, R.; et al. Kinase fusions are frequent in Spitz tumours and spitzoid melanomas. Nat. Commun. 2014, 5, 3116. [Google Scholar] [CrossRef] [Green Version]
- Yeh, I.; Tee, M.K.; Botton, T.; Shain, A.H.; Sparatta, A.J.; Gagnon, A.; Vemula, S.S.; Garrido, M.C.; Nakamaru, K.; Isoyama, T.; et al. NTRK3 kinase fusions in Spitz tumours. J. Pathol. 2016, 240, 282–290. [Google Scholar] [CrossRef] [Green Version]
- Yeh, I.; Busam, K.J.; McCalmont, T.H.; LeBoit, P.E.; Pissaloux, D.; Alberti, L.; de la Fouchardière, A.; Bastian, B.C. Filigree-like Rete Ridges, Lobulated Nests, Rosette-like Structures, and Exaggerated Maturation Characterize Spitz Tumors With NTRK1 Fusion. Am. J. Surg. Pathol. 2019, 43, 737–746. [Google Scholar] [CrossRef] [PubMed]
- De la Fouchardière, A.; Tee, M.K.; Peternel, S.; Valdebran, M.; Pissaloux, D.; Tirode, F.; Busam, K.J.; LeBoit, P.E.; McCalmont, T.H.; Bastian, B.C.; et al. Fusion partners of NTRK3 affect subcellular localization of the fusion kinase and cytomorphology of melanocytes. Mod. Pathol. 2021, 34, 735–747. [Google Scholar] [CrossRef]
- Amin, S.M.; Haugh, A.M.; Lee, C.Y.; Zhang, B.; Bubley, J.A.; Merkel, E.A.; Verzì, A.E.; Gerami, P. A Comparison of Morphologic and Molecular Features of BRAF, ALK, and NTRK1 Fusion Spitzoid Neoplasms. Am. J. Surg. Pathol. 2017, 41, 491–498. [Google Scholar] [CrossRef]
- Elder, D.E.; Massi, D.; Scolyer, R.A.; Willemze, R. World Health Organization Classification of Skin Tumours; International Agency for Research on Cancer: Lyon, France, 2018. [Google Scholar]
- Raghavan, S.S.; Peternel, S.; Mully, T.W.; North, J.P.; Pincus, L.B.; LeBoit, P.E.; McCalmont, T.H.; Bastian, B.C.; Yeh, I. Spitz melanoma is a distinct subset of spitzoid melanoma. Mod. Pathol. 2020, 33, 1122–1134. [Google Scholar] [CrossRef]
- Drilon, A.; Laetsch, T.W.; Kummar, S.; DuBois, S.G.; Lassen, U.N.; Demetri, G.D.; Nathenson, M.; Doebele, R.C.; Farago, A.F.; Pappo, A.S.; et al. Efficacy of Larotrectinib in TRK Fusion-Positive Cancers in Adults and Children. N. Engl. J. Med. 2018, 378, 731–739. [Google Scholar] [CrossRef]
- Drilon, A.; Siena, S.; Ou, S.I.; Patel, M.; Ahn, M.J.; Lee, J.; Bauer, T.M.; Farago, A.F.; Wheler, J.J.; Liu, S.V.; et al. Safety and Antitumor Activity of the Multitargeted Pan-TRK, ROS1, and ALK Inhibitor Entrectinib: Combined Results from Two Phase I Trials (ALKA-372-001 and STARTRK-1). Cancer Discov. 2017, 7, 400–409. [Google Scholar] [CrossRef] [Green Version]
- Reuther, G.W.; Lambert, Q.T.; Caligiuri, M.A.; Der, C.J. Identification and characterization of an activating TrkA deletion mutation in acute myeloid leukemia. Mol. Cell Biol. 2000, 20, 8655–8666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truzzi, F.; Marconi, A.; Lotti, R.; Dallaglio, K.; French, L.E.; Hempstead, B.L.; Pincelli, C. Neurotrophins and their receptors stimulate melanoma cell proliferation and migration. J. Investig. Dermatol. 2008, 128, 2031–2040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocco, E.; Scaltriti, M.; Drilon, A. NTRK fusion-positive cancers and TRK inhibitor therapy. Nat. Rev. Clin. Oncol. 2018, 15, 731–747. [Google Scholar] [CrossRef]
- Marchiò, C.; Scaltriti, M.; Ladanyi, M.; Iafrate, A.J.; Bibeau, F.; Dietel, M.; Hechtman, J.F.; Troiani, T.; López-Rios, F.; Douillard, J.Y.; et al. ESMO recommendations on the standard methods to detect NTRK fusions in daily practice and clinical research. Ann. Oncol. 2019, 30, 1417–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conde, E.; Hernandez, S.; Sanchez, E.; Regojo, R.M.; Camacho, C.; Alonso, M.; Martinez, R.; Lopez-Rios, F. Pan-TRK Immunohistochemistry: An Example-Based Practical Approach to Efficiently Identify Patients With NTRK Fusion Cancer. Arch. Pathol. Lab. Med. 2021, 145, 1031–1040. [Google Scholar] [CrossRef]
- Hechtman, J.F.; Benayed, R.; Hyman, D.M.; Drilon, A.; Zehir, A.; Frosina, D.; Arcila, M.E.; Dogan, S.; Klimstra, D.S.; Ladanyi, M.; et al. Pan-Trk Immunohistochemistry Is an Efficient and Reliable Screen for the Detection of NTRK Fusions. Am. J. Surg. Pathol. 2017, 41, 1547–1551. [Google Scholar] [CrossRef] [PubMed]
- Bourhis, A.; Redoulez, G.; Quintin-Roué, I.; Marcorelles, P.; Uguen, A. Screening for NTRK-rearranged Tumors Using Immunohistochemistry: Comparison of 2 Different pan-TRK Clones in Melanoma Samples. Appl. Immunohistochem. Mol. Morphol. 2020, 28, 194–196. [Google Scholar] [CrossRef] [PubMed]
- Uguen, A. Spitz Tumors With NTRK1 Fusions: TRK-A and pan-TRK Immunohistochemistry as Ancillary Diagnostic Tools. Am. J. Surg. Pathol. 2019, 43, 1438–1439. [Google Scholar] [CrossRef]
- Solomon, J.P.; Benayed, R.; Hechtman, J.F.; Ladanyi, M. Identifying patients with NTRK fusion cancer. Ann. Oncol. 2019, 30, viii16–viii22. [Google Scholar] [CrossRef] [Green Version]
- Zito Marino, F.; Pagliuca, F.; Ronchi, A.; Cozzolino, I.; Montella, M.; Berretta, M.; Errico, M.E.; Donofrio, V.; Bianco, R.; Franco, R. NTRK Fusions, from the Diagnostic Algorithm to Innovative Treatment in the Era of Precision Medicine. Int. J. Mol. Sci. 2020, 21, 3718. [Google Scholar] [CrossRef]
- Doebele, R.C.; Davis, L.E.; Vaishnavi, A.; Le, A.T.; Estrada-Bernal, A.; Keysar, S.; Jimeno, A.; Varella-Garcia, M.; Aisner, D.L.; Li, Y.; et al. An Oncogenic NTRK Fusion in a Patient with Soft-Tissue Sarcoma with Response to the Tropomyosin-Related Kinase Inhibitor LOXO-101. Cancer Discov. 2015, 5, 1049–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiesner, T.; Kutzner, H.; Cerroni, L.; Mihm, M.C., Jr.; Busam, K.J.; Murali, R. Genomic aberrations in spitzoid melanocytic tumours and their implications for diagnosis, prognosis and therapy. Pathology 2016, 48, 113–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]



| Case | Age (Years) | Sex | Re-Excision | Sentinel Lymph Node Evaluation | Events | Follow-up (Months) | Cytoplasm | Cellular Membrane | Nucleus |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 24 | F | Yes | Negative | - | 60 | + | - | - |
| 2 | 4 | F | Yes | Not Evaluated | - | 50 | + | - | - |
| 3 | 10 | F | No | Not Evaluated | - | 67 | + | - | - |
| 4 | 13 | F | No | Not Evaluated | - | 1 | + | - | - |
| 5 | 18 | F | Yes | Positive | - | 240 | + | - | - |
| 6 | 22 | F | Yes | Not Evaluated | - | 63 | + | - | - |
| 7 | 37 | F | Yes | Negative | - | 50 | + | - | - |
| 8 | 36 | F | Yes | Negative | - | 132 | + | - | - |
| 9 | 37 | F | Yes | Negative | - | 132 | + | - | - |
| 10 | 38 | F | Yes | Not Evaluated | - | 159 | + | - | - |
| 11 | 59 | F | Yes | Not Evaluated | - | 43 | + | - | - |
| 12 | 32 | M | Yes | Not Evaluated | - | 50 | + | - | - |
| 13 | 30 | M | No | Not Evaluated | - | 137 | + | - | - |
| 14 | 61 | M | Yes | Negative | - | 96 | + | - | - |
| 15 | 2 | M | Yes | Not Evaluated | - | 84 | + | - | - |
| 16 | 18 | M | Yes | Not Evaluated | - | 42 | - | - | + |
| 17 | 24 | M | Yes | Not Evaluated | - | 110 | + | - | + |
| 18 | 3 | F | No | Not Evaluated | - | 66 | + | - | + |
| 19 | 8 | F | No | Not Evaluated | - | 89 | + | - | + |
| 20 | 12 | F | Yes | Not Evaluated | - | 124 | + | - | + |
| 21 | 31 | F | No | Not Evaluated | - | 12 | + | - | + |
| 22 | 47 | F | Yes | Negative | - | 84 | + | - | + |
| 23 | 24 | M | Yes | Not Evaluated | - | 132 | + | - | + |
| 24 | 39 | M | Yes | Negative | - | 72 | + | - | + |
| 25 | 57 | M | Yes | Negative | Local Recurrence | 72 | + | - | + |
| 26 | 18 | F | Yes | Negative | Local Recurrence | 75 | - | - | + |
| Case | Site | Diameter (mm) | Thickness (mm) | Mitoses (Number/mm2) | Cytotype | Filigree-Like Rete Ridges | Lobulated Melanocytic Nests | Rosette-Like Structures |
|---|---|---|---|---|---|---|---|---|
| 1 | NA | 24 | 1.5 | 0 | Epithelioid | - | - | - |
| 2 | NA | 4 | 1.9 | 4 | Epithelioid | - | - | - |
| 3 | Leg | 10 | 1.6 | 1 | Epithelioid | + | - | - |
| 4 | Arm | 13 | 0.7 | 1 | Epithelioid | - | - | - |
| 5 | Ear | 18 | 12 | 1 | Epithelioid/Spindle | - | - | - |
| 6 | NA | 22 | 0.5 | 0 | Spindle | - | - | - |
| 7 | NA | 37 | 0.9 | 2 | Epithelioid/Spindle | - | + | - |
| 8 | Leg | 36 | 0.8 | 1 | Epithelioid | - | - | - |
| 9 | Leg | 37 | 0.7 | 0 | Epithelioid/Spindle | - | - | - |
| 10 | Thigh | 38 | 1.1 | 0 | Epithelioid/Spindle | - | - | + |
| 11 | NA | 59 | 0.7 | 1 | Epithelioid/Spindle | - | + | - |
| 12 | NA | 32 | 1.1 | 1 | Epithelioid/Spindle | - | + | - |
| 13 | Arm | 30 | 1.2 | 1 | Epithelioid/Spindle | + | - | - |
| 14 | Leg | 61 | 1.2 | 2 | Epithelioid/Spindle | - | - | - |
| 15 | Buttock | 2 | 0.9 | 2 | Epithelioid | - | - | - |
| 16 | Trunk | 18 | 4 | 1 | Epithelioid/Spindle | - | - | - |
| 17 | NA | 24 | 1.9 | 2 | Epithelioid/Spindle | - | - | - |
| 18 | Leg | 3 | 1 | 3 | Epithelioid/Spindle | + | + | - |
| 19 | Leg | 8 | 1.1 | 1 | Epithelioid | + | - | - |
| 20 | Leg | 12 | 1.1 | 1 | Epithelioid | + | - | - |
| 21 | Leg | 31 | 0.8 | 1 | Epithelioid/Spindle | - | - | - |
| 22 | Arm | 47 | 1.2 | 2 | Epithelioid | - | - | - |
| 23 | Arm | 24 | 0.5 | 0 | Epithelioid/Spindle | - | + | - |
| 24 | Hip | 39 | 1.8 | 1 | Epithelioid/Spindle | + | - | - |
| 25 | Trunk | 57 | 0.8 | 0 | Epithelioid/Spindle | - | - | - |
| 26 | NA | 18 | 0.8 | 0 | Spindle | - | - | - |
| Case | RNA-Based NGS | Real-Time RT-PCR | FISH | Fusion Partner |
|---|---|---|---|---|
| 1 | NA/I | NA/I | Positive | Unknown (NTRK1) |
| 2 | NE | Positive | Negative | Unknown (NTRK1 exon 10) |
| 3 | NE | NE | Positive | Unknown (NTRK1) |
| 4 | NA/I | NA/I | NA/I | - |
| 5 | NA/I | NA/I | NE | - |
| 6 | NE | Positive | Positive | Unknown (NTRK1 exon 10) |
| 7 | NA/I | NA/I | NA/I | - |
| 8 | NA/I | NA/I | NA/I | - |
| 9 | Positive | Positive | NE | LMNA-NTRK1 |
| 10 | NA/I | NA/I | Positive | Unknown (NTRK1) |
| 11 | NA/I | NA/I | Positive | Unknown (NTRK1) |
| 12 | Positive | Positive | Positive | LMNA-NTRK1 |
| 13 | NE | NE | Negative | - |
| 14 | NA/I | NA/I | Negative | - |
| 15 | Positive | Positive | Negative | LMNA-NTRK1 |
| 16 | Positive | Positive | Positive | ETV6-NTRK3 |
| 17 | NA/I | NA/I | Positive | Unknown (NTRK1) |
| 18 | NE | Positive | NE | Unknown (NTRK1 exon 10) |
| 19 | NE | NE | NE | - |
| 20 | NA/I | NA/I | Negative | - |
| 21 | NE | NE | Positive | Unknown (NTRK1) |
| 22 | NE | Positive | Positive | Unknown (NTRK1 exon 11–12 del) |
| 23 | NE | NE | Negative | - |
| 24 | NE | NE | Positive | Unknown (NTRK1) |
| 25 | Positive | Positive | Negative | LMNA-NTRK1 |
| 26 | NA/I | NA/I | Negative | Unknown (NTRK1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cappellesso, R.; Nozzoli, F.; Zito Marino, F.; Simi, S.; Castiglione, F.; De Giorgi, V.; Cota, C.; Senetta, R.; Scognamiglio, G.; Anniciello, A.M.; et al. NTRK Gene Fusion Detection in Atypical Spitz Tumors. Int. J. Mol. Sci. 2021, 22, 12332. https://doi.org/10.3390/ijms222212332
Cappellesso R, Nozzoli F, Zito Marino F, Simi S, Castiglione F, De Giorgi V, Cota C, Senetta R, Scognamiglio G, Anniciello AM, et al. NTRK Gene Fusion Detection in Atypical Spitz Tumors. International Journal of Molecular Sciences. 2021; 22(22):12332. https://doi.org/10.3390/ijms222212332
Chicago/Turabian StyleCappellesso, Rocco, Filippo Nozzoli, Federica Zito Marino, Sara Simi, Francesca Castiglione, Vincenzo De Giorgi, Carlo Cota, Rebecca Senetta, Giosuè Scognamiglio, Anna Maria Anniciello, and et al. 2021. "NTRK Gene Fusion Detection in Atypical Spitz Tumors" International Journal of Molecular Sciences 22, no. 22: 12332. https://doi.org/10.3390/ijms222212332
APA StyleCappellesso, R., Nozzoli, F., Zito Marino, F., Simi, S., Castiglione, F., De Giorgi, V., Cota, C., Senetta, R., Scognamiglio, G., Anniciello, A. M., Cesinaro, A. M., Mandalà, M., Gianatti, A., Valente, M. G., Valeri, B., Sementa, A. R., Ricci, C., Corti, B., Roviello, G., ... Massi, D. (2021). NTRK Gene Fusion Detection in Atypical Spitz Tumors. International Journal of Molecular Sciences, 22(22), 12332. https://doi.org/10.3390/ijms222212332